









1 Department of Cardiology, Henan Provincial Chest Hospital, 450000 Zhengzhou, Henan, China
Abstract
Background: Angiogenesis is the formation of blood vessels by sprouting from mature blood vessels and is regulated by multiple factors; however, the role of centrosome and centrosomal proteins (CEP) in angiogenesis needs further study. centrosomal protein 135 (CEP135) is an important centrosome-associated protein that can affect a variety of cellular processes, such as the cell cycle, but its effect on angiogenesis is still unknown. Methods: Tube formation and in vivo angiogenesis assays were performed to confirm the effects of CEP135 on endothelial cell (EC) angiogenesis in vitro and in mice. Cell counting kit-8 (CCK-8), 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), flow cytometry (FCM) and immunoblot assays were performed to confirm the effects of CEP135 on the proliferation and cell cycle of endothelial cells. Wound healing, transwell, and fluorescence staining were performed to confirm its effects on EC cell migration, polarization, and spindle orientation, and a tubulin turbidity assay was performed to confirm its effects on microtubule stabilization. Results: We conducted a series of experiments to explore the potential role of CEP135 in angiogenesis. CEP135 siRNA obviously inhibits angiogenesis in vivo compared with the control. CEP135 could mediate cell proliferation and the cell cycle by mediating spindle orientation. Moreover, we showed that CEP135 is involved in the regulation of angiogenesis by affecting the migration of endothelial cells using wound closure and transwell assays. We further demonstrated that CEP135 promotes endothelial polarization and microtubule stability, thus affecting cell migration. Conclusions: CEP135 was involved in the polarization of centrosomes, which is important for the migration of human umbilical vein ECs (HUVECs). These findings indicated that CEP135 may promote the polarization of HUVECs and accelerate migration, which in turn promotes angiogenesis.
Keywords
- angiogenesis
- CEP135
- migration
- polarization
- endothelial cells
Angiogenesis, which is the process relating to the formation of new blood vessels, is a highly dynamic process affecting degradation of the basement membrane, proliferation, directed migration of endothelial cells (ECs) and stabilization of new blood vessels [1, 2]. Angiogenesis generally occurs in diverse physiological and pathophysiological processes and is mediated by the balance between a number of molecules, such as vascular endothelial growth factor (VEGF) [3, 4]. During angiogenesis, proliferation and migration are critical and can promote the development of new blood vessels [5, 6]. Although several studies have reported that centrosomes can affect numerous cell activities, there is little information regarding the association between centrosomes and the regulation of angiogenesis [5, 6, 7].
Centrosomes have been found in ECs; however, their function in blood vessels remains incompletely defined [8]. Centrosomes in ECs are known to be critical in blood vessel development [9]. In addition, ECs are highly responsive to blood flow, and centrosomes are involved in sensing fluid shear stress [10]. Endothelial centrosomes may also participate in vascular mural cell recruitment by activating the Notch pathway [11]. EC centrosomes have also been reported to be involved in the detection of shear forces during vascular development, which can affect the progression of atherosclerosis by the inhibition of proatherosclerotic signaling in the aorta [12]. Although centrosomes are widely involved in the formation and maintenance of blood vessels, their possible roles and the effects of multiple centrosomal proteins on angiogenesis are still unclear [13].
The centrosomal protein 135 (CEP135) gene encodes a centrosomal protein [14]. In addition, a Bld10 mutant in Chlamydomonas reinhardtii has been shown to result in disorganized microtubules and the absence of basal bodies [15]. In cultured cells, CEP135 can localize to the cartwheel [16], and CEP135 has been reported to play a role in centriole biogenesis and particularly in central pair assembly [17]. Additionally, CEP135 gene mutations have been shown to cause primary microcephaly in mice [17]. CEP135 has also been shown to be amplified and mutated in aggressive breast tumor samples, suggesting that CEP135 may serve as a potential candidate oncogene [17]. These previous findings indicated that CEP135 is critical in the functions of the centrosome; however, its possible effects on the angiogenesis of ECs remain unclear.
In the present study, CEP135 small interfering (si)RNA-transfected cells were used to reveal the role of CEP135 in angiogenesis, which was further investigated in nude mice. The present study provides novel insight into how centrosome proteins may regulate cell processes and reports a novel mechanism for the regulation of angiogenesis.
The experiments using primary human umbilical vein ECs (HUVECs) were approved by
the Ethics Committee of the Chest Hospital of Henan Province (Zhengzhou, China).
Primary HUVECs (cat. no. 8000) were obtained from ScienCell Research
Laboratories, Inc., and cultured in RPMI-1640 medium supplemented with 10% FBS
(both from Thermo Fisher Scientific, Inc., Waltham, MA, USA) and 1% penicillin
and streptomycin in a 5% CO
The mouse CEP135 and control siRNAs were also synthesized by Guangzhou RiboBio
Co., Ltd. and used to transfect mice during the in vivo angiogenesis
assays. The sequences were as follows: siCEP135, 5
Antibodies against
The experiment was performed according to a previous study [18]. All animal procedures were approved by the Institutional Animal Care and Use Committee of the Chest Hospital of Henan Province (approval no. IRM-DWLL-2020172). A total of eight female nude mice (age, 8 weeks; weight, 22–24 g; n = 4 mice/group) were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. The experiment was performed according to a previous study [19]. The mice were sacrificed by cervical dislocation, and the lack of heartbeat was confirmed to validate death. The mice were monitored once a day before the experiment began and twice a day until the experiment ended.
Angiogenic capacity in mice was quantified with an in vivo angiogenesis assay kit (Trevigen, Inc., Gaithersburg, MD, USA). Angioreactors were filled with a small amount (20 µL) of basement membrane preparation premixed with angiogenic factors and filled with VEGF, heparin, and fibroblast growth factor-2 (FGF-2). Positive (only contains these factors) and negative controls (without these factors and siRNAs) were also set. The experimental groups included these factors and siRNAs. Mouse siRNAs at a final concentration of 2 µmol/L were then added to the angioreactors, and the transfection was finished 11 days after the transplant. The angioreactors were incubated for 1 h at 37 °C to induce gel formation and then implanted into the dorsal flank of 8-week-old nude mice. After 11 days, all of the mice were sacrificed by cervical dislocation, and the lack of heartbeat was confirmed to validate death. The angioreactors were ~5 mm long, and the mice had no adverse reactions after implantation. Cells were recovered from the angioreactors and stained with FITC-lectin at room temperature for 30 min, which stains EC glycoproteins. The relative fluorescence intensity was measured. The mRNA levels and density were calculated according to the average value of the 4 nude mice in each group.
TRIzol® (cat. no. 15596026; Invitrogen; Thermo Fisher
Scientific, Inc.) reagent was used to isolate RNA from HUVECs. Total RNA was
reverse transcribed into cDNA at 42 °C for 1 h using M-MLV reverse
transcriptase (cat. no. M1701; Promega Corporation, Madison, WI, USA). The
2
HUVECs were lysed in a buffer containing 1% Triton X-100, 150 mM NaCl and 50 mM
Tris (pH 7.5). Proteins were then transferred onto polyvinylidene difluoride
membranes (Millipore Sigma), which were blocked at room temperature for 2 h in
Tris-buffered saline containing 0.2% Tween 20 and 5% nonfat milk. The membranes
were then incubated with antibodies against CEP135 (1:500; cat. no. ab75005;
Abcam), CDK4 (1:500; cat. no. ab108357; Abcam), cyclin D1 (1:500; cat. no.
ab16663; Abcam) and
HUVECs (10
For immunofluorescence microscopy, 10
HUVECs were added to 96-well plates (1000 cells/well) and maintained for 48 h at 37 °C. Cells were subsequently treated with MTT agent for 4 h at 37 °C and washed with PBS. Formazan was then dissolved using 200 µL DMSO.
HUVECs were plated in 96-well plates to a density of 1000 cells/well. Cells were then treated with CCK-8 for 4 h at 37 °C and measured at 450 nm wavelength.
HUVECs transfected with siRNAs for 48 h were plated onto glass coverslips until 100% confluence. A 10-µL pipette tip was used to create a scratch, after which cells were washed twice with phosphate buffer saline (PBS) to remove cell debris and cultured in serum-free media for another 24 h. Images of the wound were captured using a light microscope at 0 and 24 h to determine the extent of wound closure using ImageJ 9.0 software, and the wound healing percentage was calculated as follows: Healing area/total area.
The proportion of HUVECs (10
BD Falcon inserts (BD Biosciences, Inc., NJ, USA) were used as upper
chambers, and 24-well plates were used as lower chambers. Cell culture inserts
were coated with 50 µL 20% Matrigel at 37 °C for 30 min.
Subsequently, 10
HUVECs transfected with siRNAs for 48 h were plated and grown onto coverslips until 100% confluence. The HUVEC monolayer was scratched; 3 h after scratching, the cells were fixed with 4% paraformaldehyde for 30 min.
For tubulin stabilization assays, HUVECs were placed on ice for 0 and 30 min for depolymerization, followed by immunofluorescence detection of morphology after fixing the cells were fixed with 4% paraformaldehyde for 30 min.
Microtubules (green), centrosomes (red) and nuclei (blue) were stained with
Tubulin (5 mg/mL; Cytoskeleton, Inc., Califonia, USA) and purified His-CEP135 proteins (100 nM; Guidechem, Nanjing, China) were added to a 96-well plate on ice and then transferred to a spectrophotometer. The polymerization was monitored by measuring the absorbance (350 nm). Spontaneously assembled microtubules with or without His-CEP135 (100 nM) at 37 °C were diluted with PBS, followed by shifting the temperature from 20 °C to 4 °C, according to a previous study [21].
Data were analyzed using GraphPad 8.0 software (GraphPad Software, Inc., La
Jolla, CA, USA). Error bars represent the mean
To investigate whether CEP135 is essential for angiogenesis, the present study examined the effect of CEP135 siRNAs on vascular endothelial tube formation in vitro. Two different siRNAs were used to target the CDS region of CEP135 mRNA and therefore induce the knockdown of CEP135, and both were able to effectively knockdown CEP135 expression in HUVECs (Fig. 1A,B). After plating cells for 3 or 6 h, a tubular network of interconnecting branches was observed in the control siRNA group; in contrast, fewer tubes were detected in the CEP135 siRNA groups (Fig. 1C). The accumulated tube length and node numbers at 3 h were measured as an index of angiogenesis (Fig. 1D,E). In addition, a CEP135 overexpression plasmid was transfected into HUVECs, and its efficiency was confirmed by western blotting (Supplementary Fig. 1). CEP135 overexpression had modest effects on the angiogenesis of HUVECs, with only slight changes in tube length and node numbers (Fig. 1F,G and Supplementary Fig. 2). However, overexpression of CEP135 rescued the inhibition of angiogenesis in HUVECs caused by CEP135 knockdown (Fig. 1H,I). These findings indicated that CEP135 was essential for angiogenesis.
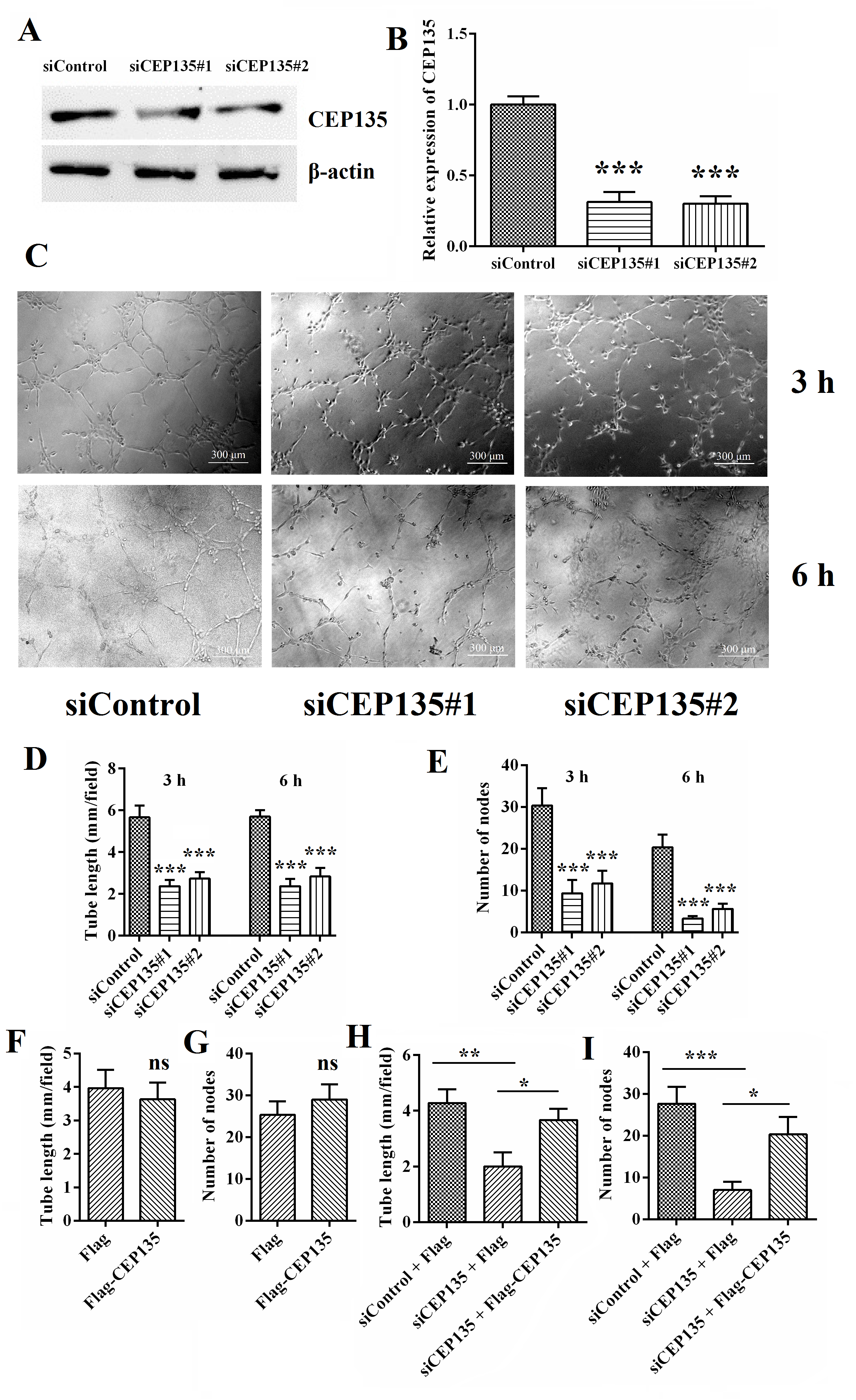 Fig. 1.
Fig. 1.CEP135 depletion leads to angiogenic defects in HUVECs. (A,B)
Western blot analysis of CEP135 and
To verify the effects of CEP135 on HUVEC angiogenesis in vitro, the present study assessed the effect of CEP135 on angiogenesis in a mouse model using semiopen angioreactors that were filled with extracellular matrix-containing angiogenic factors implanted into 4 nude mice per group (Fig. 2A). A total of 12 days after implantation, the silencing efficiency of siCEP135 was detected by qPCR (Fig. 2B). Vascular structures were identified in the angioreactors (Fig. 2C, positive control), whereas no vascular structures were detected in angioreactors without any angiogenic factors (Fig. 2C, negative control).
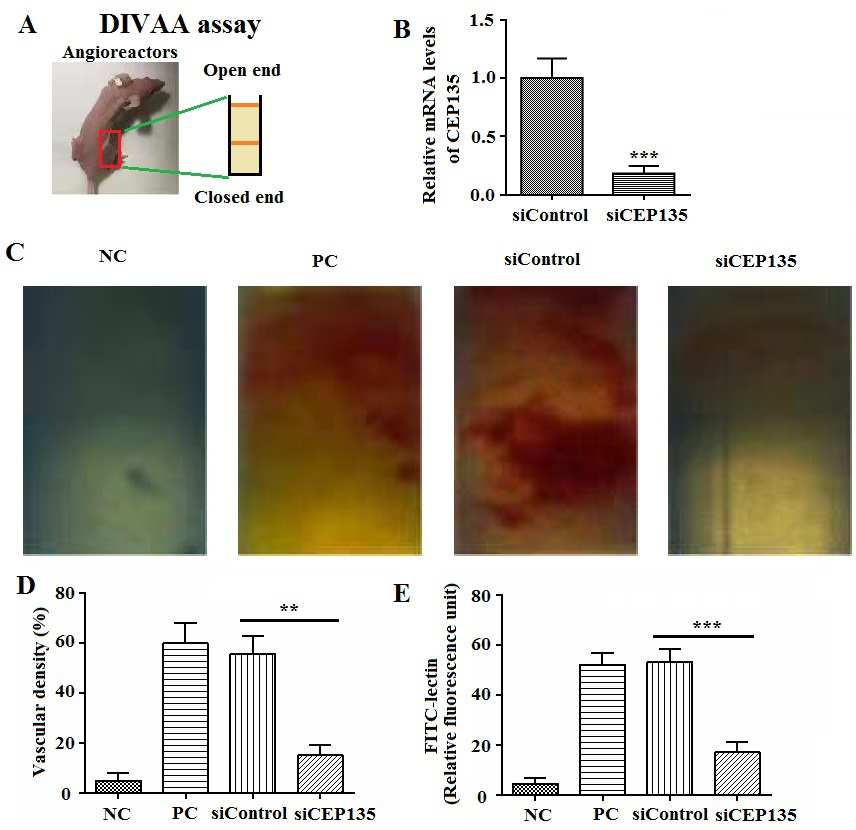 Fig. 2.
Fig. 2.CEP135 knockdown leads to angiogenic defects in vivo.
(A) Representative image of an athymic nude mouse with a semiclosed angioreactor.
(B) The efficiency of CEP135 knockdown was measured through quantitative PCR. (C)
Vascular growth for 12 days. In siRNA experiments, siCEP135 or siControl. (D)
Vascular density and (E) angiogenic responses were semiquantified by FITC-lectin
staining. Error bars indicate SEM.
Notably, CEP135 ablation in the angioreactors markedly inhibited new vascular
growth induced by heparin, VEGF and FGF-2, whereas control siRNA had no effect on
vascular growth into the angioreactors (Fig. 2C). To examine the angiogenic level
in vivo, the vascular density was assessed and was shown to be
significantly decreased in the CEP135 siRNA-treated group (Fig. 2D).
Subsequently, the cells were removed from the angioreactors and stained with
FITC-lectin, an EC marker. As shown in Fig. 2E, the fluorescence intensity of
cells in the siCEP135 group was significantly decreased compared with the control
(p
Since CEP135 affected EC angiogenesis, its effects on the proliferation of
HUVECs were assessed. Through CCK-8 and MTT assays, it was demonstrated that
CEP135 knockdown markedly suppressed the proliferation and survival of ECs, as
indicated by a decreased optical density value (Fig. 3A,B). These results
indicated that CEP135 knockdown inhibited cell proliferation. Subsequently, the
effects of CEP135 on cell cycle progression were analyzed by flow cytometry.
Notably, CEP135 knockdown led to cell cycle arrest (Fig. 3C and
Supplementary Fig. 3). In addition, the protein expression levels of
CDK4 and cyclin D1, two markers of the cell cycle, were decreased following
CEP135 knockdown in ECs (Fig. 3D–F; p
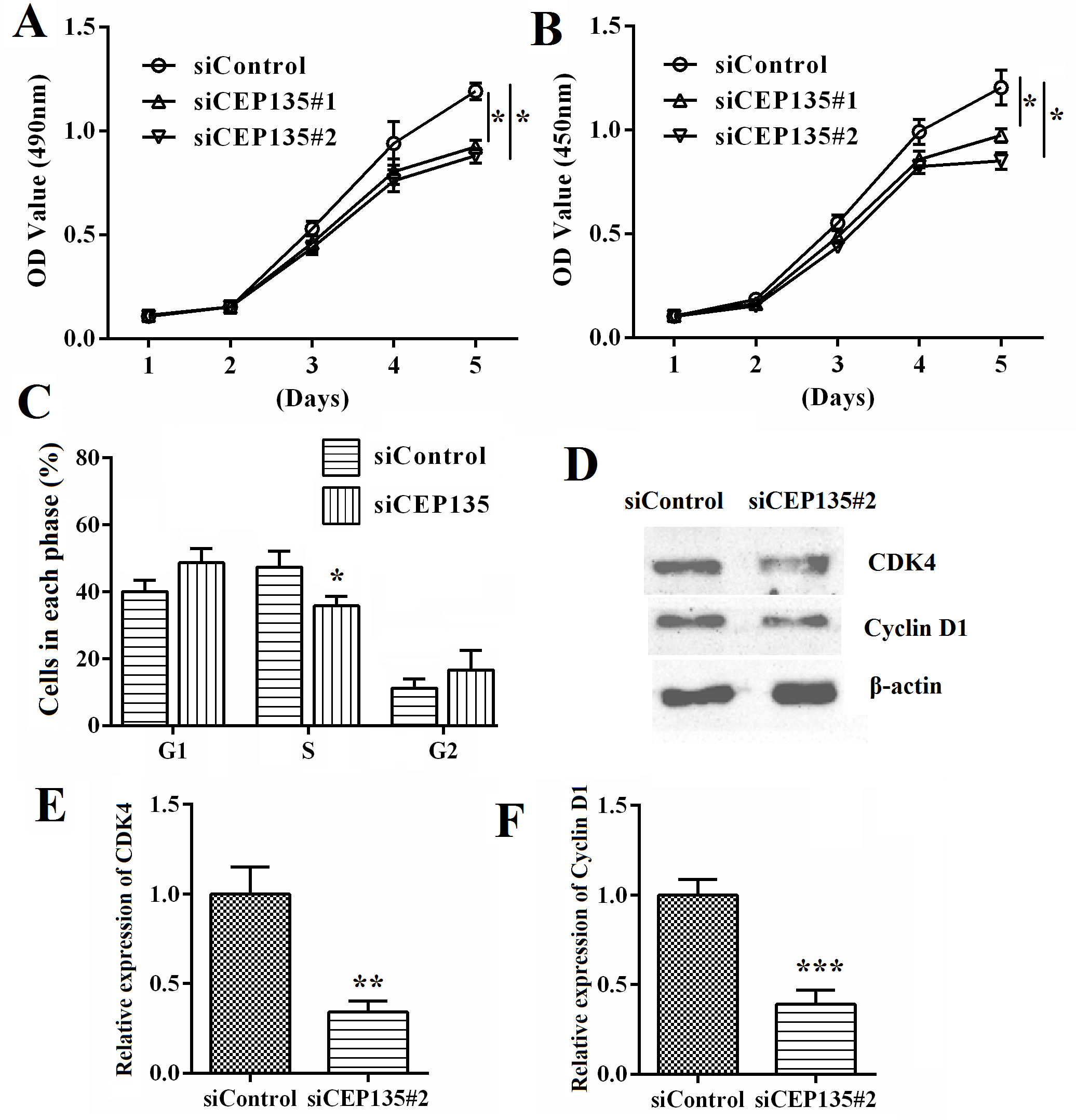 Fig. 3.
Fig. 3.CEP135 knockdown suppresses HUVEC proliferation and cell cycle
progression. (A) MTT and (B) Cell Counting Kit-8 assays revealed the effects of
CEP135 on the proliferation of HUVECs posttransfection. (C) Flow cytometry showed
the effect of CEP135 on the cell cycle progression of HUVECs posttransfection
with the transfection of control and human CEP135 siRNA#1, and the percentage of
cells at different stages of the cell cycle was analyzed. (D) Western blot
analysis showed the effects of CEP135 on the expression levels of (E) CDK4 and
(F) Cyclin D1 upon transfection with control and CEP135 siRNA#1. Each assay was
performed 3 times. One-way ANOVA was used for hypothesis testing for significance
in A and B. Error bars indicate SEM.
CEP135 is a centrosomal protein that has the potential to affect mitosis [15]. Therefore, the present study measured the effects of CEP135 on cell division. Spindle orientation is important for many developmental processes, including cell division, epithelial tissue homeostasis and regeneration. Spindle orientation was determined by measuring the angle between the spindle axis and the substratum. Notably, knockdown of CEP135 resulted in an increase in spindle angle, indicative of defective spindle orientation, whereas spindle length was not affected (Fig. 4A–E). The model was shown in Fig. 4A. These findings suggested that CEP135 depletion led to defects in spindle orientation in ECs.
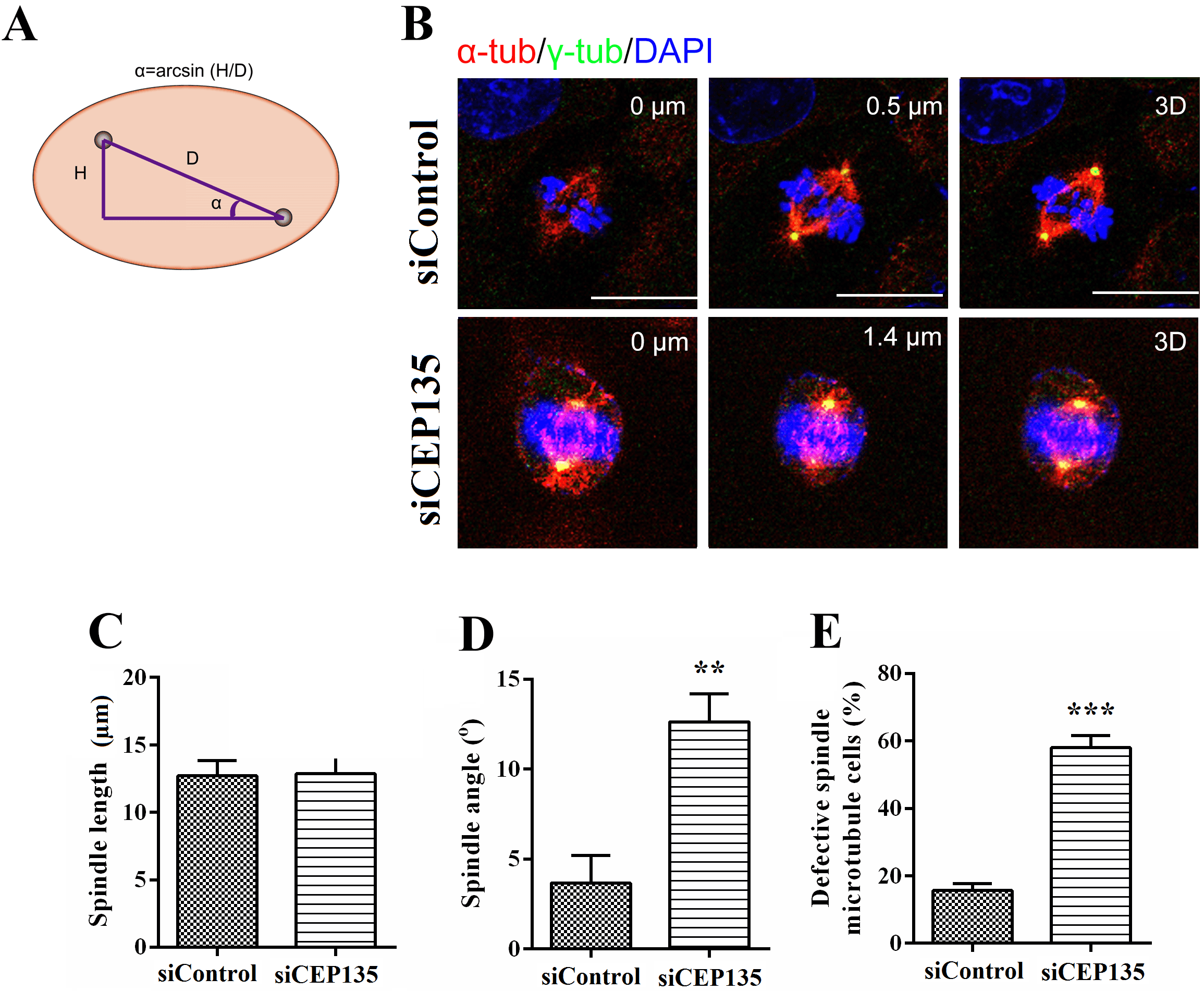 Fig. 4.
Fig. 4.CEP135 knockdown affects the spindle orientation of HUVECs. (A)
Scheme depicting spindle angle (
The present study then assessed the effect of CEP135 on HUVEC migration. A HUVEC monolayer was scratched with a pipette tip, and closure of the wounded area was documented to evaluate the capacity of cell migration. The wounded area in the control group was fully recovered after 24 h as a consequence of directed cell migration (Fig. 5A,B), suggesting the effects on cell migration. Transwell assays also revealed that CEP135 knockdown suppressed the invasion of HUVECs (Fig. 5C,D).
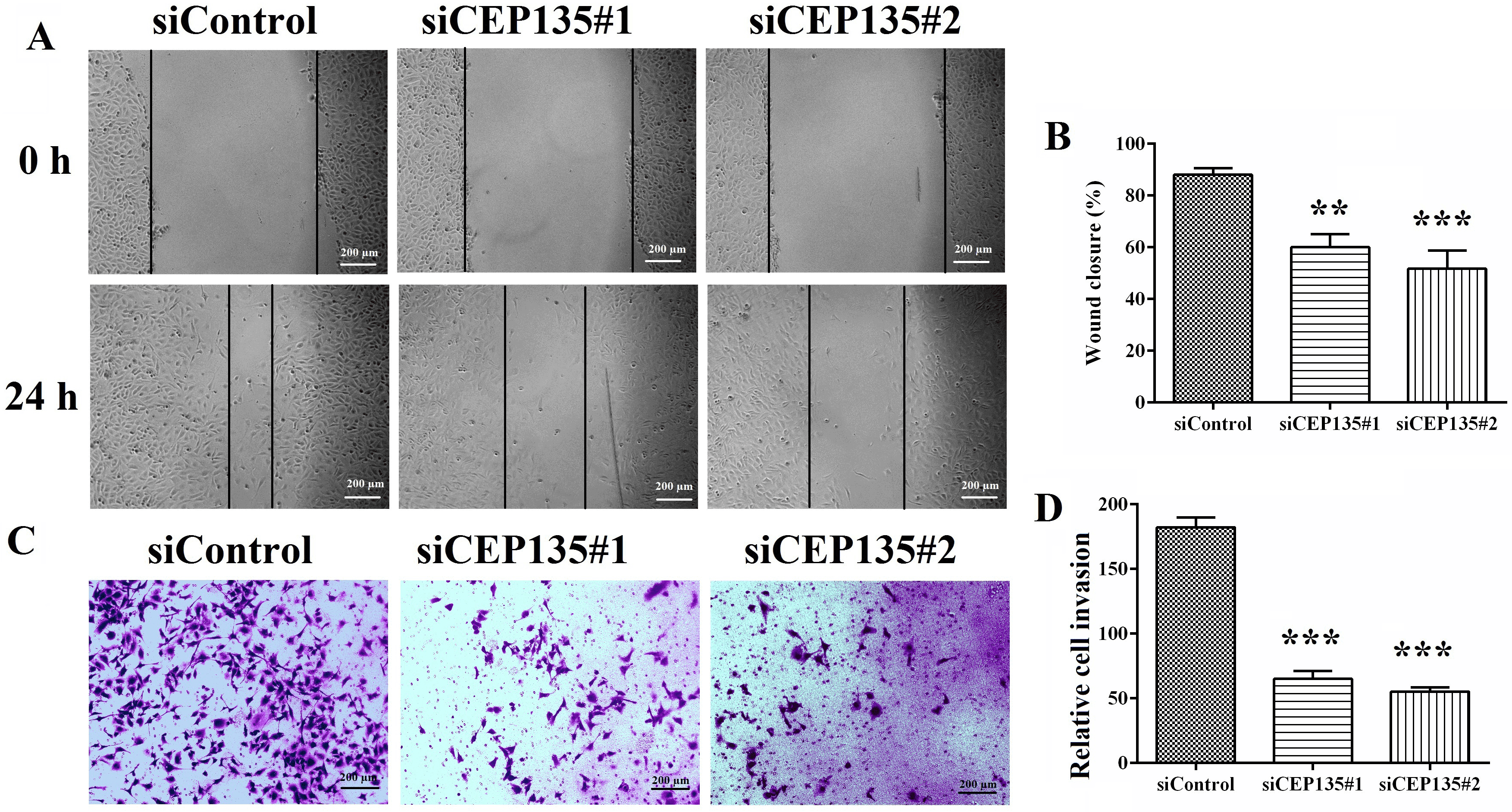 Fig. 5.
Fig. 5.CEP135 is critical for vascular endothelial cell migration. (A)
HUVECs transfected with siControl or siCEP135 were scratched, and images of the
wound margins were captured after 0 and 24 h. (B) The extent of wound closure was
semiquantified by measuring the wound area. (C) Cells invading the underside of
the insert were stained with crystal violet. (D) Semiquantification of the number
of invaded cells. Data were analyzed using one-way ANOVA. Error bars indicate
SEM. Scale bar, 200 µm.
Polarization is a critical step in cell migration that involves the rearrangement of microtubules and reorientation of the centrosome [15]. HUVECs transfected with control or CEP135 siRNAs were scratched, and the cells were fixed 3 h later. In the control siRNA group, cells at the wound margin demonstrated a typical polarized structure, with the centrosome positioned between the nucleus and the leading edge (Fig. 6A,B). In contrast, the polarized morphology was suppressed in cells from the siCEP135 group (Fig. 6A). By semiquantifying the percentage of polarized cells, it was revealed that knockdown of CEP135 expression significantly inhibited cell polarization (Fig. 6C,D). In addition, the results indicated that CEP135 was critical for HUVEC migration and polarization by affecting the polarization angles (Fig. 6D).
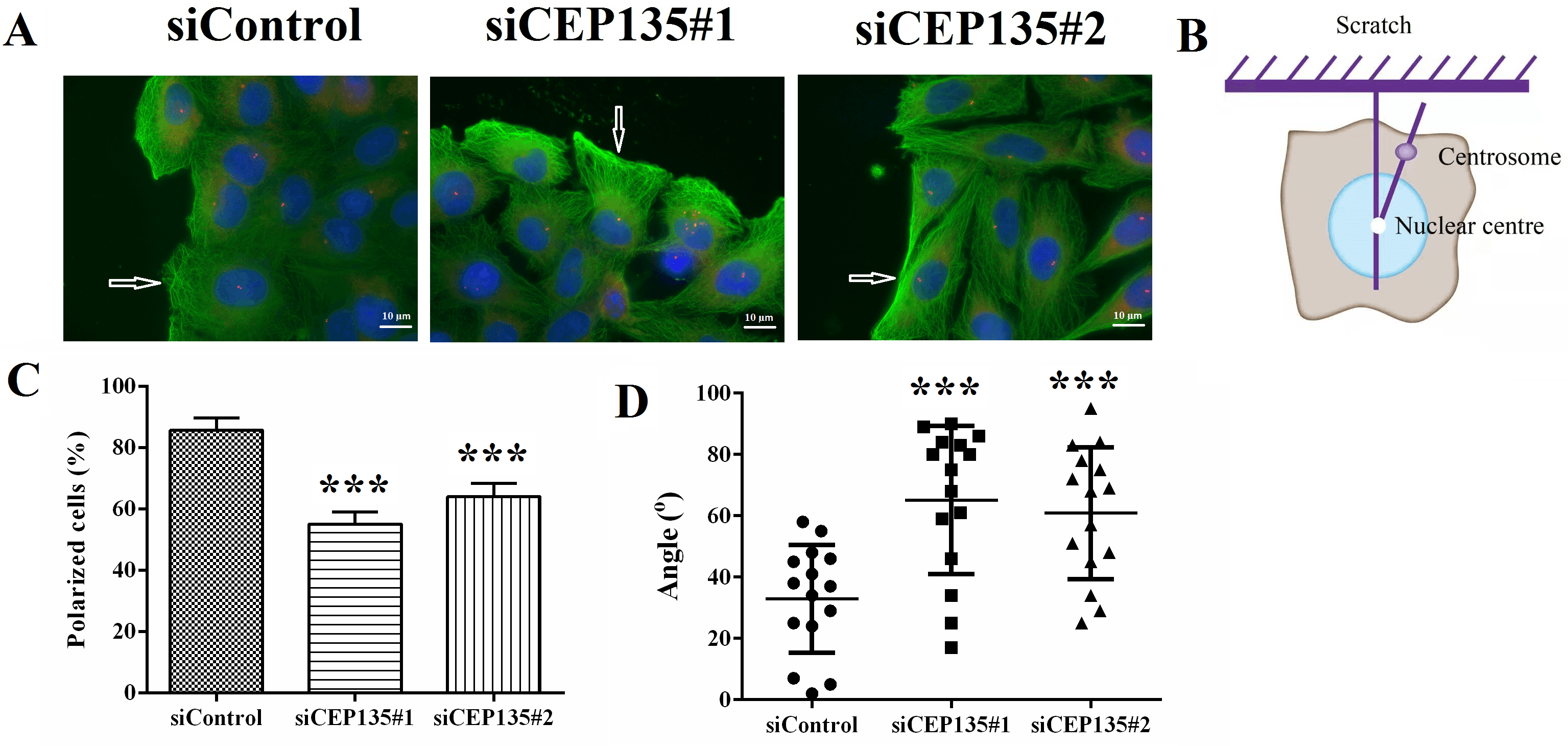 Fig. 6.
Fig. 6.CEP135 mediates vascular endothelial cell polarization. (A)
Immunofluorescence microscopy images of siControl- and siCEP135-transfected
HUVECs.
The present study assessed the mechanism underlying the effects of CEP135 on EC
proliferation and migration. The present study investigated whether CEP135 was
involved in the regulation of microtubule stabilization, which is critical for
cell polarization and migration. CEP135 knockdown significantly inhibited
microtubule stabilization when cells were placed on ice for 30 min (Fig. 7A,B;
p
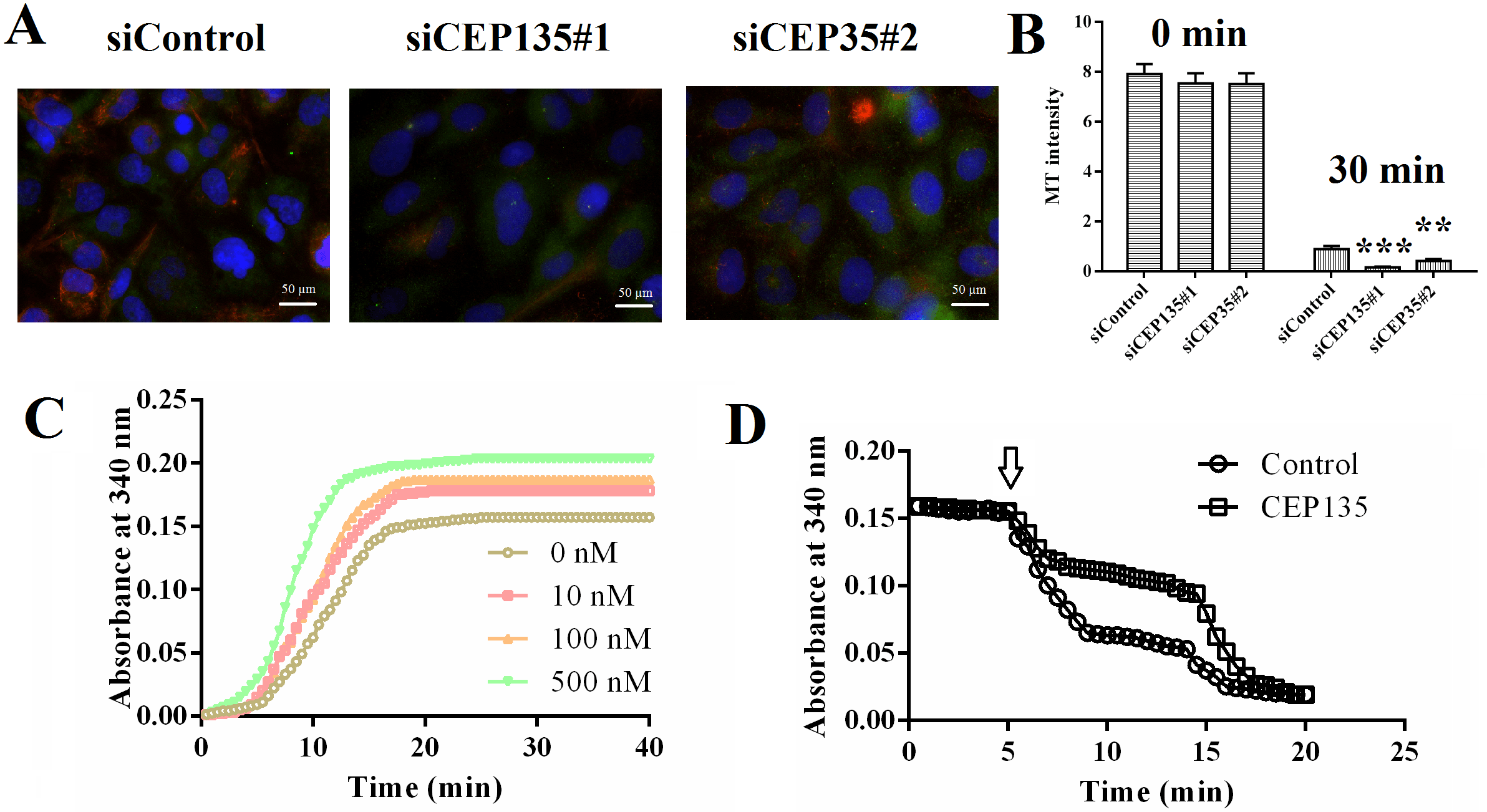 Fig. 7.
Fig. 7.CEP135 is critical for vascular endothelial cell MT
stabilization. (A) Immunofluorescence microscopy images of siControl- and
siCEP135-transfected HUVECs.
Angiogenesis refers to the process of sprouting of new blood vessels from existing vessels, which is regulated by a number of proangiogenic and inhibitory factors [4]. In adults, angiogenesis is virtually nonexistent [4]. This process often occurs in pathological processes, such as retinopathy of prematurity, tumor development and metastasis. Despite some understanding of angiogenesis, the relationship between centrosomes and endothelial angiogenesis is still poorly understood [7]. At present, some proteins that can be located in the centrosome, such as CEP70, have been reported to promote angiogenesis, and an in-depth understanding of this process is helpful to further reveal the pathogenesis of angiogenesis and related diseases [22]. The present study revealed that a centrosomal protein, CEP135, could serve as a regulator of angiogenesis.
Centrosomal proteins have multiple cellular functions in ECs [23]. They can mediate several microtubule-related processes, such as migration and directed cell division, by mediating microtubule dynamics and stabilization [8]. The present study demonstrated that a centrosomal protein, CEP135, could serve as a regulator of angiogenesis. Through in vitro and in vivo assays, it was demonstrated that knockdown of CEP135 suppressed HUVEC angiogenesis. Further in vitro assays showed that CEP135 knockdown suppressed HUVEC proliferation and cell cycle progression, possibly via alterations to the spindle. In addition, knockdown of CEP135 inhibited cell migration by mediating cell polarization. Furthermore, CEP135 was revealed to mediate microtubule stabilization, thereby mediating cell migration, proliferation and angiogenesis of HUVECs (Fig. 8). It was therefore hypothesized that CEP135 could serve as a promising angiogenic regulator.
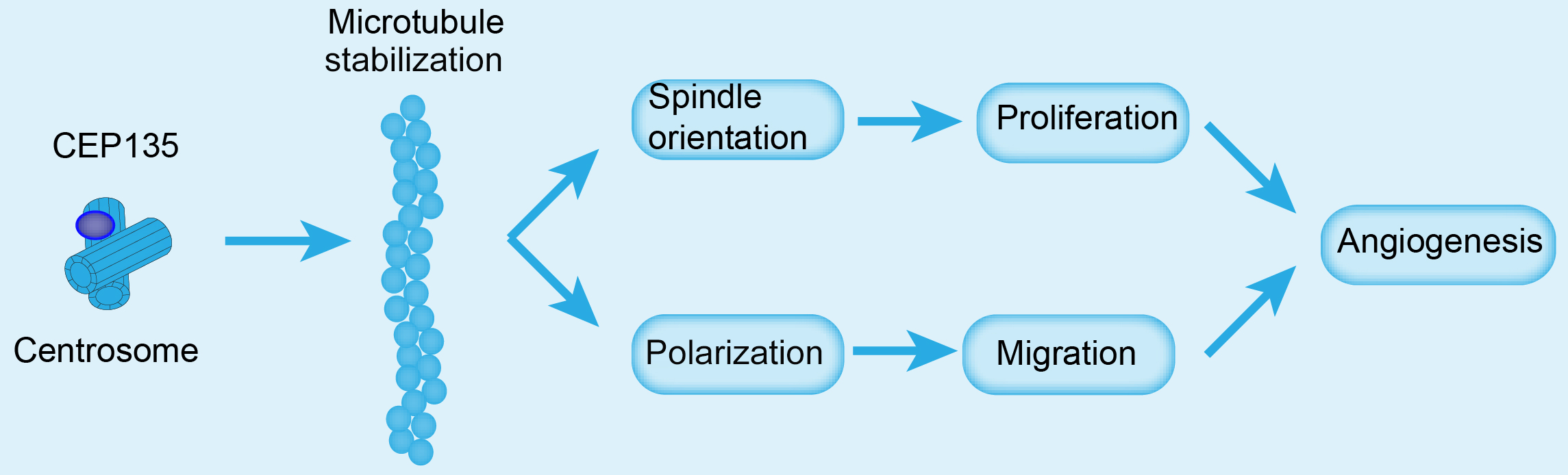 Fig. 8.
Fig. 8.Molecular model for the role of CEP135 in angiogenesis. CEP135 was involved in the polarization of centrosomes, which is important for HUVEC migration. These findings indicated that CEP135 may promote the polarization of HUVECs and accelerate migration, which in turn promotes angiogenesis. CEP135, centrosomal protein 135; HUVECs, human umbilical vein endothelial cells.
The role of CEP135 in multiple cellular processes has been well documented [16, 17]. For example, depletion of CEP135 has been reported to lead to a disorganized interphase and multiple and fragmented centrosomes with disorganized microtubules [24]. Similarly, the present study revealed that the knockdown of CEP135 led to decreased microtubule stabilization in HUVECs and that several microtubule-dependent cellular processes were further affected. The knockdown of CEP135 has been revealed to suppress cell division and cause disordered neuronal cell polarity, which is essential for neuronal migration [25]. Similarly, the present study also revealed the effects of CEP135 on HUVEC polarization. Furthermore, mutations in CEP135 were reported to cause primary microcephaly and subcortical heterotopia in a microtubule-dependent manner [24]. Other studies have also revealed the multiple cellular functions of CEP135. A homozygous CEP135 mutation was revealed to be associated with multiple morphological abnormalities of the sperm flagella [17]. In addition, CEP135 was shown to be required to establish centrosome asymmetry in Drosophila neuroblasts [26]. All these previous studies have confirmed that CEP135 could serve as a promising target for the treatment of multiple diseases.
The present study also revealed the effects of CEP135 on the regulation of
microtubule stabilization. A previous study indicated that CEP135 contains a
two-stranded coiled-coil domain, which is critical for microtubule binding [27].
A previous study indicated that the loss-of-function mutation of CEP135 limited
the localization of other centrosomal proteins, including SAS-6, CPAP and
Several studies have indicated the effects of centrosome- or microtubule-related proteins on angiogenesis; however, to the best of our knowledge, the present study is the first to demonstrate that the centrosomal protein CEP135 could affect the angiogenesis of HUVECs by mediating proliferation and migration in a microtubule-dependent manner [29, 30]. Through in vitro assays, it was revealed that CEP135 mediated HUVEC angiogenesis by mediating proliferation and migration. Similarly, the in vivo assays confirmed its effects on angiogenesis.
Notably, cell polarization is the most important link in the early stage of cell migration and is mainly regulated by MTOCs and microtubules. After cell polarization, the microtubules rearrange, resulting in the movement of intracellular substances in the direction of migration through the microtubules [29, 30]. Therefore, the present study assessed the effect of CEP135 on cell polarization and confirmed its effect on migration.
CEP135, as a centrosomal protein, plays an important role in centrosomes and MTOCs and participates in the regulation of microtubule stability. Due to the effect of CEP135 on microtubule-related functions, HUVEC proliferation and migration are further affected, both of which are regulated by microtubules.
In conclusion, the present study revealed that CEP135 mediated the angiogenesis of HUVECs in vitro and in mice. CEP135 regulated cell proliferation and migration by mediating spindle orientation and cell polarization. Furthermore, the effects of CEP135 on the stabilization of microtubules were confirmed. The present study indicated the role of CEP135 in angiogenesis and that it could serve as a critical regulator during the process of angiogenesis.
Additional supporting information may be found in the online version of the article at the publisher’s website. And the datasets used and/or analyzed during the present study are available from the author on reasonable request.
KW, XW, FZ and XZ carried out molecular biology experiments and drafted the manuscript. KW, QZ, SK and PM designed the study and performed statistical analysis. KW, GW, WW and XZ conceived the study, participated in its design, coordinated the study and drafted the manuscript. KW and XZ confirm the authenticity of all the raw data. All authors contributed to editorial changes in the manuscript. All authors have read and approved the final manuscript.
All animal procedures were approved by the Institutional Animal Care and Use Committee of the Chest Hospital of Henan Province (approval no. IRM-DWLL-2020172).
Not applicable.
This research received no external funding.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.