

 , Thuluz Meza-Menchaca 2, Oswaldo Guzmán-López 3, Eliezer Cocoletzi Vásquez 4, Jorge Ricaño-Rodríguez 5,*
, Thuluz Meza-Menchaca 2, Oswaldo Guzmán-López 3, Eliezer Cocoletzi Vásquez 4, Jorge Ricaño-Rodríguez 5,*
1 Laboratory of Agricultural Chemistry, University of Veracruz, 91040 Xalapa, Veracruz, Mexico
2 Faculty of Medicine, University of Veracruz, 91010 Xalapa, Veracruz, Mexico
3 Faculty of Chemical Sciences, University of Veracruz, 96538 Coatzacoalcos, Veracruz, Mexico
4 Faculty of Biology, University of Veracruz, 91040 Xalapa, Veracruz, Mexico
5 Center for Ecoliteracy and Knowledge Dialogue, University of Veracruz, 91060 Xalapa, Veracruz, Mexico
Abstract
In most ecosystems, plants establish complex symbiotic relationships with organisms, such as bacteria and fungi, which significantly influence their health by promoting or inhibiting growth. These relationships involve biochemical exchanges at the cellular level that affect plant physiology and have evolutionary implications, such as species diversification, horizontal gene transfer, symbiosis and mutualism, environmental adaptation, and positive impacts on community structure and biodiversity. For these reasons, contemporary research, moving beyond observational studies, seeks to elucidate the molecular basis of these interactions; however, gaps in knowledge remain. This is particularly noticeable in understanding how plants distinguish between beneficial and antagonistic microorganisms. In light of the above, this literature review aims to address some of these gaps by exploring the key mechanisms in common interspecies relationships. Thus, our study presents novel insights into these evolutionary archetypes, focusing on the antibiosis process and microbial signaling, including chemotaxis and quorum sensing. Additionally, it examined the biochemical basis of endophytism, pre-mRNA splicing, and transcriptional plasticity, highlighting the roles of transcription factors and epigenetic regulation in the functions of the interacting organisms. These findings emphasize the importance of understanding these confluences in natural environments, which are crucial for future theoretical and practical applications, such as improving plant nutrition, protecting against pathogens, developing transgenic crops, sustainable agriculture, and researching disease mechanisms. It was concluded that because of the characteristics of the various biomolecules involved in these biological interactions, there are interconnected molecular networks in nature that give rise to different ecological scaffolds. These networks integrate a myriad of functionally organic units that belong to various kingdoms. This interweaving underscores the complexity and multidisciplinary integration required to understand plant–microbe interactions at the molecular level. Regarding the limitations inherent in this study, it is recognized that researchers face significant obstacles. These include technical difficulties in experimentation and fieldwork, as well as the arduous task of consolidating and summarizing findings for academic articles. Challenges range from understanding complex ecological and molecular dynamics to unbiased and objective interpretation of diverse and ever-changing literature.
Keywords
- chemotaxis
- endophytism
- iRNA
- microbial signaling
- miRNA
- plant growth-promoting bacteria
- plant pathogenesis
- quorum sensing
- secondary metabolites
- symbiosis
In megadiverse ecosystems, many plant species coexist with a wide range of microorganisms, and their relationships can be beneficial, detrimental, or neutral. These interactions allow plants to establish connections with other microorganisms that improve or compromise their survival. Experts have examined how vegetables accurately perceive and respond to signals from their microbial environment within this context. The balance they must maintain between utilizing metabolic and defensive benefits provided by certain microorganisms and activating their defenses against antagonistic microbes has been debated for several decades [1, 2]. From this perspective, understanding the metabolic pathways in plants is essential in the field of biotechnology, as they play a key role in signal transduction and the immune mechanisms of the flora.
Over the past several decades, scientists have attempted to explain why and how plants and microorganisms interact differently in their environment. One of the main reasons for this is their genotype, which includes adaptation and survival instructions [2]. As ecosystems depend on collaboration between plants and microorganisms, this connection is based on intrinsic molecular interactions. For example, through photosynthesis, plants convert sunlight into chemical energy, producing oxygen and essential organic compounds, such as glucose. These biomolecules sustain the growth of several microorganisms. On land, bacteria and fungi decompose organic matter using specific enzymes, thereby releasing nutrients in less complex forms that plants absorb [3]. However, some plant–microorganism relationships involve other kinds of molecular dynamics. Nitrogen fixation is one example of this phenomenon. During this process, symbiotic bacteria in the root nodules of legumes convert atmospheric nitrogen into ammonia. This metabolic pathway, which is vital for plants, involves nitrogenase enzymes. In this sense, mycorrhizal fungi improve the ability of many vegetal specimens to absorb nutrients, particularly phosphorus, by employing hyphal networks that extend the root absorption area. These interactions are crucial for the nutrient cycle and maintaining balance in the ecosystem [3, 4].
Since the 1980s, significant discoveries have been made regarding the complex interplay between plants and the microbiota. As mentioned above, these findings reveal highly coordinated molecular networks that facilitate diverse biological interactions [4]. For instance, plant cells possess highly specialized structures that allow them to detect microbial characteristics. By recognizing these motifs, cells activate defenses or establish symbiotic metabolic pathways depending on the specific microbes [2, 3, 4]. These microorganisms also possess capabilities that enable RNA synthesis and the translation of peptide-mimicking molecules. From a starting point, these processes trigger reactions in plant cells, preparing the environment for harmonious coexistence and conferring an adaptive advantage to microbes. Specific differentiated plant structures, such as root nodules and symbiotic interactions with mycorrhizal fungi, function as catalytic agents at the metabolic level; thus, are considered key epicenters in organismic relationships, ranging from collaborative cooperation to biological antagonism [4]. The multifaceted nature of such relationships significantly affects the structural and functional aspects of flora-linked microbiomes, leading to adjustments and transformations in the ecological scenario within the plant organism or the immediate surrounding environment [5].
In the biological context, evolutionary trajectories have diversified over long geological periods within the botanical world, resulting in a resilient structure that enables harmonious coexistence between plants and microbes. In response to emerging biotechnological challenges, the beneficial properties of these microbial associations are ecologically responsible and promising [5]. The primary objective from both technological and sociocultural perspectives is to maximize agricultural productivity by addressing the biotic and abiotic challenges originating from living beings and the environment, respectively. When examining the complex interactions between beneficial microorganisms and their environment, a diverse and ever-changing landscape is found that requires a thorough investigation of regulatory processes in controlled settings [6]. Furthermore, the unique genetic traits of these organisms introduce uncertainty, raising questions about the defense mechanisms used by plants against harmful microorganisms and the strategies used by beneficial and antagonistic strains [4, 5, 6]. In view of the above, it is worthwhile to reflect on the following questions: How do plant immune systems adapt at the molecular and cellular levels to distinguish between beneficial and antagonists while preserving symbiotic relationships? What is the molecular basis for non-host resistance? How do resistance proteins activate cell death? How do pathogens evolve novel virulence factors? In this regard, the evolutionary history of interspecific interactions has been proposed to catalyze an understanding of how highly specialized immune mechanisms can distinguish themselves from a wide range of environmental signals and stimuli in a highly specific and efficient manner.
The extended period of evolutionary interactions between eukaryotic organisms and their symbiotic microorganisms has significantly influenced various areas of biology [7]. This relationship affects not only the well-being and functionality of plants but also the adaptability and function of microbes. From a long-term evolutionary perspective, this reciprocal phenomenon suggests a carefully regulated balance refined over time. These dynamics persistently affect natural selection mechanisms and adaptive decisions, affecting both photosynthetic organisms and microbes that maintain close symbiotic relationships [8]. From this point of view, advances in molecular genetics and related fields have deepened our understanding of the complex signaling networks that underlie plant defense mechanisms. These systems have evolved to manage interactions between individuals, as they are beneficial or pathogenic. Importantly, the crossover between biology and systems ecology has emerged as an interdisciplinary field, placing these mechanisms within an ecological and evolutionary framework [8, 9]. During plant–microbe interactions, a mutual recognition system is essential for proper physical and molecular coordination. These include the participation of Nod and Nif genes, molecules related to microbial communication, auxins and cytokinins, resistance genes (R genes), elicitors, and pathogen-associated recognition molecules (PAMPs). Since these interactions are shaped by the unique genetics of each plant specimen and the corresponding cellular adaptations, they result in regulated signal exchange. Thus, molecular connections ensure ideal functional integration in the joint environment, reflecting coevolution that has been perfected through numerous eras [5, 10].
Recent advances in genomics have revealed that plants have adapted to a wide range of biotic interactions that extend beyond their relationships with parasitic and herbivorous organisms, including beneficial symbionts. In particular, the signaling pathways that plants use to manage these relationships often show common characteristics, suggesting that their regulatory structures have evolved to defend against aggressors while improving symbiotic associations. Through several studies of this phenomenon, it is possible to understand better how plants respond to environmental factors and their consequential adaptability [11, 12].
Undoubtedly, derived from the above context, several research questions arise that are worth analyzing, among which we submit for consideration the following: What are the effects of various elements, including beneficial and antagonistic microbes, abiotic stressors, and microbial interplay, on plant–microbe interactions? How do these elements enhance our understanding of resistance mechanisms, such as effector-triggered immunity (ETI), pattern-triggered immunity (PTI), and non-host resistance? In applied science, how might we translate fundamental research into tangible advancements for new crops, particularly in understanding the molecular foundation of phenomena such as resistance protein-induced cell death? Considering the complex and dynamic inter-relations among plants, microbes, and their surroundings, how do these interactions affect plant health and disease? Furthermore, how can we leverage this knowledge to refine agricultural practices and improve crop resilience under evolving environmental conditions? Or how do these insights into binary plant-microbe interactions extrapolate to ecological contexts? As we can see, in an evolutionary framework, the driving forces behind the diversity and evolution of pathogen strategies, especially considering the varying effector numbers required by different pathogens, remain elusive.
Corresponding to the earlier questions, we propose at least five hypotheses that, in our opinion, delimit and refine part of our understanding of this context: (1) Plants have evolved intrinsic mechanisms to maintain a balance between fostering beneficial microbial relationships and defending against pathogens, significantly influenced by abiotic stress factors such as climate change; (2) insights from basic plant–microbe interaction research may be translated into practical agricultural applications, with a particular emphasis on understanding the complex and intertwined nature of the plant immune system, including the overlapping roles of effector-triggered and pathogen-triggered immunity; (3) the health and disease resistance of plants are profoundly affected by microbe–microbe interactions within their microbiome, and the diversity of pathogen effector strategies is a result of evolutionary pressures shaped by these interactions and the host environment; (4) the molecular basis of plant resistance, including non-host resistance and nucleotide leucine-rich (NLR)-mediated hypersensitive response (nucleotide-binding leucine-rich repeat), involves a variety of mechanisms that contribute to a plant’s ability to resist a wide range of pathogens; (5) the outcomes of plant–microbe interactions are contingent on the ecological context, with pathogen evolution and host adaptation driven by a combination of genetic, environmental, and ecological factors, leading to novel virulence activities and changes in disease dynamics.
Based on the above, this literature review aims to analyze and discuss the outstanding theoretical and practical background describing plant–microbe interactions in the context of molecular ecological compatibility. This was achieved through a comprehensive synthesis of the current literature, identification of research gaps and research questions, proposal of new hypotheses, delimitation of practical implications and applications, future perspectives and research directions, and their relevance to the context of biotechnology and sustainable development. Finally, we consider that the limitations in these types of studies are inherent to factors such as the difficulty in replicating environmental conditions, the complexity of microbial communities, challenges in genetic manipulation, dynamic and variable molecular–ecological interactions, limitations in imaging and visualization techniques, restrictions in modeling and computational simulation, technical problems in the analysis of massive data, ethical and legal aspects, the need for funding and obtaining monetary resources, and the impact of climate change. Nevertheless, objectively interpreting a continuously evolving array of diverse literature is equally demanding.
Understanding microbiome biology is based on the interactions between microorganisms, and as time passes, how they are studied evolves differently. Currently, scientists are studying more complicated and multi-specific microbial communities and natural environments compared to a more simplified single-species system. Due to meticulous research, biological interaction paradigms that cover a wide range of factors were identified, including competition for nutrients and intercellular communication through signaling cascades. For example, microbes coexist in biofilms and communities of bacteria that adhere to surfaces and form a matrix of polysaccharides, proteins, and DNA [12, 13]. These findings contribute to a more comprehensive understanding of microbial ecology. The importance of diversity in rhizosphere microbes cannot be understood, as it plays a crucial role in helping plants resist and outcompete other harmful microorganisms. Microbes use all types of strategies to defend themselves and simultaneously help plants. One strategy is to produce secondary metabolites, such as antibiotics, which prevent the growth of other biological entities [13].
Certain biomolecules kill undesirable bacteria (in different contexts). For instance, some Pseudomonas species produce 2,4-diacetylphloroglucinol (DAPG), a molecule with a potent biocontrol capacity due to its tricyclic phenolic structure [14]. Similarly, some Agrobacterium species produce agrocin, an effective antibiotic [15]. In addition to metabolites, many beneficial microbes compete for resources, limiting the likelihood of infection. Some release compounds that capture nutrients and metals in stressful situations and slow pathogen growth. Furthermore, the characteristics of the roots, such as reduced surface area and increased exudation, promote microbial diversity and activity in the rhizosphere. Therefore, by improving the supply of nutrients through increased root exudation and carbon content, it is possible to enrich the diversity of the microbiota further [16, 17, 18].
Another example is the biosynthesis of siderophores, iron chelators that play a vital role in biocontrol mechanisms by combining nutritional competence with the production of secondary metabolites [19]. These chelators are synthesized by a diverse array of organisms and exhibit various chemical properties, classified into several categories, i.e., hydroxamate siderophores, which contain acetylated or hydroxylated alkylated amines and are prevalent in ornithine-based fungi catechol siderophores, composed of hydroxyl and catechol units; carboxylate siderophores, which incorporate hydroxyl and carboxyl units; mixed siderophores, comprising catechol and hydroxamate components; chelators that integrate catechol and hydroxamate components [20]. In the context of iron deficiency in rhizospheres, microorganisms use siderophores to restrict the access of this mineral to pathogens. These molecules bind to iron ions and enhance their cellular uptake. For example, certain Pseudomonas species produce pyoverdin and pyochelin, which transport iron to the cell interior, thus preventing bacterial and fungal invasion of the roots of various crops [21, 22]. Although certain siderophores have a specific effect on their producing bacteria, others have a more widespread effect. Beyond their iron uptake function, these compounds also chelate other metals, including aluminum, copper, zinc, manganese, lead, and cadmium [23, 24]. Importantly, siderophores activate plant-resistance mechanisms and protect plants against various pathogens.
Microorganisms employ effective strategies to combat antagonists, mainly bacteria and fungi. One such approach is direct confrontation, largely mediated by the secretion of hydrolytic enzymes capable of degrading cellular structures. In particular, soil actinomycetes and Gram-positive bacteria parasitize and decompose fungal spores. These microorganisms interfere with the nutrient transport pathways between plants and fungi, resulting in greater bacterial colonization. The mechanism induces biotic stress in the fungus or even leads to its complete destruction through extracellular peptides. Hydrolytic enzymes (hydrolases), such as cellulases and chitinases, play a crucial role in the breakdown of essential fungal components, such as cellulose and chitin, into smaller fragments [25, 26]. Studies have confirmed these mechanisms at the molecular level, revealing that endochitinases and other proteases decrease fungal infections in crops, including cotton, wheat, and berries [27].
Certain microbes have developed strategies to combat pathogens in host plants, as demonstrated by Panwar et al. [28], whose research showed that Pseudomonas aureofaciens effectively counteracts certain strains of Gaeumannomyces graminis, a causal agent of wheat disease. This bacterium produces a toxin, phenazine-1-carboxamide (PCA), attributed to its potent antifungal activity. However, root exudates resulting from fungal infections promote the growth of Pseudomonas and other beneficial microorganisms. Furthermore, surrounding bacteria improve PCA production, producing robust biocontrol against pathogenic fungi [29]. Microbes often use the production and release of secondary metabolites to maintain a homeostatic equilibrium in their populations. Consequently, organisms that produce these molecules must regulate their release to ensure their effectiveness. In this sense, the production and detection of signaling elicitors play a critical role in controlling the release of antimicrobial agents when specific cellular concentrations are detected. This regulation is important because many organisms rely on their signaling systems to manage both the production and the activity of their antimicrobials [30, 31].
Another important secondary metabolite to consider is ralsolamycin, a lipopeptide highlighted in studies on Ralstonia solanacearum, which has a significant influence on chlamydospore formation in fungi belonging to the Mucoromycota, Ascomycota, and Basidiomycota divisions. Specifically, Fusarium fujikuroi produces bikaverin, another antimicrobial agent, and recent research suggests that ralsolamycin modifies the metabolic profile of F. fujikuroi, stimulating the production of bikaverin and beauvericin. These compounds have been shown to be effective in counteracting R. solanacearum, indicating a possible defensive role for F. fujikuroi [32]. Furthermore, ralsolamycin inhibits the cluster of imqK genes in Aspergillus flavus, favoring the production of imizoquin (an alkaloid derived from tripeptides that protect against oxidative stress and is essential for regular germination), facilitates spore germination, and reduces the population of R. solanacearum [33]. Considering the numerous studies on this topic, further research is necessary to determine the precise role of ralsolamycin.
Pyrrolnitrin (microbial pyrrole halometabolite), produced by bacteria such as Pseudomonas and Burkholderia, is another relevant biomolecule with antifungal properties and significant biotechnological value for agricultural, pharmaceutical, and industrial applications. This compound is known to be effective against plant pathogens, e.g., Phytophthora capsici and Rhizoctonia solani, at the cellular level, where it affects glycerol kinase, leading to glycerol accumulation and, ultimately, cell lysis [34, 35]. Similarly, Trichoderma spp. are particularly notable fungi with antipathogenic properties. In addition to hundreds of molecules with bioactive potential, this genus biosynthesizes signaling compounds, such as trichorzin (antimycoplasmic activity resulting from membrane permeability perturbations), peptaibols (amphipathic molecules that form voltage-dependent ion channels in cell membranes that cause perforation, leading to cell leakage and eventual death), and peptaivirins, which exhibit antimicrobial and antiviral properties against pathogens, such as cucumber mosaic virus (CMV; plant pathogenic virus of the Bromoviridae family) and tobacco mosaic virus (TMV; plant pathogenic virus of the Virgaviridae family) [36, 37].
Environmental factors, such as temperature and light, significantly influence the immune system of plants [38]. This suggests that they process information from both microbial and environmental sources to adapt to physiological responses. Recent research has focused on analyzing transcriptional alterations that occur in Arabidopsis under single and combined stresses [39, 40, 41]. These studies have revealed that the transcriptional response to multiple stressors is not simply additive. For example, the genetic response to the combination of turnip mosaic virus (TuMV; Potyvirus of the Potyviridae family that causes diseases in cruciferous plants) and abiotic stresses, such as heat and drought, deviates significantly from responses to these factors individually and in some cases, the combination even increases susceptibility to the virus [41]. Similarly, the response to the nematode Heterodera schachtii changes markedly when faced with drought stress, differing from individual responses to these conditions [39, 42].
Plant cells have been shown to reconfigure their physical structure in response to microbial stimuli, and the discovery that proteins alter their subcellular localization in response to biotic signals represents progress in understanding this field [42]. An experiment carried out by Lee et al. [43] involved the application of an elicitor known as peptide Pep13, derived from a pathogenic oomycete belonging to the genus Phytophthora, in which the protein was shown to trigger a potent immune response in cultured parsley cells. This signal activates three distinct mitogen-activated protein kinases (MAPKs) under the influence of a higher MAPK level. In particular, immunolabeling studies have shown that once activated, MAPKs tend to be located mainly in the cell nucleus. Therefore, migration of MAPKs to the nucleus enhances their ability to phosphorylate their substrates.
Similarly, the Arabidopsis resistance protein RRS1-R confers protection against R. solanacearum infection [44]. This protein, which may have arisen from gene fusion, contains a TIR-NBS-LRR domain (proteins that have an amino-terminal domain similar to that of Toll and interleukin-1 receptors), commonly present in plant resistance proteins, as well as a WRKY domain (60 amino acid region characterized by the conserved amino acid sequence WRKYGQK at its N-terminus end, accompanied by a unique zinc finger-like motif) with its carboxyl-terminal end, which is characteristic of plant transcriptional regulators. Furthermore, research using bimolecular yeast systems has revealed that RRS1-R is associated with PopP2, an avirulence protein secreted by different phytopathogenic bacteria, such as Erwinia tracheiphila and Curtobacterium flaccumfaciens pv. flaccumfaciens [45]. In this regard, transient co-expression analysis revealed that both proteins were located in the nuclei of Arabidopsis protoplasts. However, after the co-expression of RRS1-R with a PopP2 variant lacking a nuclear localization signal, both were exclusively observed in the cytoplasm. This suggested a possible preliminary interaction between the two proteins in the cytoplasm prior to their translocation to the nucleus.
The process of integrating signals from plant gene regulation remains an area of ongoing research with numerous unexplored aspects. For instance, the primary mechanism through which this integration occurs is a versatile transcription factor (TF), which is influenced by various signaling pathways, either at the gene promoter, where different TFs interact with multiple cis-regulatory elements (CREs), or at the CREs themselves, which are associated with stress-responsive genes. Although several CREs have been identified, accurately predicting gene expression in response to stress remains challenging [46]. However, research using computational tools that examine co-expression patterns in biological models, such as Arabidopsis under different stress conditions, has detected putative CREs that behave as true cis-type elements [47].
The precision in determining gene regulation under various stress factors, including salinity, ultraviolet radiation, and the presence of an elicitor of plant immune responses called flg22, has increased by analyzing a combination of CREs. This response is shaped by an intricate network of signaling that processes multiple metabolic pathways [47]. In a different context, the plant immune defense must provide resistance against pathogenic attacks and adaptability to a diverse spectrum of microorganisms, from biotrophs to necrotrophs. Therefore, maintaining ecological balance is crucial because, in contrast, exaggerated immune responses affect plant health [48, 49, 50].
Although PTI signaling is robust, its adaptive capacity appears to be lower than that of the ETI pathway [51]. To better understand this phenomenon, a formal analysis model was developed to understand the properties of PTI, such as its resilience and versatility, by integrating signal streams from various components, including jasmonic acid (JA), peptidyl arginine deiminase 4 (PAD4), and salicylic acid (SA), which are supported by extensive quantitative data and the establishment of characteristic indicators for each component in two separate instances [52]. Furthermore, the impact of exposure to microbe-associated molecular patterns (MAMPs), consisting of flg22, a peptide derived from the N-terminus of the elf18 elongation factor (thermo unstable; EF-Tu), and chitosan, was examined on the growth of two strains of Pseudomonas syringae [53]. These results represent illustrative examples of the importance of analyzing molecular effectors closely related to the integration of environmental and microbial signals in plant defense, enlightening our understanding of this process.
Regarding the crucial role of filamentous proteins in the regulation of multiple cellular processes, including the control of morphology and motility, it can be concluded that plant pathogens have devised strategies to establish symbiosis with their hosts by modulating actin cytoskeleton-related processes, as mentioned in [54, 55, 56]. In humans, intracellular pathogenic bacteria release effector proteins that modify the cytoskeleton, primarily through a type III secretion system. Although pathogenic bacteria in plants act mainly externally, they also employ this system to introduce effector molecules. For example, the AvrPto effector is essential for inhibiting callus formation in the apoplastic areas of Arabidopsis leaves in the presence of P. syringae [57]. Previous studies [58] suggested that callus accumulation at potential pathogen access points is related to actin function. Thus, AvrPto can directly or indirectly influence the host’s actin cytoskeleton.
Microbes often produce multiple MAMPs, allowing plants to modulate the activity of the PTI signaling network by recognizing different combinations of these molecules [59]. This enables the induction of a context-specific immune response. Furthermore, it is widely believed that PAD4 amplifies SA signaling. However, recent evidence suggests that SA may primarily activate PAD4 rather than directly enhance the defense against biotrophic and hemibiotrophic pathogens. This is in contrast to the notion that PAD4 plays a more significant role in the defense against bacteria than SA. To clarify these dynamics, it is crucial to investigate both PAD4 and SA further, individually and in conjunction, to understand their interactions in plant defense.
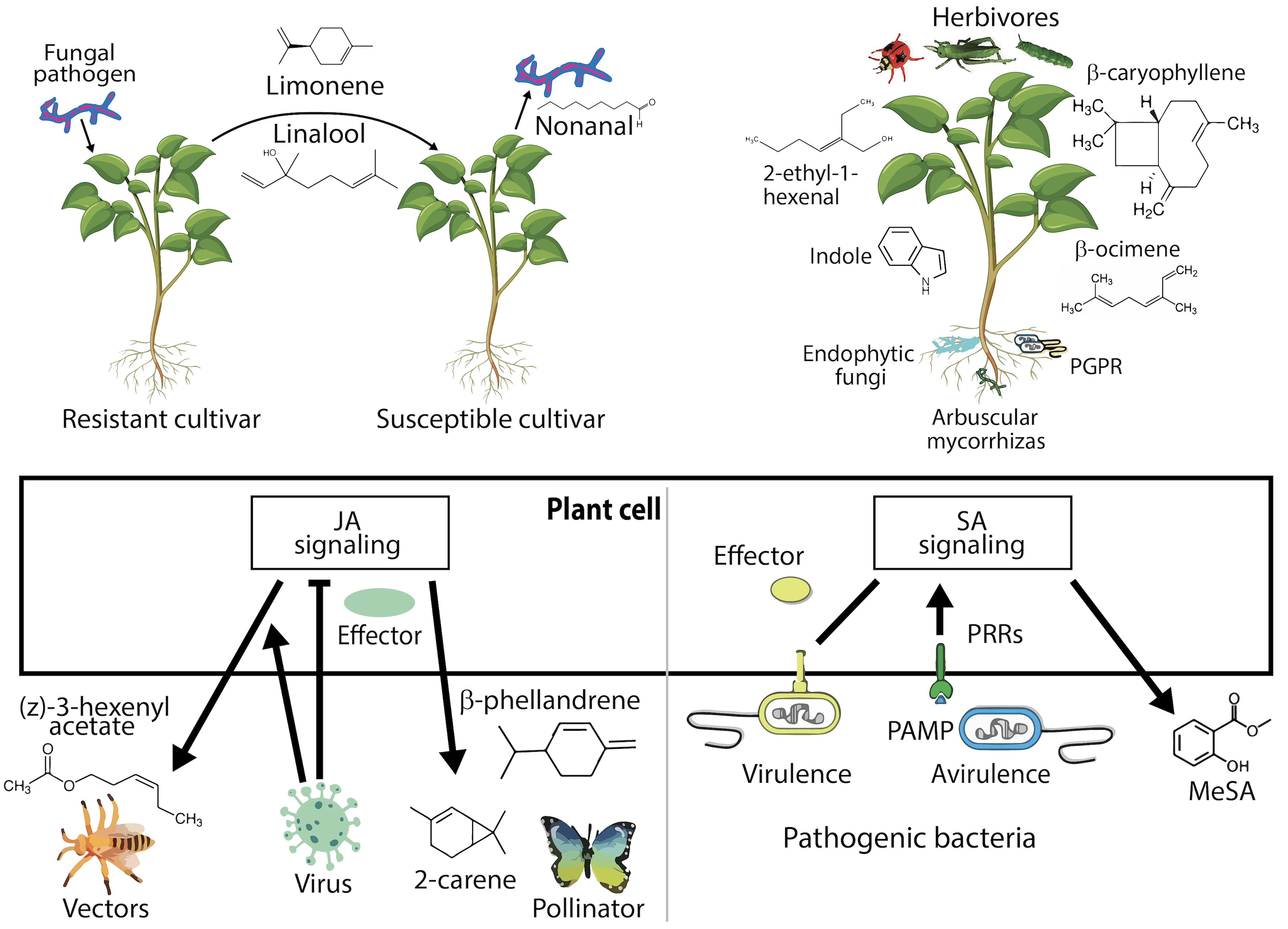
Sharifi et al. [60] suggested that host plants synthesize specific volatile compounds when attacked by antagonistic microbes. These secreted metabolites vary depending on the resistance of the plant to the invading microorganisms. Resistant plant varieties typically release many other chemical compounds, including limonene and linalool. On the contrary, a molecule called nonanal serves as a molecular marker. Similarly, exposure to exudates from resistant plants strengthens the defense mechanisms of vulnerable varieties and reduces fungal diseases. In the same way, beneficial microbes trigger the release of volatiles to prevent insects from causing damage. For instance, plants treated with plant growth-promoting (PGP) rhizobacteria metabolize biomolecules such as indole and beta-caryophyllene, which increase resistance to the antagonist. Similarly, symbiotic fungi (endophytes and mycorrhizal associations) potentiate the production of volatiles from their host plants to protect against insect threats. This process is illustrated in Fig. 1 (Ref. [60]).
 Fig. 1.
Fig. 1.Microbial pathogens affect host plants, stimulating them to release volatile compounds. This volatile production is related to the susceptibility of plants to fungal threats. Among resistant strains, emissions are rich in limonene and linalool, whereas susceptible strains are characterized by nonanal prevalence. When sensitive strains encounter volatiles from their resistant counterparts, their defenses improve, thereby reducing fungal infections. Beneficial microbes play a similar role in causing volatile release, providing protection against insect herbivory. The application of rhizobacteria, which are known to improve growth, leads plants to produce defensive volatiles, such as indole and beta-caryophyllene. Additionally, symbiotic relationships with fungi, including those living within the plant or forming mycorrhizal connections, influence these volatile patterns, offering a defense mechanism against insect invaders. JA, jasmonic acid; SA, salicylic acid; PAMP, M/pathogen-associated molecular pattern; MeSA, methyl salicylate. Adapted from [60].
Although the mechanics of bacterial chemotaxis have been fully elucidated, their underlying mechanisms remain unclear. Conventionally, chemotaxis has been defined as a bacterial strategy to search for food and optimize nutrient uptake. However, certain compounds with low nutritional values exert chemoattractant properties, whereas highly nutritious compounds do not. Chemotaxis has been shown to serve broader ecological purposes, such as directing colony expansion, identifying hosts or symbiotic partners, and promoting microbial diversity through spatial segregation in communities. Despite the diverse contexts in which chemotaxis has been studied, a substantial portion of our knowledge is derived from laboratory experiments using model organisms. This led us to consider the more refined functions of chemotaxis and how further exploration in this area could advance our understanding of microbial behavior and ecology [61].
While many microbial symbionts associated with eukaryotic hosts have been identified, recent evidence suggests the existence of symbiotic relationships between prokaryotes in nature [62]. The metabolic coupling between microbial communities offers several benefits, including enhanced motility and chemotaxis, to cope with the challenges posed by the small size of the participating organisms. Chemotaxis plays an important role in forming microbial communities in various habitats. For example, filamentous cyanobacteria that fix nitrogen, such as Anabaena spp., release signals at the edges of their heterocysts, which attract Pseudomonas spp., thus improving the fixation of this element [63]. Similarly, the sulfate-reducing bacteria Desulfonema spp. performs a sliding mobility effect to reach the mucus layer of Thioploca spp. This association promotes the reduction and reoxidation of sulfate between the two species [64].
Microorganism chemotaxis and motility represent fundamental symbiotic interactions. For example, the structure of a person’s dental plaque consists of at least 12 different microbial taxa organized in the consortia [65]. This precise organization is essential for the function and longevity of dental plaques in the oral environment. The microorganisms in these samples have been speculated to exhibit motility, possibly through flagella or type IV pili, and act primarily as episymbionts, establishing symbiosis on the surfaces of other microorganisms [66]. These examples underscore the importance of motility and chemotaxis as vital tools for microbes to facilitate the identification and formation of specific symbiotic relationships that directly affect their survival and success in the ecological realm.
It is widely acknowledged that the rhizosphere is a complex ecological environment, and the production of substances by plants in the surrounding soil, termed root exudates, significantly affects its structure [67, 68]. These exudates serve as attractants, drawing microorganisms to the roots and as a source of sustenance for those adapted to take advantage of them. The intense competitive and cooperative interactions that developed in this environment reflect a precise evolutionary balance. Plants adjust their root microbiome through selective exudate emission and changes in the rhizomicrobiome depending on the stage of development of the plant, its health status, genotype, and environmental factors [69]. Therefore, the ability of microorganisms to respond to chemical gradients is of the utmost importance. Depending on the compounds present in the exudates, microorganisms can strategically position themselves in the rhizosphere, attracting signals that indicate the availability of nutrients or moving away in response to an active plant defense [70].
The link between plants and microorganisms that live in their roots is a product of an evolutionary process. This process aims to provide mutual benefits and protect higher plant organisms primarily against diseases. Therefore, they adapt to and thrive in all types of environments. Plant exudates contain organic compounds that nourish soil microorganisms and act as signals to attract or drive different types of microbial communities [71]. Plants grow a community of these microbes in their rhizospheres by regulating and adjusting the concentrations of specific compounds, which helps promote growth and well-being [72].
Rudrappa et al. [71] suggest that complex biochemical interactions exist between Arabidopsis thaliana and its root microbiome. Their work group demonstrated how malic acid, an organic compound released by plant roots, functions as a selective signaling agent. This molecule can attract beneficial bacteria, such as Bacillus subtilis, and deter potential pathogens. This communication system is mutually beneficial for plants and bacteria. Plants protect themselves from any disease and simultaneously absorb nutrients efficiently. However, bacteria obtain a nutritional supply and a suitable environment for their growth. However, these methods are not limited to chemical signals. Previous studies have also suggested that plants use electrical gradients, called electrotaxis, to regulate microbial colonization [73]. Essentially, this is a directed movement that occurs in biological cells or organisms in response to an electric field. This adds another avenue to the complex relationship between plants and their root microbiome.
The phenomenon known as quorum sensing (QS) is a biological process in which bacteria use signaling molecules to communicate. They use this chemical exchange to monitor population density and modify gene expression [74]. An example of bacterial communication involves relationships with plants [75]. In this system, molecules known as N-acyl-L-homoserine lactones (AHLs) influence their behavior depending on their strain [76]. AHLs are common in pathogenic Gram-negative bacteria, e.g., Pseudomonas aeruginosa and Rhizobium radiobacter, but are also found in plant growth-promoting bacteria, as is the case for Burkholderia graminis [77, 78]. These molecules play key roles in microbes, including symbiosis, virulence, and antibiotic production. Additionally, plants can detect bacterial AHLs that affect gene expression in certain tissues. They also modulate development and trigger defensive responses [79].
Numerous studies have been conducted on plant–bacterial communication through AHLs and their analogs. For instance, Pérez-Montaño et al. [80] discovered that certain AHL patterns, such as N-octanoyl homoserine lactone and its 3-oxo or 3-hydroxy derivatives, play a critical role in legume interactions. Similarly, von Rad et al. [81] found that A. thaliana roots modified their transcription profile after contact with N-hexanoyl-homoserine lactone (C6-HSL). Furthermore, some plants, including soybeans and rice, release compounds analogous to AHL, including furanone signals, which modulate the behavior of QS-mediated bacterial communities [82]. These AHL analogs are structurally similar to bacterial AHLs and are capable of binding to and altering the function of the LuxR receptor (250 amino acid polypeptide) [83], thus interfering with bacterial signaling, typically orchestrated by AHLs.
Plants have developed advanced strategies for synthesizing AHL mimics that enhance their defense mechanisms against pathogenic microorganisms. Roots also biosynthesize elicitors, including flavonoids and genistein, which play a critical role in QS dynamics. These biomolecules play a fundamental role in the attraction of rhizobia to the roots of legumes and in promoting their adhesion. Furthermore, they regulate the expression of genes essential for nodulation, including Nod and Rhi, in plant tissues [84]. In view of these findings, it is important to recognize the existence of volatile organic compounds (VOCs) produced by bacteria that interfere with QS and significantly affect AHL synthesis in different types of microorganisms [85, 86]. This background is crucial for developing innovative approaches to improve the performance of plant biotechnology by optimizing interactions within the rhizosphere community [87].
Transcription factors (TFs) play critical regulatory roles in essential biological processes in plants. They interact with cis-type elements located on the promoters of target genes or through protein–protein interactions. In this regard, maintaining the optimal expression levels of crucial genes involved in biosynthetic pathways and developmental processes is imperative for the proper growth and survival of plants. TFs are central mediators of gene transcription and are not only responsible for modulating the expression of various genomic sequences but also play an important task regulating plant physiology, as their influence is notable in response to biotic and abiotic stress situations and are essential for the regulation of secondary metabolism [88].
Detecting a pathogen by the host triggers the activation and amplification of certain enzymes, hormones, and metabolites essential for its response. Gene regulation is a fundamental aspect of host–pathogen interactions in the immune cascade. Numerous studies have identified potential genes that encode key proteins involved in plant–pathogen interactions. Many of these genes have been confirmed to play direct or indirect roles in the ability of the host to identify pathogens. To date, most studies have focused on genomic and transcriptomic findings, with a particular emphasis on TFs and elements that modulate genetic material as they significantly influence the remodeling of the host cell’s transcriptomic profile. This reconfiguration improves the production of defense molecules. Interestingly, the initial stages of these defense responses often involve the expression of several stress-response genes (WRKY TFs) and reactive oxygen species (ROS)-scavenging enzymes. These TFs contain a WRKYGQK sequence domain and exhibit a characteristic zinc-finger motif [89].
Peng et al. [90] discovered the role of the OsWRKYY30 gene in
the defensive responses of rice (Oryza sativa) under various conditions.
These included exposure to JA, its derivative methyl jasmonate, SA, and two known
rice pathogenic fungi: Rhizoctonia solani and Magnaporthe
grisea. After exposure to JA and SA, a significant accumulation of
OsWRKY30 transcripts was observed. Furthermore, the expression of this
gene was positively correlated with the activation of several defense-related
genes, particularly those associated with JA synthesis and its signaling
pathways, such as lipoxygenase (LOX), allene oxide synthase 2 (AOS2), PR-3, and
PR-10. Furthermore, enhanced expression of OsWRKYY30 led to an increase
in JA levels in response to infection by the aforementioned pathogenic fungi. It
is important to mention that in addition to the expression of proteins with WRKY
domains, other related peptides have been identified, such as MYB, Sigma, and
NF-
Fungal pathogens, including Botrytis, Fusarium, and Verticillium species, are the main causes of plant diseases. To combat these threats, plants have evolved efficient immune systems via evolutionary lineages. As a result, several RNAs play critical roles in plant–fungal interactions. Recent studies have used high-throughput genomic sequencing techniques and advanced bioinformatic tools to elucidate the functions of different RNAs in plant defense or vulnerability to fungal attack. Yin et al. [91] identified microRNAs (miRNAs) and other RNAs in two varieties of cotton (Hai-7124 and Yi-11). These biomolecules showed different responses in the presence of Verticillium: Hai-7124 was resistant to this pathogen, whereas Yi-11 was susceptible to it. This study revealed the presence of more than 65 miRNAs with altered expression levels after invasion by plant pathogens. At least three miRNAs (Ptc-miR482, Ptc-miR1444, and Ptc-miR1448) modulate plant responses to biotic and abiotic stresses. Furthermore, they were found to affect genes that transcribe polyphenol oxidases as well as pathogen-resistance proteins [92].
RNA also plays a crucial role in the interactions between oomycetes and their host plants. For example, Phytophthora employs the effector PSR1 to target specific elements of the RNA pathway in plants. This process alters immunity, thereby facilitating infection [93, 94, 95]. It should be mentioned that the WY domain of PSR1 is essential to initiate infection and counteract RNA silencing in many plants [96]. A recent study explored how miRNAs are related to the target genes that confer resistance to Verticillium wilt in cotton. During this investigation, small RNA (sRNA) libraries were constructed, and genomic sequencing led to the identification of 383 miRNAs, among which GhmiR165 and GhmiR395 emerged as crucial elements in response to Verticillium dahliae. These miRNAs play a role in vascular development and secondary cell wall generation through the GhmiR165-REV pathway and sulfur incorporation through the GhmiR395–APS1/3 axis [97]. Furthermore, it was found that the interaction between GhmiR477 and CBP60A molecules is a determinant of the cotton response during the advanced stages of infection. GhmiR477 has been shown to fragment GhCBP60A mRNA at the post-transcriptional level, and inhibiting GhmiR477 reduces resistance, whereas silencing GhCBP60A improves it [98].
In addition to the effectors mentioned above, certain molecules establish direct interactions with SNARE proteins (SNAP receptors; mediators of vesicle fusion with the target membrane), thereby contributing to pathogenesis. An example of such an effector is PsAvh181 of Phytophthora sojae, which is located on the plasma membrane (PM) of plants and targets GmSNAP-1 in soybean plants. Disrupting the interaction between GmSNAP-1 and GmNSF alters immune exocytosis and reduces the availability of GmSNAP-1 molecules [99, 100]. It is also crucial to highlight the effects of the PexRD12/31 effectors of Phytophthora infestans, which directly affect vesicle trafficking and establish a significant interaction with the NbVAMP72x plant effector [101]. Upon the expression of PexRD31 in seedlings, an increase was detected in FYVE-targeted compartments (a protein motif that is preserved in different species and is defined by its high affinity and specific binding to phosphatidylinositol 3-phosphate (PI3P), a type of phosphoinositide abundantly found in early endosomes), replicating the conditions observed in plants invaded by P. infestans. Given that FYVE specifically binds to PI3P, which is abundant in early endosomes and multivesicular bodies (MVB), and considering the dimensions of these compartments of FYVE, it is plausible to assume that they are actually MVBs [101].
Furthermore, it is worth mentioning the nature of specialized feeding structures called haustoria by filamentous pathogens, such as fungi and oomycetes. These structures are derived from the invagination and expansion of the PM of the host plant [102, 103, 104]. In contrast, a constant recycling process involving the flagellin receptors FLS2-GFP and GFP-SYP121 was observed between the PM and endosomes in the extrahaustorial matrix. This space is located between the fungal and plant membranes, and this observation leads to the hypothesis that filamentous pathogens employ MVBs to extend the plant PM. Table 1 (Ref. [105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123]) shows representative miRNAs involved in plant–microbe interactions.
| miRNA | Pathogen type | Defensive role | Pathogen species | Target | Reference |
| miR156 | Bacterium | Brassica oleracea | Xanthomonas campestris pv. campestris | ARF | [105] |
| miR160 | Bacterium | Arabidopsis thaliana | Pseudomonas syringae | ARF10 | [106] |
| miR167 | Bacterium | Arabidopsis thaliana | Pseudomonas syringae | ARF8 | [107] |
| miR390 | Bacterium | Brassica oleracea | Xanthomonas campestris pv. campestris | ARF | [108] |
| miR393 | Bacterium | Arabidopsis thaliana | Pseudomonas syringae | TIR1, AFB2 | [109] |
| miR472 | Bacterium | Arabidopsis thaliana | Pseudomonas syringae pv. tomato | CC-NBS LR | [110] |
| miR1448 | Fungus | Populus sp. | Botryosphaeria dothidea | ABC Trs | [111] |
| miR1450 | Fungus | Populus trichocarpa | Botryosphaeria dothidea | NBS-LRR | [112] |
| miR156 | Fungus | Triticum aestivum | Erysiphe graminis | SPL | [113] |
| miR164 | Fungus | Gossypium hirsutum | Verticillium dahliae | NAC100 | [114] |
| miR164a | Fungus | Oryza sativa | Magnaporthe oryzae | NAC60 | [115] |
| miR396a | Oomycete | Nicotiana tabacum | Phytophthora nicotianae | GRF | [116] |
| miR482 | Fungus | Gossypium hirsutum | Verticillium dahliae | DS protein | [117] |
| miR9664 | Fungus | Triticum aestivum | Puccinia striiformis f. sp. tritici | CLP1 | [118] |
| miR156 | Mutualistic microbe | Solanum lycospersicum | Pochonia chlamydosporia | SPL | [119] |
| miR10186 | Nematode | Glycine max | Heterodera glycines | Rhg4 | [120] |
| miR159a | Virus | Nicotiana tabacum | Potato virus Y | HC-Pro | [121] |
| 17 20–21 nt sRNA | Pathogenic | Triticum aestivum | Puccinia striiformis | NBS-LRR | [122] |
| 21 nt sRNA | Pathogenic | Solanum lycopersicum | Botrytis cinerea | ATG2 | [123] |
Note: miRNA target abbreviations: ARF, auxin response factors; TIR1, f-box auxin receptor; AFB2, f-box auxin receptor; CC-NBS-LR, disease-resistance proteins domains; ABC Trs, ATP-binding cassette; NBS-LRR, nucleotide binding site–leucine rich repeat; SPL, SQUAMOSA promoter-binding protein-like; NAC 100/60, miRNA regulatory system; GRF, growth-regulating factor; DS, drought stress; CLP1, multifunctional kinase encoded protein; Rhg4, resistance to Heterodera glycines 4; HC-Pro, viral suppressor of RNA silencing; ATG2, key regulator of autophagy induction.
Transcriptional flexibility provides an additional layer of complexity in the evolutionary process and is not limited to genetic changes inherited from previous generations. The genotype of a particular organism leads to various transcriptional phenotypes depending on the environmental conditions under which it develops. This finding suggests that genes subjected to intense environmental stress often exhibit differences in expression levels between different populations, species, or even between isolated individuals. The speed and level of gene expression play crucial roles in virulence, indicating that plant pathogens take advantage of this transcriptional flexibility to improve their infection strategies in host organisms [124].
Adaptation of different isolates enables plant pathogens to show various transcriptional profiles, achieving varying degrees of efficacy when infecting the same host. For example, various fungal strains of Blumeria graminis exhibit significant differences in effector gene expression when invading the host [125]. In the case of Zymoseptoria tritici, up to 30% of genes undergo alterations in their regulation during infection by a single host, which could explain the observed virulence discrepancies [126]. Recent studies on experimental evolution in yeast have indicated that variations in gene expression related to a selected trait confer remarkable benefits in terms of survival and adaptation. Although epigenetic alterations are fundamental in this process, discerning the molecular mechanisms underlying transcriptional flexibility in plant pathogens remains challenging.
In plant pathogens, as in plants themselves, epigenetic mechanisms such as DNA methylation and histone alterations are essential to establish routes of infection and physiological development [127]. There is growing evidence that pathogens utilize epigenetic tools to enhance virulence [128, 129]. In relation to plant pathogens, epigenetic adaptations that affect avirulence effector genes have been suggested to act as regulatory systems, allowing pathogens to evade host alerts [130]. However, the role of these epigenetic factors in host–pathogen interactions has yet to be explored.
Phytophthora ramorum has a wide range of host plants, with more than 100 recorded species [131]. Although it exhibits limited genetic diversity in North American forests, it shows remarkable phenotypic variation among isolates. These variations include changes in the colony morphology, senescence, and virulence. Correlations were identified between these characteristics and the species of origin of isolates. For example, most California coastal oak strains (Quercus agrifolia) show increased activity by transposable elements (TEs) and decreased expression of the crinkler effector gene (CRN), which are essential for pathogenicity. This led to a decrease in the viability of the isolates. In contrast, isolates of the California laurel (Umbellularia californica) exhibited a marked increase in CRN expression. This suggests that interactions with oak trees induce stress in the pathogen by altering TE silencing, thereby activating them. Given that TEs are often regulated by epigenetic silencing mechanisms, it is plausible to consider that these epigenetic processes may be responsible for phenotypic variations in different isolates of P. ramorum, depending on the host plant of origin [128, 129, 130, 131].
Plant pathogens employ specialized effector molecules to infect their hosts successfully. These molecules facilitate host invasion and interfere with the host immune system [132, 133]. Proper expression of these effectors requires precise transcriptional regulation to balance the interference with host immunity and the potential risks of triggering defensive responses or adverse transcriptional effects [134, 135]. In eukaryotic transcription, TFs bind to promoter regions to activate DNA-dependent RNA polymerases, culminating in the production of mRNA [136]. Several transcriptional regulators have been identified as essential for effector gene expression in pathogenic plant fungi [137]. The Sge1 regulator in Fusarium oxysporum f. sp. lycopersici is essential in regulating the expression of specific transcripts during tomato infection; meanwhile, the absence of Sge1 compromises the virulence of this fruit [138]. Sge1 homologs have also been documented to have similar roles in regulating pathogenicity-related genes in several plant antagonists [139].
For example, the TF Pf2 regulates genes linked to pathogenicity in different necrotrophic pathogenic fungi [140]. On the contrary, in Magnaporthe oryzae, the G-protein signaling regulator RGS1 acts as a transcriptional regulator, suppressing the expression of several effector genes before colonizing the plant [141]. Although these regulators generally coordinate the expression of effector genes during infection, it is crucial to recognize that they are activated at specific stages of infection. The analysis of fungal chromatin from an epigenetic perspective in fungal infections has also been extensively studied, although challenges remain due to the disproportionate biomass between the pathogen and the host plant. Consequently, plant-transcribed genomic sequences are often predominant [142], highlighting the need for more sophisticated regulatory mechanisms to ensure the timely expression of effector genes during infection. Therefore, using a multidisciplinary approach, specifically in the context of pathogenic plant fungi, is imperative for an in-depth understanding.
Some patterns associated with microbial invasion, in addition to MAMPs, include structural components known as damage-associated molecular patterns (DAMPs) generated by the host plant in response to pathogen-induced damage. Furthermore, pathogens secrete proteins or metabolites that function as pattern recognition or effector molecules during invasion. Microorganisms also adapt to staying out of sight by secreting proteins that interfere with recognition or altering molecules that are detectable by the host. These mechanisms allow pathogens to avoid detection and establish successful infections [133, 143]. In return, plants have developed receptors to counteract the expansion of pathogens. This mutual adaptation establishes a continuous evolutionary race between plants and pathogens. As a result, variation in a single gene significantly impacts host–pathogen interactions, potentially transforming them from compatible to incompatible and vice versa [144, 145, 146].
Although most of these metabolic pathways have been almost completely
elucidated, the complexity of the induced immune responses in plants remains to
be determined. Moreover, the basic pathway of the defense cascade mechanism has
yet to be discovered. This process begins when a plant detects an antagonistic
organism. The first cellular reaction involves altered plasma membrane
permeability, Ca
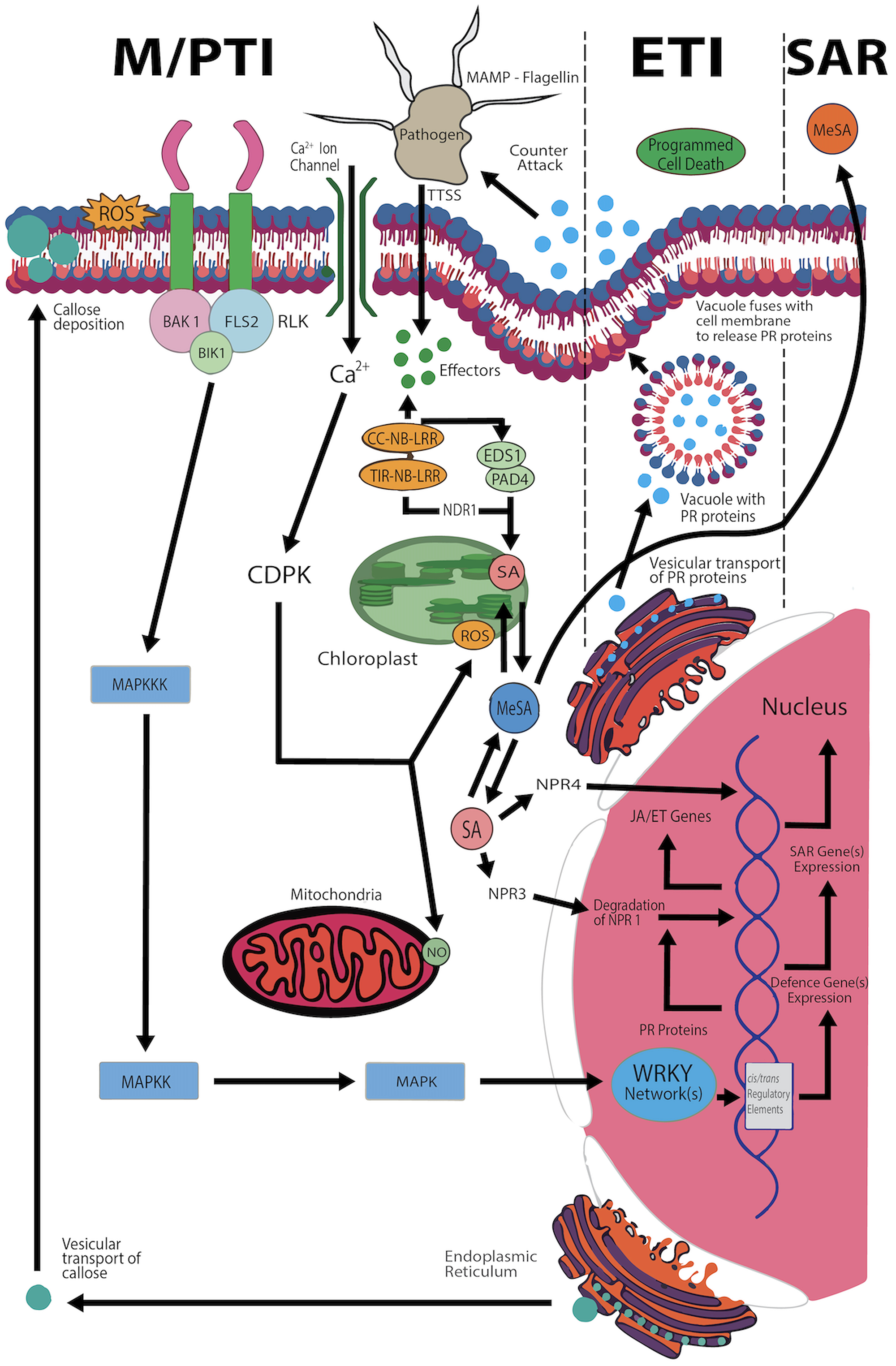
 Fig. 2.
Fig. 2.Diagram of plant immune response dynamics at the cellular level. The relationship between innate mechanisms and various immunity and defense strategies when encountering microbes is described. The sequence of M/pathogen-associated molecular pattern (PAMP) recognition triggers M/pattern-triggered immunity (PTI) to initiate effector-triggered immunity (ETI), resulting in systemic-acquired resistance (SAR). The image includes key components, including BAK1, BIK1, and CC–NB–LRR complexes. This illustration is based on the work of Muthamilarasan and Prasad [147] and was adapted from [148]. ROS, reactive oxygen species; FLS2, Arabidopsis receptor kinase; RLK, receptor-like kinase; CDPK, calcium-dependent protein kinases; MAPKKK/MAPKK, mitogen-activated protein kinase kinase/kinase; MAMP, microbe-associated molecular pattern; TIR-NB-LRR, toll interleukin 1 receptor domain found in nucleotide-binding (NB) leucine-rich repeat resistance proteins and their truncated homologs; NDR1, non-race specific disease resistance-1; EDS1, enhanced disease susceptibility-1; PAD4, lipase-like expression profile; SA, salicylic acid; NPR, nonexpressor of pathogenesis-related; JA/ET, jasmonic acid and ethylene expression profiles; WRKY, plant-specific transcription factor group in plants; PR, cytosolic pH homeostasis.
Although the intricacies of secondary metabolite secretion in plant–microbe interactions are not yet fully understood, they are unequivocally associated with cell polarity. Consequently, the insights obtained from studies on other polar systems, such as cilia, epithelial barriers, and cell division in yeast, may be of significant utility in future research [147, 148, 149, 150, 151]. Establishing and maintaining polarity requires a bifurcated approach: Initially identifying target membrane regions and then unevenly distributing secretory vesicles to those areas. The initial signals for the recruitment of membrane markers such as PEN1 arise from detecting microbes. These vesicles are discernible by their electrically charged molecules and probable interactions with enveloping proteins [152].
There is increasing evidence that plant–microorganism interactions exhibit similar structural features. For example, fungal pathogenic interactions characterized by an appressorium share similarities with symbiotic interactions hosted by an arbuscule. Structures designed to host symbionts such as bacteria or fungi have also revealed comparable patterns. For instance, a developing arbuscule triggers an infection thread that hosts rhizobia, and an arbuscule bud shares similarities with the symbiosome [153]. Beyond these visible similarities, it is possible that common mechanisms of cell polarity exist and that discoveries in one area may complement those in another [154].
The host cell faces the challenge of coordinating the secretion of proteins necessary for both defense and symbiosis. As a case in point, when a nitrogenous bacterium is present, the cell has two potential membranes at its disposal: the already existing plasma membrane and the membrane of the developing symbiosome [155]. This raises the question of how cells ensure that symbiotic materials, such as nodule cysteine-rich (NCR) peptides, are directed only to the symbiosome, thereby preventing their accidental release into the extracellular space. A similar challenge arises in the interactions with both symbiotic and pathogenic fungi. Components, such as membrane proteins, cell wall elements, and possibly secreted proteins, are located at the point of interaction. Given this selectivity, these mechanisms allow the host cell to create and maintain a unique interface that fuses easily with the plasma membrane [156].
Examination of the nature of host cells involved in interspecific interactions is often directed toward a phenomenon known as endophytism. Endophytes are microbial communities that inhabit the internal tissues of plants and are often referred to as microbiomes [156, 157]. However, the concept of pathobiome [158, 159] can be extended to include these communities, eventually leading to the term holobiont [160]. Pathobiomes are particularly important in recognizing that diseases could arise from symbiotic combinations of eukaryotic, microbial, and viral entities present in the plant, as well as from surrounding biotic factors that affect host health [161]. Thus, microorganisms coexist with the plant throughout its life or only at specific stages, and their interactions range from mutualism and commensalism to amensalism and parasitism [162].
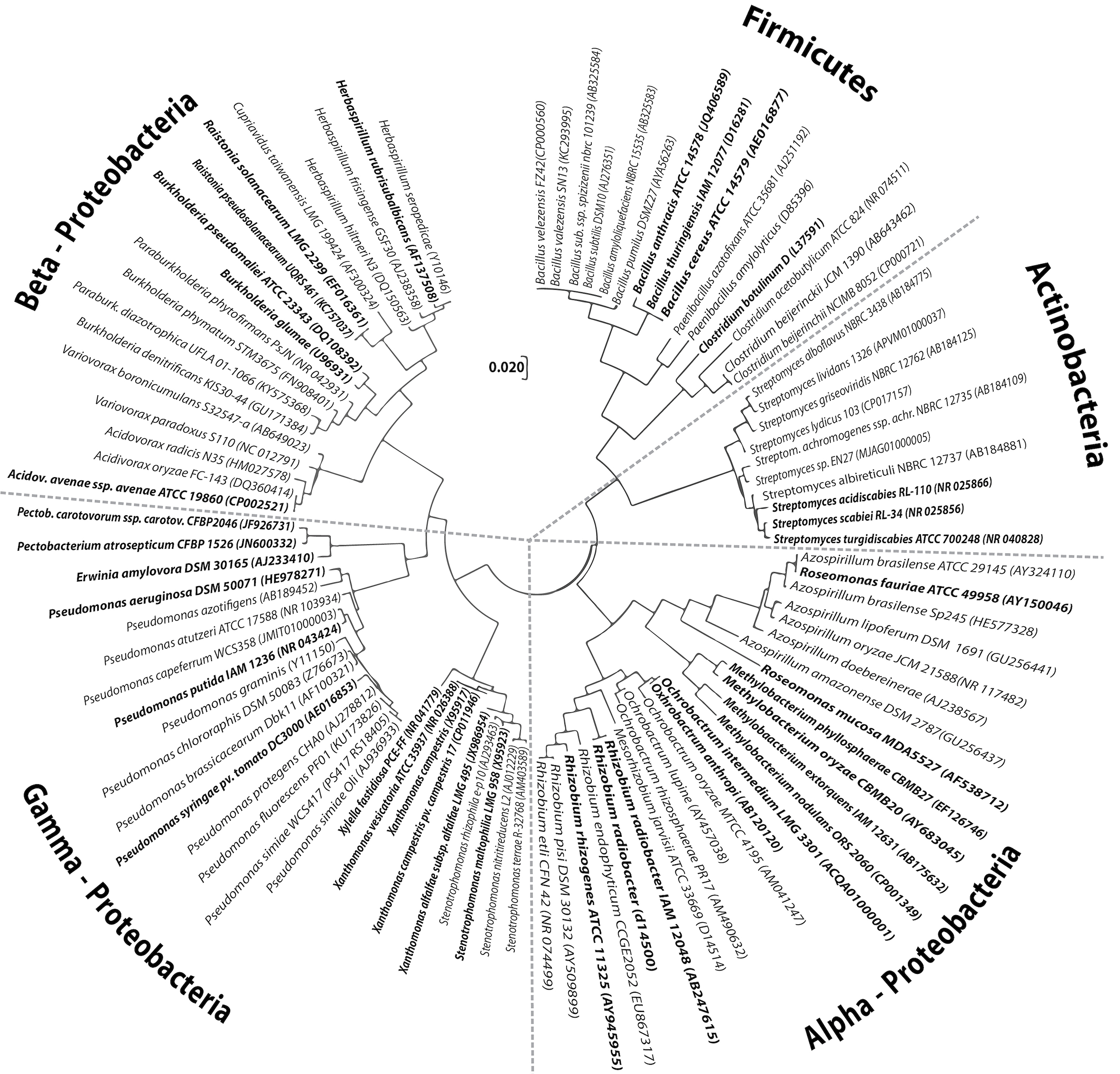
As previously mentioned, multiple interactions between plants and microbes are regulated by various molecular mechanisms, and only a portion of these interactions has been revealed. For example, endophytes have developed strategies that allow them to bypass the immune defenses of plants and prevent colonization [163]. This is not a unique phenomenon, as pathogens employ similar tactics in their successful colonization efforts. As Queiroz and Santana [164] and Rodriguez et al. [7] mentioned, endophytic bacteria and their mutualistic counterparts infiltrate plant tissues as pathogens do, yet do not cause damage and often provide advantages. These common needs lead to host-associated microbiomes in diverse phyla, where species are closely related but divergent in their impact (Fig. 3, Ref. [7, 164]).
 Fig. 3.
Fig. 3.Selected plant growth-promoting bacteria exhibit evolutionary ancestry with certain pathogenic bacteria. The diagram shows the evolutionary relationships [164] between taxa descended from common ancestors and the distances between them. At least five main clades were observed, which were subdivided into subclades that originated from a common group. Evolutionary reconstruction was conducted using the maximum likelihood method based on the Tamura–Nei model. Adapted from [7].
It is of utmost importance to note that many organisms that are labeled pathogens exhibit phases in their life cycle in which they cohabit within their hosts and behave as endophytes. Several investigations have suggested the existence of pathogenic microorganisms that live in a dormant state, either within the endosphere of healthy plants, such as endophytes, or opportunists [165, 166, 167, 168]. Many fungi have been shown to coexist as potential pathogens without causing considerable damage to their hosts [169, 170]. Manzotti et al. [165] identified six fungal species in tomato roots, each species triggering pathological symptoms after inoculation. These findings highlight the importance of maintaining a balanced microbiome in plants, where potential pathogens are regulated by the synergistic action of other microbes [170].
Ramularia collo-cygni is a notable example of a passive pathogen that infects barley crops without causing any immediate symptoms. This fungus can colonize barley seeds and spread throughout the vascular system of the plant, manifesting disease symptoms only when it reaches the leaves [171, 172]. A study by Stam et al. [173] suggested that this organism may have recently adopted a pathogenic role, as it lacks genetic markers commonly found in other pathogens. Species in the Mycosphaerellaceae family, including Zymoseptoria tritici in wheat [174, 175], Pseudocercospora fijiensis in banana [176], Cercospora beticola in sugar beet [177], and Fulvia fulva together with F. oxysporum in tomatoes [178, 179], exhibit a subtle infection pattern. They initially establish themselves within the host organism asymptomatically and later demonstrate their virulence through the production of spores. It should be noted that this behavior is not unique to ascomycetes, as some basidiomycetes, for example, those of the genus Ustilago, also exhibit similar patterns [180].
Certain microorganisms have been proposed to have evolved from an endophytic lifestyle to a pathogenic profile while maintaining beneficial characteristics. However, some species, such as V. dahliae, have been suggested to have the potential to transition into endophytes [181]. Recent findings indicate that infection with a specific mycovirus leads to the transformation of the Sclerotinia sclerotiorum pathogen into a beneficial endophyte [182]. Furthermore, regulating hormone signaling in the plant defense is of utmost importance in determining the outcome of plant–microbe interactions. Several plant pathogens manipulate phytohormone levels to facilitate infection [183, 184, 185]. Moreover, endophytes significantly affect their host plants, particularly under abiotic stress conditions [186, 187]. Certain molecules have been discovered to mediate interkingdom communication, primarily to fortify pathogenic or commensal relationships. Some necrotrophic pathogens produce sRNA signaling molecules that suppress host defense, a process that depends on interfering RNA (iRNA) [188]. These molecules also appear to be involved in regulating endophytic signaling [189].
Multiple mechanisms through which endophytic organisms overcome plant defenses have been demonstrated by research on evolutionary pathways for pathogenic virulence, and valuable insights have been obtained from these studies. Sacristán et al. [190] investigated the determinants of evolutionary virulence and identified key mechanisms, including mutations, mobile genetic elements, gene duplication, neofunctionalization, and horizontal gene transfer. Transposable elements have attracted significant attention within the scientific community, particularly in relation to comparative analyses of endophytic and pathogenic organisms. In this regard, Fusarium oxysporum is noted for possessing additional chromosomes, referred to as supernumerary chromosomes [191].
Although the evolutionary mechanisms underlying endophytism and its genetic context remain debatable, several theories have been proposed to explain it. For instance, endophytic lineages were proposed to have arisen multiple times from phytopathogenic ancestors. However, alternative hypotheses suggest that phytopathogens may have evolved from endophytic ancestors [190]. Although the transitions between parasitic and endophytic modes of life are likely to vary between lineages, the specific genetic structure of a fungal lineage determines its adaptive potential [192]. One of the difficulties in determining the ecological functions of horizontally transmitted endophytes (HTEs) is their inherent variability. Members of the ascomycetes group possess endophytic representatives, suggesting a range of potential host or substrate interactions that are specific to each endophyte. Interestingly, this refers to the “endophytic continuum theory”, which suggests that fungal endophytes vary in their physiological status, infection methods, colonization patterns, secondary metabolism, life-history strategies, developmental stages, and evolutionary phases. This diversity extends to various fungal and host taxa that participate in symbiotic relationships, including the nature of plant–fungus interactions, ranging from mutualism to parasitism, which is influenced by factors such as fungal species, host genetics, and environmental conditions [193].
The symbiotic relationship between legumes and rhizobia is characterized by sophisticated molecular interactions. Legumes release flavonoids into the surrounding soil [194, 195], which leads rhizobia to produce specialized lipo-chitooligosaccharides called Nod factors. These molecules initiate nodule formation in plant roots [196, 197] and are detected by plant receptors equipped with LysM domains [198]. This detection leads to a cascade of transduction events within the plant cell nucleus, resulting in the activation of TFs and the regulation of gene expression. This, in turn, oversees nodule formation [199, 200, 201]. Similarly, carbohydrate-based signals are responsible for directing bacterial penetration into roots by utilizing LysM domain-binding receptor kinases. These receptors recognize bacterial exopolysaccharides [199] and play a critical role in promoting bacterial infection through the infection thread [200]. Plant NIN TFs (which appear to have different functions in the root epidermis and root cortex) are known to perform a wide range of functions in the association between legumes and rhizobia, ranging from the initiation of infection to the nitrogen fixation process within nodules [200, 201, 202, 203, 204, 205, 206]. In this regard, genetic approaches have evolved to understand the key TFs necessary for nodulation [207, 208].
Interestingly, plant pathogens employ contrasting strategies when interacting with host plants compared to symbiotic signals. These pathogens transmit proteins to plant cells to favor colonization or suppress host defense responses. Gram-negative bacterial pathogens use a type III secretion system (T3SS) to transfer effector proteins to plant cells [209]. Some of these effectors, such as Xanthomonas and Ralstonia TALs, target the nucleus of the plant, bind to specific DNA sequences, and stimulate the transcription of genes that increase the susceptibility of plants to infection [210, 211, 212]. Oomycetes differ from other microorganisms because they introduce specific protein sequences, such as RxLR or LXLFLAK, which function as keys for accessing plant cells [213, 214, 215, 216, 217]. Although the RxLR segment is cleaved within the pathogen before secretion, the mechanism through which these effectors cross the membrane and gain access to the cells is not fully understood. Research on P. infestans has shown that approximately 25% of the analyzed RxLR proteins are located in the nucleus or nucleolus [218]. Consequently, pre-mRNA splicing is a vital process in eukaryotic gene expression since it eliminates non-coding sequences and splice-encoding regions to generate mature RNA [219]. The spliceosome, a complex consisting of RNA and proteins, directs this process [220].
Alternative splicing is a widely documented regulatory mechanism in plants that responds to various types of stresses [221, 222, 223]. Significant changes in the splicing of numerous genes have been observed in situations such as interactions with mycorrhizae, nodulation, or Ralstonia infections [224, 225, 226]. It should be noted that at least one type III effector of Ralstonia, RipS3, is localized to specific subnuclear domains [227]. This enriches our understanding of how splicing influences cell function and interactions with external agents such as pathogens or beneficial symbionts. Phytophthora species employ specialized structures known as haustoria to introduce effectors into the plant cells [228, 229]. An emblematic example is P. sojae, which causes root rot in soybeans. This species releases PsAvr3C effectors to enhance infection [213, 230]. It should be noted that PsAvr3 binds to soybean proteins rich in serine, lysine, and arginine (GmSKRP). These proteins, which are vital components of the plant spliceosome, undergo a normal splicing process that is altered by interactions with PsAvr3 [231].
Pathogens, such as P. infestans and P. sojae, have been found to manipulate the host’s iRNA process by releasing various effectors. For example, P. infestans biosynthesizes at least nine molecules that interfere with the tomato splicing system [232]. Similarly, the effector P. sojae PSR1, which is also involved in suppressing iRNA in the host plant, improves the virulence of this species in soybeans by inhibiting ribonucleic acid interference and promoting the formation of lesions [233]. An association between PSR1 and PINP1 plant RNA helicases has also been identified [233, 234]. Similarly, PINP1 has been found to be analogous to PRP16, a modulator of exon assembly in the spliceosome [235, 236, 237]. Overexpression of PSR1 and inhibition of PINP1 in Arabidopsis have been shown to trigger changes in alternative splicing of defense-associated genes [236]. Therefore, PSR1 could interfere with PINP1 activity. Through this process, the pathogen appears to modulate the host splicing mechanisms to neutralize its natural defense mechanisms.
Bioinoculants, a consortium of beneficial microorganisms, play a vital role in promoting plant health through molecular symbiotic exchanges, which are essential for plant growth. In this context, mycorrhizae improve nutrient absorption, and nitrogen-fixing bacteria increase plant nitrogen levels. These interactions enhance plant resistance to pathogens and significantly increase crop yields, making bioinoculants crucial in developing sustainable agricultural practices. Although introducing bioinoculants into the soil is technically feasible, a comprehensive understanding of the dynamics of microbial invasion remains elusive. Therefore, the physiology of beneficial terrestrial microbes and their interactions within the microbiome must be comprehensively studied [238]. Furthermore, their biosynthetic metabolism has been extensively explored. For example, Zaidi et al. [239] characterized a unique chemotactic phenomenon in B. subtilis, demonstrating that these colonies move through soil particles as functional clusters that target plant roots. These findings highlight the importance of conducting inquiries that improve our understanding of the nature of bioinoculants. In this respect, new functions for various secondary metabolites have been discovered daily [240, 241].
Bioremediation is a clear example of the interaction between plants and microbes within a biotechnological context. As we know, the increase in industrial activity and pesticide use (including inorganic compounds) in recent decades has profoundly impacted environmental conditions since human activities, xenobiotics, and natural processes affect plant life, soil quality, and human health through direct and indirect pathways. Although conventional remediation methods exist, their costs, toxicity, and scalability often limit their applications. Plant–microbe interactions are constant in nature and have evolved for applications in various domains. In this regard, growth-promoting rhizobacteria possess traits that influence flora and soil and thus play a role in purification processes both directly and indirectly. These bacteria drive the breakdown of numerous organic and inorganic substances, positioning them as viable agents for sustainable mitigation of pesticide pollution [238, 239, 240, 241].
The study of plant–microbe–metal exchanges has shed light on the crucial role of plant–microbe consortia in the biogeochemical renewal of metals and their utilization in phytoremediation. Selecting an appropriate combination of vegetation and microbial life is critical in remediation efforts to maximize the phytoremediation efficiency. This synergy aims to enhance the benefits of in situ microbial technologies. Interactions ranging from macroscopic (higher plants) to microscopic (microbes) and involving heavy metals are essential to understanding the mechanisms of plant-metal uptake during geobiological interactions [241]. Advances in understanding how plants and microbes coexist and address metal stress, as well as their collective contribution to the heavy metal biogeochemical cycle, including the dynamics of mobilization, immobilization, translocation, and metal conversion, have been crucial for refining metal bioremediation methodologies [242, 243, 244].
Considering the aforementioned points, the functions of plant growth-promoting microbes (PGPMs), including bioaccumulation, bioleaching, and bioexclusion, play a critical role in the evolution of microbial communities that are capable of adapting to, withstanding, or tolerating environments containing high levels of heavy metals. Acidification, chelation, and protonation typically result in the release of metals into the environment. Nevertheless, metal immobilization is often a consequence of precipitation, alkalization, and complexation processes. In particular, chemical transformations significantly influence the mobilization and immobilization of these elements [245, 246].
Kumar et al. [247] conducted a comprehensive examination of key plant and microbial characteristics and plant-microbe interaction processes in the context of agriculture and climate change. Their investigation included the following areas: (1) an in-depth analysis of biochemical and molecular frameworks that shed light on the plant–microbe dialogue, potentially influencing the evolutionary trajectory of microbial assemblages; (2) the presence of microbial candidates possessing stress mitigation traits, which imparted stress resilience to their plant hosts and acted as bioprotectors; (3) expanded insights into leveraging synergistic plant–microbe links to develop microbial inoculants that enhance plant growth while preserving biodiversity integrity and ecosystem safety, similar to biofertilizers.
If we consider the importance of studying these three areas, future research should focus on dissecting molecular plant–microbe interaction mechanisms under harsh environmental conditions, validating the impact of concurrent inoculations under combined biotic/abiotic stresses, identifying functional genes within beneficial microbes essential for the promotion and processing of organic and inorganic molecules, refining methodologies for large-scale deployment at contaminated sites, and exploring the commercial production of bioinoculants aimed at environmental decontamination [246, 247].
In recent decades, the study of molecular interactions between plants and microbes has undergone significant discoveries, as technological and intellectual advances that represent the state-of-the-art have allowed us to address questions at a level of detail that appeared unattainable two decades ago. Although many of these inter-relations benefit plant health, microbial colonization, and the environment, complex biochemical pathways are sometimes spontaneously activated and have opposing effects. Progress in biotechnological techniques, particularly those related to molecular genetics, has enriched our understanding of this type of dynamics among species belonging to different kingdoms. When our knowledge of other mechanisms and molecules involved in this phenomenon is further deepened, it will be possible to elucidate the evolutionary history of the individuals under study. Incoming scientific areas, such as proteomics, transcriptomics, metabolomics, and other omics sciences, would offer an opportunity to maximize the understanding and societal advantages of the plant–microorganism relationship in a theoretical and practical sense. Based on the above, future research on plant–microbe interactions should focus on developing specific biofertilizers, increasing genetic resistance to pathogens, studying the soil microbiota and its influence on agriculture, applying omics techniques, innovating biological control, analyzing the impact of climate change on these interactions, and exploring endophytic microbiota including the identification of new metabolic pathways, genes, and proteins. However, it is imperative to focus on addressing key inquiries that have not yet been comprehensively deciphered, for example, how various factors, such as beneficial and harmful microorganisms, abiotic stress, and inter-microbial dynamics, impact plant–microbe interactions and how these insights can further our understanding of resistance mechanisms, such as ETI, PTI, and non-host resistance? Similarly, in the field of applied sciences, can methodologies effectively transition basic research findings into practical applications for emerging crops and elucidate the molecular basis behind phenomena, including resistance protein-triggered cell death? In this regard, it is worthwhile to question how the complex and dynamic interactions between plants, microbes, and their environment influence plant health and disease and how this knowledge could be harnessed to improve agricultural practices and crop resilience in the face of changing environmental conditions. Finally, in an evolutionary context, what drives the evolution and diversity of pathogen strategies remains unclear. Given the variance in the number of effectors required for different pathogens, it is important to posit how these findings on binary plant–microbe interactions extend to complex ecological settings. Undoubtedly, despite the scientific limitations in this field of study, such as the complexity of both field and laboratory experimentations, as well as the difficulty in understanding the multivariate molecular–ecological dynamics between different species, questions such as these await future answers from the academic community.
CCC, TMM, and OGL performed the literature search and wrote the manuscript. ECV contributed to the literature search and figure design. JRR provided oversight, literature search, writing, and direction for the manuscript. All authors have participated sufficiently in the work to take public responsibility for appropriate portions of the content and agreed to be accountable for all aspects of the work in ensuring that questions related to its accuracy or integrity. All authors read and approved the final manuscript. All authors contributed to editorial changes in the manuscript.
Not applicable.
The authors thank the colleagues and reviewers who kindly read the manuscript and made valuable suggestions to improve its quality and content.
This research received no external funding.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.