






 , Marla Xhani 1,3,†, Benedetta Sprovera 1, Brigitta Buttari 4, Giorgia Abballe 1, Flaminia Desideri 1, Emiliano Panieri 5,6, Luciano Saso 5, Marzia Arese 1,*
, Marla Xhani 1,3,†, Benedetta Sprovera 1, Brigitta Buttari 4, Giorgia Abballe 1, Flaminia Desideri 1, Emiliano Panieri 5,6, Luciano Saso 5, Marzia Arese 1,*1 Department of Biochemical Sciences “A. Rossi Fanelli”, Sapienza University of Rome, 00185 Rome, Italy
2 Department of Biosciences, Biotechnologies and Biopharmaceutics, University of Bari “Aldo Moro”, 70121 Bari, Italy
3 Istituto Zooprofilattico Sperimentale del Lazio e della Toscana “M. Aleandri”, 00178 Rome, Italy
4 Department of Cardiovascular, Endocrine-Metabolic Diseases and Aging, Italian National Institute of Health, 00161 Rome, Italy
5 Department of Physiology and Pharmacology “Vittorio Erspamer”, Sapienza University of Rome, 00185 Rome, Italy
6 Section of Hazardous Substances, Environmental Education and Training for the Technical Coordination of Management Activities (DGTEC), Italian Institute for Environmental Protection and Research, 00144 Rome, Italy
†These authors contributed equally.
Academic Editor: Josef Jampílek
Abstract
Background: Bisphenol A (BPA) and perfluorooctanoic acid (PFOA) are
synthetic compounds widely utilized in industrial activities devoted to the
production of daily life plastic, metal products, and packaging from which they
are able to migrate to food and water. Due to their persistence in the
environment, living organisms are chronically exposed to these pollutants. BPA
and PFOA have adverse effects on tissues and organs. The aim of this study was to
identify the molecular targets and biochemical mechanisms involved in their
toxicity. Methods: HepG2 and HaCaT cells were treated with BPA or PFOA,
and the trypan blue exclusion test and 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT) assay were performed to define the
conditions for subsequent investigations. We conducted quantitative PCR and
western blot analysis to evaluate the expression of proteins involved in nitric
oxide (NO) signaling. Cell-based assays were carried out to evaluate reactive
oxygen species (ROS) production, nitrite/nitrate (NOx) accumulation,
3-nitrotyrosine (3-NT) formation, and mitochondrial membrane potential (MMP)
determination in treated cells. Results: HepG2 and HaCaT cells incubated
for 24 h with subtoxic concentrations of BPA or PFOA (50 and 10
Graphical Abstract
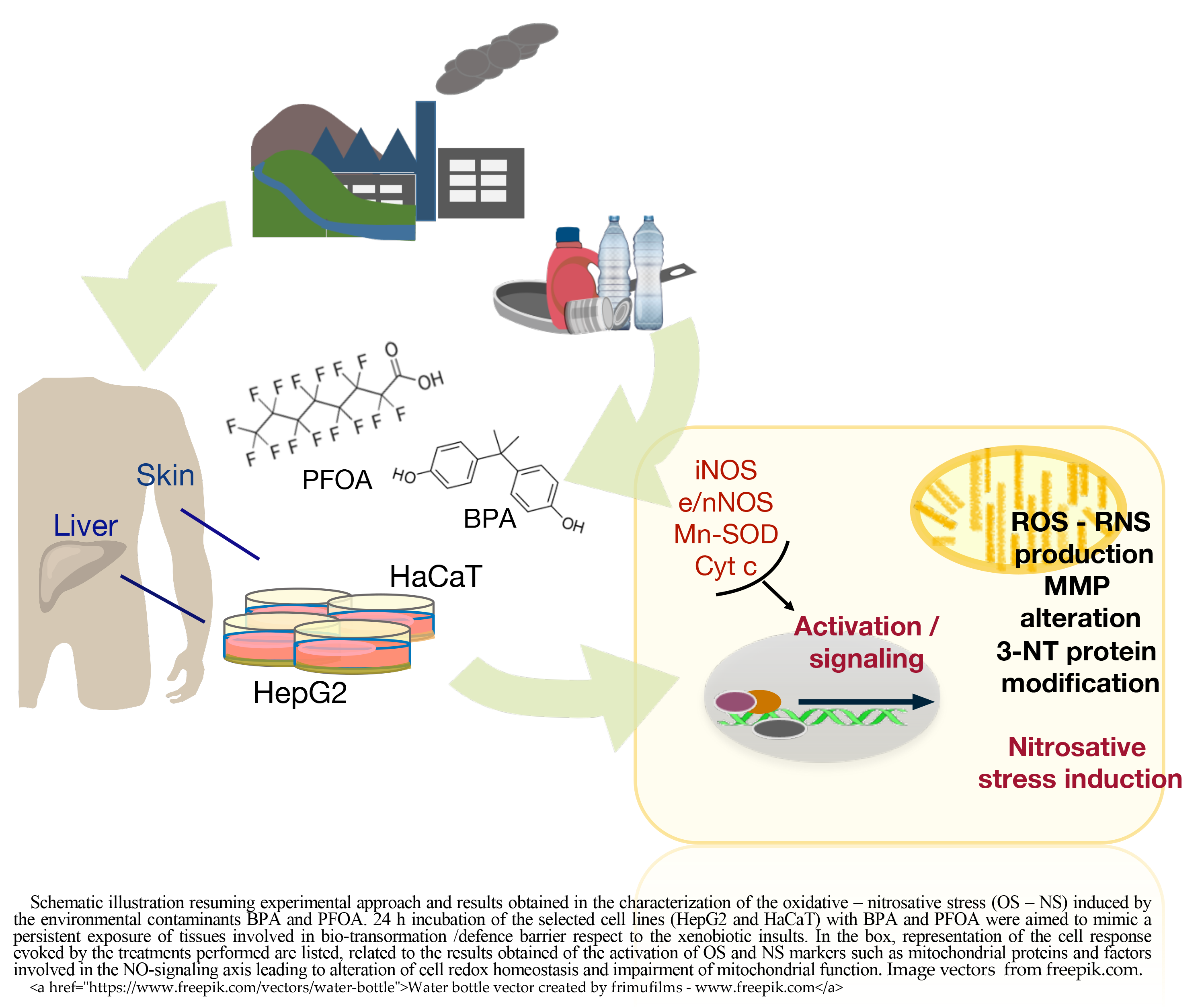
Keywords
- emerging contaminants
- endocrine disruptors
- nitric oxide signaling
- nitro-oxidative stress
- mitochondrial dysfunction
In healthy cells, the redox balance is maintained through a complex network of adaptive responses that include the continuous generation and elimination of reactive oxygen/nitrogen species (ROS/RNS). The fine-tune regulation of redox signaling events is activated by incoming insults (physical, chemical, or biological) to restore the physiological equilibrium [1, 2, 3, 4, 5].
Mitochondria are one of the main endogenous sources of ROS due to the formation of superoxide ion (O
The short-lived, highly reactive peroxynitrite (ONOO
A number of xenobiotics and related chemicals (e.g., atmospheric pollutants and industrial contaminants) are involved in the production of intracellular ROS/RNS, and the persistence of a given prooxidant stimulus may specifically affect redox homeostasis leading to chronic oxidative stress (OS) [4, 15].
Diffuse OS is a common alteration in several pathological conditions including hypertension, diabetes, neurodegeneration and cancer [1, 6, 19, 20]. Among the exogenous sources of nitro-OS, there has been growing interest in the chemical derivatives of synthetic compounds utilized in industrial manufacture. These chemicals, widely utilized as plasticizer and waterproof materials for packaging, utensils and pharmaceuticals, are now classified as emerging contaminants (ECs) as they are dispersed in the environment at alarming concentrations [21, 22].
Bisphenol A (BPA) and perfluoro-octanoic acid (PFOA) are ideal representatives of this class of pollutants. Due to their intrinsic high stability, resistance to environmental conditions and long half-life, both have been widely used as components of utensils and personal care tools to confer durability, plasticity and impermeabilization [23, 24, 25].
The documented migration of BPA and PFOA from products to soil, water and food, and the consequent bioaccumulation of these ECs have established a condition of human chronic exposure to these pollutants through ingestion, inhalation and dermal absorption [26, 27, 28, 29]. Great efforts are being made to clarify the adverse effects that both BPA and PFOA may have on living organisms, and to better understand the mechanisms underlying their toxic effects [27, 28, 30, 31, 32, 33, 34].
BPA and PFOA are endocrine disruptors (EDs), i.e., molecules able to interfere with hormone-driven processes [35]. Several studies have shown correlations between exposure to EDs and decreased fertility, as well as an increased incidence of cancers such as breast, ovarian, thyroid, lung, and testis [33, 35, 36, 37, 38].
Both BPA and PFOA are thought to target the mitochondria of exposed cells, thus inducing mitochondrial dysfunction and promoting an increase of both ROS and RNS with consequent onset of OS [39, 40, 41, 42, 43]. Glutamatergic neurons have increased expression of both iNOS and nNOS, resulting in the elevation of ROS and RNS and 3-nitrotyrosine (3-NT) production induced by chronic exposure to BPA [44], whereas 24 h PFOA exposure results in increased ROS and antioxidant enzyme expression in the HepG2 human hepatoma cell line [45]. The proposed mechanisms of BPA and PFOA toxicities are linked to their structural and chemical properties, which favor cell permeability and point to the interactions with hormone receptors (i.e., estrogen receptor and peroxisome proliferator activated receptor alpha, respectively) in exposed cells [46, 47, 48]. Recent studies showed that PFOA was able to interact with superoxide dismutase (SOD) directly, inducing OS and apoptosis [49], and may lead to the production of proinflammatory cytokines, ROS/RNS increase and mitochondrial dysfunction in rat cell model [40].
In this work, the mechanisms of toxicity of the EDs BPA and PFOA were investigated, focusing on their ability to interfere with the NO signaling. To this end, two human cell models were chosen to assess the effects of BPA and PFOA treatments: the HepG2 hepatocellular cancer-derived cell line, which is the gold standard model for xenobiotic metabolism and cytotoxicity studies; and the HaCaT immortalized keratinocyte cell line, which is a model system to study the skin as the outer barrier to environmental cytotoxic and genotoxic agents [50, 51, 52].
The expression of regulators involved in NO metabolism such as the NOS isoforms (eNOS in HepG2, nNOS in HaCaT and iNOS in both cell lines) was assessed at both the mRNA and protein levels, in association with evaluation of the expression of the mitochondrial proteins manganese SOD (MnSOD) and cytochrome c (cyt c). As a relevant parameter for the analyses of oxidative and nitrosative stress (NS), the level of ROS production in cells treated with BPA and PFOA was determined, as well as the accumulation of nitrites/nitrates (NOx), reflecting the increased production of NO compared to untreated cells. Furthermore, as a downstream effect of altered ROS/RNS homeostasis, we evaluated the level of 3-NT modification in proteins and alterations of the mitochondrial membrane potential (MMP).
Upon cell treatment with low
As members of a wide class of synthetic pollutants (ECs, EDs), both BPA and PFOA have analogues for which a comparable level of toxicity has been reported [27, 28, 38, 39, 49]. Further work is needed to clarify the effective EC pressure to which living organisms are exposed and related redox dysregulation is generated. We postulate that a comprehensive understanding of the specific contribution of abundant ECs (e.g., BPA or PFOA) to alterations in NO signaling and the promotion of OS and NS will provide a concrete basis by which to define the harmful effects of these pollutants. Of relevance, the major involvement of NO signaling in the toxicity exerted by the ECs BPA and PFOA and a novel remarkable effect of PFOA on the regulation of NOS isoforms were observed in this study. Within the limitation of the model system (cell lines) and application of selective incubation conditions, which do not represent the complexity of the whole organism, we found this cell line approach as an opportunity to evaluate the parameters related to altered ROS and RNS bioavailability.
The results of this study may provide insight into the effects derived from an impaired antioxidant cell response, which leads to an ROS/RNS increase, bioenergetic dysfunction and inflammation at the tissue level, a condition favoring the onset of pathological states [1, 20, 53].
The HepG2 human hepatocellular cancer-derived cell line (HB-8065TM; ATCC, Manassas, VA, USA) was maintained in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 2 mM L-glutamine (Aurogene SRL, Rome, Italy), 10% heat-inactivated fetal bovine serum (FBS) and 1% antibiotics of 50 U/mL penicillin and 50
Cells were incubated in the presence or absence of BPA and PFOA at different concentrations and times. When necessary, cells were harvested by trypsinization and centrifugation (1000
DMEM and FBS were from Invitrogen Life Technologies (Gibco, Paisley, UK) and PAA Laboratories (Linz, Austria). BPA, perfluorooctanoic acid, JC-1, MTT, and trypan blue solution were from Merck Life Science. PFOA was dissolved in sterile dimethyl sulfoxide (DMSO; Merck Life Science) and further diluted in water; the administration of 10
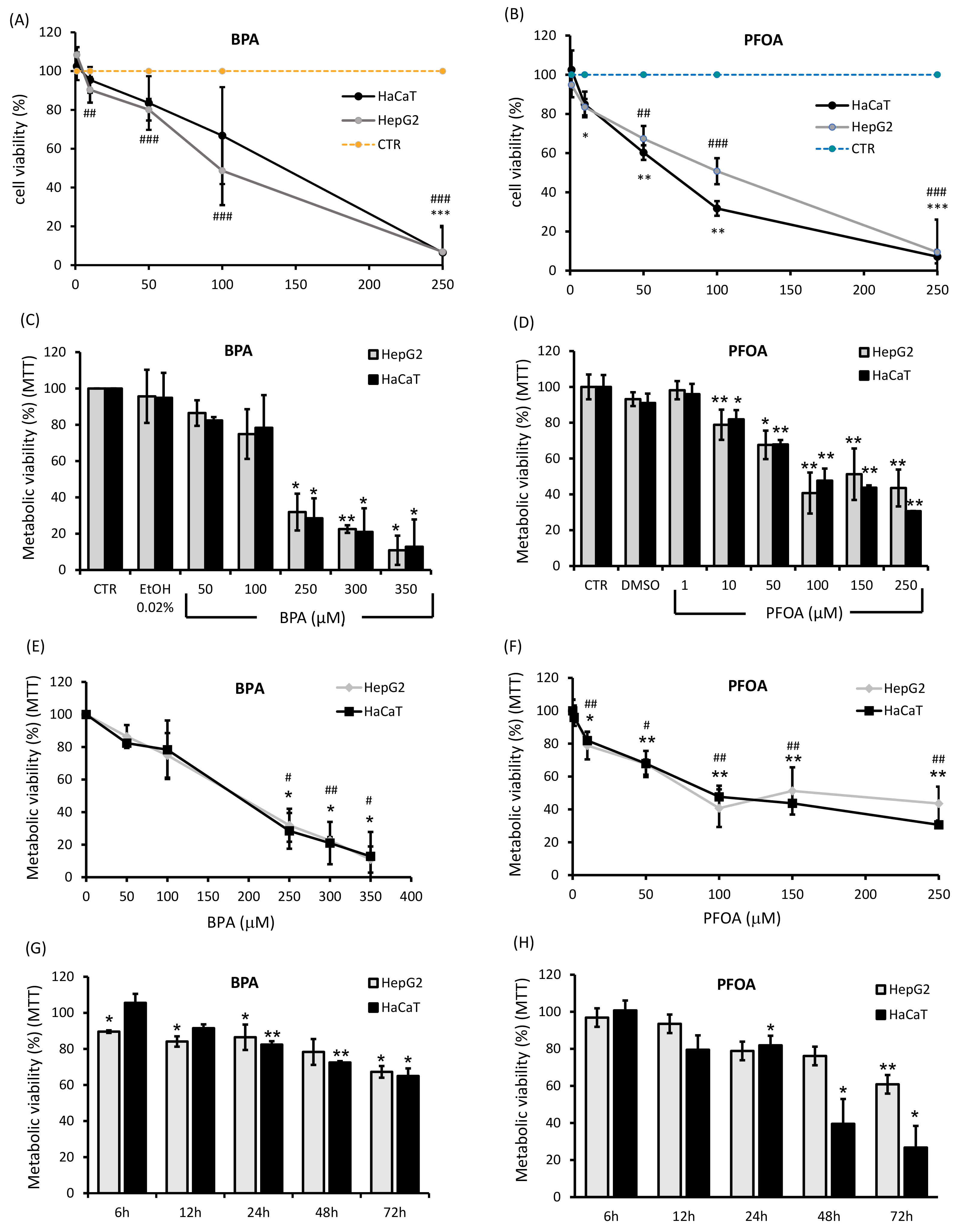 Fig. 1.
Fig. 1.Viability of HepG2 and HaCaT cells after exposure to BPA and PFOA. The percentage of living cells undergoing treatments with BPA or PFOA was assayed by the trypan blue exclusion test in a concentration range of 0–250
The viability of HepG2 and HaCaT cells was assessed by the trypan blue exclusion test and MTT [3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide] reduction assay as previously described [54]. Briefly, cells seeded on 35 mm dish or 6-well plates were incubated for 24 h with BPA or PFOA (from 1 to 250
Quantitative PCR (qPCR) was carried out in HepG2 and HaCaT cells. Briefly, cells were incubated for 24 h with BPA (50
HepG2 and HaCaT cells (3
Reactive oxygen species (ROS) generation was assessed in living cells using two fluorescent probes with a slight difference in ROS specificity for independent evaluations: red-fluorescent MAK145 (Merck Life Science) and 2
The total NOx was evaluated as a quantitative measure of NO production. The accumulation of NOx was assessed in the culture medium of HepG2 and HaCaT cells (~2.5
The evaluation of 3-NT modified proteins was used as marker of peroxynitrite- mediated NS in HepG2 and HaCaT cells treated with BPA 50
MMP was measured by flow cytometry (Accuri C6 Flow Cytometer®; Becton Dickinson, Franklin Lakes, NJ, USA) to detect the accumulation of the cationic fluorescent probe JC-1 (5,5’,6,6’-tetrachloro-1,1’,3,3’ tetraethylbenzimidazolylcarbocyanine iodide) (Abcam) into the mitochondrial negatively charged matrix according to the manufacturer’s instructions [55]. Briefly, after treatment with BPA 50
Data are reported as the mean
The cytotoxicity of BPA and PFOA in HepG2 and HaCaT cells was determined by the trypan blue exclusion and MTT assays (Fig. 1). Dose-response analyses were performed by incubating both cell lines with increasing amounts of BPA or PFOA. As shown in Fig. 1A–C, 24 h exposure to BPA induced similar effects on HaCaT and HepG2 cells, namely, a decrease in cell viability of about 20% in a concentration range of 50–100
The results of the viability assays allowed us to identify a suitable concentration and incubation time for further experiments: 50
The mRNA and protein levels of the different isoforms of NOS (iNOS and tissue-specific NOS, eNOS produced in hepatocytes [HepG2], and nNOS expressed in keratinocytes [HaCaT]) were determined after incubation with BPA or PFOA for 24 h. Compared to controls, BPA induced a two-fold increase in both iNOS (p = 0.015 vs ctr) and eNOS (p = 0.023 vs ctr) mRNA in HepG2 cells; the effect of PFOA on iNOS activation was higher (four-fold; p = 6
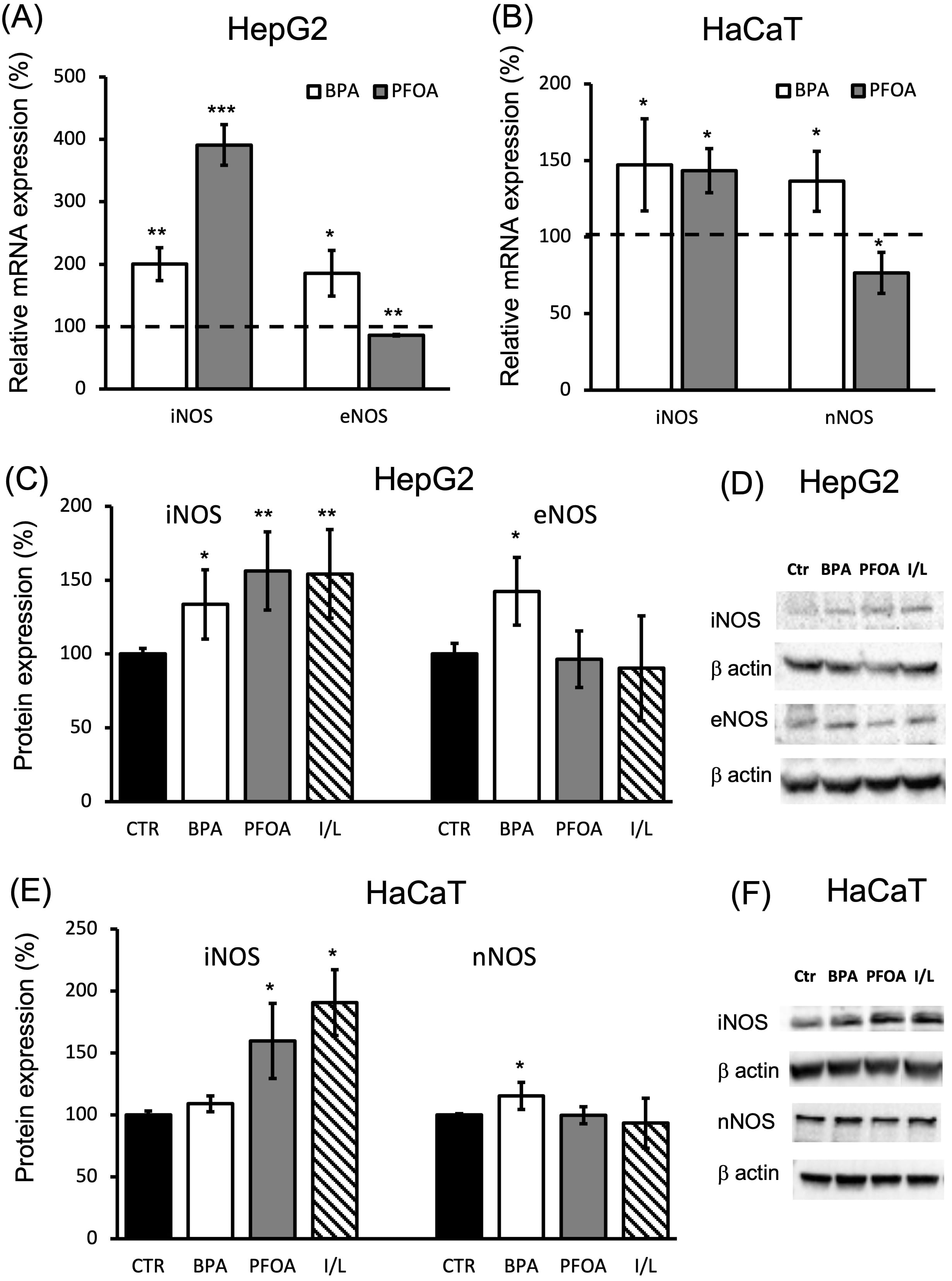 Fig. 2.
Fig. 2.NOS isoform expression in HepG2 cells and HaCaT cells treated with BPA or PFOA. (A–B) qPCR analysis was carried out (A) in the presence of iNOS/eNOS specific primers and cDNA purified from HepG2 cells treated for 24 h with 50
The mRNA expression levels of the mitochondrial proteins MnSOD and cyt c were detected following treatment of HepG2 and HaCaT cells with BPA or PFOA under the conditions previously described. The level of MnSOD mRNA was not significantly altered in HepG2 after 24 h of BPA exposure, and was slightly induced by PFOA (1.25-fold; p = 0.014 vs ctr) (Fig. 3A), although at the protein level, the significant upregulation of MnSOD was induced by both EDs (40% BPA increase; p
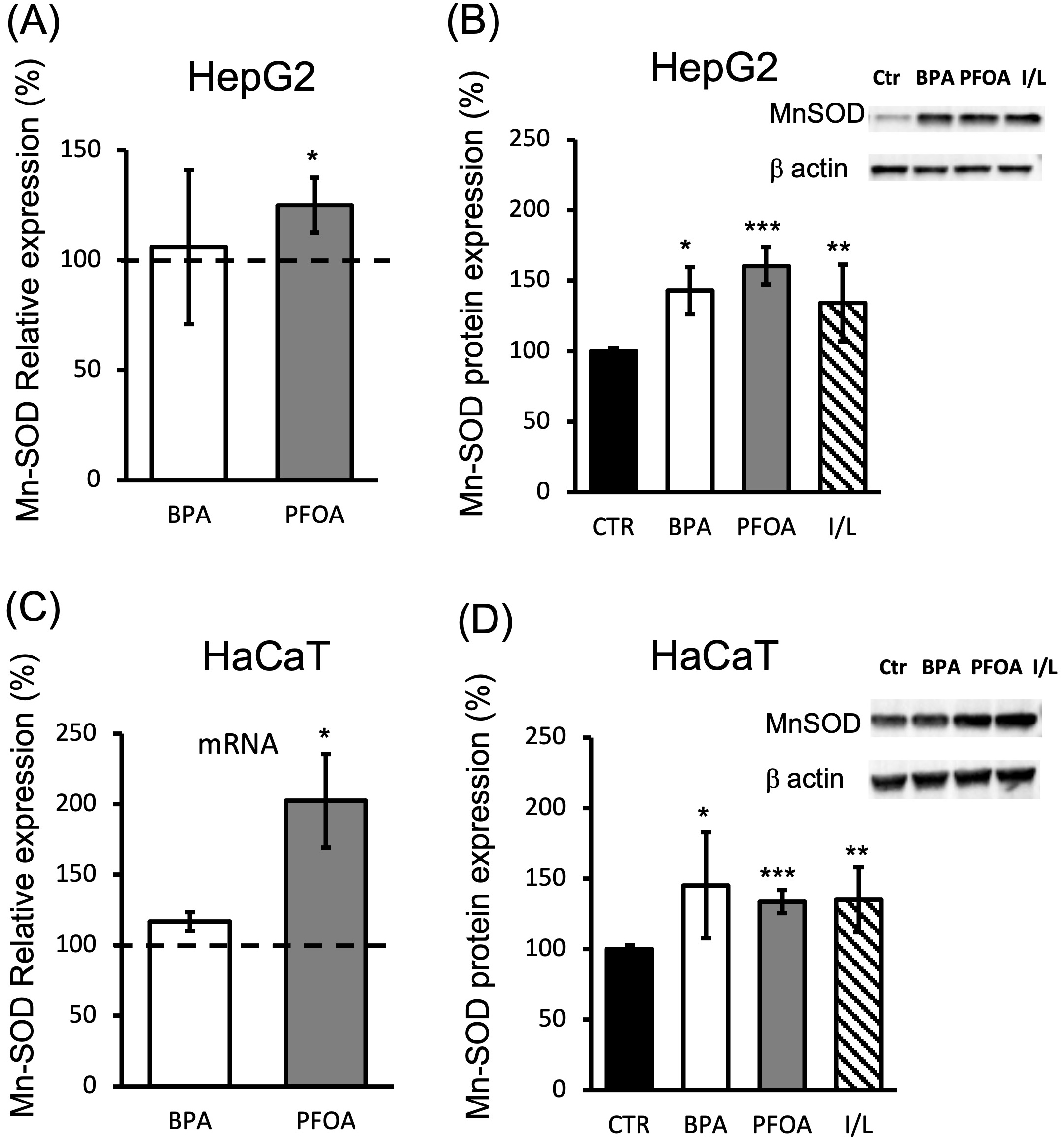 Fig. 3.
Fig. 3.MnSOD expression in HepG2 and HaCaT cells treated with BPA and PFOA. (A,C) MnSOD mRNA evaluation by qPCR analysis was carried out in the presence of MnSOD-specific primers (see methods) and cDNA purified from (A) HepG2 or (C) HaCaT cells treated for 24 h with 50
The analysis of cyt c expression showed mRNA induction specifically driven by PFOA treatment of both cell lines, with a stronger effect observed in HepG2 cells (above two-fold, p = 0.0036; Fig. 4A) compared to HaCaT (~1.8-fold, p = 0.045; Fig. 4C). BPA treatment led to a small but significant decrease in cyt c mRNA (35% decrease; p = 0.036), with the effect limited to HaCaT cells, as in HepG2 cells, there was no significant change in cyt c mRNA expression after BPA treatment compared to the control (Fig. 4A). As shown in Fig. 4B, cyt c protein expression detected by Western blot analysis in HepG2 cells was ~1.4-fold higher in response to both BPA (p = 0.0087) and PFOA (p = 0.01) treatment, whereas the response of HaCaT cells regarding cyt c protein expression (Fig. 4D) was reproduced only in the case of PFOA treatment (~1.5-fold increase; p = 0.0013) but not by BPA. In keratinocytes, indeed, BPA led to no changes in cyt c protein expression under the condition assayed.
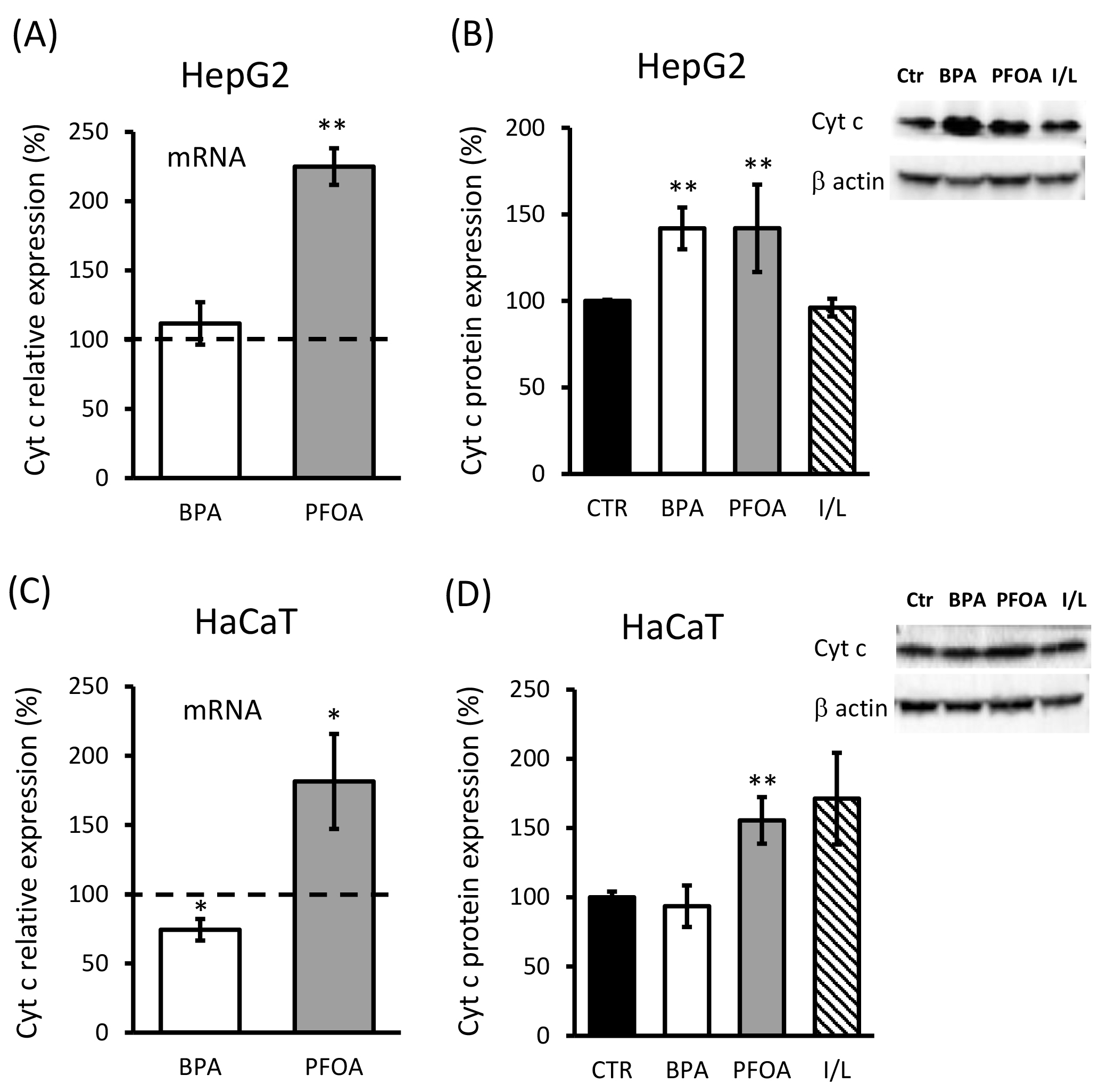 Fig. 4.
Fig. 4.Cyt c expression in HaCaT cells treated with BPA and PFOA. (A,C) Cyt c mRNA evaluation by qPCR analysis carried out in the presence of cyt c-specific primers (see methods) and cDNA purified from (A) HepG2 or (C) HaCaT cells treated for 24 h with 50
Overall, in HaCaT cells, 24 h treatment with BPA showed a lower sensitivity in terms of cyt c and MnSOD expression with respect to HepG2 and compared with the effects exerted by PFOA.
The production of ROS, the accumulation of NOx and the amount of NT modifications in proteins (3-NT) were measured to assess the OS and NS levels in HepG2 and HaCaT cells after 24 h treatment with BPA or PFOA. ROS determination was carried out taking advantage of two different ROS-targeting probes, for independent evaluations: DCFDA (Fig. 5A), mainly targeting hydrogen peroxide (H
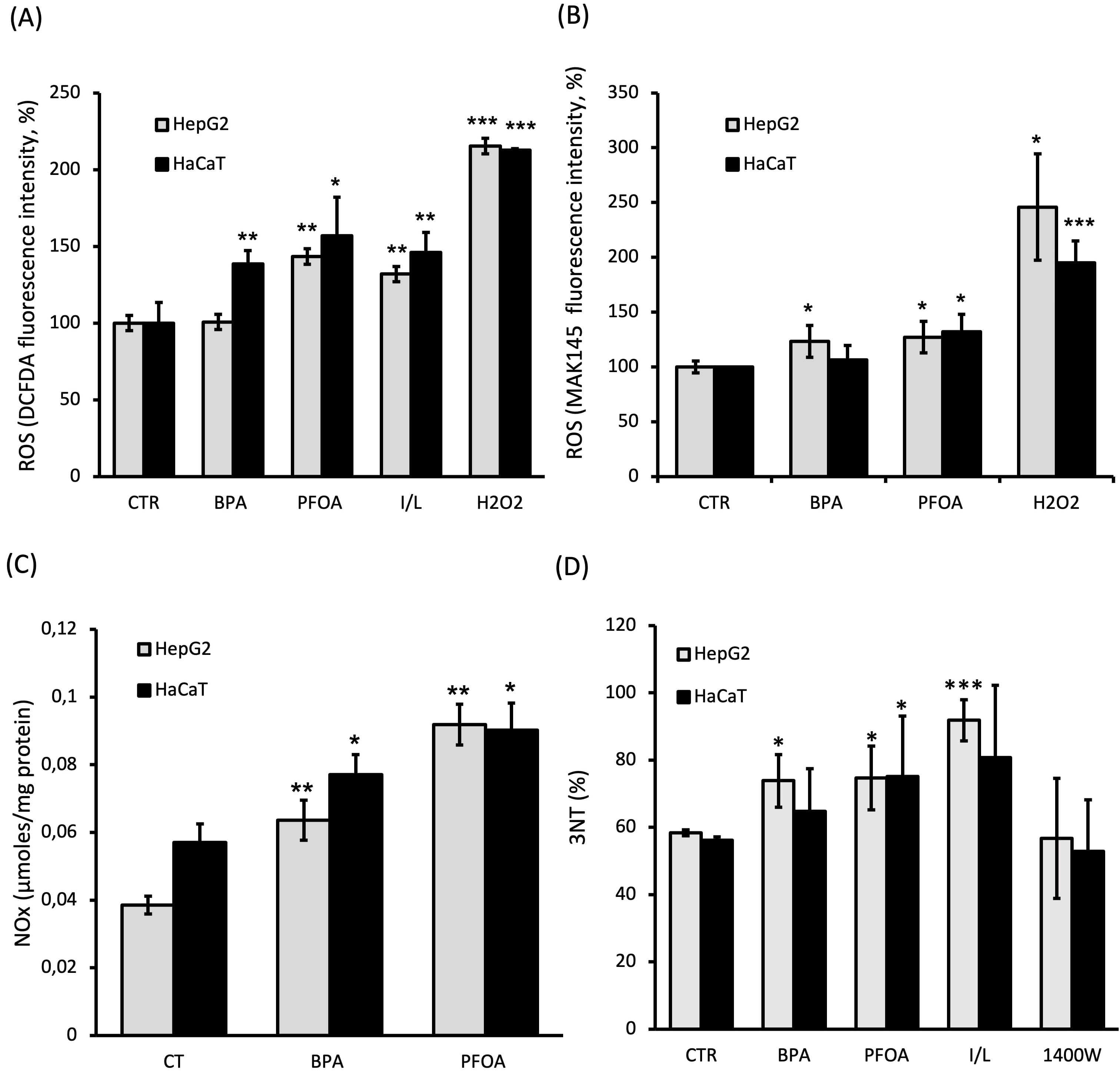 Fig. 5.
Fig. 5.OS and NS induced by BPA and PFOA. Assays were carried out following 24 h incubation of both HaCaT and HepG2 cells with BPA 50
The levels of NO end products, NOx, were found to be increased by treatment with both BPA and PFOA in the two cell lines tested (Fig. 5C). The effect was more evident in PFOA-treated cells, resulting in the increased accumulation of NOx by about two-fold and 1.5-fold for HepG2 (p = 0.0012) and HaCaT (p = 0.05) cells, respectively. A smaller but clearly significant induction of NOx was also achieved by BPA, by about 1.5-fold in both cell lines (p HepG2 = 0.009 and p HaCaT = 0.045 vs controls). It is worth noting that the result of increased NOx accumulation was in good agreement with the observation of increased NOS expression (especially iNOS) observed upon ED treatments.
As shown in Fig. 5D, the amount of NT modification in proteins was increased of about 25% in HepG2 cells following treatment of both BPA (p = 0.029 vs ctr) and PFOA (p = 0.018 vs ctr). Similar changes were seen in HaCaT cells exposed to PFOA (p = 0.021 vs ctr), whereas only a small irrelevant increase was observed for BPA (lower than 20%) under the assayed conditions. It is worth mentioning that a 24 h incubation of IFN-
The overall mitochondrial functional state was investigated by measuring the MMP of BPA- or PFOA-treated cells, by evaluating the mitochondrial import of the fluorescent probe JC-1 and the formation of the red J-aggregates. As shown in Fig. 6A, after 24 h incubation with BPA, the MMP was significantly lowered in HepG2 (~20% decrease; p = 0.0115) and HaCaT (~40% decrease; p = 0.023), whereas no significant difference was induced by PFOA. Surprisingly, the response of the MMP to the two-step addition of the ionophores nigericin and valinomycin, indicated a condition of hyperpolarization determined by the incubation of cells with PFOA. The hyperpolarization was significant in HepG2 (~30% decrease; p = 0.032) with respect to the polarization level of the untreated cells, whereas a small non-significant depolarization was observed for BPA treatment (Fig. 6B). The MMP alterations described, specifically induced by BPA or PFOA, were observed in the two cell lines assayed.
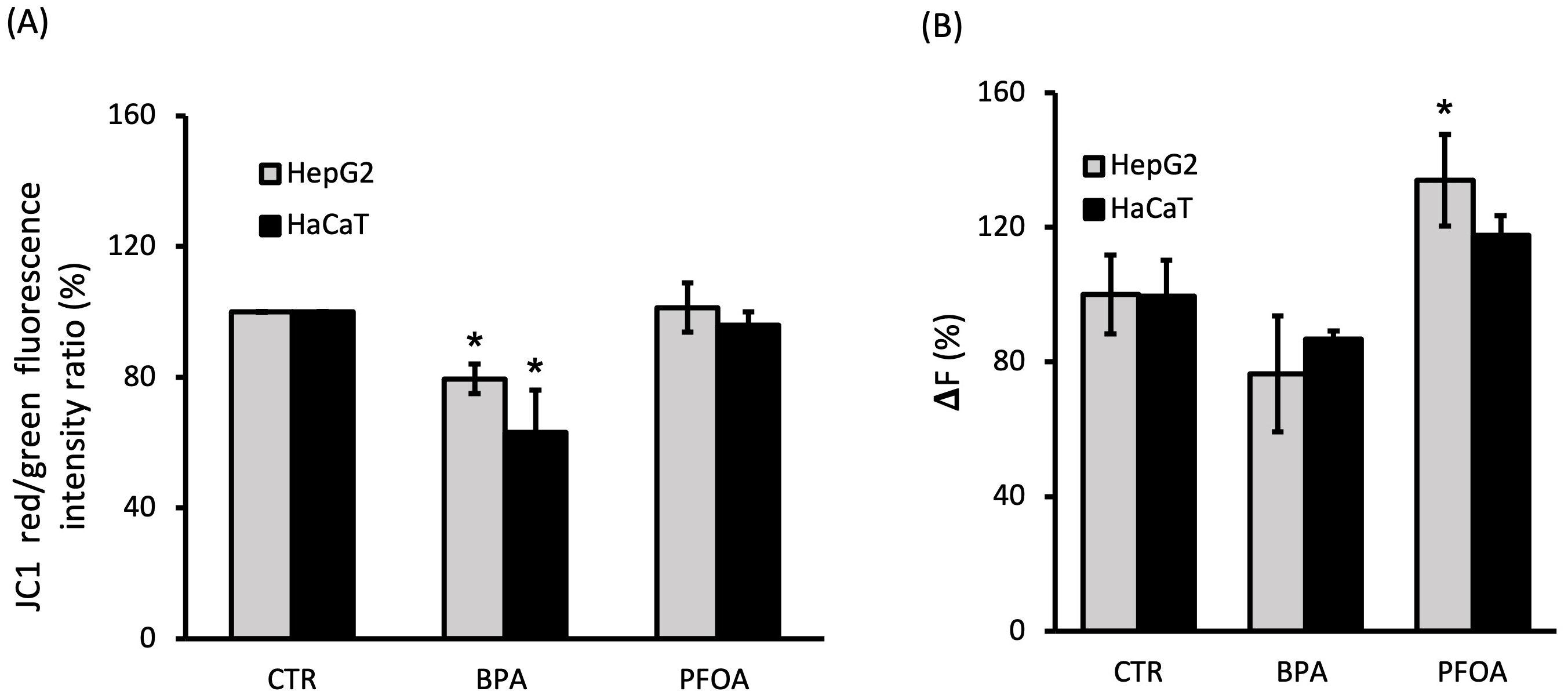 Fig. 6.
Fig. 6.MMP in cells incubated with BPA and PFOA. (A) The percentage of intensity ratio between red and green fluorescence of JC-1 was measured by flow cytometry in HepG2 and HaCaT cells after 24 h incubation with BPA 50
Environmental diffusion of the ECs BPA and PFOA, compounds widely utilized in industrial manufacturing processes [21, 22], has resulted in alarming concentrations detected in water and soil as well as bioaccumulation, as demonstrated by the detection of both BPA and PFOA in human body fluids with documented mother to fetus transfer [56, 57]. The adverse effects exerted by chronic exposure of BPA and PFOA have been described over the last several years [27, 32, 35]. Of relevance, the ED action shown for both compounds [33, 35, 36], and related alterations of the physiological hormonal signals are strictly connected to the perturbation of cellular pathways related to redox homeostasis. The involvement of BPA in OS induction has been previously suggested based on its interaction with estrogen receptors, although the mechanisms of this process need to be elucidated [27, 31, 41, 44, 58].
This paper reports a detailed characterization of the biochemical parameters related to OS and NS induced by BPA, showing a correlation between alterations of ROS and RNS signaling and establishment of a cell redox imbalance with consequences at the mitochondrial level. Furthermore, the novelty of the specific involvement of PFOA in the perturbation of the NO signaling axis is proposed here, with activation of iNOS and production of peroxinitrite, suggesting a specific effect exerted by PFOA toward NS induction, under the condition assayed.
The limitations of the study with respect to the biomedical relevance of the findings must be taken into account, with particular reference to the cell models and the concentration of ECs assayed. As transformed/immortalized cells, the HepG2 and HaCaT cell lines utilized in this study may display altered metabolism and viability respect to normal cells. However, it must be considered that the cell lines utilized represent a model system for toxicological studies and that the experimental condition utilized (low glucose adaptation, FBS and phenol red starvation) were adopted to minimize eventual deviations in the results. The screening of dose-response and time dependency of the ECs carried out by the trypan blue exclusion test and MTT assay (Fig. 1) was conducted to establish a suitable concentration and time window for further investigations regarding OS and NS in cell models of interest.
At the concentrations of 50
The increased expression of the “physiological” tissue-specific NOS isoform (eNOS in HepG2 and nNOS in HaCaT) induced by BPA may indicate the activation of adaptation pathways that possibly stimulated a specific cell stress response counteracting the detrimental activity of iNOS to limit the increase of highly reactive molecules to an acceptable range in terms of OS/NS. Under the condition assayed (50
At the mitochondrial level, BPA administration led to a decrease in MMP in both cell lines (Fig. 6A). In HaCaT cells, this result was associated with the low involvement of cyt c with respect to BPA-induced modulatory events (Fig. 4C,D), revealing a depressed mitochondrial function. In view of the endocrine disruptive activity of BPA, under the conditions herein explored, we clearly showed that alteration of the NO axis [43, 44] and dysregulation of the mitochondrial parameters [41], which have been previously proposed, are indeed strictly correlated and may be associated with the induction of nitro-OS [31, 33].
The results observed upon PFOA treatment (10
BPA and PFOA are both able to influence and alter the NO signaling pathway and promote the alteration of mitochondrial function. In hepatocytes, PFOA determines a proinflammatory condition, characterized by the high level of iNOS expression, increased ROS, significant alteration of proteins at the nitro-tyrosine level, and mitochondrial hyperpolarization. In the same cells, BPA was active in deregulating physiologic eNOS/iNOS crosstalk at the expression level, leading to elevation of NT modification with consequences at the mitochondrial level, as suggested by the decreased MMP. The keratinocyte response to insult arising from contaminant administration in the case of PFOA resulted in lower involvement of NO signaling compared to HepG2, but still with indications of an altered MMP and MnSOD activation at the mitochondrial level. The observed (mild) induction of iNOS mRNA driven by BPA was not accompanied by increased iNOS protein in keratinocytes, with the overall effect of a minor involvement of the NO axis under the tested conditions. However, BPA was able to induce persistent mitochondrial depolarization in keratinocytes, in accordance with the endocrine-disruption physiological alterations induced by this compound.
The experimental work herein presented shows that BPA and PFOA may target different components of the NO signaling pathway, with downstream effects mostly related to alteration of cell redox homeostasis. Further investigations are needed to fully understand the bioenergetic alterations and cell response linked to the nitro-OS induced by these pollutants. The findings reported will help clarify the mechanisms of action of BPA and PFOA, which will aid in the management of pathological relapses linked to chronic exposure to these environmental contaminants.
3-NT, 3-nitrotyrosine; BCA, Bicinchoninic acid; BPA, Bisphenol A; Cyt c, Cytochrome c; DAN, 1,1’,3,3’-tetraethyl-5,5’,6,6’-tetrachloroimidacarbocyanine iodide; DCFDA, dichlorodihydrofluorescein diacetate; DMSO, Dimethyl sulfoxide; DTT, Dithiothreitol; ECs, Emerging contaminants; EDs, Endocrine Disruptors; eNOS, Endothelial nitric oxide synthase; ER, Estrogen receptor; iNOS, Inducible nitric oxide synthase; JC-1, 1,1’,3,3’-tetraethyl-5,5’,6,6’-tetrachloroimidacarbocyanine iodide; MMP, Mitochondrial membrane potential; MnSOD, Manganese superoxide dismutase; MTT, 3(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; nNOS, Neuronal nitric oxide synthase; NO, Nitric oxide; NOx, Nitrogen oxides; OS, Oxidative stress; OXPHOS, Oxidative phosphorylation; PFOA, Perfluoro-octanoic acid; qPCR, Quantitative PCR Reaction; RNS, Reactive nitrogen species; ROS, Reactive oxygen species; SOD, Superoxide dismutase.
MCM and MA designed the research study. MX, BS, GA and FD performed the research. MCM and BS analyzed the data. MA, MCM and BB wrote the manuscript. EP and LS provided advice on data finalization and supervised manuscript preparation. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript.
Not applicable.
We warmly acknowledge Prof. Paolo Sarti (Department of Biochemical Science “A. Rossi Fanelli”, Sapienza University of Rome) for promoting and supporting this project. Dr. Mario Carere (Department of Environment and Health, Istituto Superiore di Sanità, Rome, Italy) is acknowledged for fruitful discussion and methodological advices on environmental pollutants. Prof. Luciana Mosca (Department of Biochemical Science “A. Rossi Fanelli”, Sapienza University of Rome) is gratefully acknowledged for providing us the HaCaT cell line.
Work supported by Regione Lazio of Italy (FILAS-RU-2014-1020 to prof. Paolo Sarti). Sapienza Progetti di Ateneo to M.A. RP11715C819AF6BA. Ph.D. fellowship of M.Xh. was funded by the Enrico and Enrica Sovena Foundation (Italy).
Luciano Saso is serving as one of the Guest Editors of the special issue “Modulation of oxidative stress: Biochemcal and pharmacological aspects”, in this journal. We declare that Luciano Saso had no involvement in the peer review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process of this article was delegated to Josef Jampílek.
Apart from what is mentioned above, all the authors declare that there is no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.