







1 Department of Medical Laboratories, College of Applied Medical Sciences, Qassim University, 51452 Buraydah, Saudi Arabia
Abstract
Biofilms, which consist of microorganisms enclosed in an extracellular polymeric material (EPS), hold immense importance in the fields of environmental research, industry, and medicine. They play a significant role in ecosystem dynamics and stability, but they also pose issues such as biofouling, corrosion, and pollution. Biofilms in medical environments are linked to persistent infections and elevated healthcare expenses. The EPS matrix plays a crucial role in maintaining the structural integrity and antibiotic resistance of these structures. The research primarily investigates the role of the EPS matrix in facilitating horizontal gene transfer among biofilm communities, with a particular emphasis on EPS and its impact on this process. The process is recognized as a pivotal mechanism in the emergence of antibiotic resistance, underscoring the crucial function of EPS in the dynamics of biofilms. The analysis also highlights the significant financial constraints caused by biofilms in several industries. Biofilm-associated infections in the healthcare sector result in escalated treatment expenses and extended hospitalization periods. In an industrial context, biofilms have a role in increasing maintenance expenses and product contamination, emphasizing the need for efficient management solutions. This review presents the most recent progress in biofilm research, emphasizing the utilization of sophisticated imaging tools and molecular methodologies. In addition to conventional imaging techniques, the research explores the utilization of sophisticated molecular tools, such as DNA and RNA sequencing, in conjunction with proteomics. These approaches are essential for assessing the genetic and metabolic mechanisms that regulate biofilm development and antibiotic resistance. The review underscores the significance of employing an interdisciplinary methodology in the study of biofilms. By incorporating a range of approaches, such as sophisticated imaging and molecular analysis, a comprehensive understanding of biofilm dynamics may be achieved. This approach also opens up possibilities for developing novel solutions to address the negative impacts of biofilms on health, industry, and the environment.
Keywords
- biofilms
- bacteria resistance
- tolerance
- biofilm control
Biofilms are intricate arrangements of microbial life that form elaborate structures, proliferating across various habitats and surfaces. These communities are composed of one or more types of microorganisms. They are enveloped by a self-produced extracellular polymeric substance (EPS) matrix [1], which provides both structural support and robust resistance to diverse stressors, including antibiotic treatments [2, 3].
The indispensability of biofilms is manifested in their essential role in various contexts. Biofilms in industrial cooling systems and aquatic ecosystems, for instance, demonstrate their dynamic flexibility by adapting their microbial composition and functionality to withstand the deleterious effects of pollutants [4, 5]. The medical industry is profoundly impacted by the adaptability of biofilms, which pose significant challenges in the treatment of infections, particularly those caused by bacteria like Staphylococcus aureus and Streptococcus mutans. These organisms create biofilms that are both robust and contribute to the persistent nature of infections due to their viscoelastic properties [6, 7, 8].
Biofilms are colonies of bacteria that possess a remarkable ability to display both fluid-like and solid-like characteristics under mechanical stress [9]. This unique viscoelastic behavior is due to an EPS matrix surrounding the bacterial cells [10]. Because of their viscosity, biofilms are highly resistant to flow and deformation [11, 12], yet they also possess elastic properties that allow them to return to their original shape after deforming [13]. These viscoelastic properties play a crucial role in biofilms’ development, stability, and response to their environment. They are closely linked to the structural constituents present in the EPS matrix [9, 14].
The existence of biofilms relies heavily on their composition and structural characteristics. Research has identified that factors such as rough surface texture, extracellular DNA (eDNA), polysaccharide, and protein materials are crucial in their ability to collect infections and resist elimination [15, 16, 17]. Various approaches, such as phage therapy and phototherapy, have been proposed to combat these attributes, showing promise in the dispersion and prevention of biofilm formation [18, 19].
Biofilms pose significant challenges to environmental management, especially in agricultural streams where controlling them is crucial for ecological management tactics. Furthermore, it is essential to understand the micro-scale habitat conditions and interactions within biofilms to advance research and promote environmental stewardship [20]. Biofilms also impact coral reef ecosystems, which change composition when exposed to long-term nutrient enrichment, suggesting a complex interaction between biofilms and their surroundings [21].
Microbial extracellular polymeric substances serve the purpose of not only ensuring structural stability but also protecting the local bacteria from external factors such as antibiotics. This contributes to their reputation as a difficult obstacle in eradicating persistent infections [1, 22]. Different functions of microbial extracellular polymeric substances are presented in Fig. 1.
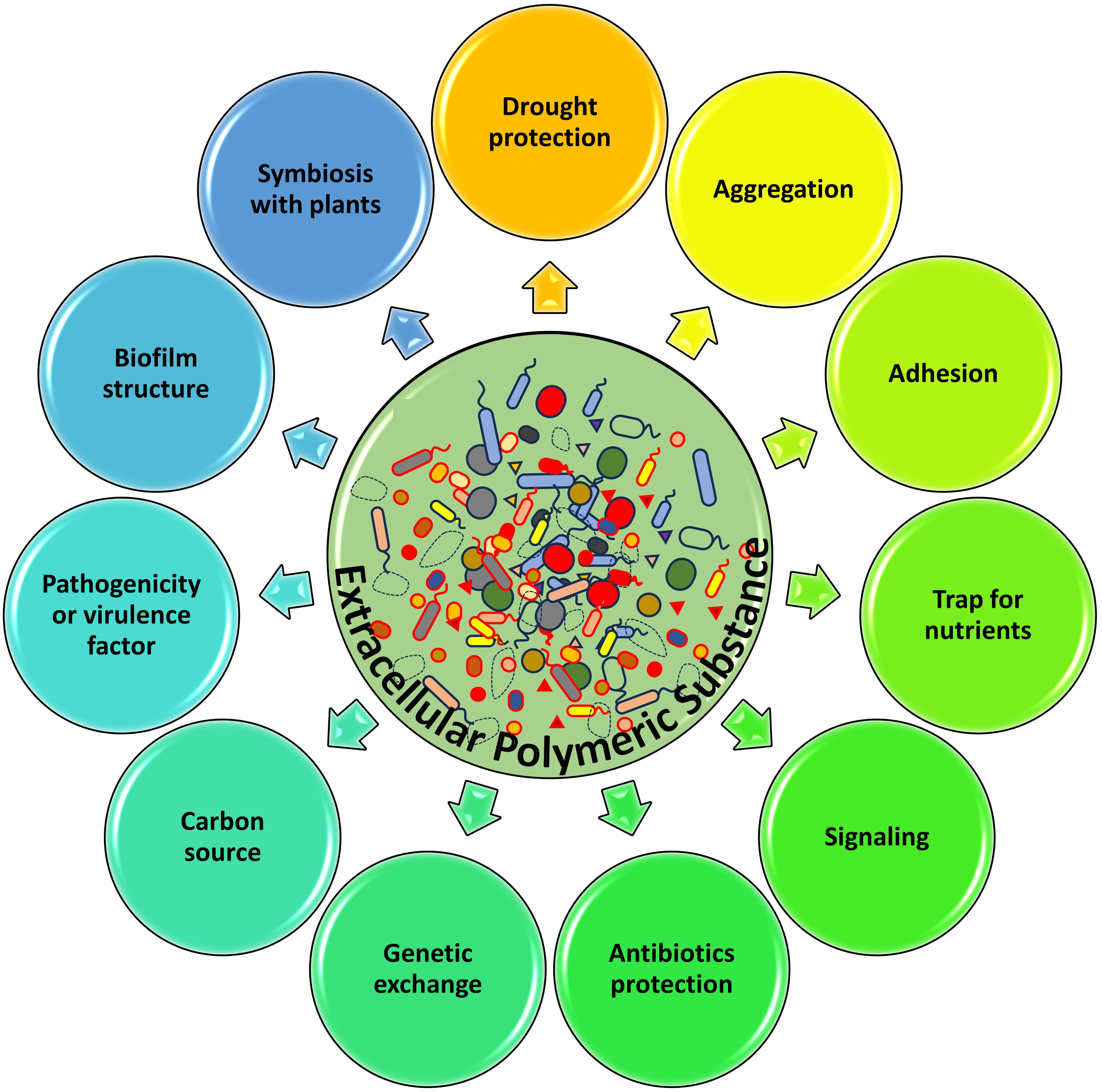 Fig. 1.
Fig. 1.Diagramatic representation of different functions of microbial extracellular polymeric substance.
Biofilms play a significant role in various illnesses, particularly those associated with medical equipment, in the clinical setting. S. aureus, for instance, can form biofilms on these devices, leading to prolonged and potentially hazardous infections [23, 24]. Chronic diseases are often associated with the existence of biofilms, and the inability of current treatments to effectively combat bacteria residing within them underscores the urgent need for innovative therapeutic approaches [25].
Biofilms have undergone extensive research concerning specific medical conditions. For example, studies on S. aureus biofilms in cystic fibrosis have yielded valuable insights into their importance, techniques used to analyze them, and approaches to prevent their development [26]. Biofilms’ involvement in endodontics and chronic rhinosinusitis has also been investigated, revealing their role in developing both conditions [27].
This review delves into the complexities of biofilms, with a focus on the significance of EPS, molecular methodologies, cutting-edge imaging methods, and the ongoing battle against antibiotic resistance. Biofilms’ enduring traits and diverse impacts across multiple domains emphasize the importance of interdisciplinary research and the need for innovative strategies to manage and prevent their formation efficiently.
Biofilms have a significant and versatile influence that can be observed in various industries, environments, and healthcare facilities, presenting challenges and opportunities for innovation and improvement.
Biofilms have a significant and varied impact on many industrial processes and infrastructure, both in industrial and environmental settings. These films, found on natural and man-made surfaces, play a crucial role in the strength and survival of bacteria [28], making them a challenging presence in industrial environments that can hinder cleaning and sanitation protocols.
In industrial settings, biofilms have been found to impede the effectiveness of cleaning and sanitization methods that rely on physical or chemical agents. The resilience of these surfaces against conventional cleaning techniques presents a significant challenge in achieving and sustaining cleanliness and sterility [29]. Also, biofilms can cause changes at the metal-environment interface, exacerbating the corrosion process [30]. This underscores the importance of addressing this factor to ensure the longevity and efficacy of infrastructure and equipment in various industries.
Gaining a comprehensive understanding of how biofilms affect industrial processes and infrastructure is essential in addressing these challenges. The engineering practice of biofilms in bioplugging has become prominent in various industries, environmental management, and water research [31].
Biofilms are an innovative engineering technique that utilizes microbial communities to regulate and manipulate fluid movement in various systems [32]. These structures consist of consortia of microbes that adhere to surfaces and generate a defensive matrix. Biofilms are commonly found in damp environments and are being developed to block or seal small openings and cracks in various systems, known as bioplugging. As the biofilm grows, it impedes the passage of fluids or gases within these cavities [31].
Bioplugging has numerous benefits for the oil and gas sector, particularly improving oil recovery. It achieves this by obstructing undesired water pathways in reservoirs, diverting more oil onto extraction wells, and enhancing total productivity [31, 33]. Additionally, bioplugging is helpful in water management and treatment, effectively sealing leaks in distribution systems and reducing water loss and contamination [34, 35]. Environmental remediation also utilizes bioplugging, where biofilms serve as biological barriers to confine or break down pollutants [36].
Additionally, it is vital to consider the internal complexities of biofilms when predicting their responsiveness, particularly in bioclogged porous systems found in industrial and environmental settings. This underscores the importance of a deeper understanding of biofilm composition and behavior in diverse industrial settings [37].
Biofilms are well-known in the healthcare industry for their involvement in persistent infections and their ability to resist antibiotics [38, 39]. These biofilms can shield bacteria from the effects of antibiotics, making treatment more complex and leading to prolonged illnesses and increased healthcare costs. Biofilms that develop on medical devices, such as endotracheal tubes [40], pose a significant threat of hospital-acquired infections, a severe issue in clinical settings. The process of bacteria colonizing surfaces and the subsequent development of biofilms, as described by Wang et al. [41], is a significant concern, particularly in the medical field. Biofilm formation plays a critical role in the bacterial life cycle, affecting host interactions and aiding in the emergence of drug resistance and chronic infections [42].
Although biofilms can have detrimental impacts, they can also benefit in specific contexts, like agriculture and environmental management [43]. Within the agricultural industry, biofilms can bolster plant growth and enrich soil health. Meanwhile, in ecological management, biofilms can be utilized in bioremediation methods to eradicate pollutants.
Highmore et al. [44] reported that biofuel formation has significant economic implications and estimated the total annual cost of biofilms to be around USD 5 trillion. These financial repercussions extend to crucial sectors like healthcare, food and water safety, marine industries, and other industrial operations [44]. Furthermore, Dutta et al. [45] found that biofilms can significantly reduce the efficiency and productivity of many industrial processes, leading to considerable financial losses and operational obstacles.
The process of biofilm production is complex and nuanced, with many genetic, regulatory, and signaling factors at play. Extensive research has emphasized the significance of genes responsible for efflux pumps, pili formation, and quorum sensing in the establishment of biofilms [46]. Different molecular mechanisms involved in biofilm formation are described in Fig. 2.
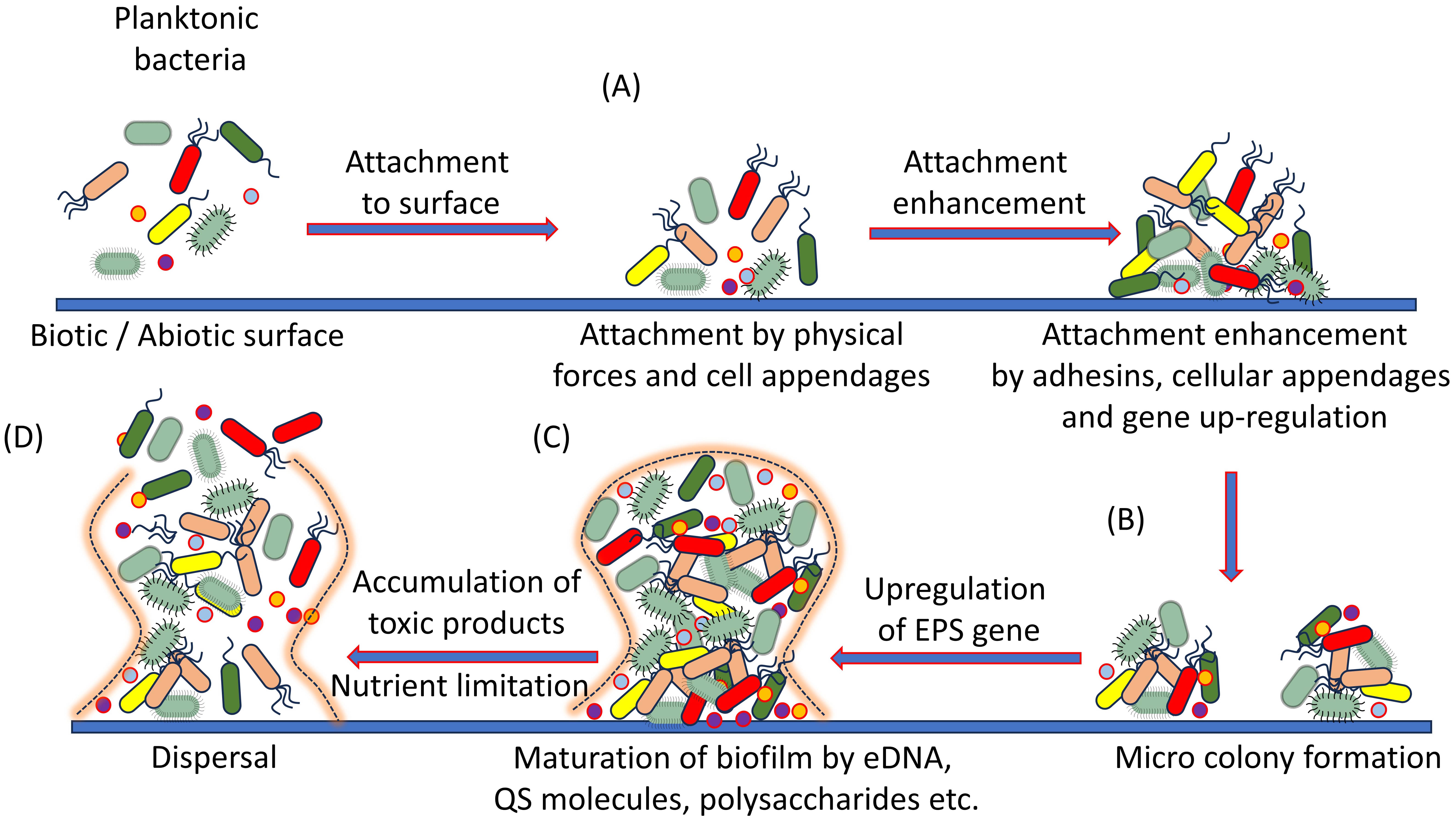 Fig. 2.
Fig. 2.Step by step biofilm formation mechanism. The four stages of biofilm formation include: (A) On compatible surface reversible attachment of free-living bacteria by their cell appendages. It is followed by enhanced irreversible attachment by adhesins. (B) Discrete micro colony formation with around 100 bacteria/cluster by the gene upregulation for maintained attachment. (C) Upregulation of extracellular polymeric material (EPS), release of Qureum Sensing (QS) molecules, polysaccharides and eDNA promote the maturation of biofilm. (D) after some time, depletion of nutrients and accumulation of toxic substances push the biofilm colonies for dispersal of bacteria.
In-depth transcriptome studies on certain bacterial species, such as Vibrio Vulnificus, have revealed the critical role of novel genes in establishing biofilm and rugose colonies [47]. Rugose colonies often exhibit a bumpy or rough surface texture. This ability to form rugose colonies and biofilm is commonly observed in vibrios, and a rugose colony phenotype typically indicates increased synthesis of exopolysaccharides [47, 48]. Activating specific genes, namely cabH and brpN, is critical for forming robust biofilms and rough colonies in Vibrio vulnificus [47]
Furthermore, specific genes like fbe, aap, and icaA are emphasized for their importance in bacterial biofilm formation in Staphylococcus epidermidis [49]. The significance of quorum sensing mechanisms in the inception of biofilm formation highlights the crucial role of cellular communication in these processes [50]. Additionally, studies have shed light on the role of transcription factors, including Mac1p and the FLO gene family, in regulating biofilm formation in Saccharomyces cerevisiae [51, 52]. This supports the notion of a complex genetic and regulatory network involved in this process. Recent research has provided further insights into the specific genes that impact the composition of the biofilm matrix and the transition between biofilm and planktonic stages, leading to a deeper understanding of the biofilm lifecycle [53, 54].
In the development of biofilms, quorum sensing serves as a crucial regulatory mechanism that enables bacteria to communicate with one another. This intricate process involves N-acyl homoserine lactones (AHLs), which function as signaling molecules in Gram-negative bacteria to control population density and synchronize activities [55]. The generation, identification, and reaction to external chemical messengers, known as autoinducers, are essential for this process [55]. Pseudomonas aeruginosa and other Gram-negative bacteria rely on AHLs for quorum sensing to regulate gene expression, biofilm formation, and virulence factors [56]. Additionally, AHLs facilitate synchronized group behavior within a population of Gram-negative bacteria [57]. The LuxR protein plays a pivotal role in detecting AHLs as signaling molecules in the quorum-sensing system of Gram-negative bacteria [58].
Gram-positive bacteria use small autoinducing peptides (AIPs) for quorum sensing, unlike Gram-negative bacteria. Quorum sensing in Gram-positive bacteria relies on peptides, specifically AIPs signaling molecules, to regulate collective characteristics, such as pathogenicity [59]. These peptides are crucial in controlling group behaviors and related characteristics based on cell density [60]. To communicate between cells, Gram-positive bacteria secrete oligopeptides known as quorum sensing peptides (QSPs) [61]. Additionally, they express and identify antimicrobial peptides, specifically cecropin B-like peptides, which are vital to their defense systems [62]. These findings highlight the significance of peptide-based quorum sensing modulators in Gram-positive bacteria and their potential for treating bacterial illnesses.
Research has shown that the levels of phosphorylated Spo0A, a key regulator, impact the initiation of biofilm formation. High concentrations of Spo0A~P promote sporulation, while lower levels kickstart the biofilm formation process in Bacillus subtilis [63, 64]. Furthermore, mild malnutrition indirectly spurs the development of biofilms by stimulating the expression of genes that control the extracellular matrix in a specific cell group via the SinI-SinR-SlrR network [65]. Additionally, Spo0A has a negative regulatory effect on AbrB, a transcription factor that also negatively impacts the process of biofilm formation in B. subtilis [66].
The concentration of phosphorylated Spo0A within bacterial cells is a deciding factor in developing spores or biofilms. When the levels of phosphorylated Spo0A are high, spore formation is promoted, while moderate levels encourage the development of biofilms [67]. Additionally, the disruption of Spo0A binding to the accDA promoter region has been observed to impact biofilm production, indicating the significant role of genes regulated by Spo0A in this process [68].
Adenosine triphosphate (ATP) has been identified as a critical player in bacterial adherence and biofilm formation by promoting cell lysis and releasing eDNA [69]. The presence of ATP-rich microenvironments within biofilms has been suggested as a means to specifically target and eliminate bacteria, emphasizing the importance of ATP in biofilm elimination [41]. Furthermore, c-di-AMP, a second messenger molecule produced from ATP, has been recognized as essential for biofilm formation [70]. Studies have also demonstrated the significant role of ATP in creating biofilms in Gram-Negative Bacteria found in Antarctica, further highlighting its influence on biofilm development [71].
Transcriptome analysis has revealed the involvement of ATP and the ABC transport system in regulating biofilm development in Vibrio parahaemolyticus [72]. Moreover, inhibiting the MsrA drug efflux pump in Staphylococcus saprophyticus has been found to limit biofilm production and the quorum-sensing system, and this inhibition has been associated with ATP [73].
The impact of proteins and extracellular molecules on biofilms is significant. For instance, in P. aeruginosa biofilms, the attachment of matrix protein CdrA to Psl enhances biofilm production and aggregate stability [74]. Similarly, stress-related proteins like LexA, Hfq, and DnaK have been found to reduce biofilm formation in Clostridioides difficile mutants [75]. Moreover, proteins such as TasA within the biofilm matrix are capable of forming fibers that create the rough surface of the biofilm, which is crucial to its structure [76]. Additionally, OprF, a protein found in P. aeruginosa biofilms, is a significant matrix protein that affects the levels of biofilm matrix eDNA based on nutrient availability [77].
It has been discovered that proteins and eDNA play a role in creating the biofilm matrix and enhancing resistance against anti-biofilm methods in Prevotella species [78]. A study on the pathogenic yeast Candida albicans examined the role of proteins, such as Pra1 and Zrt1, in the biofilm formation process, revealing their importance [79]. The extracellular aminopeptidase PaAP is also highly prevalent in the P. aeruginosa biofilm matrix [80]. These findings highlight proteins’ vital role in generating and maintaining biofilms. Furthermore, large-scale transcriptome research has identified proteins involved in quorum sensing, such as Npr, PlcR, PapR, ComX, and CSF, which have been found to regulate biofilm production in various types of bacteria [81].
Forming biofilms is a complex process influenced by various factors, such as growth conditions, growth media, and the presence of specific matrix components. Recent research conducted by Sauer et al. [82] has revealed that the structure of biofilms in P. aeruginosa may vary depending on the growth circumstances and medium used. It has also been observed that bacterial biofilms can develop pillars and mushroom-like structures, indicating that biofilm architecture is diverse and intricate [83].
According to research by Hartmann et al. [84], mechanical contacts between cells are responsible for developing three-dimensional organization and structure in expanding biofilms. This process can be regulated by controlling the production of specific matrix components. Yan et al. [85] have also suggested that global mechanical instabilities play a role in biofilm structure formation, emphasizing the mechanisms’ complexity. Bacterial biofilm structure is significantly impacted by eDNA, which contributes to the formation, composition, and function of biofilms [86].
The intricate relationship between mechanical instability and the development of biofilm architecture highlights the dynamic and adaptable nature of microbial communities [85]. During the initial stages, shear stress in fluid environments is necessary to facilitate the attachment of microbial cells to surfaces. Fluctuations in fluid dynamics create a microenvironment where bacteria can either adhere effectively or encounter difficulties in making initial contact. This essential first step sets the foundation for subsequent biofilm formation [87, 88].
As biofilms progress in their development, mechanical forces play a crucial role in facilitating microbial attachment and shaping the structure of these communities. The movement of fluids can produce distinct patterns within the biofilm matrix, including streamers and pillars [82, 89]. This dynamic response underscores the biofilm’s remarkable ability to adapt and reorganize itself in response to an array of mechanical stimuli, revealing the intricate nature of biofilm architecture. It has been noted that mechanical shear stress significantly influences biofilm properties such as thickness, density, permeability, and the spatial distribution of live cells within the biofilm [90].
Mechanical cues are critical in regulating the extracellular matrix comprising polysaccharides, proteins, and eDNA. Elevated shear stresses prompt bacteria to increase their EPS production. The heightened production of biofilm matrix strengthens its stability, creating a defensive barrier that protects the microbial community from external stimuli. The control of EPS generation demonstrates the biofilm’s ability to respond and adapt to the mechanical properties of its environment [84, 91].
Additionally, varying oxygen levels are recognized as a crucial element in forming the final structure of Aspergillus fumigatus biofilms, highlighting the ever-changing process of biofilm development [83]. As A. fumigatus biofilms mature, oxygen gradients naturally occur and play a critical role in shaping their architecture. As studies have shown, these low-oxygen environments promote the growth of filamentous fungal biofilms and enhance their resistance to antifungal treatment [92]. The biofilm structure of A. fumigatus is regulated by a gene family known as biofilm architectural factor A (bafA) which is produced heterogeneously. This particular gene family affects the growth of the fungus in low oxygen conditions and the appearance of colonies, ultimately impacting its ability to form biofilm and cause disease [83].
The development and structure of biofilms are complex processes influenced by various factors, such as microbial composition, components of the EPS, and environmental conditions [86]. Biofilms are organized communities of microorganisms surrounded by an EPS, which protects cells from external and chemical threats, maintains the integrity of the biomass, and facilitates hydration and nutrient uptake [86].
Understanding biofilms’ spatial arrangement and structural changes is crucial for comprehending bacterial interactions and the collective functional abilities of biofilms that surpass those of individual cells. The formation and characteristics of biofilms are determined by the bacterial composition and the surrounding environmental conditions [17]. To gain a thorough understanding of bacterial interactions and the emergent functional capabilities of biofilms, it is essential to comprehend the spatial organization and structural dynamics of biofilms. General composition of biofilm matrix in common microbial communities are explained through Fig. 3.
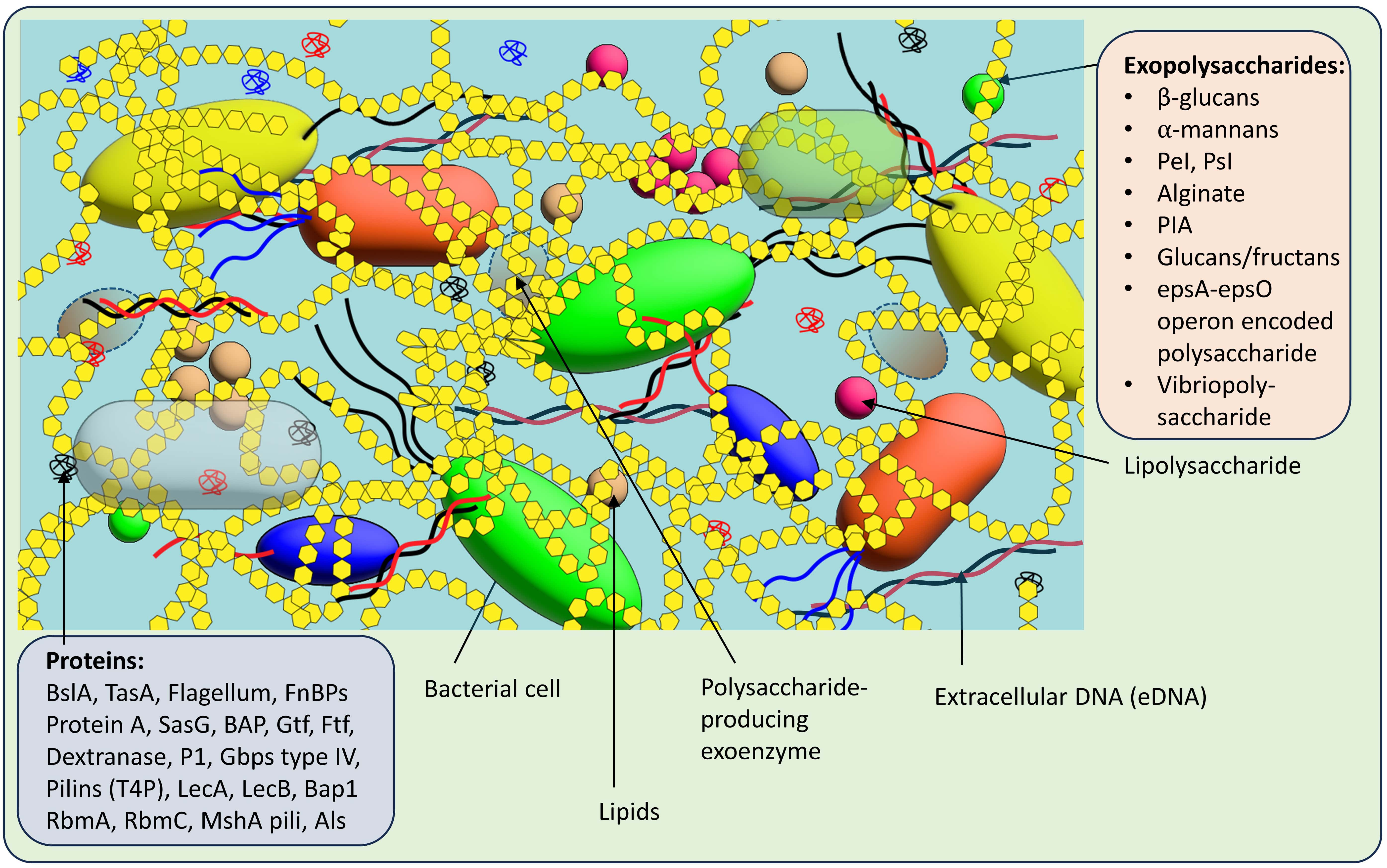 Fig. 3.
Fig. 3.General composition of biofilm matrix in common microbial communities. The biofilm can have same or different types of bacteria, embedded in different exopolysaccharides and proteins. The biofilm also contains lipids, lipopolysaccharides, eDNA and polysaccharide producing exoenzyme, etc.
The process of biofilm formation involves a series of distinct stages, namely initial adhesion, microcolony creation, maturation, and dispersion [93, 94].
Understanding the early reversible stage of biofilm formation is a crucial step toward developing effective strategies to prevent or control biofilm growth. This stage involves the reversible adhesion of bacterial cells to surfaces, which is transient and dynamic, allowing cells to detach and reattach. The temporary nature of this process is essential in establishing the early contact between bacteria and the surface, which can significantly impact the subsequent stages of biofilm development [82]. Recent research conducted by Giacomucci et al. [95] has shed light on the significance of reversible attachment in the biofilm formation process, particularly in species such as Vibrio cholerae. This knowledge can help us develop better approaches to managing biofilm formation and its associated problems [95].
Various factors influence the attachment and aggregation of bacterial cells on surfaces during the initial stages of biofilm formation. Flagella, fimbriae, and pili are among the elements that play a significant role. For instance, the single polar flagellum and Type 4 pili (T4P) are crucial for the development and maturation of biofilms in Gram-negative opportunistic pathogens like P. aeruginosa [96]. Alterations in the flagellar hook protein FlgE can impact the organization of P. aeruginosa biofilms and their ability to withstand antibiotics [97]. Additionally, the structural characteristics of the biofilm-associated ancient Chaperone-Usher pilus CupE from P. aeruginosa offer valuable insights into the function of pili in cellular adhesion and biofilm generation [98]. Research conducted on S. maltophilia revealed that a strain lacking flagellin significantly reduced swimming capability and impaired adhesion and biofilm formation [99]. Similarly, studies on Burkholderia pseudomallei have demonstrated that flagella, fimbriae, and type IV pili are crucial in promoting adhesion, aggregation, and biofilm formation [100]. These findings highlight the importance of flagella, fimbriae, and pili in the early stages of biofilm formation in different types of bacteria.
Structures are delicate in the initial phase of biofilm formation, and bacterial metabolisms are highly active. Given their vulnerability to antimicrobials, it is essential to manage and control biofilms within a specific timeframe [101]. Researchers have found that DNase can hinder the early development of biofilms in some bacterial strains, highlighting the potential for targeting genetic and molecular elements in the early stages of biofilm formation [102]. Moreover, studies have explored the impact of environmental factors such as fluid shear stress and bacterial adhesion conditions [88], as well as natural substances like eucalyptol on the early phase of biofilm formation [103]. These findings suggest that regulating these parameters can be a strategic approach to managing the creation and growth of biofilms. The reversible early phase of biofilm formation is a critical step with significant implications, and a deeper understanding of this stage can lead to more effective methods for controlling and preventing biofilms. By analyzing influential factors and exploring ways to modify these parameters, it is possible to develop more precise and efficient approaches to manage the challenges associated with biofilm.
The transformation from reversible attachment to permanent adhesion marks a crucial juncture in the development of biofilms. This shift involves a move away from a transient, loosely bound state to a more lasting, firmly established community. Surface characteristics such as roughness, charge, and stiffness play an essential role in the adhesion process and the resulting formation of biofilms [104, 105].
Exploring P. aeruginosa biofilms, particularly transitioning from reversible to irreversible attachment, yields valuable insights into potential methods for preventing biofilm formation. Targeting the reversible attachment phase could disrupt the biofilm formation process before it advances into a more mature and resistant stage [106].
The microcolony phase in developing a bacterial biofilm is a vital turning point. It marks the transition from the initial, temporary attachment of individual cells to the formation of organized, long-lasting communities that serve as the groundwork for the growth of a fully developed biofilm. This stage is known for its complex nature, which is influenced by genetic, molecular, and environmental factors.
Researchers have highlighted the crucial role of eDNA in the early stages of microcolony development. When released into the surrounding environment, eDNA acts as a trigger that influences the initial aggregation of bacterial cells. This step is vital for the spatial arrangement of the microcolony and sets the stage for eventual biofilm formation [107]. A recent study delves deeper into the complexity of this phase by using computer simulations to analyze the initial growth of bacterial biofilms. The study reveals that the interaction between diffusion and growth rates significantly impacts the creation and durability of biofilm microcolonies [108].
The formation of biofilms during the microcolony stage is affected by various environmental factors such as temperature, pH levels, and nutrient availability. Research has shown that these variables have a direct impact on the growth and aggregation behaviors of bacteria, ultimately influencing the overall structure of the biofilm [4, 109]. Furthermore, oxygen distribution within the biofilm plays a crucial role, particularly in larger microcolonies where a decrease in oxygen levels in the core can affect the biofilm’s structure and function [110]. This study of oxygen gradients within biofilms sheds light on how different microenvironmental conditions can result in varying growth patterns within a single biofilm.
Molecular and genetic factors further complicate the microcolony stage. Research highlights the importance of cyclic-di-GMP and quorum sensing in regulating different aspects of biofilm growth, including adhesion and the production of EPS, which affect microcolony formation and development [111]. Additionally, Pincott et al. [112] have shown that heat shock proteins, nitric oxide, and pH changes play a significant role in the initial attachment and subsequent formation of small bacterial colonies.
In vitro models, such as the one developed by Sherman et al. [113], provide crucial insights into the cellular ssteractions and environmental conditions that promote the formation of organized biofilm communities. These models are essential in understanding the physiological significance of microcolony formation, a crucial stage in biofilm development that involves dynamic interactions of genetic, molecular, and environmental factors. Understanding these complexities is vital for developing effective techniques to control biofilms, particularly in medical and industrial settings with significant challenges. By investigating the molecular mechanisms and environmental factors that regulate this stage, researchers can uncover innovative ways to interrupt biofilm development, leading to creative approaches to managing biofilm-related issues.
Forming biofilms is a complex and dynamic process involving the production of an EPS, which provides structural support for the three-dimensional framework of the biofilm. During this stage, columnar structures are established, with a vertical gradient of viscoelasticity influenced by hydrodynamic shear resulting from fluid flow in the surrounding environment [112]. The extent of shear stress is a critical factor in determining the physical properties of the biofilm, affecting its resilience and organization.
As per research conducted by Gupta et al. [114], the maturation phase of biofilms relies on eDNA generation and adhesin preservation to enhance stability and integrity, ensuring the survival and functionality of the microbial community within. Wei & Yang’s study [115] focused on the impact of hydrodynamic conditions and microscale surface roughness on fully-developed multilayer biofilms, particularly on shear stress and thickness. Environmental factors play a crucial role in determining the physical attributes of biofilms as they mature. A three-phase model for A. fumigatus biofilm formation was introduced, highlighting the mature stage’s resistance to antifungal treatments and its role in disease progression [116].
During the maturation and matrix generation phase, biofilm morphology undergoes a crucial transformation, resulting in the expansion of the biofilm as a three-dimensional formation [112]. This phase is responsible for the evolution of biofilms from basic cell aggregations to complex, organized communities. Water channels play a vital role in biofilms’ proper function and structural integrity. In Bacillus subtilis biofilms, wrinkles serve as pathways for water, allowing nutrients to move across different subgroups of the biofilm [43]. Similarly, Escherichia coli biofilms’ intra-colony channels act as nutrient-absorbing systems, emphasizing the channels’ vital role in sustaining the biofilm’s survival [117]. The channel networks are crucial for maintaining biofilm survival by facilitating the transportation of nutrients and chemicals [118].
Recent research has shown that external fluid flows play a significant role in biofilms’ intricate structure and three-dimensional morphology. Fluid dynamics directly impact biofilms’ growth, which can result in the formation of unique shapes such as towers and mushroom-like formations [84, 119]. Furthermore, mechanical instability and wrinkling in expanding biofilms indicate biofilm growth’s dynamic nature and further emphasize water channels’ involvement in this process [120].
Scientists have developed bioinspired artificial nanochannels that mimic the functions of natural biofilms. These nanochannels leverage the properties of water channels to achieve high permeability and selectivity [121]. Additionally, researchers have explored 3D printing technology to create structured E. coli biofilms, which could be utilized for organizing bacteria and water channels to support nutrient and oxygen exchange [122].
As cells mature, the EPS matrix plays a crucial role in enhancing their attachment and unity, forming tightly packed cell clusters and facilitating the accumulation of microbes. This, in turn, results in developing a well-organized and firmly bound biofilm [123]. The transition of biofilm models from static to dynamic systems, as explained by Sánchez et al. [124], emphasizes the importance of surface characteristics, bacterial movement, and hydrodynamic conditions in biofilm development. The intricate network that governs the various stages of the biofilm’s life cycle underscores the complex nature of biofilm maturation and its significance in multiple settings, from natural ecosystems to healthcare facilities [124].
The final stage of biofilm growth, known as dispersion, is a critical component of the biofilm life cycle. During this phase, cells actively disperse from the biofilm structure and attach themselves to other surfaces, establishing new biofilms in different locations. Recent research has shown that biofilm dispersion is a complex and intentional process involving the purposeful release of cells from the structure [125]. Additionally, studies have suggested that this process is coordinated and synchronized, with cells detaching and dispersing deliberately [126]. These findings indicate a high level of organization and control within the biofilm, essential for the survival and spread of its constituent cells.
Research has shown that nitric oxide is crucial in triggering the dispersal of P. aeruginosa biofilms, shedding light on the complex regulatory mechanisms involved. This study highlights that biofilm dispersion can be controlled through substances that hinder or promote it, thereby providing potential strategies for managing biofilm formation in diverse settings [127]. In a similar vein, Bridges et al. [128] identified unique signaling pathways, matrix-digesting enzymes, and motility components that regulate the dispersal of V. cholerae biofilms.
A recent study delved into the transition from individual planktonic cells to complex multicellular biofilms, highlighting the importance of cyclic di-GMP, quorum sensing, and bacterial proteins in the dispersion of biofilms [129]. This underscores the intricate regulatory networks involved in the life cycles of biofilms and the sophisticated communication mechanisms that govern their behavior. The study also found that berberine, when used at sub-inhibitory levels, impedes biofilm dispersal in S. aureus [130], offering potential therapeutic avenues against biofilms through the use of antibacterial drugs.
Moreover, the identification of the DNA-specific endonuclease EndA as a crucial factor in the dispersal of P. aeruginosa biofilms, as proposed by Cherny & Sauer [125], implies that breaking down the biofilm structure is vital for efficient dispersion. This knowledge is critical for devising strategies to encourage or hinder biofilm dispersion, depending on the desired outcome in medical, industrial, or environmental contexts. The dispersion stage of biofilm development is a tightly regulated and dynamic process that ensures the dissemination and survival of bacteria that form biofilms. Understanding the molecular mechanisms and environmental stimuli that govern this phenomenon is crucial for developing practical approaches to manipulating biofilms, whether by facilitating their dispersal in favorable circumstances or impeding it to mitigate the risks of disease and biofouling.
The EPS matrix is an essential component of biofilms, and studies have shown that it plays a significant role in antibiotic resistance [131]. Comprised of proteins, eDNA, and polysaccharides, the EPS matrix has a complex chemical composition that creates a barrier, hindering the diffusion of antibiotics into the biofilm [132]. This barrier reduces the speed at which antibiotics penetrate the biofilm, making it more resistant to antibiotic therapy [133, 134]. The EPS matrix not only prevents the infiltration of antibiotics but also resists phagocytosis by immune cells, further enhancing the resistance of biofilms against antibiotics [135]. The limited ability of antibiotics to permeate the biofilm matrix and the extended duration required for antibiotics to infiltrate the biofilm has been identified as contributing factors to the relationship between antibiotic resistance and biofilm formation [136].
The microenvironments created by biofilms have a notable impact on the efficacy of antibiotics. These habitats contain varying pH levels, oxygen, and nutrients that can influence how antibiotics interact with and penetrate biofilms. Altered conditions within biofilms, such as lowered pH and oxygen levels, can hinder the ability of antibiotics to enter and remain within the biofilm, ultimately promoting antibiotic resistance [137, 138, 139].
The slow growth and dormancy of bacteria that form biofilms contribute significantly to their increased resistance to antibiotics. According to Pu et al. [140], inactive bacteria within biofilms are associated with reduced metabolic and reproductive rates, making them more resistant to antibiotics. This sluggish growth hinders the ability of antibiotics to penetrate the biofilm, resulting in decreased effectiveness of antibiotic [141, 142]. Additionally, the varied control of metabolic activity and growth rate of cells within biofilms, including inactive and dead bacteria, further enhances the resistance of biofilms to antibiotics [142].
Efflux pumps play a critical role in both the development of biofilms and the resistance to antimicrobial agents. These pumps can regulate the internal conditions of biofilms, protect cells from antibiotic attacks, and contribute to multi-drug resistance [143]. Efflux pump inhibitors have been identified as a promising strategy to disrupt biofilm formation and enhance the efficacy of antibiotics, thereby reversing pathogen resistance to these drugs [144]. Understanding the functions of efflux pumps in biofilm formation can pave the way for innovative therapeutic approaches that target their activity, dismantle biofilms, and improve the treatment of infections [145]. Numerous studies have demonstrated the efficacy of efflux pump inhibitors in reducing biofilm production in a variety of bacteria, including E. coli, P. aeruginosa, S. aureus, and Klebsiella pneumoniae [145, 146]. Efflux pumps not only mediate antibiotic resistance but also contribute to biofilm development and several other physiological processes [147]. Moreover, research suggests that efflux pump inhibitors can impede biofilm production by regulating drug efflux pumps in S. saprophyticus [73].
Studies have shown that bacteria forming biofilms exhibit higher levels of antibiotic resistance due to genes associated with biofilm formation [148]. Biofilms are also related to the spread of antibiotic-resistant genes across bacterial species and the promotion of resistance transmission [149, 150]. The dense cellular population and extracellular matrix present in biofilms facilitate the genetic mutations and transfer of antibiotic-resistance genes in biofilms [151, 152]. The increased cellular density also leads to higher levels of exogenous DNA, aiding the acquisition of mobile genetic elements and the horizontal transfer of genes, which contributes significantly to antibiotic resistance dissemination [152]. In addition, the genetic and physical heterogeneity within biofilms plays a crucial role in enabling bacteria to withstand the effects of antibacterial medications [153]. B. subtilis researchers have also examined the genetic variants and gene expression levels of essential genes responsible for biofilm formation and matrix creation in ionizing-radiation-resistant bacteria [154], providing insight into the genetic factors behind biofilm development and resistance. Furthermore, bacteria capable of building biofilms possess specific genes related to biofilm formation, which enhance their resistance, and the development of biofilms is linked to a more extraordinary occurrence of genetic factors that confer resistance to quinolone antibiotics [150, 155].
Biofilms respond to stress in various ways, including shear dynamics, electrical oscillations, and metabolic and electric processes. According to Kurz et al.’s study [156], shear stress induces intermittent behavior in preferential flow channels within biofilms in porous media, indicating a non-Newtonian response. Electrical oscillations in biofilms may help them resist light-induced stress [157]. Meanwhile, Martinez-Corral et al.’s research [158] reveals that biofilms react to nutrient-limitation-driven stress from the core to the periphery through metabolic and electric processes.
Furthermore, research has shown that environmental stress can cause existing biofilms to restructure, highlighting the significant impact of stress on the arrangement of biofilm structures [159]. Stress reactions play a crucial role in developing biofilms and acquiring antibiotic resistance in pathogenic bacteria. The c-di-GMP molecule regulates various aspects of biofilm formation, including resistance to antimicrobial medicines and other stress-induced reactions [160]. The extracellular matrix and changes in the physiological state of cells within the biofilm help biofilms withstand stress [161]. Moreover, genes associated with biofilm development are activated in response to stress, further supporting their contribution to stress resistance [162].
Persister cells are essential in conferring resistance to antimicrobial agents in bacterial biofilms. Persister cells have been discovered in different types of bacteria, including Mycobacteria, Borrelia, S. epidermidis, P. aeruginosa, Acinetobacter baumannii, and C. albicans [110, 163, 164]. Persister cells inside biofilms trigger toxin/antitoxin systems, suppressing protein synthesis, inducing dormancy, and developing antibiotic tolerance [164]. In addition, studies have demonstrated that the creation of biofilms is associated with a significant increase in resistance to antifungal treatments and the facilitation of the development of persister cells that are tolerant to antifungal agents [165]. Furthermore, the prolonged survival of the biofilm community when exposed to antibiotics enables the development of persister cells through mechanisms associated with inherent protection against antibiotic-induced harm [166]. Biofilm persister cells have been proposed to exhibit resistance to antibiotics up to 1000 times higher than the most minor inhibitory concentrations [167]. Persister cells are formed within biofilms to increase the tolerance of biofilm cells. These cells are a tiny subset called “transiently resistant” cells linked to biofilms’ unique growth pattern [168].
The development of bacterial biofilms on implantable medical devices presents complex challenges in the healthcare industry. Biofilm on implants increases vulnerability to infections and illnesses, which can be long-lasting and ongoing compared to infections caused by freely moving bacteria [169]. This results in extended patient suffering, increased healthcare costs, and potential complications. Antibiotic resistance is becoming a growing concern in the creation of biofilms [170]. The protective extracellular matrix of biofilms acts as a barrier, making it difficult for antimicrobial drugs to penetrate and efficiently target the bacterial community. As a result, treating biofilm-associated illnesses can be very difficult [171].
Another significant concern is the impact on the functionality of implantable medical devices. Biofilms can compromise the optimal performance of medical equipment, such as catheters, prosthetic joints, or cardiac implants, ultimately leading to device malfunction or failure [170, 172]. This not only affects the well-being of patients but also necessitates additional medical interventions, such as the removal and replacement of faulty equipment [173].
The presence of biofilm can trigger chronic inflammation, wherein the body’s immune response to the persistent infection can lead to prolonged inflammation and tissue damage, worsening the overall impact of the biofilm-related illness. This persistent inflammatory condition poses a challenge in inpatient treatment, requiring a sophisticated strategy for infection prevention and inflammation control [174, 175, 176].
Furthermore, accurately detecting illnesses related to biofilms presents its challenges. Traditional diagnostic techniques may not be precise enough to identify biofilms, which can lead to delayed or insufficient treatmen [177, 178]. Therefore, it is necessary to adopt advanced diagnostic methods to facilitate early detection and prompt intervention in biofilm growth on implants.
To address these challenges, recent advancements in polymer coatings have shown promise in combating bacterial adhesion and biofilm formation on medical equipment [179]. A viable approach to preventing the formation of biofilms on medical devices is through specialized coatings that deter bacteria from adhering to the surface. Antifouling and bioinspired self-adhesion coatings, for instance, have successfully inhibited bacterial attachment on surfaces [170, 180].
Crystal violet staining has become a fundamental technique in biofilm research, recognized for its effectiveness in measuring biofilm biomass and evaluating biofilm formation. Its versatility has been demonstrated in various investigations, such as the classification of Stenotrophomonas maltophilia strains based on their biofilm production ability [181] and the quantification of biofilm biomass in high-throughput workflows [182].
Crystal violet staining is also applied to examine the effects of biofilm removal, including the quantification of biomolecules in bacterial cells and extracellular polymeric compounds [183, 184] and the evaluation of the inhibitory impacts on biofilm formation [185, 186].
In addition to these applications, crystal violet staining has also been crucial in other scientific fields. For instance, one study utilized it to evaluate biofilm formation capacity in canine cystitis [187]. Crystal violet staining has proven to be a valuable technique in studying Pseudomonas syringae and assessing biofilm biomass [188]. The method has also played a crucial role in evaluating the impact of various substances on biofilm formation, including silver nanoparticles [189], propolis [190], and norspermidine [191]. The adaptability and dependability of this method in providing both quantitative and qualitative information about biofilm biomass and formation make it an essential tool in biofilm research. Its extensive utilization in diverse settings, from examining bacterial varieties to testing antibiotic substances, underscores its fundamental contribution to advancing our knowledge about biofilms.
Scientists have extensively researched ConA, a protein that binds to carbohydrates, for its ability to visualize biofilms. It is well-known for its strong attraction to mannose and glucose molecules [192]. When used in conjunction with other fluorescent stains in Confocal Laser Scanning Microscopy (CLSM), ConA has been proven effective in examining live bacterial biofilms and observing the structural elements of the biofilm matrix [193, 194]. Moreover, researchers have successfully utilized ConA to modify mesoporous silicon-based biosensors for label-free optical detection of bacteria in real-time mode [195]. In addition, ConA has been identified as a critical element in developing nanosystems for treating infections, underscoring its crucial role in biofilm research [196].
SYTO 9 and propidium iodide (PI) use in biofilm research has been extensively documented in scientific literature. The LIVE/DEAD BacLight™ Bacterial Viability Kit frequently employs these fluorescent dyes to differentiate between live and dead cells by assessing the integrity of their membranes [197, 198, 199]. SYTO 9 and PI staining have been utilized in a variety of research endeavors, including investigating biofilm production in different strains of A. baumannii [199], evaluating the antibacterial and antibiofilm activities against S. mutans [200], and assessing the impacts of antiseptic agents on S. aureus biofilm [201]. Additionally, SYTO 9 and PI staining have been used to examine the implications of eugenol, trans-cinnamaldehyde, citronellol, and terpineol on the management of E. coli biofilm [202]. The SYTO 9 and PI combination has also been used to determine the viability of S. mutans biofilm [203], evaluate the effectiveness of antiseptic agents against staphylococcal biofilm [204], and assess the bacterial viability of chlorine- and quaternary ammonium compounds-treated Lactobacillus cells [205]. However, it is worth noting that PI staining, despite its widespread use, has been found to underestimate the vitality of adhering bacterial cells [206]. Fig. 4 described different methods of staining biofilms.
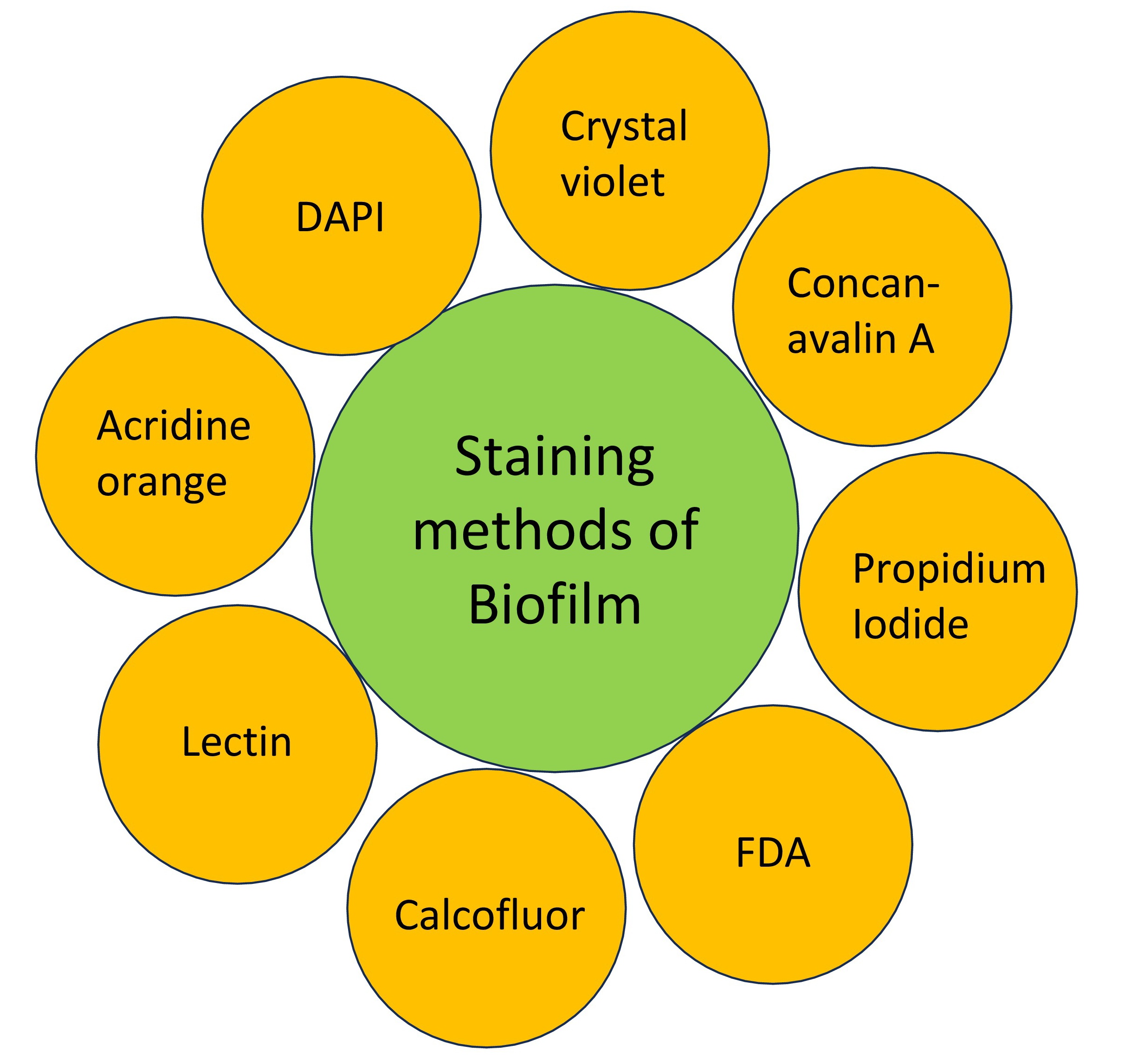 Fig. 4.
Fig. 4.Different methods of staining biofilms. The different stains include lectin staining, Acridine organge staining, Calcofluor staining, DAPI staining, fluorescein diacetate staining, crystal violet staining, Concanavalin A staining, and Propidium iodide staining.
DAPI staining, which uses 4
Acridine orange staining is a widely accepted process for visualizing and measuring biofilms. Its application spans multiple studies, such as one conducted by Hamdoon et al et al. [214], which analyzed biofilm formation on different orthodontic retainer materials. In another study, Hrynchuk et al. [215] utilized acridine orange fluorescence staining to assess the survival of bacterial cells within fully formed biofilms of S. aureus, while investigating the effects of an adamantane derivative. Acridine orange staining is a crucial tool for investigating biofilm kinetics and composition. Monmeyran et al. [216] explored the efficacy of the inducible chemical-genetic fluorescent marker FAST to monitor bacterial biofilm dynamics, in contrast to classical fluorescent proteins. Schiessl et al. [217] highlighted the importance of understanding biofilm metabolism and its impact on antibiotic resistance. Their findings demonstrated the contribution of phenazine synthesis to antibiotic tolerance and metabolic variability in P. aeruginosa biofilms [217]. These results suggest that integrating acridine orange staining, fluorescent markers, and metabolic profiling can provide valuable insights into the intricate mechanisms underlying bacterial biofilms and their antibiotic resistance.
Researchers have discovered that lectins can be practical tools for studying the composition of biofilm matrices, as they selectively bind to extracellular matrices [218]. Lectins have been used to identify polysaccharides in biofilms of P. aeruginosa [219], and to visualize and analyze carbohydrate-containing EPS in biofilms of Sulfolobus acidocaldarius [220]. They have also been employed to investigate changes in biofilm composition by labeling extracellular proteins and polysaccharides, indicating that lectins can potentially explore the dynamics of biofilm matrices [221].
However, it is vital to recognize the limitations of lectin staining. A study by Domingue et al. [222] cautions that while lectins can label biofilms formed under non-living conditions, they cannot distinguish between carbohydrates from microbes and those from humans. This limitation makes it difficult to accurately determine the molecular source of a biofilm matrix in living organisms. As macromolecules, lectins have limited ability to penetrate biofilms, which could impact their effectiveness in certain situations [223]. It is crucial to consider these limitations when interpreting results obtained from lectin staining of biofilms.
Numerous papers have extensively reported on using calcofluor white stain in
biofilm research. This fluorochrome specifically attaches to
The use of Fluorescein diacetate (FDA) staining is a well-established technique in assessing microbial activity and the formation of biofilms. Through FDA staining, researchers can observe biofilm bacteria attached to surfaces and quantify their activity level [230]. The adaptability of this staining method is demonstrated by its application in investigating the biofilm formation of pathogens, including P. aeruginosa and S. mutans [231, 232, 233]. Moreover, using FDA staining alongside other dyes for dual viability staining has increased its effectiveness in evaluating the viability of microorganisms [234]. The staining technique has also been refined to facilitate a quantitative assessment of biofilm formation, highlighting its significance in measuring the extent of biofilm growth [235].
Recent studies have shown a growing interest in using peptide nucleic acid (PNA) probes for investigating biofilms. PNA fluorescence in situ hybridization (FISH) has proven effective in differentiating bacterial populations within biofilms, providing insights into their spatial arrangement and metabolic processes [236, 237]. This technique offers a more comprehensive understanding of biofilm structure and functional progression [238, 239]. PNA probes have also been utilized to quantify cystic fibrosis multispecies biofilms, demonstrating their capacity to evaluate specific populations within complex biofilms of multiple microbial species [240]. Moreover, PNA-FISH has been successfully employed to examine biofilms in diverse settings, including septic arthritis models and wound care, highlighting its versatility across various research domains [241, 242, 243]. PNA probes exhibit a strong affinity for complementary nucleic acids and boast biological stability due to their lack of charge and peptide bond-linked backbone [239], making them highly useful for investigating biofilms. PNA-FISH probes have been specifically designed to detect pathogens such as Gardnerella vaginalis [244] and Atopobium vaginae [245] in the context of bacterial vaginosis. These probes confirm the presence and impact of these bacteria within biofilms. Biofilm stains and their applications are presented in Table 1 (Ref. [187, 188, 192, 196, 207, 215, 217, 218, 219, 227, 228, 230, 231, 237, 240, 246, 247, 248]).
| Stain | Description | Examples | References |
| Crystal Violet | A primary dye that uniformly dyes the entire biofilm. Frequently employed in tests for quantifying biomass. | Measuring the amount of living matter in Staphylococcus aureus biofilms; Evaluating the growth of biofilms on medical devices such as catheters. | [187, 188] |
| Concanavalin A [ConA] | ConA, which has been tagged with a fluorescent marker, attaches to glucose and mannose molecules present in the EPS matrix. | Examining the EPS matrix in biofilms. | [192, 196] |
| SYTO 9 and Propidium Iodide | Employed in conjunction to distinguish between viable and non-viable cells inside the biofilm. | Evaluating the feasibility of mixed-species biofilms after treatment; Investigating the impact of antibiotics on the survival of biofilms. | [246, 247] |
| DAPI (4’,6-diamidino-2-phenylindole) | A fluorescent dye that specifically attaches to DNA, employed to stain the nuclei of cells present in biofilms. | Applying dye to EPS biofilms; Examining the arrangement of bacteria in environmental biofilms. | [207, 248] |
| Acridine Orange | Attaches to DNA, RNA, and acidic polysaccharides, resulting in the coloring of both viable and non-viable cells. | Distinguishing between viable and non-viable bacteria within a biofilm; Analyzing the cellular makeup of biofilms in industrial water systems. | [215, 217] |
| Lectins | Fluorescently labeled lectins stain specific polysaccharides in the EPS matrix. | Determining the particular polysaccharides present in biofilms; valuable for viewing the composition of the biofilm matrix. | [218, 219] |
| Calcofluor White | A cellulose- and chitin-specific fluorescent dye employed as a stain for fungus or polysaccharide matrices. | Observing the EPS of biofilms, enabling the targeted staining of polysaccharides present in the biofilm structure. | [227, 228] |
| Fluorescein Diacetate | Used for assessing the metabolic activity of cells in biofilms. | Evaluating microbial activity and the production of biofilms; evaluating the viability of microorganisms. | [230, 231] |
| Peptide Nucleic Acid (PNA) Probes | Used for FISH to identify specific bacterial species within biofilms. | Characterizing distinct bacterial species within heterogeneous biofilms; Identifying pathogenic bacteria present in biofilms linked to persistent cystic fibrosis. | [237, 240] |
Confocal Laser Scanning Microscopy (CLSM) is an essential technology for observing and investigating biofilms with precision. Researchers have shown that this technique effectively uncovers the complex structure, spatial patterns, and bacterial quantities in mixed-species biofilms [249]. CLSM is applied to evaluate the efficacy of chemical treatments in combating biofilms, with a specific focus on eliminating bacteria and removing biofilms [250].
Additionally, it is crucial in overseeing the early stages of biofilm formation, such as attaching to and colonizing minerals [251]. CLSM has been extensively used in numerous studies to examine biofilm structures. For example, it has been used to evaluate the effectiveness of marine coatings in preventing biofilm formation [252], validate the layer-by-layer biofilm elimination theory [253] and investigate the variability in biofilm formation among different microorganisms [254].
In biofilm research, CLSM has proven to be indispensable in evaluating the effects of various therapies on biofilm production. For instance, probiotics have been studied for their potential to prevent biofilm formation [255], while the impacts of antimicrobial drugs on biofilms [256] and the degradation of biofilm structures by certain chemicals [257] have also been investigated. Furthermore, CLSM has been instrumental in examining the structural modifications of biofilms, particularly those treated with disinfectants like sodium hypochlorite [258, 259].
Moreover, CLSM has proven to be effective in monitoring biofilms grown on different metallic surfaces, providing non-intrusive three-dimensional imaging capabilities essential for understanding microbial biofilms’ complex characteristics [260]. Given its ability to provide comprehensive insight into the formation, structure, and response to treatments of biofilms, CLSM is a critical tool in this field. The following section outlines various biofilm detection methods, which are also illustrated in Fig. 5.
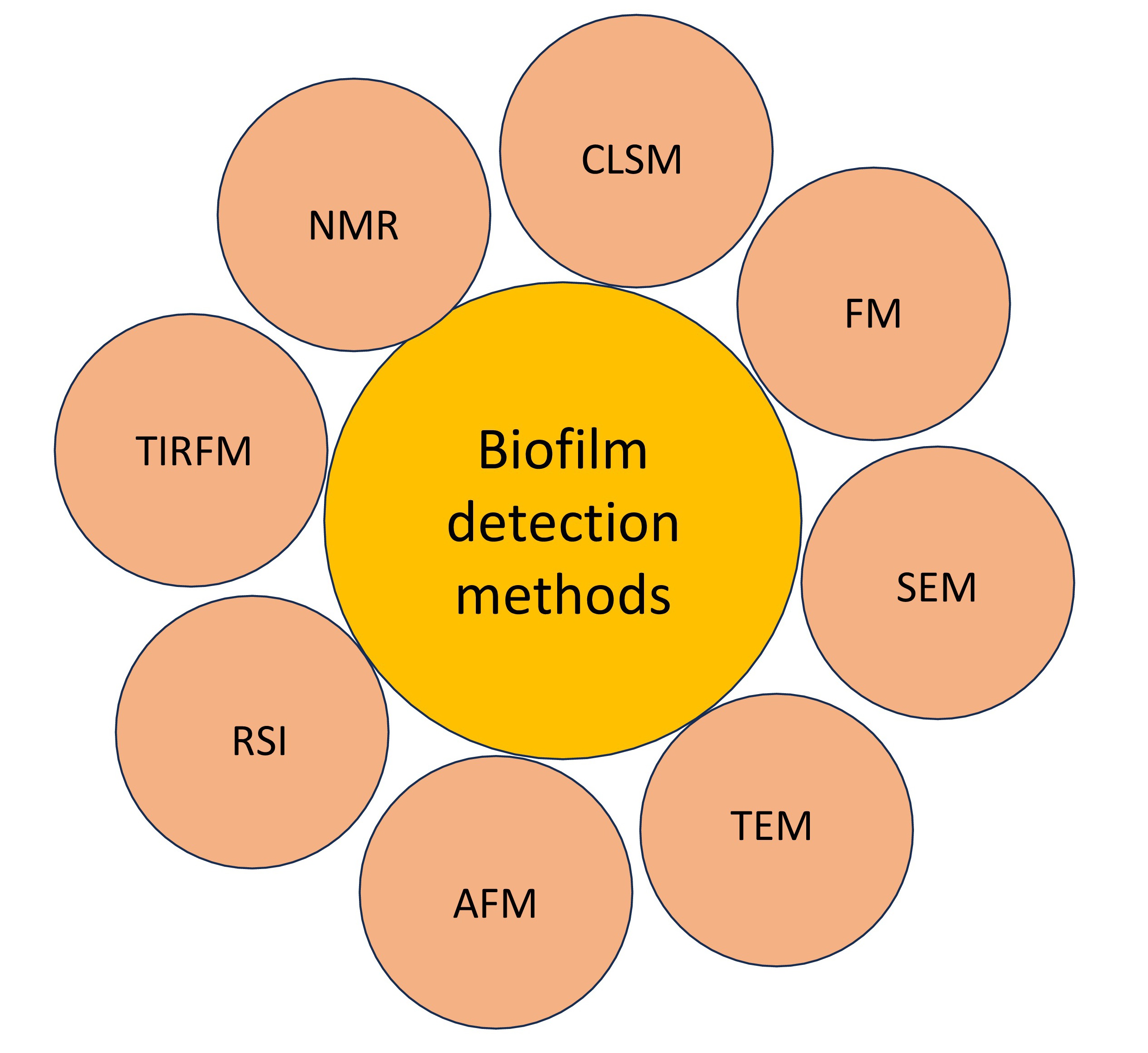 Fig. 5.
Fig. 5.Different biofilm detection methods. Abbreviations: CLSM, confocal laser scanning microscopy; FM, fluorescence microscopy; SEM, scanning electron microscopy; TEM, tunneling electron microscopy; AFM, atomic force microscopy; RSI, Raman spectroscopy and imaging; TIRFM, total internal reflection fluorescence microscopy; OCT, optical coherence tomography; NMR, nuclear mangnetic resonance.
Fluorescence microscopy is a vital tool for studying biofilms as it offers a precise 3D view of their arrangement and movement [261]. It is beneficial for tracking the development of biofilms from a single cell to a complex community [262]. However, identifying authentic biofilm signals from mineral reflection and autofluorescence in geological samples can be challenging, which is an essential factor to consider in these settings [261]. Moreover, fluorescence microscopy may have difficulty visualizing individual cells within dense biofilms due to light penetration constraints [261].
Fluorescence microscopy provides high-resolution, 3D imaging of biofilm structures, allowing for the examination of their composition and structure in both quality and quantity [263]. It can also identify regional differences in biofilm characteristics and genetic makeup [264, 265, 266]. Understanding the complexity of biofilms and evaluating measures to combat them, such as analyzing the efficacy of biofilm reduction approaches using fluorescence microscopy, is critical for significant progress in this field [267, 268].
Scanning electron microscopy (SEM) is an indispensable tool in biofilm research, providing highly detailed imagery that reveals biofilms’ intricate architecture and composition [269]. With its remarkable ability to magnify images to the nanoscale, SEM enables an exhaustive analysis of biofilm topography [270, 271]. The applications of this technique in biofilm research are diverse, encompassing the assessment of biofilm formation [272], the characterization of ultrastructure [273], the analysis of architectural alterations [271], and the imaging of structural features [274]. In addition, SEM plays a critical role in illustrating the effects of various treatments on biofilms [247, 275], investigating the mechanisms of biofilm destruction [183], and examining the components and growth patterns of biofilms [276]. By shedding light on the intricate architecture of bacteria and EPS, SEM emphasizes the importance of preserving the complex structures of biofilms during analysis [277].
Transmission Electron Microscopy (TEM) is an essential method for observing the
intricate structure of biofilms at a high resolution. The applications of biofilm
research are vast, assessing various agents’ effects on reducing biofilms [278]
and identifying amyloid-
Furthermore, TEM has played a vital role in evaluating the microscopic harm caused by antimicrobial peptides [283], investigating the impact of propolis on bacteria [284], and characterizing the structural features of dental plaque [285]. Additionally, it has provided valuable information on the release of cytoplasmic molecules from biofilm cells [286], and the influence of cholic-acid-derived amphiphiles on fungal biofilms [287].
Moreover, the efficacy of TEM is further enhanced when combined with other imaging techniques. It has been combined with SEM [284, 288], CLSM [289], and atomic force microscopy (AFM) [290] to examine the ultrastructure of biofilms thoroughly. It has also been used to track the evolution of biofilm properties over time [291] and evaluate the mechanical characteristics of bacterial biofilms [292].
Atomic Force Microscopy (AFM) is crucial in biofilm research, particularly in understanding their reactions to different treatments. For instance, Ebadi et al. [293] employed AFM to illustrate the eradication of biofilms after exposure to zinc oxide nanoparticles. Similarly, El-Baz et al. (2021) [294] used AFM to demonstrate the impact of plant essential oil exposure on biofilm formation, as indicated by changes in cell adhesion and surface roughness. Additionally, Đukanović et al. [295] utilized AFM to evaluate the antibiofilm efficacy of Frangula emodin against S. aureus biofilms, providing comprehensive observations on their structure and form. These studies underscore the importance of AFM in evaluating biofilm reactions to various treatments.
Moreover, AFM has proved essential in uncovering structural changes in bacterial cells and decreases in the rigidity and ability to stick together of biofilms treated with ceragenins [296, 297]. Handorf et al. [298] also utilized AFM to investigate the effects of plasma-treated water on Listeria monocytogenes biofilms, observing significant modifications in the structural integrity of the bacterial biofilm.
In addition, AFM has been used to analyze the formation of biofilms on various surfaces such as glass, polystyrene, steel, ceramic, and rubber [299]. This study provided significant knowledge regarding the ability of marine bacteria to produce biofilms. Furthermore, it was used to evaluate the mass and morphology of biofilms formed on various surfaces, providing insight into the deterioration of materials resulting from biofilm formation [300]. The wide range of applications highlights the flexibility and significance of AFM in studying biofilms.
Raman spectroscopy is a valuable and non-invasive approach to thoroughly examining biofilms. Its ability to decipher biofilms’ intricate makeup and structure makes it an ideal method for analysis. This technique is instrumental in studying crucial components of biofilm matrix, like exopolysaccharides and minerals. Additionally, it can accurately identify specific organic compounds present in biofilms [289, 301]. Raman spectroscopy has proven to be highly effective in exploring the formation and organization of minerals in biofilms, especially in the detection of crystalline calcium carbonate and calcite in P. aeruginosa biofilms [22]. The technique also exhibits remarkable proficiency in identifying precise chemical interactions and monitoring dynamic changes in biofilm composition over time, as demonstrated in Vibrio parahaemolyticus’s investigation of biofilm formation [302].
The integration of Raman spectroscopy and CLSM has proven to be a powerful combination, expanding the capabilities of both methods. This has allowed for a more comprehensive examination of the structure and distribution of biofilm architecture and matrix localization [22]. Recent dental research has demonstrated the effectiveness of this approach in identifying bacterial populations in oral subgingival biofilm models, revealing its potential for analyzing complex, multi-species biofilms [303]. Furthermore, its use in clinical settings, such as evaluating S. aureus clinical isolates’ crystal violet biofilm test outcomes, highlights its significant contribution to biofilm analysis in medical contexts [304].
The technique of Total Internal Reflection Fluorescence Microscopy (TIRFM) has proven highly effective in biofilm research due to its ability to produce high-resolution images of biofilm dynamics and structures. Its adoption in this field has dramatically enhanced our understanding of biofilm formation and activity, enabling us to track the development of biofilms from a single initial cell to a fully formed, three-dimensional community with precision [262]. This level of detail is essential for comprehending the intricate mechanisms involved in biofilm formation.
TIRFM is also adaptable for both qualitative and quantitative evaluations of biofilms. It has been used to assess essential properties, including bacterial biomass, surface coverage, and the composition of EPS, providing comprehensive insights into the composition and structure of biofilms [267]. This underscores its crucial role in understanding their resilience and behavior.
As one of the most widely used fluorescence microscopy techniques in biofilm research, TIRFM is particularly popular for assessing the spatial arrangement of biofilms, including the detailed observation of cell dispersion within the biofilm matrix [305]. Its unique features make it an indispensable instrument for scientists exploring the tiny realm of biofilms, providing a glimpse into their intricate and ever-changing characteristics.
Optical Coherence Tomography (OCT) has become an essential tool in biofilm research thanks to its non-invasive approach and ability to provide immediate imaging while maintaining the integrity of the biofilm structure [250, 306]. OCT’s ability to assess biofilm structure organization by measuring their height and visualizing their intricate features makes it a trustworthy imaging method for investigating biofilms [306]. This method’s far-reaching applications include analyzing biofilm thickness and structure in marine ecosystems and bioelectrochemical systems [307, 308]. OCT has proven successful in measuring biofilm growth on various surfaces, such as anodes and chitosan-based materials in marine environments [307, 309]. It has also played a crucial role in tracking how biofilms respond to antibiotic treatments and their removal from dentures [310, 311].
OCT has sparked creative advancements in modeling methods that utilize OCT biofilm images to generate maps of non-Newtonian viscosity. Progress in this field has opened up opportunities to gain deeper insights into the mechanical characteristics of biofilms [312, 313]. Moreover, OCT’s ability to explain the impact of divalent ions and polyphosphate on the composition, structure, and stiffness of simulated drinking water biofilms highlights its crucial role in environmental and water research [15]. The numerous applications showcased here demonstrate the extensive capabilities of OCT in biofilm research and other scientific inquiries.
Nuclear Magnetic Resonance (NMR) microscopy has become a popular method for exploring biofilms, providing valuable insights into their structure and metabolism [314]. This technique is especially effective for studying biofilms in porous settings and flow cells, enabling researchers to examine water dynamics and biofilm growth across various time and length scales [315]. Furthermore, NMR spectroscopy’s ability to analyze small-molecule metabolites within and outside live cells has played a critical role in understanding biochemical processes [316].
The versatility of NMR is further demonstrated by its use in analyzing the makeup of biofilms produced by different microbes. For example, it has been successfully applied to biofilms of E. coli, V. cholerae, and A. fumigatus [317]. NMR’s efficacy in metabolomics has garnered significant attention for its potential to diagnose and identify new targets for preventing and managing biofilm-related illnesses, particularly those caused by P. aeruginosa [318]. The ability of NMR to accurately detect and measure crucial biofilm compounds significantly enhances our understanding of biofilm metabolism [314]. In addition, NMR has been used to assess the impact of contrast agents on biofilms, particularly in terms of their visibility in porous substrates [319]. The wide range of NMR microscopy applications highlights its significant contributions to biofilm research, encompassing metabolic analysis and advanced structural imaging. This underscores its importance in both scientific inquiry and practical implementation for biofilm control and treatment. Moreover, advanced imaging techniques for studying biofilms are presented in Table 2 (Ref. [223, 260, 267, 270, 278, 280, 289, 295, 307, 311, 314, 315, 320, 321, 322, 323, 324, 325, 326]).
| Imaging Technique | Description | Examples | References |
| Confocal Laser Scanning Microscopy (CLSM) | 3D imaging by laser scanning and fluorescence capture. | Visualizing microbial community structure in environmental biofilms. | [223, 260, 320] |
| Fluorescence Microscopy | Uses fluorescent dyes or proteins for visualization. | Studying gene in biofilms using fluorescent reporter genes. | [321, 322] |
| Scanning Electron Microscopy (SEM) | High-resolution imaging of biofilm surfaces. | Observing the surface architecture of dental plaque biofilms. | [270, 323] |
| Transmission Electron Microscopy (TEM) | Ultrastructure imaging of microbial cells in biofilms. | Analyzing cell-to-cell interactions within bacterial biofilms. | [278, 280] |
| Atomic Force Microscopy (AFM) | Studying physical properties like surface topography. | Measuring mechanical properties of biofilms under different environmental conditions. | [295, 324] |
| Raman Spectroscopy and Imaging | Chemical composition analysis without dyes or labels. | Chemical mapping of antibiotic penetration in biofilms. | [289, 325] |
| Total Internal Reflection Fluorescence Microscopy (TIRFM) | High-res imaging of biofilm interface. | Studying the initial stages of biofilm formation on surfaces. | [267, 326] |
| Optical Coherence Tomography (OCT) | Non-invasive real-time imaging of thicker biofilms. | Monitoring biofilm thickness and structure in wastewater treatment systems. | [307, 311] |
| Nuclear Magnetic Resonance (NMR) Microscopy | Internal structure and water content analysis. | Studying water channels and transport mechanisms in biofilms. | [314, 315] |
The study of DNA sequencing and metagenomics have proven to be effective methods in exploring biofilms, revealing valuable information on the microorganisms present, their potential functions, and how the community evolve [327]. Metagenomic sequencing has played a crucial role in uncovering new biosynthetic gene clusters, CRISPR-Cas systems, and the microbial makeup of biofilms, providing researchers with a wealth of further information [327]. In addition, metagenomic methods have been employed to investigate the clinical significance of polymicrobial biofilms, yielding helpful insights into potential methods for eliminating them [328]. Additionally, metagenomic research has been utilized to explain the microbial mechanisms and functional genes linked to anti-biofilm activities in marine settings, offering valuable insights into biofouling and biofilm resistance [329].
Metagenomic sequencing has emerged as a crucial tool for uncovering the remarkable microbial diversity, specific interactions, and community succession within biofilm ecosystems. As a result, our comprehension of biofilm development and the ecological interconnections between organisms has become more comprehensive [330, 331]. This research highlights the unique nature of various stages in biofilm development and the convergence of sequential pathways, providing invaluable insights into the adaptive strategies of biofilm communities.
Metagenomic sequencing has transformed biofilm research, comprehensively understanding microbial diversity and how it responds to environmental changes. This approach has shed light on biofilm growth and maturation mechanisms, monitoring gene expression, and temporal metabolic processes [332, 333, 334].
In particular, high-throughput DNA sequencing has been instrumental in studying the structural and functional characteristics of biofilms associated with microplastics, as well as revealing the significant functional diversity among microbial populations in wastewater treatment plants [335, 336]. These findings have provided valuable insights into the ecological roles of specific microbial groups within biofilms and the environmental consequences of biofilm formation.
Understanding the genetic control of biofilm formation in bacterial species requires transcriptomic analysis. Dubois et al. [224] utilized this method and discovered that the genes codY, ccpA, and spo0A were upregulated in C. difficile biofilms, indicating their involvement in biofilm development. Similarly, Penesyan et al. [337] found that the AdeABC efflux pump was upregulated in biofilm samples treated with ciprofloxacin, highlighting the rapid microevolution of biofilm cells in response to antibiotics. Transcriptomic analysis also revealed the relBE toxin-antitoxin system as a crucial regulator of biofilm formation in P. aeruginosa, suggesting its potential as a target for combating biofilms [338].
Nassar et al. [339] performed a transcriptome investigation on S. aureus biofilms and identified consistently elevated genes essential for biofilm formation at various stages of maturity. Finally, Shenkutie et al. [340] conducted a comparative transcriptome study on A. baumannii biofilms treated with sub-minimum inhibitory doses of imipenem and colistin, shedding light on the expression of antibiotic resistance and virulence genes specific to biofilms.
RNA sequencing, also called RNA-seq, has become the favored method for studying gene expression patterns in biofilms. It is a cost-effective option that enables the quick analysis of numerous samples [341]. This innovative methodology has yielded significant knowledge on gene expression and functional patterns in biofilm samples, examining biological mechanisms in free-floating bacteria and biofilms [342]. Metatranscriptome analysis is an essential approach for investigating gene expression and metabolic activities in microbial communities, providing valuable insights into the functional activity of a microbiome [343]. In a recent study, Radzieta et al. [344] utilized RNA sequencing to identify the primary microbe responsible for diabetes-related foot osteomyelitis and examined the microorganism’s activity level in functions linked to disease development and biofilm formation pathways. Additionally, metatranscriptomics has been used to investigate clinically significant biofilms, specifically examining gene expression in dental biofilms linked to periodontitis [345].
Metatranscriptomics offers several advantages compared to DNA-based amplicon sequencing. It selectively captures living organisms and is less prone to amplification biases, making it a valuable tool for identifying microbial species and subspecies in mixed communities [346]. Furthermore, metatranscriptomic analysis provides insights into the species actively engaged in metabolic processes within a community, whether in a healthy or diseased state. This level of information cannot be obtained using 16S ribosomal RNA profiling [347]. Additionally, metatranscriptomics has been employed to monitor biofilm-associated effectors in biofilms related to dairy products, showcasing its capacity to enhance food safety by elucidating active regulatory mechanisms [348].
Numerous investigations have been conducted to scrutinize protein manifestation in various categories of biofilms to comprehend their proteome makeup. Yung et al. [349] employed exometabolomics and proteomics techniques to discover reverse diauxie in P. aeruginosa biofilm. Meanwhile, Isa et al. [350] found that the interaction between C. albicans and P. aeruginosa biofilm led to proteome alterations in both organisms. Pereira et al. [309] discovered that the concentration of proteins in the EPS of biofilms was greater in intermittently operated bioanodes than in continuously operated bioanodes, further emphasizing the ever-changing characteristics of biofilm proteomics and the impact of external stimuli on protein expression.
Proteomic analysis has also been employed to compare strains of different bacteria that can produce biofilms with those that cannot. Sá et al. [351] presented the initial comparative proteomic analysis of strains of Corynebacterium pseudotuberculosis isolated from goats, distinguishing between those capable of generating biofilms and those that cannot. Similarly, Lawal et al. [352] discovered that S. saprophyticus from clinical and environmental sources have different biofilm compositions. These findings suggest that variations in proteins play a significant influence in the production of biofilms.
In addition, the study of proteomics has provided valuable insights into the mechanisms behind biofilm resistance and inhibition. For example, Piras et al. [353] identified 14 proteins expressed differently in the biofilm and planktonic forms of S. aureus and linked the production of these proteins in biofilms to antibiotic resistance. Similarly, Akbari et al. [354] demonstrated that the extracellular biofilm matrix acts as a physical barrier that impedes the penetration of antibiotics into the biofilms, thereby hindering their efficacy.
The scientific study of metabolomics, which involves a detailed analysis of the tiny molecules involved in biological processes, has proven to be a powerful tool for understanding the formation and metabolism of biofilms. Numerous studies have utilized metabolomics to investigate the metabolic changes during biofilm production. For instance, Pisithkul et al. [355] conducted a study that found a strong correlation between metabolomic, transcriptomic, and proteomic measures, suggesting that the changes in metabolism during biofilm growth are primarily regulated at the transcriptional level. Additionally, Salvatore et al. [356] used Partial Least Squares Discriminant Analysis (PLS-DA) on GC-MS metabolomic data to differentiate between planktonic cultures, single-species biofilms, and dual-species biofilms. They achieved this by analyzing the types and quantities of various primary and secondary metabolites released into the culture medium [356].
Metabolomics has been utilized to investigate the impact of various therapies on biofilm metabolism. One study examined the antifungal micafungin’s effects on P. aeruginosa’s metabolome and biofilm development using a comprehensive mass spectrometry-based metabolomics method. The findings indicated micafungin’s potential as a quorum-sensing inhibitor [193]. Additionally, Leggett et al. [357] employed an untargeted NMR-based metabolomics approach to compare the metabolic activities of suspended P. aeruginosa cultures and biofilms. Results showed distinct metabolic differences between the two phenotypes.
Furthermore, metabolomics has played a fundamental role in identifying crucial metabolites and metabolic pathways associated with biofilm development. For instance, researchers used a technique called 13C tracing to study the metabolic processes of bacteria as they grew in biofilms. The results provided insights into maintaining microbial fluxomics and how resources are distributed for biomass increase during biofilm development [358].
The study of lipid profiling in biofilms has become a crucial area of research, shedding light on the composition and significance of lipids in biofilm development, drug resistance, and microbial adaptation. Key constituents of EPS in mycobacterial biofilms, such as keto-mycolic acids and polysaccharides, have emphasized the importance of lipids in the construction and stability of biofilms [359]. Additionally, research on C. albicans biofilms has revealed variations in phospholipid, sphingolipid, and sterol species depending on the growth phase, highlighting the ever-changing nature of lipid composition during biofilm development [360]. Furthermore, changes in lipid metabolism have been linked to the emergence of antibiotic tolerance in bacterial biofilms. Thickening of the cell membrane due to lipid metabolism activation hinders the penetration of antibiotics, resulting in low drug concentrations within the interior of the biofilm [361].
Lipidomic profiling has been utilized to examine the lipid compositions of dental plaque samples and in vitro biofilms, providing valuable insight into the lipid profiles of biofilm matrices [362]. In addition, lipid alterations have been identified in biofilms of non-tuberculous mycobacteria, emphasizing the importance of lipid metabolism in establishing and maintaining biofilms [363]. Furthermore, research on lipids (lipidomic analysis) has played a vital role in understanding the link between iron deficiency and the increased susceptibility of drug-resistant Mycobacterium tuberculosis to chemotherapy. This research has illuminated the significance of lipids in the pathogenicity of biofilms and the development of drug resistance [364].
The lipid composition of biofilms is associated with bacterial adaptation and survival, as demonstrated by the molecular relationship between the composition of membrane phospholipids and biofilm formation in E. coli [365]. This suggests that modifying lipid biosynthesis may be a potential strategy for controlling biofilm formation and other characteristics of multicellular organisms. Additionally, it has been recognized that maintaining lipid homeostasis is crucial for the production of mycobacterial biofilms and their ability to adapt to challenging conditions. This underscores lipid metabolism’s significant role in biofilm biology [366].
Utilizing multi-omics techniques has gained significant recognition in the study of biofilms due to the detailed insight they provide into complex microbial populations and their interactions. These techniques incorporate a range of omics technologies, including genomics, transcriptomics, proteomics, and metabolomics, to gain a comprehensive understanding of the intricate mechanisms involved in biofilm formation and function [367, 368, 369]. By utilizing multi-omics techniques, researchers can obtain a more profound understanding of the diverse antimicrobial processes exhibited by substances such as polyphenols on the microbial communities in the gut [370]. Additionally, including multi-omics data enables a more comprehensive understanding of the characteristics of mutations that disrupt metabolic processes in biofilms [371].
Using multi-omics methodologies has proven valuable for tackling the complexities inherent in synthetic multi-species biofilms. This underscores the importance of conducting comprehensive analyses at multiple levels [372]. Furthermore, the development of cutting-edge computational techniques, including artificial intelligence and machine learning, has enabled the integration of biofilm imaging data with extensive multi-omics datasets, presenting exceptional opportunities to explore the relationship between structural patterns and functional properties in biofilms [369].
Moreover, multi-omics analysis can reveal synergistic interactions within biofilms, offering insights into the spatially organized microbial communities and their distinct activities [373, 374] The incorporation of ‘omics’ expression data into genome-scale metabolic models further enhances the representation of pathways relevant to the biofilm setting, providing vital knowledge about the critical processes involved in biofilm formation [375]. Additionally, the MPLEx technique has expedited the measurement of many omics, thereby improving our understanding of the dynamics of microbial communities in biofilms [376].
7.2.8.1 DNA Methylation Analysis
DNA methylation is an essential epigenetic modification on cytosine nucleotides, typically within CpG islands, and is linked with age [377]. This modification is the primary mechanism of epigenetic regulation in bacteria and significantly impacts therapeutically relevant traits, such as biofilm formation [378]. DNA methyltransferases (DNMTs) facilitate this process, and research shows that DNA methylation interacts with histone modifications, leading to changes in gene expression [379].
Biofilm development is a complex process that involves multiple elements, and one of them is managing biofilm production to increase stress tolerance by gene regulation [380]. Gene expression, such as the zapD gene, creates biofilms in Uropathogenic Proteus mirabilis [381]. The MarR family regulator OsbR governs biofilm formation, anaerobic nitrate respiration, and oxidative stress response in Chromobacterium violaceum [382]. The gapB gene also contributes to biofilm creation, and removing it increases extracellular DNA in biofilms [383].
Epigenetic modifications, including DNA methylation, histone modification, and non-coding RNA regulation, are crucial for the reversible and heritable changes in genomic DNA, ultimately affecting gene expression [384]. These modifications are also associated with the progression of other ailments, such as cancer, and have been suggested as potential methods for diagnosis and treatment [385].
7.2.8.2 Chromatin Immunoprecipitation (ChIP) Techniques
Researchers have utilized chromatin immunoprecipitation (ChIP) techniques to investigate the control of biofilm formation at the gene expression level [386]. By applying the Chromatin ImmunoPrecipitation-on-chip technique, they were able to identify the specific promoter areas that are bound by transcription factors during the process of biofilm development [386, 387]. Through genome-wide transcriptional profiling and ChIP approaches, the authors discovered an intricate and interconnected transcriptional network within mature biofilms, uncovering the presence of critical transcriptional regulators and their associated target genes [387, 388]. Another study conducted ChIP-qPCR experiments to assess the interaction between variant histone H3 and the promoters of genes that are either activated or repressed during biofilm formation [388]. These works demonstrate the usefulness of ChIP methods in revealing the regulatory mechanisms responsible for biofilm formation.
Microarrays have proven to be a valuable tool for investigating biofilms, providing researchers with significant insights into gene expression and regulation in various microbes. Subroto et al. [389] emphasized the importance of microarray and RNA-sequencing techniques in investigating biofilms generated by A. fumigatus, revealing a lack of research in this area. Additionally, DNA microarray analysis plays a crucial role in understanding gene expression and adhesion genes in biofilm-associated bacteria, particularly Methicillin-Resistant Staphylococcus aureus (MRSA) [390, 391]. RNA sequencing analysis can also help comprehend gene expression patterns during different phases of in vitro biofilm creation [392].
In addition, Wong et al.’s study [393] highlights the ability of polymer microarrays to identify new polymers that resist biofilm formation, demonstrating this technology’s fast and effective nature. Another study uses a polymer microarray to show how surface chemistry affects bacterial adhesion and biofilm formation, emphasizing the bacteria’s susceptibility to even minor changes in polymer chemistry [394].
In addition, Fajriyah’s article provides a comprehensive overview of microarray technologies, highlighting their extensive use in generating data for bioinformatics research, particularly in the life sciences and biotechnology fields [395]. Moreover, RNA-seq has enhanced sensitivity and a more comprehensive range than microarrays, making it well-suited for investigating biofilms and persister cells that may produce RNA at low levels [341].
Studies have revealed that the CRISPR-Cas system affects the development of biofilms in various bacterial species, including P. aeruginosa [396], Salmonella Typhi [397], and A. baumannii [398]. The presence or absence of the CRISPR-Cas system has been linked to the sensitivity of P. aeruginosa clinical isolates to antimicrobial agents and their biofilm-forming ability [396] The CRISPR-Cas system has also been associated with regulating gene expression and biofilm development [399].
The CRISPR-Cas system is a molecular mechanism in prokaryotic microorganisms, intricately linked with the DNA repair system in bacterial cells [400]. Furthermore, the CRISPR-Cas system has been found to play a role in acquiring virulence genes and controlling biofilm production in Enterococcus faecalis isolates [401]. Notably, the CRISPR-Cas system has demonstrated its ability to control the development of biofilms, revealing its crucial role in bacterial biology.
Quantitative polymerase chain reaction (qPCR) is a powerful technique widely used to analyze biofilms and has been applied extensively in various research fields to quantify and characterize biofilm communities. For instance, Malone et al. [402] utilized DNA sequencing and qPCR techniques to evaluate the effect of different treatment durations with Cadexomer Iodine on chronic biofilm infections in diabetic foot ulcers. Similarly, Zayed et al. [403] employed the method of applied viability DNA extraction and qPCR analysis to investigate changes in the structure and composition of biofilms after undergoing oral biofilm cryotherapy. Additionally, Bravo et al. [404] used qPCR to examine both the overall bacterial counts and the counts of specific bacteria within biofilms that developed on dental implants.
Furthermore, qPCR has been applied to evaluate biofilm treatment. For example, Wang et al. [405] assessed the efficacy of radezolid in killing S. aureus bacteria and preventing the formation of biofilms. Similarly, Y. Shen et al. [406] utilized qRT-PCR to determine the proportion of S. mutans in oral biofilms, demonstrating the usefulness of qPCR in investigating distinct microbial populations within biofilms.
Moreover, qPCR has been used to evaluate interventions’ impact on biofilms’ production and composition. For instance, Praseetha et al. [407] showed how linalool therapy modified the gene expression associated with the harmfulness and production of biofilms in Streptococcus pyogenes. Similarly, Veerapandian and Vediyappan employed semi-quantitative PCR to assess the variation in gene expression of specific genes associated with biofilms in Streptococcus gordonii and C. albicans. This study highlights the importance of qPCR in investigating the effects of treatments on gene expression related to biofilms [408].
Next-generation sequencing (NGS) has proven to be a powerful tool in investigating biofilms. This method provides a comprehensive understanding of microbial populations’ taxonomic composition and community dynamics, making it a valuable resource for researchers [409]. When paired with qPCR, NGS has been extensively utilized to evaluate biofilms without culturing. This approach allows for a more in-depth comprehension of biofilms’ complex nature and the microbial community’s organization [410]. NGS has been particularly useful in studying the microbiome of dental caries, revealing its potential to decipher the intricate microbial makeup of biofilms related to oral health [411]. Additionally, NGS has played a critical role in analyzing subgingival biofilms, providing insights into changes in microbial communities’ composition in healthy and periodontitis-affected areas across multiple generations of subcultures [412]. Finally, NGS has been employed to examine marine biofilms’ microbial diversity and functional capabilities, revealing previously unknown microbial diversity present in marine microorganisms that form biofilms [327].
NGS has been instrumental in examining the microbiome of biofilms in various environments, including extracorporeal membrane oxygenator catheters and drinking water distribution systems. This has provided valuable insights into the taxonomic composition and metabolic behavior of biofilm communities [413, 414]. NGS has also been employed to evaluate the impact of environmental variables, such as sulfonamide concentrations, on the composition and functioning of freshwater biofilm communities [415].
In addition, NGS has proven to be an effective tool for evaluating the efficacy of antimicrobial coatings and identifying areas for potential development of new formulations. This is achieved by enabling the assessment of biofilm formation on different surfaces [416]. Furthermore, NGS has been utilized to investigate the effects of simulated microgravity on the capacity for biofilm production, providing valuable insights into the impact of environmental variables on biofilm growth [417].
Although the investigation of biofilms at the individual cellular level has historically received little attention, recent scientific inquiries have made notable strides in understanding the complexities of biofilm development at this level. For example, refined deep-learning techniques have enabled the latest progress in single-cell segmentation, allowing for monitoring cell lineages and quantifying growth rates in bacterial biofilms [418]. With this methodology, growth rates of individual cells can be quantified throughout space and time during biofilm formation, leading to a deeper understanding of the internal dynamics of biofilms. Additionally, Jeckel et al. [87] conducted a study utilizing experiments, simulations, and statistical analysis to identify common biophysical principles governing the initial development of biofilm architecture at the individual cell level in various bacterial species. This study highlights shared characteristics in biofilm growth across many species and provides significant insights into the initial stages of biofilm establishment [87]. Furthermore, the proliferation of individual cells enclosed within self-produced EPS has been identified as a crucial catalyst for biofilm development [419].
Additionally, the dislodging of individual cells from biofilms has been linked to factors that weaken or erode the biofilm’s structural integrity. These isolated cells produce more enzymes that break down the biofilm matrix than cells that are either planktonic or part of the biofilm [125]. This discovery underscores the importance of understanding the processes that govern biofilm dispersion at the cellular level. Recent research has made significant strides in unraveling the dynamics of biofilms at the individual cell level. This has yielded valuable insights into the shared biophysical mechanisms that drive biofilm formation, the tracing of cellular lineages, the role of EPS in biofilm formation, and the factors that influence biofilm dispersion.
Flow cytometry has become essential for analyzing biofilms, providing valuable insights into microbial communities and their properties. It plays a crucial role in biofilm research by offering comprehensive information about biofilm structure, microbial composition, and functional characteristics. Flow cytometry can investigate the temporal variations in cell counts within biofilms and bulk water, providing insights into the progression of biofilm formation [420]. Additionally, it can assess the functions of key extracellular matrix components in microbial competition for producing floating biofilms, leading to a more comprehensive understanding of biofilm ecology [421]. Flow cytometry is also helpful in measuring and describing dual-species biofilms, providing a thorough evaluation of the populations and interactions within the biofilms [422]. This approach accurately measures the different groups of cells embedded inside biofilms, providing vital knowledge about the arrangement and makeup of cells within biofilms [423].
Furthermore, flow cytometry is used with other methodologies, such as surface plasmon resonance, to observe and track the interactions and movement of nanoparticles inside biofilms in real time [424]. This allows for the acquisition of data on the various stages of biofilm growth and its characteristics. Additionally, it serves as a prompt and accurate substitute for microscopic assessment of bacteria in both free-living and biofilm forms in aquatic samples [425]. Flow cytometry has also been used to monitor the structural modifications of oocysts in biofilm systems [426]. Molecular techniques and application in biofilm reserch are presented in Table 3 (Ref. [200, 327, 339, 344, 350, 352, 355, 356, 364, 372, 381, 390, 391, 399, 400, 405, 411, 418, 427, 428, 429]).
| Method | Application in Biofilm Research | Example | Citation |
| DNA Sequencing and Metagenomics | Identifying the microbial constituents of biofilms and their genetic characteristics. | Sequencing DNA from dental plaque biofilms to study oral microbiota and cystic fibrosis. | [427, 428] |
| Transcriptomics | Examining gene expression in biofilms to elucidate the impact of environmental alterations on biofilm behavior. | Investigating the gene expression in biofilms that develop on medical devices in order to comprehend the reasons behind antibiotic resistance. | [339, 429] |
| RNA Sequencing and Metatranscriptomics | Examining the RNA profiles of biofilms, specifically in intricate, multi-species communities. | Sequencing RNA from diabetes-related foot osteomyelitis. | [344] |
| Proteomics | Analyzing the protein composition of biofilms to identify pivotal proteins responsible for the creation and upkeep of biofilms. | Analyze the proteome of Pseudomonas aeruginosa biofilms to discover potential targets for novel therapeutic interventions. | [350, 352] |
| Metabolomics | Examining the metabolites generated by biofilms in order to comprehend their metabolic pathways and reactions to environmental stresses. | Analyzing the metabolic characteristics of biofilms in relation to various antibiotic therapies. | [355, 356] |
| Lipidomics | Investigation of lipid patterns in biofilms is crucial for comprehending membrane dynamics and signaling. | Investigation of lipids in of Pseudomonas Aeruginosa biofilms. | [364] |
| Multi-Omics Approaches | Utilizing genomes, transcriptomics, proteomics, and metabolomics to comprehensively analyze biofilms and gain a holistic knowledge. | An integrated investigation of multi-species biofilms. | [372] |
| Epigenetics | Explores the impact of epigenetic modifications on the process of biofilm development and behavior. | Investigations on the epigenetics of Proteus mirabilis biofilms. | [381] |
| Microarrays | Analyzing gene expression patterns in biofilms can provide insights into the regulatory networks that contribute to biofilm formation. | Employing microarrays to evaluate alterations in gene expression in MRSA | [390, 391] |
| CRISPR-Cas Systems | Manipulating genetic material to investigate the functions of genes in the process of biofilm creation and upkeep. | Employing CRISPR-Cas9 to disable genes that are believed to contribute to the ability of biofilms to withstand adverse conditions. | [399, 400] |
| Quantitative PCR (qPCR) | Quantifying particular DNA sequences in biofilms is a common method for assessing the abundance of specific bacteria groups or genes. | Quantifying the prevalence of antibiotic resistance genes in biofilms developed chronic infections in diabetic foot ulcers, examining the effectiveness of radezolid in killing S. aureus biofilm. | [344, 405] |
| Next-Generation Sequencing (NGS) | An in-depth examination of the genetic composition in biofilms, offering valuable understanding of their varied and intricate characteristics. | Examining the microbiome of dental caries; and marine samples. | [327, 411] |
| Single-Cell Genomics | Analyzing the genetic material of individual cells within biofilms to uncover the variation within the population. | Analyzing genetic heterogeneity among cells within a single biofilm structure. | [418] |
| Flow Cytometry | Examining the physical and chemical attributes of individual cells inside biofilms is valuable for evaluating cell survival, diversity, and reaction to interventions. | Quantifying the survivability and physiological conditions of biofilms when exposed to various antimicrobial treatments. | [200] |
The application of microtiter plate-based model systems in biofilm research has gained significant attention due to its adaptability and relevance in diverse research domains. Microtiter plate-based techniques have been utilized to investigate the development of biofilms in various microorganisms, including A. baumannii, S. aureus, P. aeruginosa, and E. coli [430, 431, 432, 433]
The efficacy of these methodologies in examining biofilm formation, assessing the impact of different conditions on biofilm growth, and screening antimicrobial drugs has been demonstrated [434, 435]. Microtiter plate-based tests have been employed with other techniques, such as confocal laser scanning microscopy, to assess biofilm formation comprehensively [436].
Furthermore, microtiter plate-based model systems have been utilized to study the inhibitory effects of different chemicals on biofilm growth. For instance, the effectiveness of gold nanoparticles in combating biofilm formation by A. baumannii was evaluated using this technique [437]. Additionally, the microtiter plate technique has been employed to examine the influence of antibiotics on biofilm, thereby demonstrating its usefulness in investigating the efficacy of antimicrobial medicines [432]. Moreover, microtiter plate-based assays have been utilized to study the biofilm inhibitory effects of natural products and synthetic drugs [433, 435]. A schematic representation for the different techniques of common in vitro models for use in biofilm studies are presented in Fig. 6.
 Fig. 6.
Fig. 6.Some commonly used in vitro models for biofilm studies.
The CDC biofilm reactor is an invaluable tool for investigating the complex process of biofilm formation and evaluating the effectiveness of antimicrobial drugs. It recreates the environmental conditions observed in clinical settings, including continuous nutrition supply and shear stress, allowing for controlled biofilm cultivation [438]. The reactor has been extensively utilized to examine the impact of biomaterial characteristics on biofilm growth and investigate the formation of biofilms on various surfaces by different pathogens [439, 440, 441, 442]. Additionally, it has been employed to cultivate a diverse range of oral microbiota for scientific investigation purposes [443]. The CDC biofilm reactor has been confirmed as a reliable and consistent system for studying several aspects of biofilm physiology, morphology, growth dynamics, and antibiotic susceptibility profiles [248, 444]. Computational fluid dynamics has also been utilized to examine the shearing stresses within the reactor [445].
Furthermore, the reactor has been observed to facilitate the development of more intricate and resilient biofilm formations compared to alternative techniques, making it a valuable instrument for investigating biofilm architecture and antimicrobial resistance [438, 446].
Capillary biofilm reactors (CBRs) have shown potential in cultivating high-density biofilms due to their high surface area-to-volume ratio and exceptional cell densities [447]. Nevertheless, one major challenge in implementing CBRs is the extended period required for biofilm formation, which can take up to 5 weeks [448]. Additionally, research indicates that the continuous operation of CBRs promotes microbial growth, resulting in denser biofilms and higher current densities [449].
Moreover, the use of CBRs for the continuous oxidation of cyclohexane to cyclohexanol has been explored, revealing the potential of employing specific microorganisms at high cell densities [450]. This study emphasizes the importance of evaluating the technique in expanding biofilm-based capillary reactor systems [451]. This reactor can assess biofilm development, including biofilm dispersion, where individual cells exit the biofilm and return to a planktonic state [129]. This highlights the fluid nature of biofilms and the significance of comprehending their life cycle for efficient reactor design and operation.
Extensive research has been conducted on using flow cell reactors for cultivating biofilms, focusing on investigating key factors such as biofilm development, biomass production, and reactor performance. For instance, a recent study employed light-sheet microscopy to reveal a coordinated flow, akin to a fountain, that promotes the sideways spread of the biofilm by transferring cells toward its leading edge [262]. This discovery highlights the critical role flow dynamics play in the growth and spread of biofilms. Moreover, it was found that hydrodynamic conditions significantly impact the plasmid copy number in both planktonic and biofilm cells, underscoring the importance of fluid dynamics in genetic expression within biofilms [452]. Other studies have focused on using drip-flow biofilm reactors to simulate the growth of biofilms under consistent laminar flow conditions. These studies have emphasized the need to maintain controlled flow conditions in biofilm research [453, 454]. A study examining the flow dynamics of a reverse fluidized bed biofilm reactor has provided valuable insights into how the speed of air near the surface affects the behavior of the biofilm [32].
The potential of rotating disk reactors has been widely explored across various fields, including biofilm research, photocatalysis, and wastewater treatment [455]. One study investigated the formation of drinking water biofilms in a rotating annular reactor, highlighting the impact of turbulence on biofilm development [455]. Similarly, another study emphasized the importance of rotation speed in a photocatalytic degradation process using a photochemical rotor-stator spinning disk reactor [456]. These findings underscore the crucial role of rotation in regulating biofilm formation and mass transfer within the reactor.
Furthermore, researchers have examined the effects of mechanical stresses on biofilm elimination and cyclohexanone conversion in rotating biofilm reactors. They have found that high shear rates produced by rotational speed are crucial for biofilm growth and developing a structure suitable for biocatalytic applications [454, 457]. Another study presented a rotating disk bioelectrochemical reactor (RDBER) capable of cultivating both cathodic and anodic biofilms, with the added benefit of being autoclavable and allowing for observation of biofilm development through optical coherence tomography [458].
A drip-flow biofilm reactor is an essential tool for exploring the behavior and development of biofilms in various scenarios. This reactor model has been utilized in numerous studies to examine biofilm formation in chronic wound infections [459], investigate oral biofilms [460], evaluate the efficacy of biofilm management in wound infections [461], and monitor treatment responses in models of periodontal multispecies biofilms [453]. It has also been used for cultivating, treating, collecting, and examining P. aeruginosa biofilms [462]. The unique feature of the drip-flow biofilm reactor is that it allows for the growth of biofilms at the interface between air and liquid while maintaining a constant and steady flow. This creates a model that resembles a problematic wound environment [453, 461]. Additionally, it has been utilized to study the reactions of anaerobic biofilms to various electron acceptors, which affect the effectiveness of selenium removal and the characteristics of the biofilm [463]. The reactor has also cultivated single and mixed flow-cell biofilms on various metal surfaces at the air-liquid interface [464].
The widely used Robbins method has been updated to provide a more effective way of studying biofilm development and evaluating antibiofilm agents. The technique involves using a specialized device that can rapidly create and mold biofilms in a liquid, offering promising results for antibiotic lock treatment in removing biofilms from colonized surfaces [465]. Among the several biofilm reactors used in biofilm labs, the modified Robbins device is crucial to biofilm research [466].
The laminar flow chamber of the modified Robbins device comes equipped with suspended substrates to examine biofilm growth under controlled experimental settings. It is highly promising and indicative of in vivo conditions [182, 467]. Moreover, it has been used to recreate biofilms of P. aeruginosa and MRSA, demonstrating its adaptability in studying different biofilm varieties [468]. The updated Robbins device now comes with more sampling ports for analysis, allowing for the simultaneous collection of multiple biofilm samples at different time points throughout biofilm growth [467].
The modified Robbins method’s significance is further emphasized by its inclusion in many biofilm models, such as the Calgary biofilm device and the CDC rotating biofilm reactor. This indicates its broad adoption and utilization in biofilm research [469, 470]. Additionally, this technique has been used to investigate bacterial attachment to various surfaces in static and dynamic environments, demonstrating its versatility in different experimental scenarios [471, 472].
The annular biofilm reactor has proven to be an invaluable tool in a wide range of research, demonstrating its significance in studying biofilm formation, microbial activity, and the impact of various substrates and environmental factors on biofilm growth. Lee et al. [473] utilized this method to investigate the effect of pipe material on biofilm formation and structure in drinking water distribution systems.
Moreover, it has been utilized to replicate the shear stress experienced by sewer biofilms during flow designed for self-cleaning purposes, making it highly relevant to environmental science and wastewater treatment [474]. The lab-scale rotating annular reactor has also been used to assess the spatial arrangement of antibiotic resistance genes and integrons in biofilms, underlining the importance of studying microbial behavior and biofilm resistance [475].
In addition, a rotating annular reactor has been employed to investigate biofilm growth in diverse flow patterns, demonstrating its effectiveness in comprehending the process of biofilm formation under distinct circumstances [455]. Furthermore, the reactor has been used to assess the endurance of naturally occurring biofilms, highlighting its significance in evaluating the longevity and stability of biofilms [476]. Table 4 (Ref. [129, 432, 434, 435, 440, 446, 452, 453, 455, 456, 460, 461, 468, 471, 472, 473, 475, 477, 478]) is a summery of these reactors.
| Model System/Reactor | Description | Example in Biofilm Research | Citation |
| Microtiter Plate-Based Model Systems | Compact platforms designed for efficient investigation of biofilm growth at a large scale, while considering different environments. | Evaluating the development of biofilms in various microorganisms; Examining the influence of different conditions on biofilm development, and screening antimicrobial drugs. | [432, 434, 435] |
| CDC Biofilm Reactor | The Centers for Disease Control and Prevention (CDC) has developed a reactor specifically designed to cultivate biofilms on detachable coupons. | Investigating the effectiveness of antimicrobial agents in combating biofilms inside healthcare environments; Examining the formation of biofilms by different pathogens on diverse surfaces. | [440, 446, 477] |
| Capillary Biofilm Reactor | Utilizes capillary tubes to achieve uninterrupted fluid flow and facilitate the creation of biofilms, replicating the natural environment seen in water pipes. | Assessing the formation of mixed biofilms by pathogenic bacteria and the efficacy of antimicrobial treatments; Investigating the process of biofilm dispersion. | [129, 478] |
| Flow Cell Reactor | An apparatus that enables the examination of biofilms in a fluid environment, typically with the ability to capture images in real-time. | Examining the structure of biofilms and gene expression alteration in relation to different flow velocities; Simulating the growth of biofilms in a consistent laminar flow. | [452, 453] |
| Rotating Disk Reactor | A biofilm growth reactor equipped with revolving disks to regulate shear stress and promote the formation of biofilms. | Analyzing the influence of shear stresses on the architecture and resilience of biofilms; Examined the proliferation of drinking water biofilms in a rotating annular reactor under various flow patterns. | [455, 456] |
| Drip Flow Reactor | A slow-flowing liquid reactor commonly employed for the investigation of biofilms on medical devices. | Simulating the growth of biofilms in wound infection; Investigate oral biofilms. | [460, 461] |
| Modified Robbins Device (MRD) | An apparatus that facilitates the cultivation of biofilms in a regulated flow environment on detachable coupons. | Examining various biofilm varieties; Investigate the attachment of bacteria to various surfaces in both stationary and dynamic environments. | [468, 471, 472] |
| Annular Biofilm Reactor | A cylindrical reactor designed to facilitate the cultivation and analysis of biofilms on various surfaces in the presence of flowing conditions. | Examining biofilm removal strategies in industrial water systems; Investigating microbial behavior and resistance in biofilms. | [473, 475] |
Recently, interest in utilizing 3D-printed devices for examining biofilm behavior has been a noticeable surge in interest. The ability of 3D printing to generate consistent and reproducible biofilms has been highly advantageous in evaluating preventive measures against biofilms. 3D printing in biofilm research offers exciting prospects for future exploration and progress. Recent reports have highlighted the development of biofilm growth systems created solely with 3D printing technology, including a flow device used to study chronic wound infections [479]. The use of 3D-printed microfluidic gadgets for studying biofilm activity is undoubtedly compelling. 3D printing has significantly transformed microfluidics, providing new opportunities and capabilities for examining biofilms. A study by Waheed et al. [480] serves as a prime example of the capabilities of 3D printing technology in producing a customized microfluidic flow cell for studying biofilms in their natural environment in conjunction with microscope analysis.
3D printing technology has revolutionized biofilm research, allowing scientists to conduct meticulous analyses and comprehend biofilms’ intricacies by generating elaborate structures that replicate actual environmental conditions [481]. By utilizing 3D printing, researchers can precisely control variables such as surface roughness, porosity, and topography to faithfully recreate conditions, which is particularly useful for investigating how different surfaces impact the development and expansion of biofilms [482]. For instance, Hall et al. [483] conducted a study using biofilm quantification, surface topography analysis, and 3D projections to gain insights into bacterial attachment to 3D-printed surfaces.
Integrating 3D printing technology is essential in developing flow cell systems that can replicate the fluid dynamics of various settings, such as pipelines or medical devices. Through microfluidic devices, researchers can investigate the growth of biofilms under specific conditions, like shear stress, with exact control of environmental factors, temperature, and nutrient flow [484, 485, 486].
Within drug delivery research, 3D printing presents a unique opportunity to fabricate devices capable of accurately administering medications to biofilms. This approach enables researchers to evaluate the efficacy of antimicrobial drugs and explore how they penetrate biofilm structures. By utilizing 3D-printed drug delivery devices, researchers can assess the effectiveness of various medications in interrupting biofilm development and growth [487, 488].
The precision and adaptability of 3D printing offer researchers a powerful tool for creating tissue scaffolds with precise geometries and architectural characteristics. By accurately replicating the microenvironments of various tissues, these scaffolds provide a more genuine portrayal of physiological conditions and offer an enhanced correlation with human physiology [489]. This is particularly important in biofilm research, where the structure of live tissues can be accurately replicated, enabling a more precise investigation into the interaction between biofilms and host tissues and their response to different stimuli [490].
3D printing technology has enabled researchers to understand biofilm structures better, more precisely, and efficiently. By creating transparent models of these structures, surveillance and examination of growth and behavior are simplified, providing valuable insight into the process of biofilm creation [491, 492]. The transparency of these 3D-printed structures enhances imaging techniques, such as microscopy, allowing for accurate analysis of the biofilm structures. This level of precision is crucial for obtaining comprehensive data on the composition and interactions of biofilms [491, 492].
Biofilms are a complex microbial community that requires constant monitoring to understand their growth and behavior. Recently, nanoparticle sensors have emerged as a dynamic and quantitative method for studying these intricate microbial communities. These sensors are designed with materials that exhibit precise reactions to parameters associated with biofilm growth [493]. Various types of nanoparticles, such as quantum dots, gold nanoparticles, or magnetic nanoparticles, are utilized for their unique features and ease of customization [494].
To allow for selective binding to particular biomolecules, ions, or environmental variations linked to the development of biofilms, the surface of nanoparticles is typically modified with chemicals that interact specifically with biofilm components [495]. Once introduced into the biofilm environment, either by being dispersed directly into the growth medium or incorporated into biofilm substrates, the nanoparticles interact with the biofilm matrix and react to changes in the microenvironment [496].
The precise data generated by observing nanoparticle sensor characteristics supplies researchers with valuable insights into the growth and behavior of biofilms. Researchers can quantify various factors, including the thickness of biofilms, the amount of living matter, and the rate of metabolic processes [493, 497]. This allows for a better understanding of how the biofilm community functions and changes over time. Nanoparticle sensors enable uninterrupted and extended surveillance of biofilm dynamics, enabling researchers to scrutinize the progression of biofilms over time, their reactions to environmental stimuli, and their responses to antimicrobial interventions [498, 499].
In the realm of biofilm research, in silico techniques utilize computer methods, simulations, and models to explore biofilm production and behavior complexities. These computational tools provide a virtual platform for researchers to investigate biofilm biology without the limitations of physical experiments.
Among these techniques, Agent-Based Modeling (ABM) targets individual agents, representing various organisms, such as bacteria or cells, within a simulated environment. ABMs allow for a detailed analysis of interactions and behaviors at a small scale, providing valuable insights into the emergence of characteristics and the development of intricate biofilm structures [500]. Notably, ABMs have played a crucial role in stimulating the growth of biofilms and accurately representing the complex structure that arises from the interactions between individual cells and their surroundings [501]
Computational Fluid Dynamics (CFD) is employed to replicate the movement of fluids within biofilm environments, providing valuable insights into how nutrition transport, shear stresses, and fluid dynamics influence the growth and spread of biofilms [467, 486]. This technique is essential for comprehending the effects of different flow conditions on the production of biofilms. Xia et al. [502] developed a three-dimensional individual-based biofilm model by combining CFD with the discrete element method (DEM). Their objective was to predict the impact of EPS on the deformation, recovery, and detachment of bacterial biofilms in flowing conditions [502]. Genome-scale metabolic Modeling (GEMs) involves using computational models that can analyze and predict cellular behavior on a large scale by encompassing an organism’s entire metabolic network [375]. This technique helps explore the complexities of biofilm metabolism by creating computational models of the metabolic processes in microorganisms that generate biofilms. These models can provide valuable insights by examining the impact of changes in nutrition supply or environmental factors on biofilms’ growth and metabolism [503]. Altay et al. [504] demonstrated the power of GEMs by using them to uncover the changes in metabolism that occur during the formation of biofilms in Burkholderia cenocepacia. Their study examined the metabolic alterations associated with biofilm creation, shedding light on the metabolic reconfiguration of B. cenocepacia during this process [504].
Utilizing molecular dynamics (MD) simulations, it is possible to accurately replicate atomic and molecular interactions within biofilm structures on a molecular scale. This method provides valuable insights into biofilm components’ dynamics, interactions, and structural stability, ultimately leading to a deeper understanding of their mechanical properties [505, 506]. In a recent study by Sharma et al. (2023) [507], MD simulations were utilized to investigate the stability of protein and ligand complexes, providing valuable information on the molecular dynamics of biofilm-related events.
Bioinformatics and computational biology play a critical role in the study of biofilms. Using computational methods, large amounts of genomic, transcriptomic, and proteomic data can be analyzed to identify essential genes, pathways, and regulatory networks associated with biofilm development. This valuable insight into the molecular foundations of biofilm dynamics provides a deeper understanding of these microorganisms and their behavior [508, 509].
The application of network modeling in biofilm research has proven to be a dependable technique that yields a thorough understanding of the intricate connections within a biofilm. This methodology involves the creation of interaction networks that portray the complex interrelationships between various components within the biofilm, including microbial species, genes, and proteins [509, 510]. In these networks, nodes represent distinct entities, while edges signify interactions, dependencies, or regulatory connections [511].
Dealing with biofilms is a challenging task, especially when it comes to combating antibiotic resistance. These films are notorious for their ability to confer resistance to multiple antibiotics, creating a significant obstacle in clinical settings [512]. Their resilient structure, restricted antibiotic penetration, and enhanced resistance transfer all contribute to the difficulties associated with antibiotic therapy [513]. Moreover, biofilm cells exhibit rapid microevolution in response to antibiotics, resulting in resistance levels far surpass those observed in planktonic cultures [337]. The bacteria’s ability to acquire resistance against conventional antimicrobial treatments that rely solely on antibiotics only exacerbates the issue [514]. The significant resistance of biofilms to various antimicrobials worsens the ongoing antibiotic crisis [515]. Protective EPS molecules encasing biofilms create microenvironments that pose challenges in dispersing them with conventional antibiotics [516].
Biofilms significantly enhance microbial cell antibiotic resistance, yet the specific mechanisms that facilitate this ability are not fully known [517]. Alternative therapies are urgently needed due to the lack of effective treatment options for antibiotic-resistant bacteria in biofilms [518].
Numerous factors contribute to the resistance of biofilms to antibiotic treatment, including their multicellular nature, EPS barrier, activation of efflux pumps, synthesis of enzymes that degrade antibiotics, heterogeneous modulation of metabolic activity and growth rate, and presence of highly drug-tolerant persister cells [519, 520]. Additionally, the varying occurrence of resistance resulting from biofilm penetration could help explain why some antibiotic treatments with limited ability to penetrate biofilms may depend on bacterial management to inhibit resistance development [521]. Effectively addressing biofilm-associated illnesses and antibiotic resistance requires a comprehensive understanding of the mechanisms involved in antibiotic resistance in biofilms. Comprehending and controlling biofilms, particularly concerning antibiotic resistance, presents complex challenges that require a thorough understanding of these mechanisms.
The comprehension and control of biofilms present a complex challenge due to their many components and nuanced attributes. Sauer et al. [82] proposed an extended conceptual framework that offers a unified basis for enhancing our understanding of biofilm formation and devising customized antibiofilm approaches. However, the intricate nature of biofilms poses a substantial obstacle in developing efficient antibiofilm techniques, perpetuating their persistence as a problem across various domains [82].
To comprehend biofilms, it is essential to understand the spatial and temporal arrangement of the biofilm matrix [522], which is crucial for their perception and control. However, this continues to be a significant obstacle due to the complexity of the samples and the limited range of methods currently available. Additionally, the wide range of glycoside hydrolases found in biofilms, as examined by Ellis et al. [523], creates challenges in anticipating or understanding the enzymes that may have potential therapeutic uses in the future. The intricate nature of these enzymes adds to the challenge of creating effective therapies for illnesses associated with biofilms.
The characteristics of biofilm-associated infections, such as their resilience and endurance, have become a significant obstacle to public health [524]. The presence of such resistance adds complexity to managing these infections. Suarez et al.’s research [119] further emphasizes the importance of biofilm thickness in shaping microbial communities, making it challenging to predict their structure accurately. Additionally, Jana et al.’s investigation [525] into the non-linear rheological properties of bacterial biofilms consisting of a single species demonstrates variability, potentially due to the diversity of ingredients in the extracellular matrix. Understanding the connection between phenotypic traits, diverse development settings, and community characterization is crucial in establishing structure-property relationships in oral biofilms [526].
The extent of variation in different strains of biofilms and their vulnerability to EPS-breaking enzymes [527] still needs to be better understood, underscoring the need for further investigation. Staats et al. [528] stress the significance of thorough comprehension of biofilms at a molecular level in a shared environment to counteract biofilm development in periprosthetic joint infections. Overall, these issues highlight the necessity for continuous investigation and innovative strategies to gain comprehensive comprehension and control of biofilms.
Dealing with biofilms in the context of chronic infections is a complex and persistent challenge in modern medicine. Biofilms are renowned for their resilience and ability to resist traditional treatment approaches, making biofilm-associated infections an ongoing and formidable obstacle [524]. One specific challenge is the limited effectiveness of certain therapeutic approaches, such as the restricted ability of photosensitizers to penetrate biofilms coupled with high levels of biofilm glutathione that consume reactive oxygen species. This reduces the efficacy of photodynamic therapy for treating biofilm infections [529]. Exploring alternate or more efficient therapeutic approaches to combat biofilms effectively is crucial.
Biofilm-related infections, especially in chronic wounds, pose a significant global challenge. Developing tailored antimicrobial therapies to improve wound healing is essential [530, 531, 532, 533]. These infections require specific care due to unique challenges, such as bacteria associated with biofilms having increased resistance to antibiotics. The notion of “biofilm tolerance”, which refers to the heightened resistance provided by biofilm lifestyles, adds complexity to disease management [148, 164, 534]. Biofilm presence in various therapeutic scenarios, such as infections related to orthopedic devices, poses significant therapy challenges [535, 536]. The correlation between microorganisms that create biofilms and infections in prosthetic joints affects all aspects of infection management, including diagnosis, treatment, and prevention techniques.
It is important to consider the resilience of biofilms to cleaning agents, disinfectants, and antibiotics. This knowledge can help us develop better strategies to effectively address diseases caused by biofilms and mitigate their impact [537]. Additionally, the variation within biofilm communities of the same species, which enhances individual biofilm populations’ ability to survive and thrive, adds an extra layer of complexity to biofilm management [538]. Biofilms present long-lasting challenges in understanding and controlling diseases. Due to their tenacious nature, increased resistance to traditional treatment approaches, and correlation with diverse clinical disorders, they pose a difficult challenge in the healthcare field. To overcome these problems, it is crucial to develop customized antimicrobial strategies, enhance our understanding of biofilm production, and explore alternative treatment methods to combat illnesses associated with biofilms effectively.
Identifying and evaluating biofilms present complex challenges across numerous fields, particularly in modern medicine and clinical practice. One of the biggest hurdles in clinical settings is accurately detecting biofilm infections. Silva et al. [539] noted that the absence of a universally accepted diagnostic protocol poses significant difficulties in pinpointing illnesses related to biofilms. This lack of standardized diagnostic procedures can lead to inconsistencies and complications regarding treatment.
Traditional methods of detecting biofilms, such as culture, microscopy, and biochemical assays, have notable limitations. These techniques may need to be more accurate in the total number of biofilm populations due to the presence of viable but non-culturable (VBNC) cells [540, 541]. In addition, the methods commonly used to tackle biofilm-related issues often result in inaccurate adverse outcomes and lack precision and accuracy [542, 543]. Addressing the complexities associated with comprehending and managing biofilms involves many challenges, such as diagnosis, identification, evaluation, and the development of effective treatment strategies. A multidisciplinary approach, standardized protocols, and advanced assessment methodologies are required to manage biofilm-related concerns effectively. The mysterious attributes of EPS further complicate the intricate nature of biofilms. Seviour et al. [544] have highlighted that these compounds are essential for biofilm formation and operation. The limited knowledge of EPS compounds contributes to the difficulties of accurately targeting and controlling biofilms in various environments [544].
Moreover, ongoing research has found that pinpointing the bacteria responsible for biofilm formation remains challenging [545]. Effective biofilm management is paramount due to their propensity to develop in various environments. However, assessing biofilms in clinical settings requires intricate techniques, as Magana et al. [249] emphasize the need for comprehensive approaches to characterize, monitor, and quantify biofilms while also creating efficient imaging and sensing tools.
Additionally, discovering new drugs, particularly antifungal agents, to counteract biofilm production in clinical care is challenging [546]. Even preventing and disrupting biofilms in settings such as contact lens cases presents methodological difficulties [547]. The resistance of bacteria residing in biofilms to antibiotics and disinfectants, demonstrated by species like Salmonella, makes it challenging to determine the effectiveness of these treatments [548].
The practical application of antibiofilm peptides (ABPs) for therapeutic purposes and their evaluation using murine biofilm models pose considerable challenges [549]. Furthermore, evaluating biofilm formation on diverse surfaces, ranging from marine to dental surfaces, requires a comprehensive approach to assess the impact of surface attributes on biofilm growth [550, 551].
The management of biofilms in environmental settings is challenging due to the resilient nature of these microbial communities. Biofilms exhibit exceptional resistance to antimicrobial agents and ecological stresses, which can pose significant risks to public health and environmental sustainability [149, 552].
Within water distribution networks and aquatic ecosystems, biofilms are essential in managing microbial communities and pharmaceutical contaminants, as evidenced by numerous studies [553, 554]. However, the adaptability of biofilms to their surroundings adds further complexity to understanding and controlling these organisms in the environment [369, 500].
Controlling biofilm infections presents a significant challenge due to their resilience to various antimicrobial drugs, including antifungal therapies. The development of low-oxygen microenvironments within the fungal biofilm structure highlights the complex nature of these infections [92, 555]. Moreover, biofilms can adapt and shield themselves against stress factors, underscoring their durability and presenting additional obstacles to the creation of effective control strategies [1].
Biofilms significantly impact both human health and environmental processes, including biogeochemical cycling and the breakdown of environmental pollutants—however, their ability to form organized colonies and withstand environmental stress challenges ecological systems. To fully grasp the environmental risks associated with biofilms and their effects on complex populations, it is crucial to understand their impact on microbial communities and behavior in aquatic environments [556].
Addressing the impact and management of biofilms requires a multidisciplinary approach. This approach should include improved detection techniques, a thorough understanding of how biofilms behave in natural environments and developing and implementing effective control strategies. Addressing these intricate issues can decrease biofilms’ adverse effects on environmental systems, human health, and industrial processes.
The persistence of biofilms presents a significant challenge in both medical and environmental contexts due to their inherent toughness and resilience to conventional antimicrobial therapies. Extensive research, as discussed earlier [557, 558], has demonstrated the resistance of biofilms to antibiotics and antimicrobials, underscoring the difficulty of eliminating infections caused by these structures. This resistance makes biofilm control more challenging and raises serious concerns about the efficacy of current treatment approaches.
Another study highlighted the restricted availability of efficacious therapeutic choices as a crucial element in the difficulty of eliminating biofilms [558]. This limitation contributes to difficulties in early detection, inadequate distribution of traditional medications within the biofilm structure, altered conditions within biofilms, and the rapid adaptation and emergence of resistance in bacterial populations [171].
A comprehensive approach has been explored to tackle the challenge of biofilms. A proactive strategy has been developed to create novel antibacterial techniques to hinder biofilm production [559, 560]. A more holistic approach also involves integrating biological, physical, and chemical methods to manage and eliminate biofilms [561]. By utilizing combination therapies that specifically target critical proteins involved in biofilm development and control, there is promising potential to disrupt the entire lifecycle of biofilms [562].
Moreover, a study on developing new materials and techniques to eliminate biofilms suggests a shift towards more targeted and efficient treatment approaches. Nonthermal plasmas have shown promise in laboratory settings for combating bacteria associated with biofilms, providing a unique physical method for controlling biofilms [563, 564, 565]. To effectively eliminate biofilms, a thorough and multifaceted strategy is necessary. Addressing the issue of biofilm elimination requires the development of new antibacterial techniques, combination medications, and innovative bio-remedial procedures. The resilience and flexibility of biofilms necessitate the improvement of existing treatments and the creation of new methods that can successfully dismantle biofilm structure and resistance mechanisms. This comprehensive strategy exemplifies the collaborative effort of academics and healthcare professionals in their pursuit to address and control biofilm-related illnesses and environmental consequences efficiently.
The study of biofilms is a complex and interdisciplinary field that poses significant research gaps. Due to biofilm activity’s intricate nature and its wide-ranging effects on sectors like healthcare, environmental management, and industrial practices, collaborative research efforts spanning multiple disciplines are crucial for advancing our comprehension and control of biofilms.
In the medical realm, biofilms can have significant consequences for human health, particularly in the context of medical device-related infections. A recent study underscores the importance of collaboration between medical researchers and engineers to develop efficient approaches for detecting, managing, and eliminating biofilms in intricate infections [566, 567]. The manipulation of materials and surfaces is crucial in preventing or facilitating biofilm growth, highlighting the need for interdisciplinary cooperation.
Biofouling is a similarly complex problem in environmental research and industrial applications, affecting water treatment systems, shipping industries, and other sectors. Thorough research is necessary to develop detection methods that can be universally applied across different fields of study [567]. Microbiologists, environmental scientists, and materials engineers must collaborate due to the intricate nature of biofouling to create inventive and environmentally friendly, efficient, and long-lasting solutions.
The study of biofilms from an interdisciplinary perspective can yield new insights into their spatial arrangement. Collaborative studies on biofilm structure have enhanced our understanding of microbial interactions and survival strategies within these communities [568]. This knowledge is critical for developing effective strategies to break down or utilize biofilms, depending on the intended objective.
Biofilm engineering has opened up exciting possibilities in the field of material science. In their research, Ng et al. [569] demonstrate the potential of biofilms to generate catalytic devices, highlighting the promising intersection of microbiology and materials science for innovative applications. The convergence of biofilms and technology presents unique opportunities but requires a deep understanding of biofilm biology and engineering principles.
Integrating machine learning into biofilm research has been suggested to harness the possibilities of interdisciplinary collaboration [570]. Machine learning has the potential to provide powerful tools for identifying patterns and predicting the development and behavior of biofilms. However, achieving this goal requires a strong partnership between biofilm and data science experts to translate complex biological mechanisms into computer models.
Despite advancements in the biofilm field, significant shortcomings remain areas for improvement, particularly in translating academic research into practical applications in industries. Highmore et al. [44] highlighted the disparity between experimental results obtained in a controlled laboratory setting and their implementation in real-life scenarios. To bridge this gap, it is essential to foster collaborations between academics and industry to expedite the application of biofilm research findings into tangible solutions.
In veterinary medicine, there has been a noticeable lack of direct research on the therapeutic significance of microbial biofilms [571]. This presents an interdisciplinary area for investigation that has yet to be fully explored. This underscores a broader issue: understanding human biofilm infections could be more effectively utilized in veterinary settings, representing a missed opportunity for transferring knowledge between different fields.
Furthermore, interdisciplinary research is necessary to explore the economic dimensions of biofilms [572]. The economic impact of biofilms spans several industries, requiring a comprehensive approach that integrates scientific research and financial analysis.
There are numerous multidisciplinary gaps in biofilm research, including machine learning, veterinary medicine, biochemical engineering, and economic analysis. Collaborative research addressing these knowledge gaps is crucial to developing a deeper understanding of biofilms and using scientific findings practically. Incorporating multiple disciplinary perspectives will be essential in formulating groundbreaking solutions to the complex issues presented by biofilms.
The field of biofilm research and control is a rapidly evolving area that continually presents us with new interdisciplinary methods to address biofilm production, detection, and elimination issues. The future of biofilm research looks promising as it encompasses a wide range of possibilities, including the development of new and advanced antimicrobial substances, innovative therapeutic approaches, and the exploration of alternative remedies for infections associated with biofilms. In particular, creating medicines that target multiple aspects of the complex biofilm microenvironment is very promising.
Furthermore, there is a growing potential for investigating new technologies specifically targeting biofilms with the help of studies conducted in living organisms or outside the body. Future biofilm research may investigate alternative methods to eliminate pre-existing biofilms or prevent biofilm formation by utilizing alternative antimicrobial agents derived from natural sources. This could lead to the development of novel control techniques involving chemical synthesis and using quorum-sensing molecules. There is also great potential in exploring alternative antimicrobial approaches to manage oral biofilm-associated infections, such as using natural substances and alternative antimicrobial agents.
In addition, advancing technology that explicitly targets biofilms and creating efficient techniques to address the challenging problem of bacterial persistence in aquatic ecosystems are promising avenues for future biofilm research and control. Overall, the future of biofilm research and control holds great promise for addressing issues related to biofilm production, detection, and elimination, and we can look forward to exciting new developments in the field.
Through the integration of advanced imaging techniques and detailed molecular investigations, this integrative study has provided a comprehensive understanding of the intricate dynamics of biofilms. Our examination of the complex structure of biofilms, the intricacies of EPS, and the mechanisms driving antibiotic resistance has been successful. The use of confocal laser scanning microscopy, electron microscopy, and fluorescence microscopy has revealed the intricate composition of biofilms, demonstrating the essential role played by EPS in maintaining their stability and defense mechanisms. Additionally, molecular techniques such as DNA and RNA sequencing, proteomic and metabolic investigations have uncovered the genetic and metabolic networks that support biofilm communities and enhance their ability to withstand antimicrobial treatments. Of significant importance, this study has highlighted the role of EPS in not only maintaining biofilm structure but also enabling genetic exchanges that result in heightened antibiotic resistance. This knowledge is critical in the medical profession, where biofilm-associated infections present a challenging task, as well as in environmental and industrial settings, where effective biofilm management is crucial. This study’s broad perspective represents progress in our ongoing efforts to comprehend and control biofilm behaviors for improved health outcomes and environmental management. As we delve deeper into these microbial cities, it becomes increasingly apparent that a comprehensive approach is necessary for devising effective solutions to the challenges posed by biofilms.
AA was responsible for the entire preparation of this manuscript.
Not applicable.
Researcher would like to thank the Deanship of Scientific Research, Qassim University for funding publication of this project.
This research received no external funding.
The author declares no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.