





 , Zhigang Shang 1,2,*, Liaofeng Wang 1,2, Zuohao Ma 1,2, Jiajia Li 1,2
, Zhigang Shang 1,2,*, Liaofeng Wang 1,2, Zuohao Ma 1,2, Jiajia Li 1,21 School of Electrical and Information Engineering, Zhengzhou University, 450001 Zhengzhou, Henan, China
2 Henan Key Laboratory of Brain Science and Brain-Computer Interface Technology, 450001 Zhengzhou, Henan, China
†These authors contributed equally.
Abstract
Background: Exploring the neural encoding mechanism and decoding of motion state switching during flight can advance our knowledge of avian behavior control and contribute to the development of avian robots. However, limited acquisition equipment and neural signal quality have posed challenges, thus we understand little about the neural mechanisms of avian flight. Methods: We used chronically implanted micro-electrode arrays to record the local field potentials (LFPs) in the formation reticularis medialis mesencephali (FRM) of pigeons during various motion states in their natural outdoor flight. Subsequently, coherence-based functional connectivity networks under different bands were constructed and the topological features were extracted. Finally, we used a support vector machine model to decode different flight states. Results: Our findings indicate that the gamma band (80–150 Hz) in the FRM exhibits significant power for identifying different states in pigeons. Specifically, the avian brain transmitted flight related information more efficiently during the accelerated take-off or decelerated landing states, compared with the uniform flight and baseline states. Finally, we achieved a best average accuracy of 0.86 using the connectivity features in the 80–150 Hz band and 0.89 using the fused features for state decoding. Conclusions: Our results open up possibilities for further research into the neural mechanism of avian flight and contribute to the understanding of flight behavior control in birds.
Keywords
- pigeon
- flight state
- FRM
- LFP
- functional connectivity
The switching of external motion states exhibited by organisms, such as sudden acceleration and deceleration, often reflects changes in the internal state of the brain [1, 2]. Effectively decoding the typical states has significant theoretical and practical implications for the understanding of internal movement intention encoding mechanisms, prosthetic control, and functional rehabilitation. It is also crucial for the development of future closed-loop brain computer interface technologies based on internal neural information decoding and external stimulation encoding. The decoding of neural information related to motor intentions or motion states in the brain has been extensively studied in various species including rats [3], primates [4, 5], and birds [6, 7].
Previous studies have indeed demonstrated that rhythmic information present in the brain can effectively represent different motor intentions or states [8, 9]. For instance, Ahmadi et al. [10] successfully detected resting and force-generating time segments in the rat primary motor cortex using gamma (50–100 Hz) local field potentials (LFPs) during a key pressing task and accurately decoded the discrete rest/force states as well as continuous values of the force variable. Zhuang et al. [11] recorded LFPs using microelectrode arrays in the primary motor cortex (M1) of monkeys performing reaching and grasping tasks. Their mutual information and decoding analysis revealed that higher frequency bands (e.g., 100–200 and 200–400 Hz) carried the most information about the examined kinematics, suggesting the utility of LFPs, especially high-frequency bands, for controlling reach and grasp movements. Bundy et al. [12] used a hierarchical partial-least squares (PLS) regression model to predict hand speed, velocity, and position based on electrocorticography (ECoG) signals recorded from epileptic patients performing a 3D center-out reaching task. Their findings highlighted the importance of beta band power changes for classifying movement and rest, as well as local motor potential and high gamma band power changes for predicting kinematic parameters. Although a series of evidence shows that the movement state of organisms can be decoded by recording neural activities in the brain and extracting relevant rhythm features [13], most studies have been restricted by the signal acquisition equipment and the ability to examine only specific scenarios within the laboratory, leading to a large disparity between research and practical application.
Compared with rodents and primates, the pigeon possesses exceptional flying capabilities and is not constrained by ground environments [14], making it an ideal animal model for studying natural motion states, especially flight states. Notably, studies have demonstrated that micro-current stimulation in the formation reticularis medialis mesencephali area (FRM) of the pigeon brain can induce turning actions [15]. During flight, the direction of flight can be regulated by applying electrical stimulation to this specific brain area [16]. However, it is important to consider the animal’s autonomous movement intentions to avoid potential negative effects on regulation efficacy or physical fitness when external stimulation instructions do not align with the animal’s movement intentions. In this regard, it is crucial to investigate and understand the neural decoding of autonomous movement intentions during flight. However, it is worth noting that despite these promising findings, most of the existing research on decoding the different motion states of pigeons is restricted by the equipment used for signal acquisition and experiments can only be conducted in specific laboratory settings [17], allowing only limited exploration of flight processes.
Thanks to the advancements of integrated circuit technology and microcontroller
technology, signal recorders suitable for small animals in natural conditions
have been developed for use in related research. In 2019, Massot et al.
[18] introduced a new tool called ONEIROS, which enabled sleep research in small,
freely moving animals. They demonstrated the practicality of the device by
successfully recording a pigeon in an 8
We therefore used pigeons, and the FRM associated with their flight as the target region, to investigate the autonomous flight intentions in the bird brain during natural flight in this study. We used our wearable signal recording system to simultaneously record the neural signals in the FRM and the posture information of three pigeons during their free flying. Four typical flight states, including baseline, accelerated take-off, uniform flight, and decelerated landing, were defined via analysis of global positioning system (GPS) data. Using coherence analysis, brain function connectivity networks were constructed based on the multi-channel LFP signals corresponding to the various states. Based on the topological characteristic analysis results of the functional connectivity networks, distinct features in the gamma frequency band (80–150 Hz) were extracted for decoding the four typical flight states and the results were compared with the other bands. To the best of our knowledge, this is the first study focused on the neural information decoding of different outdoor flight states of pigeons, providing a valuable reference for further research on the neural mechanism of avian flight, navigation, and behavior control in birds.
In this study, a total of six adult homing pigeons (Columba livia, 450–500 g, unknown sex) obtained from a local supplier (Gongchuang Pigeon Co., Zhengzhou, Henan, China) were used. All pigeons were housed in a loft, provided with regular feeding by professionals and free drinking water in their normal living conditions. The pigeons’ diet was restricted during flight tasks until completion of the experiment. All experimental procedures adhered to the guidelines outlined in the Animals Act, 2006 (China), and aimed to ensure the ethical care and use of laboratory animals. The study protocol was approved by the Life Science Ethical Review Committee of Zhengzhou University (No. SYXK 2019-0002).
Once it was established that the pigeons were capable of reliably completing the
flight task, we implanted microelectrode arrays into the FRM of pigeons, in which
eight individually insulated tungsten microwires (California Fine Wire, Grover
Beach, CA, USA) with a 35-µm inner diameter and 150-µm spacing
between the wires (Kedou Brain Computer Technology, Suzhou, Jiangsu, China) were
used. The surgical procedure followed a similar protocol as described in our
previous study [20]. Initially, pigeons were anesthetized using a 3% sodium
pentobarbitone solution (at a dose of 0.12 mL/100 g body weight,
C
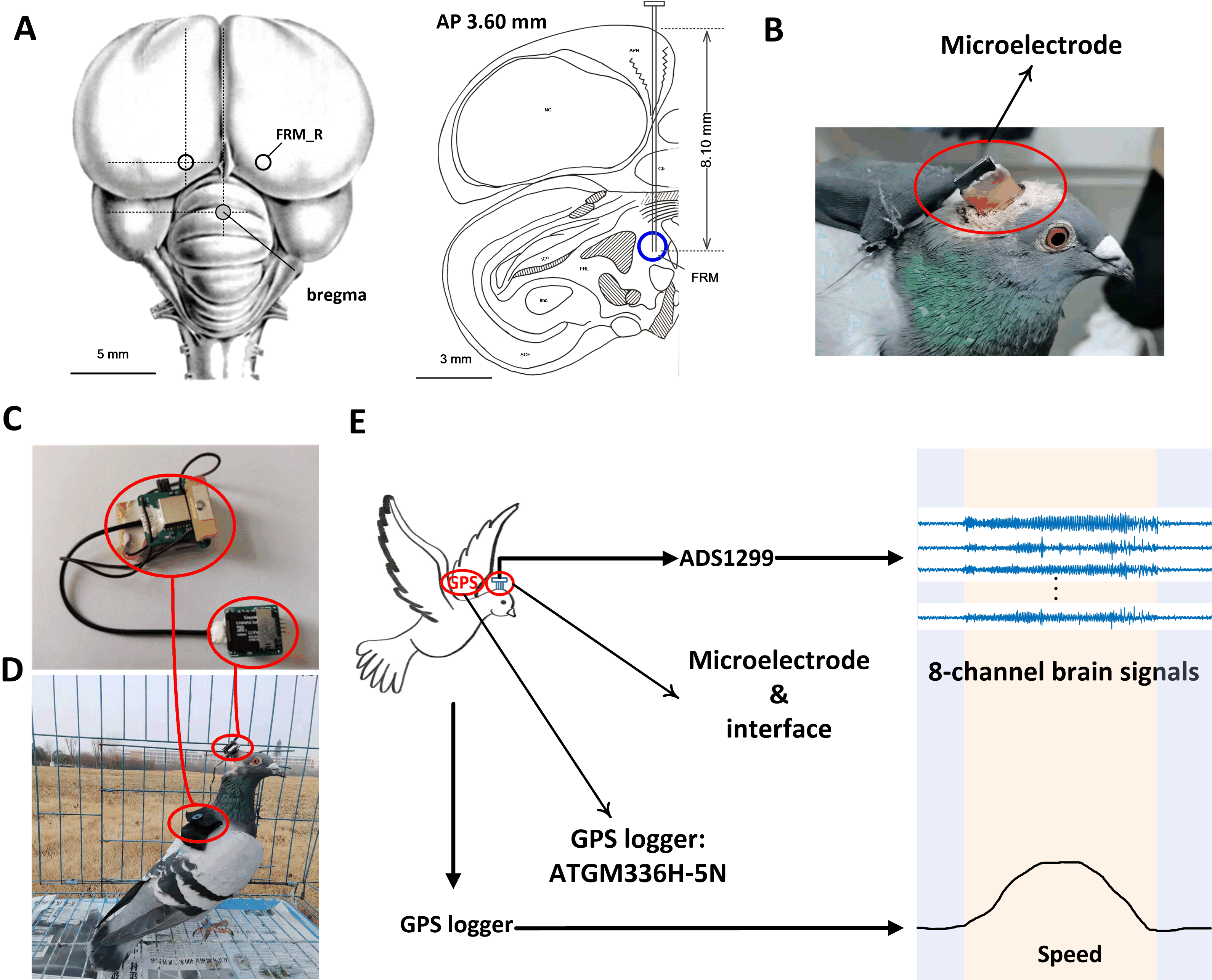 Fig. 1.
Fig. 1.Electrode implantation and data acquisition. (A) Location of microelectrode implantation. (B) Pigeon implanted with a microelectrode array. (C) Wearable data recording device. (D) Pigeon wearing the data recording device. (E) Diagram of data recording during free flight, in which the 8-channel LFP signals and the GPS data were recorded. AP, anteroposterior; FRM, formation reticularis medialis mesencephali; GPS, global positioning system; LFP, local field potential.
To simultaneously record the flight state related GPS data of pigeons and their
neural signals in the FRM, a wearable data recording device was developed, which
was described extensively in our previous publication [19] and is shown in Fig. 1C. The self-made device (Henan Key Laboratory of Brain Science and
Brain-Computer Interface Technology, Zhengzhou, Henan, China) weighed 13.6 g and
consisted of a GPS module (ATGM336H-5N with sampling range of 1–10 Hz,
positional accuracy
Before recording, the pigeons were trained to perform in the study by wearing backpacks to increase their capacity for bearing weight for 2 weeks. The study mainly consisted of a pre-experiment and a formal experiment. In the first week, the pigeons were loaded with 20-g weight during both walking and flying to adapt to the burden of the device. In the second week, the pigeons were released daily on short-distance (2 km) flights away from the loft. After the pre-experiment, the pigeons wore our data recording device as shown in Fig. 1D. During the whole flight process, including baseline, accelerated take-off, uniform flight, and decelerated landing, 8-channel neural signals and GPS data were recorded synchronously as depicted in Fig. 1E.
We performed GPS and LFP data acquisition during the flight of pigeons from the release site to the home loft as shown in Fig. 2. The sampling rate of GPS data is 10 Hz and that of the LFP signal is 1000 Hz. The GPS data provides essential information regarding the pigeon’s latitude and longitude coordinates during free-flying, as well as velocity data. The acceleration of the pigeon during flight can be further calculated based on the velocity data as follows:
When pigeons are flying outdoors, the neural signals can be prone to interference from baseline drift and noise generated by their flapping wings. To address these challenges, it is important to eliminate the trend terms from the neural signals and account for the non-stationary nature of the signal. Firstly, to accomplish baseline drift, discrete wavelet transform (DWT) was employed. The DWT is defined as follows:
where
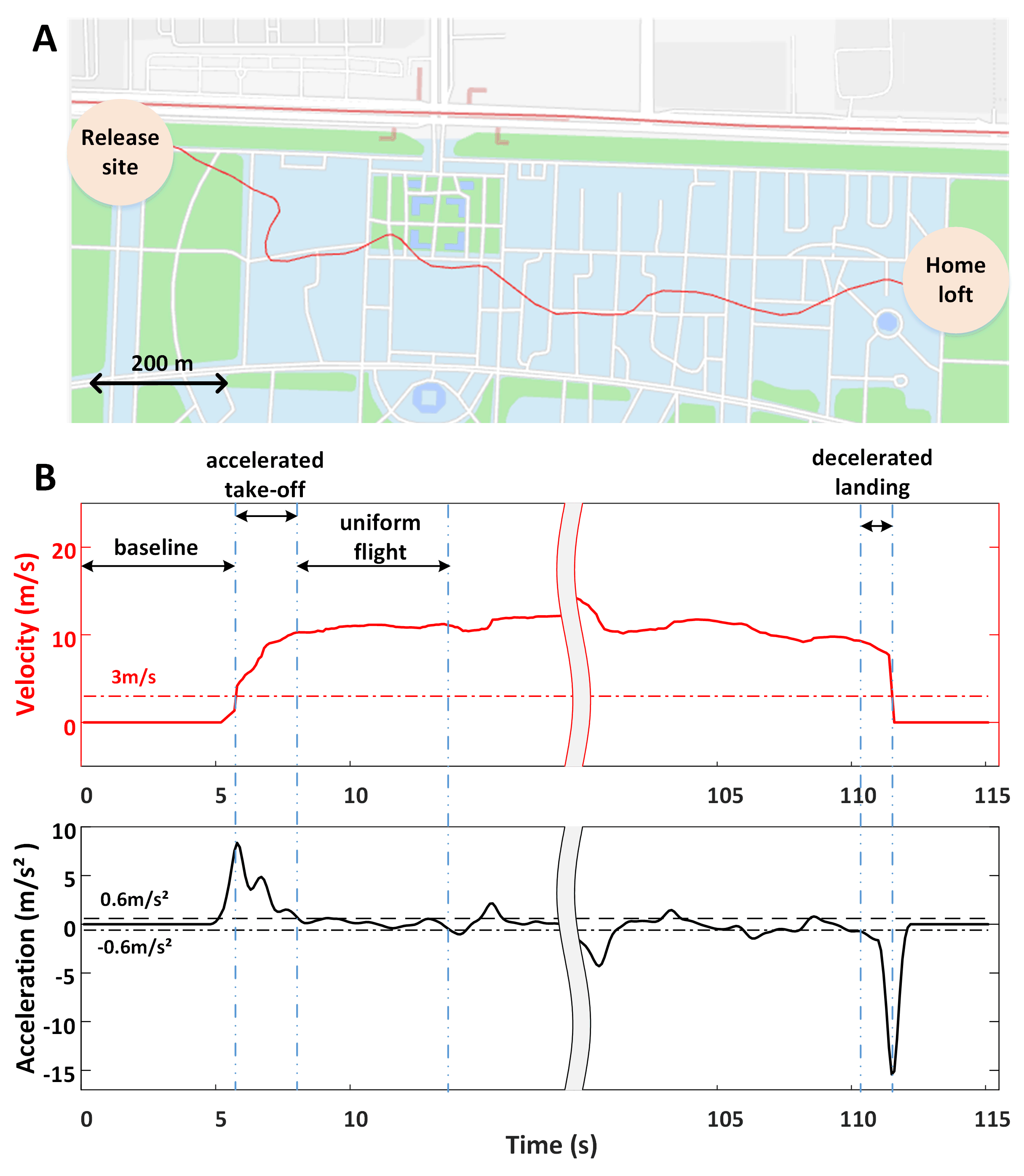 Fig. 2.
Fig. 2.Recorded GPS track and definition of typical states. (A) The track is displayed on the map, where the red line indicates the actual flight route from the release site to the home loft. (B) The velocity and acceleration curve calculated after pre-processing the GPS data, in which the solid red line represents the change of speed, and the solid black line represents the corresponding change of acceleration, marking four typical motion states based on the velocity and acceleration curves.
To address the issue of flapping noise in the neural signals, the primary method
used is priori variational modal decomposition (VMD). This technique aims to
extract intrinsic mode functions or modes of oscillation from the signal without
relying on fixed functions for analysis. This makes it particularly effective for
processing non-smooth and non-linear neural signals. The fundamental concept
behind VMD is to construct and solve variational problems [22]. Previous studies
have indicated that pigeons exhibit vibrations in the frequency range of
approximately 3–10 Hz during flight [23, 24]. According to this prior knowledge,
modal components within this frequency range are discarded. The remaining modal
components are then reconstructed to obtain the processed LFP signal. In the
current study, our multi-channel LFPs were filtered using a zero-phase bandpass
filter to obtain signals in the following four frequency bands:
In this study, we focused on four typical flight states of pigeons: baseline,
accelerated take-off, uniform flight, and decelerated landing. Previous studies
have commonly used GPS speeds greater than 5 or 10 km/h to identify pigeons in
flight [25, 26]. However, in this study, we considered the actual movement of the
pigeons during flight and the accuracy of the sensors to define the above typical
states. For small fluctuations in speed within a certain range (
(1) Baseline: This state represents a continuous process in which the pigeon’s speed is less than 3 m/s. It typically corresponds to the pigeon being at rest on the ground.
(2) Accelerated take-off: This state represents a continuous process in which
the pigeon’s speed is greater than 3 m/s, and the acceleration is greater than
0.6
(3) Uniform flight: This state represents a continuous process in which the
pigeon’s speed is greater than 3 m/s, and the absolute value of the acceleration
is less than 0.6
(4) Decelerated landing: This state represents a continuous process in which the
pigeon’s speed is greater than 3 m/s, and the acceleration is less than –0.6
We then explored the connectivity between multiple channel LFPs in the FRM to find effective features on the scale of local network connections for decoding the four states. This is accomplished by generating a connection matrix that represents the relationship between different channels or brain regions. Subsequently, the topological properties of the brain function connectivity network are analyzed using principles from graph theory. In this study, the connectivity network was established by quantifying the coherence between channels. The coherence coefficient is calculated as follows:
where
To measure the topological characteristics of the connectivity network, the clustering coefficient (Cc), global efficiency (Ge), and average path length (Apl) were calculated specifically. The clustering coefficient quantifies the tendency for neighboring channels to form interconnected clusters [27]. It provides a measure to evaluate the level of network integration and can be defined as follows:
where N represents the total number of nodes in the network,
The global efficiency provides insights into the efficiency of information flow across the network and is defined as the average value of the path length between two nodes in the network as follows:
where
The average path length represents the average number of edges required to traverse between any two channels in the network. It provides a measure of the global efficiency of information transmission or integration across the entire network and can be defined as:
where
We analyzed the differences in connectivity characteristics of the four typical states and extracted effective features for the decoding analysis. Support vector machine (SVM) models can better solve the problem of small sample classification and have been successfully applied in many fields [28]. Thus, an SVM was used in this study for typical flight state decoding and the performance was assessed through ten-fold cross-validation.
Our results are given in the form of mean
We effectively collected the data of three pigeons (P02, P03, and P06), while three other animals (P01, P04, and P05) dropped out of the study due to electrode implantation failure or poor signal quality. The pigeons were released from a location about 2 km away from the loft, and one whole flight process recorded by GPS data of P02 is shown in Fig. 2A. The speed and acceleration information of pigeons was calculated based on the GPS data to better characterize their specific states. Four typical motion states including baseline, accelerated take-off, uniform flight, and decelerated landing were then defined according to the above calculations, which are shown in Fig. 2B.
For the recorded neural signals, we have shown a pre-processing example of 5 s of data during the study of P02. Fig. 3A shows the raw signal obtained from one of the acquired channels, in which there is a baseline drift. We then applied the DWT to remove the drift, and the result is shown in Fig. 3B. Fig. 3C shows the time-frequency analysis results of the LFPs, and there is a noticeable wing coupling component with a frequency range of approximately 5–10 Hz. Next, we employed VMD to mitigate the wing-flapping noise; Fig. 3D displays the signal and time-frequency analysis results after VMD.
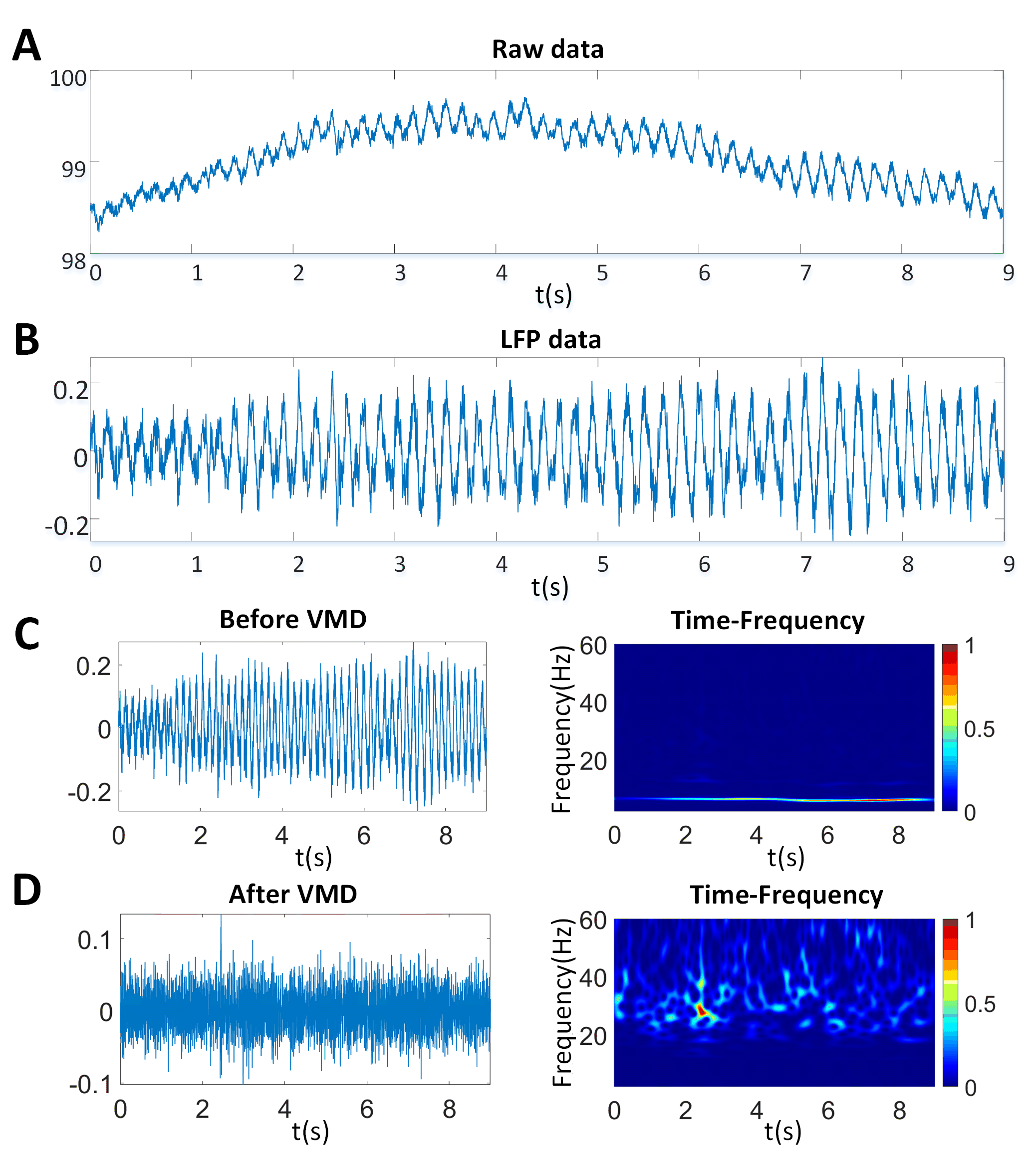 Fig. 3.
Fig. 3.Neural signal pre-processing. (A) Raw neural signals. (B) LFP signal processed to remove baseline drift by wavelet technology. (C) LFP signals before VMD in time and time-frequency domain. (D) The LFP signals after VMD in time and time-frequency domain. VMD, variational modal decomposition.
Each pigeon underwent a different number of sessions, with 12, 13, and 15 sessions respectively in the current study (each session represents one flight experiment). According to the definition of the typical states, we obtained the corresponding neural signals and divided them into segments of 500 ms windows. Table 1 shows the experimental data for three pigeons.
| Pigeon | No. of channels | No. of sessions | No. of signal segments | |||
| Baseline | Accelerated take-off | Uniform flight | Decelerated landing | |||
| P02 | 8 | 12 | 86 | 66 | 94 | 83 |
| P03 | 8 | 13 | 116 | 92 | 97 | 89 |
| P06 | 8 | 15 | 106 | 108 | 99 | 90 |
We first compared the results of the power spectrum analysis at different
frequency bands in four typical states and the results are shown in Fig. 4A. We
observed distinct power spectrum differences among these four typical states,
with the most notable discrepancies occurring in the
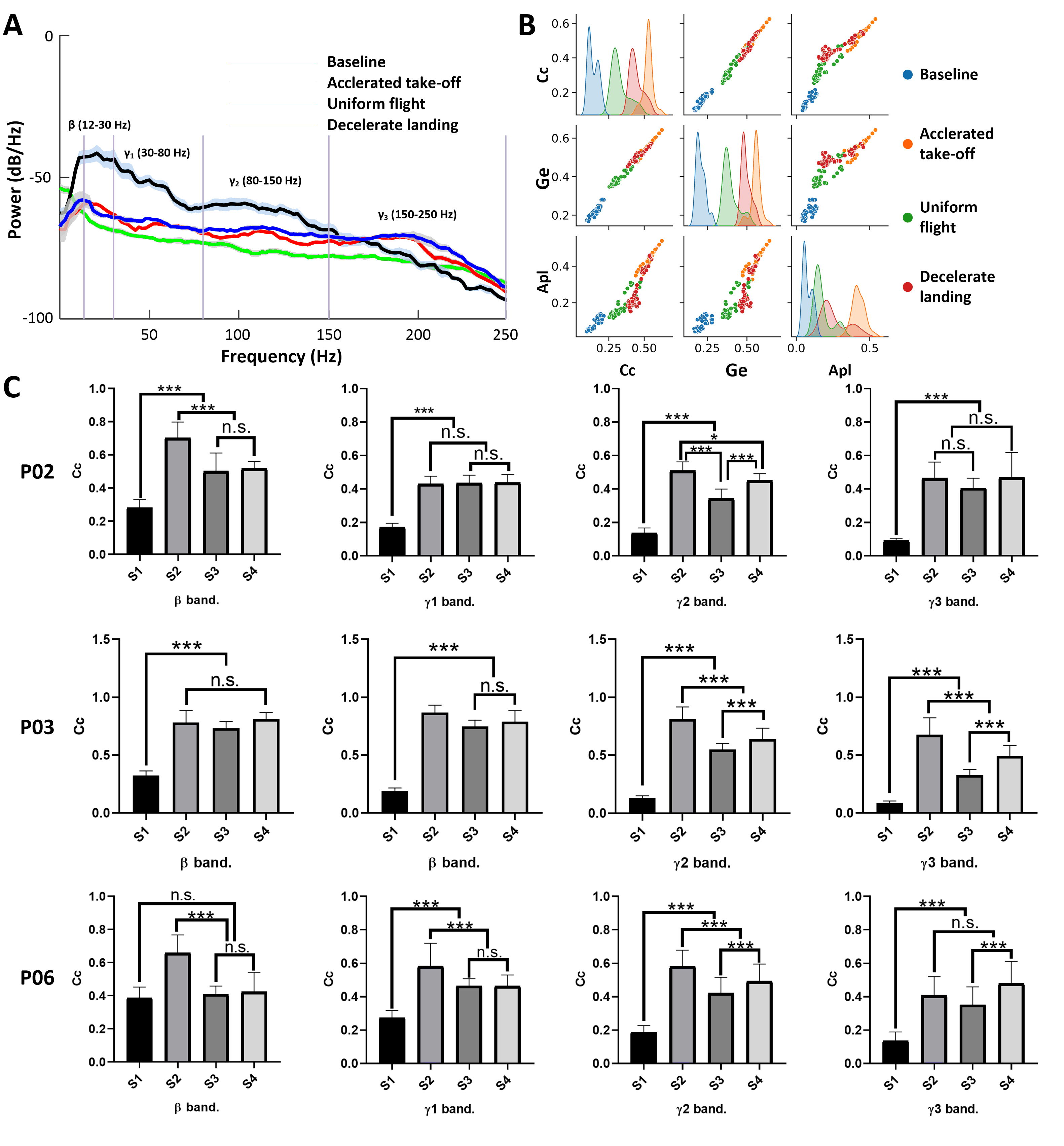 Fig. 4.
Fig. 4.Power spectrum and functional connectivity analysis results.
(A) Power spectrum analysis of the LFPs during four typical states. (B)
Functional network characteristic distributions of the
We then calculated the clustering coefficient, global efficiency, and average
path length of the functional networks for the four states of three experimental
pigeons using graph theory. The statistical analysis results of the clustering
coefficient for the bands are shown in Fig. 4C. For the inter-state comparisons,
it is apparent that the clustering coefficients for all three typical flight
states of the three pigeons are significantly higher (Kruskal-Wallis test,
p
The functional connectivity characteristics from the FRM showed variability
across four typical states. We therefore sought to ascertain with what accuracy
could the pigeon’s current states during flight be determined based on the
functional connectivity features. To answer this question, a multi-classified SVM
was used to decode the pigeon’s states and the analysis results are shown in Fig. 5 (detailed results are shown in Supplementary Table 1). The typical
flight state decoding accuracy of three pigeons is shown in Fig. 5A. The results
indicate that the accuracy varies across the four different frequency bands. The
best average decoding results for P02 and P06 are achieved in the
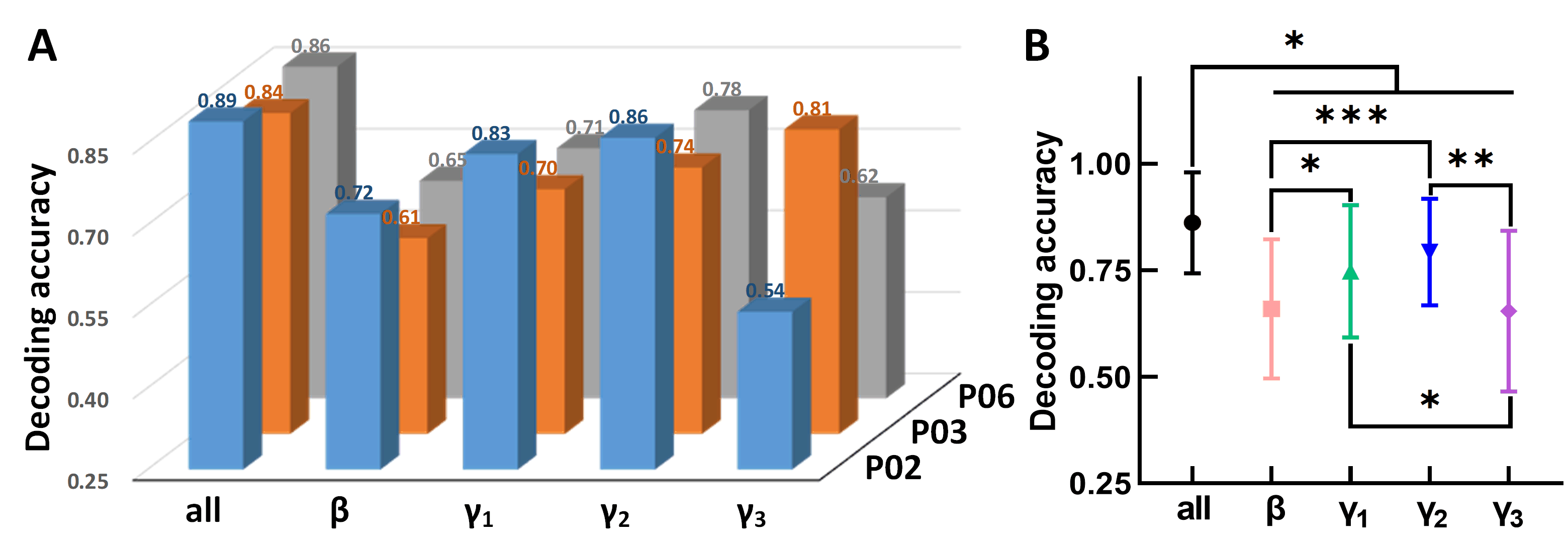 Fig. 5.
Fig. 5.Decoding analysis results using functional connectivity
features. (A) Decoding accuracy of typical flight states of three pigeons. (B)
Decoding results across all pigeons (* p
The statistical decoding results across all three pigeons are shown in Fig. 5B.
We observed the best decoding performance using all features in the four bands
across the three pigeons, which was significantly higher than those using the
features in other single frequency bands (0.86
This study explored the neural activity analysis and state decoding of pigeons during flying under natural outdoor conditions. The results revealed that the FRM of pigeons encoded the flight related information, particularly the state switching related information. We found that high-frequency functional connectivity in the FRM, especially in the gamma band (80–150 Hz), better characterized the flight state of the pigeon compared with the other bands. The decoding analysis results also provide insights into the neural processes underlying different states during pigeons’ flight. It should be noted that previous research has linked high-frequency rhythm activity to multiple functions including motor, attention, and decision-making [29, 30, 31]. Our findings align with previous studies in other animal species, suggesting the relevance of the gamma band in locomotion [10, 11].
Despite the findings, there are limitations in the study, including but not limited to verifying the validity of the data, behavioural control based on flight decoding, and further optimization of site localization by staining slices. First, our study contains data for analysis from only three pigeons, constrained by the difficulty in constructing avian flight experimental paradigms in large-scale natural environments and the difficulty in collecting and denoising neural signals under free-flying conditions. In addition, incomplete experimental data problems as mentioned caused by difficult electrode surgical implantation and postoperative recovery, as well as accidental loss or death of pigeons, also pose challenges for us to expand the sample and data size. Hence, more pigeons and data are needed to further improve the accuracy and robustness of our results. Furthermore, it is known that the direction of flight can be regulated by applying electrical stimulation to this specific brain area during flight [15, 16, 32]. Applying different stimulation parameters to pigeons can also induce varying turning angles [33]. By stimulating individuals, researchers can study flapping flight and collective behaviour in pigeon flocks [34]. Further study focused on the behavioural control of birds based on flight state recording and decoding should therefore be carried out. Additionally, further detailed exploration of the structural characteristics of the FRM with the help of stained slices, together with examination of the specific functions of the left and right hemispheres [35, 36] of the FRM and their relationship with autonomous steering intention, could enhance the understanding of motion state decoding during bird flight.
Regarding the remaining issues and areas for further study, it is indeed important to consider factors beyond speed changes and incorporate different steering information during flight [37]. In terms of brain regions and physiological measurements, it is worth noting that other brain regions may provide targets for future movement intention decoding. For example, the activity in the opticus principalis thalami (OPT) has been previously linked to computing the distance-to-collision of approaching surfaces in pigeons [35]. Furthermore, considering correlations between other kinds of recordings apart from GPS tracking measurements alongside oscillatory recordings could provide valuable complementary information. In addition, there are also potential limitations in the experimental period regarding the individual characteristics of the pigeon and environmental factors, the data pre-processing process of artifact removal, and the decoding process using the SVM. We aim to eliminate these negative influencing factors to better optimize our future studies.
Overall, our results contribute to filling a knowledge gap and provide valuable support for further studies on avian motion state decoding and flight behaviour control, supporting closed-loop brain-computer interfaces for birds that consider both internal neural encoding and external behavioural decoding. In addition, our results show that our recording device can provide valuable insights for long-distance flight and navigation related research.
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
LY, ML, and ZS designed the study. LY and EM performed the research. LY and LW designed and assembled the device. LY, EM, LFY, and ZM conducted experiments. LY, LFY, and ML analyzed the data. LFY and JL performed the validation and visualization. LY and ML performed the investigation. LY and ML prepared the original draft. ML and ZS reviewed and edited the draft. All authors have read and agreed to the published version of the manuscript. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
The study was conducted in accordance with the Declaration of Helsinki, and the protocol was approved by the Life Science Ethical Review Committee of Zhengzhou University (No. SYXK 2019-0002).
Not applicable.
This work was supported by the National Postdoctoral Researcher Program (GZC20232447), the National Natural Science Foundation of China (62301496), and the Key Scientific and Technological Projects of Henan Province (232102210098, 222102310223).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.