












1 Department of Pathology, Kunming Medical University, 650500 Kunming, Yunnan, China
2 Department of Pathology, First Affiliated Hospital of Kunming Medical University, 650032 Kunming, Yunnan, China
3 Department of General Surgery, Xinqiao Hospital, Army Medical University, 400037 Chongqing, China
†These authors contributed equally.
Abstract
Background: Breast cancer is the commonest global malignancy and the primary cause of carcinoma death. MCM6 is vital to carcinogenesis, but the pathogenesis of MCM6 remains unclear. Methods: MCM6 expression in patients with breast cancer was examined through The Cancer Genome Atlas (TCGA) database, immunohistochemistry, Quantitative Real-Time PCR (qRT‒PCR) and Western blotting. The prognostic factors were assessed by the Kaplan‒Meier method and Cox regression. On the basis of the key factors selected by multivariable Cox regression analysis, a nomogram risk prediction model was adopted for clinical risk assessment. The TCGA database was utilized to determine how MCM6 is correlated with chemotherapy sensitivity, immune checkpoint-related genes (ICGs), tumor-infiltrating immune cells, along with tumor mutation burden (TMB) and methylation. The impact of MCM6 on carcinoma cells was investigated in terms of proliferation, cell cycle as well as migrating and invasive behavior through CCK assays, flow cytometry, wound healing assays, Transwell assays and xenotransplantation experiments. Results: MCM6 expression was upregulated, which is closely associated with the size of the tumor (p = 0.001) and lymph node metastasis (p = 0.012) in patients with breast cancer. Multivariate analysis revealed MCM6 to be an independent risk factor for prognosis in patients with breast carcinoma. The nomograph prediction model included MCM6, age, ER, M and N stage, which displayed good discrimination with a C index of 0.817 and good calibration. Overexpression of MCM6 correlated with chemotherapy sensitivity, immune checkpoint-related genes (ICGs), tumor-infiltrating immune cells, tumor mutation burden (TMB), and methylation. Silencing MCM6 significantly inhibited proliferation, prolonged the G1 phase of the cell cycle, and restrained the proliferation, migration and invasive behavior of cancerous cells and inhibited tumor growth in vivo. Conclusions: Our research shows that MCM6 is highly expressed in breast cancer and can be used as an independent prognostic factor, which is expected to become a new target for the treatment of breast cancer in the future.
Keywords
- breast cancer
- MCM6
- prognosis
- nomogram
- drug sensitivity
- immune cell
Breast cancer is the most common global malignancy and the primary cause of carcinoma death, severely damaging the health of the female population [1]. Breast cancer is a kind of disease with high heterogeneity in morphology, molecular biology, clinical manifestations and therapeutic response that leads to prevention and treatment remaining challenging worldwide [2, 3, 4]. In recent years, many genes related to the progression of breast cancer have been discovered according to its pathogenesis and drug resistance mechanism. Traditional chemotherapy combined with targeted therapy and immunotherapy achieved better effects than before. For instance, trastuzumab, a monoclonal antibody utilized as a standard treatment for malignancy in the case of Human epidermal growth factor receptor-2 (HER2) overexpression in cancer cells, has an objective response rate of 26% in clinical treatment [5]. The combined medication of cetuximab and cisplatin could inhibit Epidermal Growth Factor Receptor(EGFR) and prolong progression-free survival by 1.5 to 3.7 months and overall survival (OS) time extended from 9.4 to 12.9 months [6]. Targeted therapy is of vital importance for the overall prevention and treatment of malignancy. However, continued treatment results in a loss of target expression, and gene mutations downstream of the target gene itself lead to acquired resistance [7, 8, 9], which are the greatest challenge facing targeted therapies. Immune checkpoint inhibitors (ICIs) have entered the treatment regimen for metastatic breast cancer. Biomarkers such as PD-L1 expression and tumor infiltrating lymphocytes (TIL) are commonly used for immunotherapy, yet only a subset of patients benefit from ICIs and immune-based combination therapy [10]. Immunotherapy alone has been shown to increase the risk for early death compared with other treatments. Immunotherapy combined strategy can prevent early death [11]. Based on the combination of ICIs with each other or with other anticancer drugs, including systemic chemotherapy and poly ADP-ribose polymerase (PARP) inhibitors, targeted therapy has become a widely used strategy and new direction in cancer immunotherapy [12]. Identification of biomarkers predicting response to immune combinations is the key to progress [13]. Therefore, the discovery of novel diagnostic and therapeutic biomarkers could be a breakthrough in the treatment of the disease.
MCM6 is a key regulator of DNA replication and hence plays an undeniable role in maintaining the cell cycle [14]. MCM6 expression is upregulated in many cancerous cells, and the overexpression of MCM6 promotes the occurrence and development of carcinoma [15, 16, 17, 18]. Relevant studies proved that MCM6 regulates proliferation, immune response and maintenance of the DNA replication system and tumor cell proliferation and invasion and other physiological and pathological processes [19, 20]. MCM6 promotes metastasis though the MEK/ERK pathway and serves as a novel serum biomarker for early relapse in hepatocellular carcinoma [21]. MCM6 is highly expressed in tumors with higher histological grades, especially in clinically aggressive breast cancers (e.g., luminal B, HER2-positive, and triple-negative subtypes) [16]. Therefore, MCM6 might represent a sensitive and specific biomarker for cancer prognosis. Inhibition of MCM6 is likely to be an effective treatment. Nevertheless, the function and related mechanism are unclear in breast carcinoma.
This study aimed to explore the correlation between the expression level of MCM6 and clinicopathological features and to evaluate whether MCM6 can be used as a prognostic factor for breast cancer. Then, the related signaling pathways and chemotherapy sensitivity with MCM6 overexpression in patients with breast cancer were evaluated via gene set enrichment analysis (GSEA) and Genomics of Drug Sensitivity in Cancer (GDSC) database. Additionally, The Cancer Genome Atlas (TCGA) database was employed to determine the correlation between MCM6 and tumor-infiltrating immunocytes. The functional role of MCM6 was further investigated in vitro and in vivo in breast cancer cells. This study provides an important basis for further exploring the role of MCM6 in diseased cells and for targeted treatment of breast carcinoma.
The human breast cancer cell lines MCF-7, BT549, T47-D, MDA-MB-231, and HCC-1806
and the human breast epithelial cell line MCF-10A were obtained from
The Cell Bank of Chinese Academy of Sciences
and ATCC and American Type Culture Collection (ATCC), and cultured at 37
°C with 5% CO
TRIzol reagent (15596-026, Invitrogen, Carlsbad, CA, USA) was utilized for the
extraction of the aggregate RNA, which was reverse transcribed to cDNA according
to the requirements of the RevertAid First Strand cDNA Synthesis Kit (K1622,
Thermo Scientific, Waltham, MA, USA). PCR amplification was conducted using the
SYBR Green Select Master Mix System (4472908, Invitrogen, Carlsbad, CA, USA) on a
CFX96TM Real-Time PCR Detection System. The target mRNA expression was normalized
to the mRNA expression of GAPDH. The primers were as such follows: MCM6: forward,
5′-TGTCAGTGGTGTTGATGGATATG-3′, reverse
5′-GCTGTCTGTTCCTCATCTCTG-3′; GAPDH: forward,
5′-GGCAACAATATCAGCAACTTC-3′, reverse 5′-GGTCCTTAGGTGTGGTATC‑3′.
All qRT‒PCR experiments were conducted no less than three times. MCM6 expression
was examined via Western blotting. Briefly, the cells were lysed for 20 min using
RIPA buffer (89900, Thermo Fisher Scientific, Waltham, MA, USA), and all cell
proteins were extracted after centrifugation. The BCA Protein Assay Kit (23227,
Pierce, Waltham, MA, USA) was adopted for the quantification of cell lysate protein. The
protein solution was separated using SDS–PAGE and transferred to a
polyvinylidene fluoride (PVDF) membrane, which was treated with evaporated milk
(5%) at 4 °C for a night. The PVDF membrane was cultured with anti-MCM6
antibody (13347-2-AP, Proteintech, Rosemont, IL, USA) at 1:1000 at approximately
23 °C and treated with secondary antibody at a 1:10,000 dilution. The
internal control used was
IHC staining was performed in conformity with the manufacturer’s protocol (Dako
REAL EnVision Detection System; Dako, Santa Clara, CA, USA). To elaborate, the tissue segments
underwent a baking process at 68 °C for 2 h. The slices were dewaxed
with xylene, gradient alcohol and distilled water. Then, the slices were boiled
in citric acid buffer (pH 6.0, 0.01 M) for 2 min at high temperature and pressure
for antigen repair. The slices were incubated with 0.3% hydrogen peroxide to
block endogenous peroxidase activity. Subsequently, the slices were cultured with
MCM6 antibody (diluted 1:400; 13347-2-AP, Proteintech) at 4 °C overnight
and goat anti-mouse IgG-HRP antibody. Diaminobenzidine tetrahydrochloride (DAB)
was used to observe the reaction. IHC staining was scored by two independent
investigators. The aggregate score was obtained by multiplying the proportion of
positive tumor cells [scored as 0 (
The RNA-seq data and relevant information about the research subjects were acquired from the 2022 version of The Cancer Genome Atlas database (TCGA) (https://www.genome.gov/Funded-Programs-Projects/Cancer-Genome-Atlas) from the official website. The subjects were grouped into two groups according to expression level, namely, the median MCM6 mRNA expression of each sample. The clinicopathological data of the subjects were collected, including age, clinical classification of primary focus, regional lymph node and distant metastasis, represented by TNM collectively, endogenous molecular classification (Luminal A, Luminal B, basal like, HER2 overexpression or normal type) and histological type. The expression level of MCM6 protein in breast cancer patients was obtained in UALCAN database (http://ualcan.path.uab.edu/).
The online websites Kaplan‒Meier Plotter (http://kmplot.com/analysis/) and PrognoScan (http://dna00.bio.kyutech.ac.jp/PrognoScan/) were selected for the prognostic analysis of MCM6 in patients with breast carcinoma. The “rms” R package was used to combine MCM6 expression with clinicopathological features to construct a nomogram risk prediction model for the prediction of OS, and the predictive performance was measured using calibration curves and the C-index [22, 23].
The h.all.v7.5.symbols.gmt [Hallmarks] gene set database was selected. GSEA was deployed to elaborate the functional differences of the gene between the two expression groups, and the MCM6 expression level was considered a label of phenotype. The arrangements of gene sets were arranged out by 1000 times per every analysis. The False Discovery Rate (FDR) q value, the normalized enrichment score (NES) and the nominal p value were established to classify each phenotypic enrichment signaling pathway.
With the RNA-seq data, the MCM6-ICG relations to explore the immune genes
related to MCM6 expression were analyzed using the R package, which includes
limma, reshape2, ggplot2, ggpubr, and corrplot. The correlation threshold was set
to a correlation coefficient
Diseased cell proliferation was measured with a sulforhodamine B (SRB) assay. The cells were seeded in a 96-well plate (100 µL/well) and fixed with cold 10% trichloroacetic acid for half an hour at room temperature. The SRB solution (0.4%, Sigma S9012, Wappingers Falls, NY, USA) was treated with deionized water and then added for dyeing for approximately 10 min. Tris solution (10 mM) was incubated at 23 °C for 5 min, and the optical density (OD) of the cells was determined at 490–530 nm wavelengths [24].
The cell samples were collected using trypsin, fixed with 80% ethanol at 4 °C in a dark environment overnight, and then stained in the dark with propidium iodide (0.1 mg/mL, BD Biosciences, Franklin Lake, NJ, USA) and RNase A (1 mg/mL) for 30 minutes. The cell cycle of each group was tested via flow cytometry. The data were analyzed by FlowJo 7.6 (San Carlos, CA, USA) [25].
The collected cells were resuspended in binding buffer, cultured with Annexin V-FITC (BD Biosciences) for 10 minutes in a dark environment at room temperature and subsequently stained with binding buffer and PI (BD Biosciences) in darkness for 30 minutes at 4 °C. Cell measurement was conducted using flow cytometry [25].
The cells were inoculated into 6-well culture plates in complete medium until they were 80% confluence. The cells were scratched with a 200 µL pipette tip. Then the medium was changed to a serum-free medium. The migration process was carefully observed at 0, 24 and 48 h on an inverted microscope (Olympus, Tokyo, Japan). ImageJ (National institutes of Health, Bethesda, MD, USA) was employed to calculate the migration area [25].
The cell suspension was cultured in serum-free medium into the upper layer of the experimental plates coated with Matrigel (BD Biosciences). The serum-containing medium was placed in the lower layer. The cells that invaded the lower layer received methanol treatment and Giemsa staining and were photographed microscopically [26].
The animal experiments won approval from the authoritative organization of the
hospital. Six-week-old female BALB/C nude mice were maintained in a pathogen-free
environment. MDA-MB-231 cells (1
For data analysis, SPSS 22.0 software (IBM Corp., Armonk, NY, USA) was adopted.
The expression difference between breast tumor and normal breast tissues was
analyzed by the Wilcoxon signed-rank test. Univariate and multivariate Cox
analyses were selected to analyze the influence of MCM6 expression along with
other factors on mouse survival. Logistic regression analysis was performed to
determine how MCM6 was related to other clinical factors. One-way analysis of
variance was employed to make comparisons between two groups. p
We first explored MCM6 expression in a wide range of human malignancies. The mRNA expression of MCM6 increased in various cancers from TCGA database analysis (Fig. 1A,B), and MCM6 expression in breast cancer considerably exceeded that in normal tissues (Fig. 1C,D). The protein expression level of MCM6 in patients with breast cancer outstripped that in normal tissues from the online database UALCAN (http://ualcan.path.uab.edu/) (Fig. 1E). Consistent with the expression pattern from the online database, immunohistochemical staining outcomes indicated higher MCM6 expression in diseased tissues than in adjacent breast tissue. Among 100 cases of breast cancer tissues, 92 cases (92.0%) had overexpression of MCM6, and the rest (8.0%) had low expression of MCM6, while there were 45 cases (45.0%) with overexpression of MCM6 and 55 cases (55.0%) with low expression of MCM6 in adjacent tissues (Fig. 2A–C). In comparison with MCF-10A human breast epithelial cells, MCM6 expression in breast cancer cell lines (MCF-7, BT549, T470, MDA-MB-231, HCC1806) was significantly increased (Fig. 2D,E). These results suggest that MCM6 was overexpressed in patients with breast carcinoma.
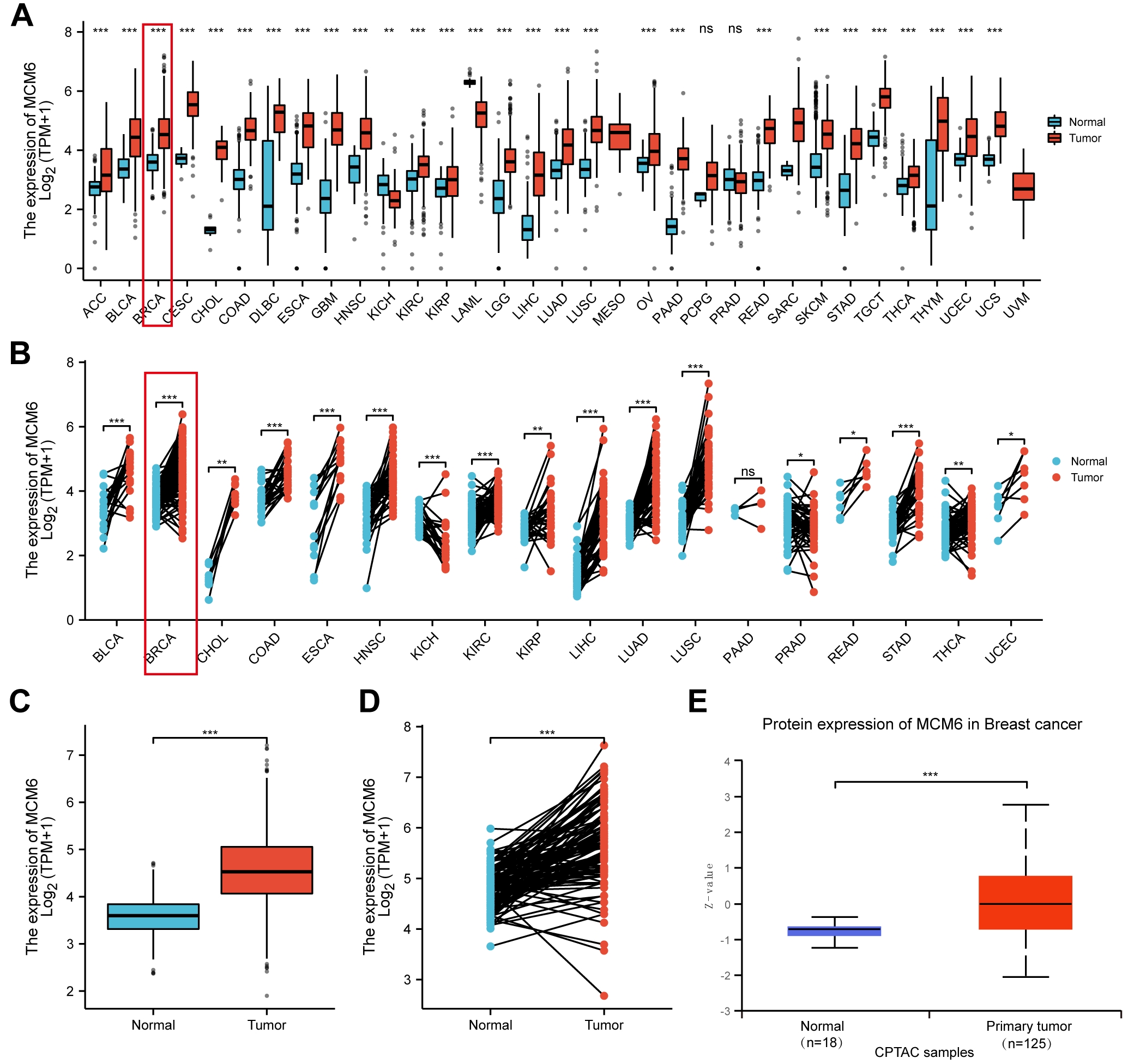 Fig. 1.
Fig. 1. MCM6 was significantly upregulated in breast cancer
based on database analysis. (A,B) MCM6 expression was increased in a variety of
tumors according to The Cancer Genome Atlas (TCGA) database analysis. (C) Box plot of TCGA database
describes the different expression levels of MCM6 between normal tissues and
cancerous tissues. (D) Based on the same number of samples, MCM6 expression in
normal and diseased tissues was compared. (E) MCM6 protein expression was higher
in normal breast tissue according to the UALCAN database. *p
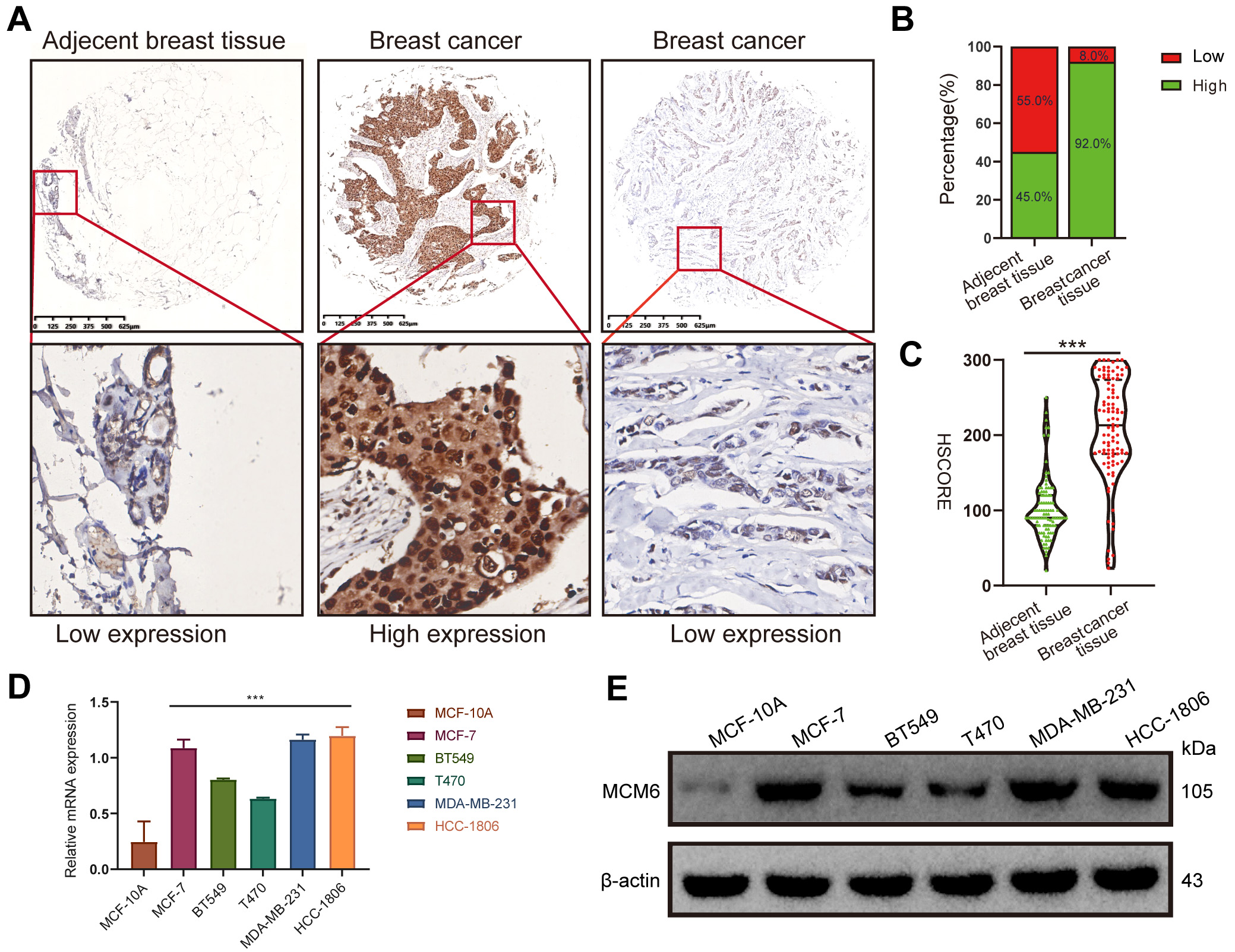 Fig. 2.
Fig. 2.MCM6 is significantly up-regulated in tissues and cells of
breast cancer based on experimental verification. (A–C) Immunohistochemical
staining of MCM6 expression in adjacent breast tissue and breast cancer tissues,
representative immunohistochemical images and statistical results (n = 100).
(D,E) Quantitative Real-Time PCR (qRT‒PCR) and western blotting analyses of MCM6 expression in MCF-10A breast
cells and diseased cells, including MCF-7, BT549, T470, MDA-MB-231, and HCC1806
breast cancer cells. Statistics are presented as the means
We further explored how MCM6 expression is correlated with clinicopathologic parameters (age, histological grade, size of tumor, lymph node metastasis, vascular invasion, distant metastasis, and molecular typing). The outcomes demonstrated that MCM6 expression was associated with the size of tumor (p = 0.001) and lymph node metastases (p = 0.012) (Table 1). An analysis of the TCGA database revealed that the expression of MCM6 varied according to age, and there were differences between Asians and Caucasians White (Fig. 3A,B); MCM6 expression in estrogen receptors (ER) and progesterone receptors (PR)-negative breast cancer patients was higher than that in ER- and PR-positive breast cancer patients (Fig. 3C,D); MCM6 expression in HER2-positive subjects was more obvious than in negative breast cancer patients (Fig. 3E); MCM6 expression was elevated in luminal B, basal and HER2 breast cancer (Fig. 3F); MCM6 expression in invasive lobular carcinoma was higher than that in invasive lobular carcinoma tube carcinoma (Fig. 3G); no notable difference in the expression of MCM6 was found in each clinical stage (Fig. 3H); MCM6 expression was different only in the T1 and T2 stages; no considerable difference was shown in MCM6 expression between the N stage and the M stage, which T represents tumor size stage, N represents lymph node metastasis stage, and M represents distant metastasis stage (Fig. 3I–K). Based on the analysis from the PrognoScan online website, breast cancer patients with high MCM6 expression had reduced OS, relapse-free survival (RFS) and distant metastasis-free survival (DMFS) (Fig. 4A–C), which is consistent with the results of the Kaplan-Meier Plot online prognostic analysis (Fig. 4D–F).
| Characteristics | MCM6 expression | p value | ||
| Low (%) | High (%) | |||
| Age | 0.309 | |||
| 7 (63.6) | 69 (77.5) | |||
| 4 (36.4) | 20 (22.4) | |||
| History grade | 0.427 | |||
| 1 | 1 (9.1) | 2 (2.2) | ||
| 2 | 8 (72.7) | 65 (73.0) | ||
| 3 | 2 (18.2) | 22 (24.7) | ||
| Tumor diameter | 0.001 | |||
| 7 (63.6) | 14 (15.7) | |||
| 3 (27.3) | 58 (65.2) | |||
| 1 (9.1) | 17 (19.1) | |||
| Lymph node metastasis | 0.012 | |||
| With | 8 (72.7) | 30 (33.7) | ||
| Without | 3 (27.3) | 59 (66.3) | ||
| Vascular invasion | 0.547 | |||
| With | 6 (54.5) | 40 (44.9) | ||
| Without | 5 (45.5) | 49 (55.1) | ||
| Distant Metastasis | 0.074 | |||
| With | 10 (90.9) | 57 (64.0) | ||
| Without | 1 (9.1) | 32 (36.0) | ||
| Surrogate subtypes | 0.836 | |||
| Luminal A-like | 2 (18.2) | 9 (10.1) | ||
| Luminal B-like | 4 (36.4) | 42 (47.2) | ||
| Her-2+ | 3 (27.3) | 22 (24.7) | ||
| Triple negative breast cancer | 2 (18.2) | 16 (18.0) | ||
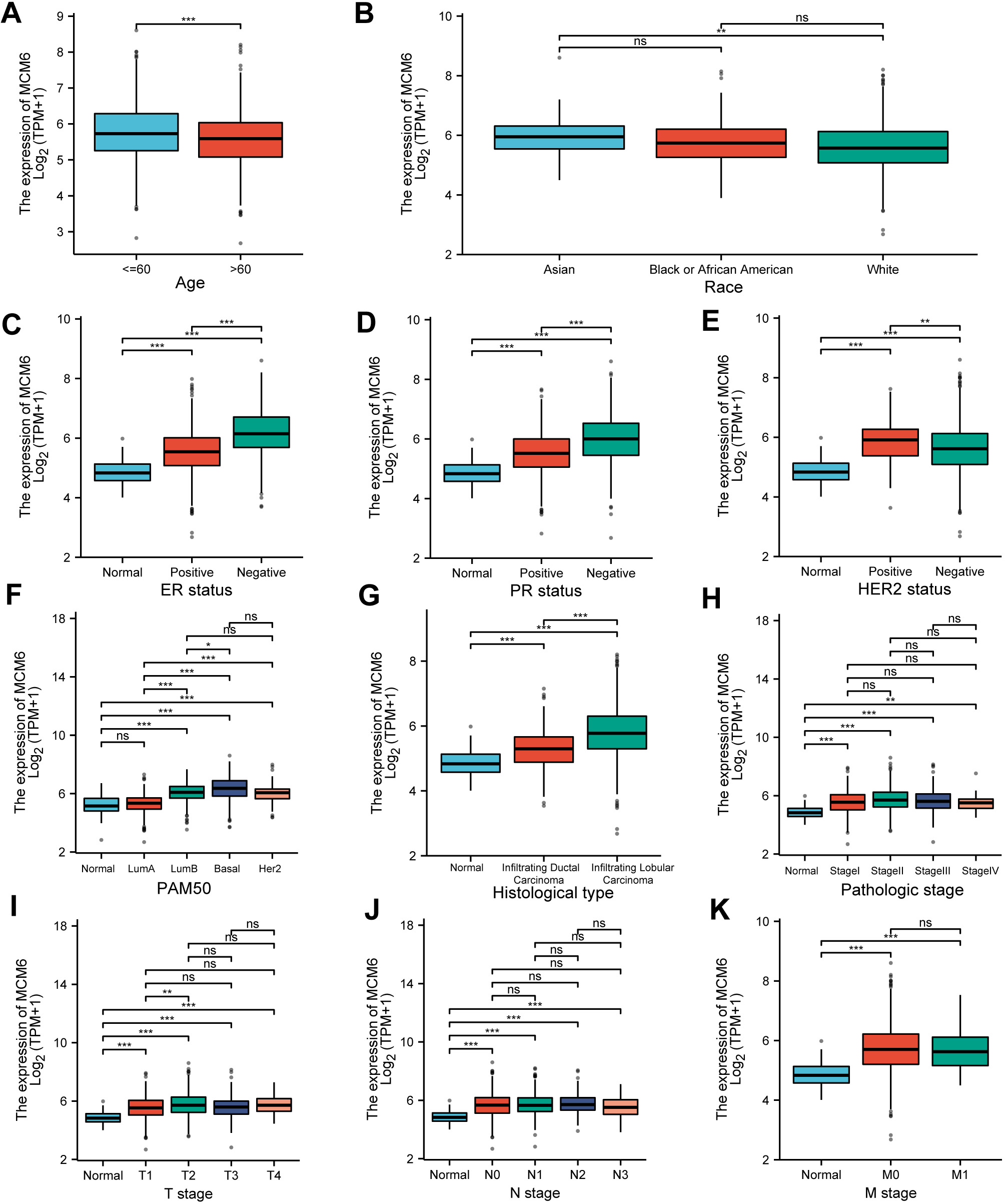 Fig. 3.
Fig. 3.Expression of MCM6 in different clinicopathological
features. (A) MCM6 expression in people
under 60 years old and over 60 years old. (B) MCM6 expression in Asian, Black or
African American and White individuals. (C–E) Relationship between MCM6
expression and Estrogen Receptors (ER), Progesterone Receptors (PR), and Human Epidermal Growth Factor Receptors 2 (HER2) status. (F) Relationship between MCM6 expression
and different cancerous types of molecules. (G) Relationship between MCM6
expression and different histological subtypes of breast cancer. (H) MCM6
expression in different pathologic stages. (I–K) MCM6 expression in the T, N and
M phases. Statistics are presented as the means
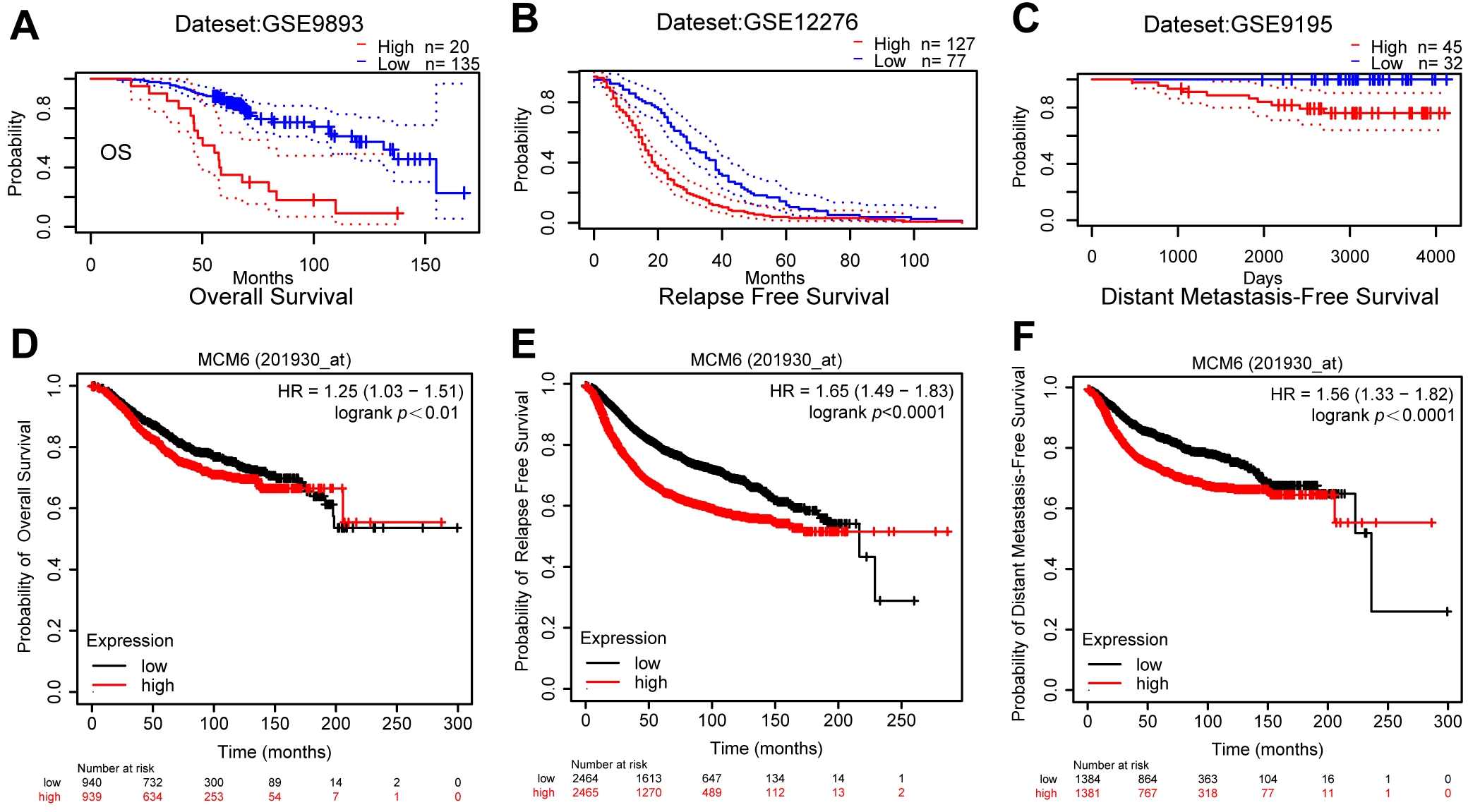 Fig. 4.
Fig. 4.Breast cancer with high MCM6 expression has a poor prognosis. The PrognoScan online website was selected to analyze the prognostic role of MCM6 in breast carcinoma, and (A), (B), and (C) represent Overall Survival (OS), Relapse Free Survival (RFS) and Distant Metastasis Free Survival (DMFS), respectively. The Kaplan Meier Plotter online website was also utilized for the analysis of the prognostic effects of MCM6 in breast cancer, and the results suggested reduced survival time in patients with overexpression of MCM6 in terms of all three indicators mentioned above, and (D), (E), and (F) represent OS, RFS and DMFS, respectively.
In addition, univariate Cox regression analysis revealed that the expression of ER, PR, stage, T stage, M stage, N stage and MCM6 was associated with a low survival rate of patients (Fig. 5A). In multivariate Cox regression, after adjusting for other confounding factors, the expression of age, ER, M, N and MCM6 was an independent indicator (Fig. 5B). Furthermore, various indicators (age, ER, M, N and MCM6 expression) were included in the nomogram prediction model by multivariate Cox regression. The total score was obtained by age, expression of ER, M stage, N stage, MCM6, and prediction of survival probabilities at three years and five years (Fig. 5C). The C-index for the prediction nomogram was 0.817. Calibration curves displayed the predictive power of the predictive model in terms of survival time in patients with breast carcinoma (Fig. 5D,E). These results suggest that overexpression of MCM6 is a dismal prognostic factor for breast cancer.
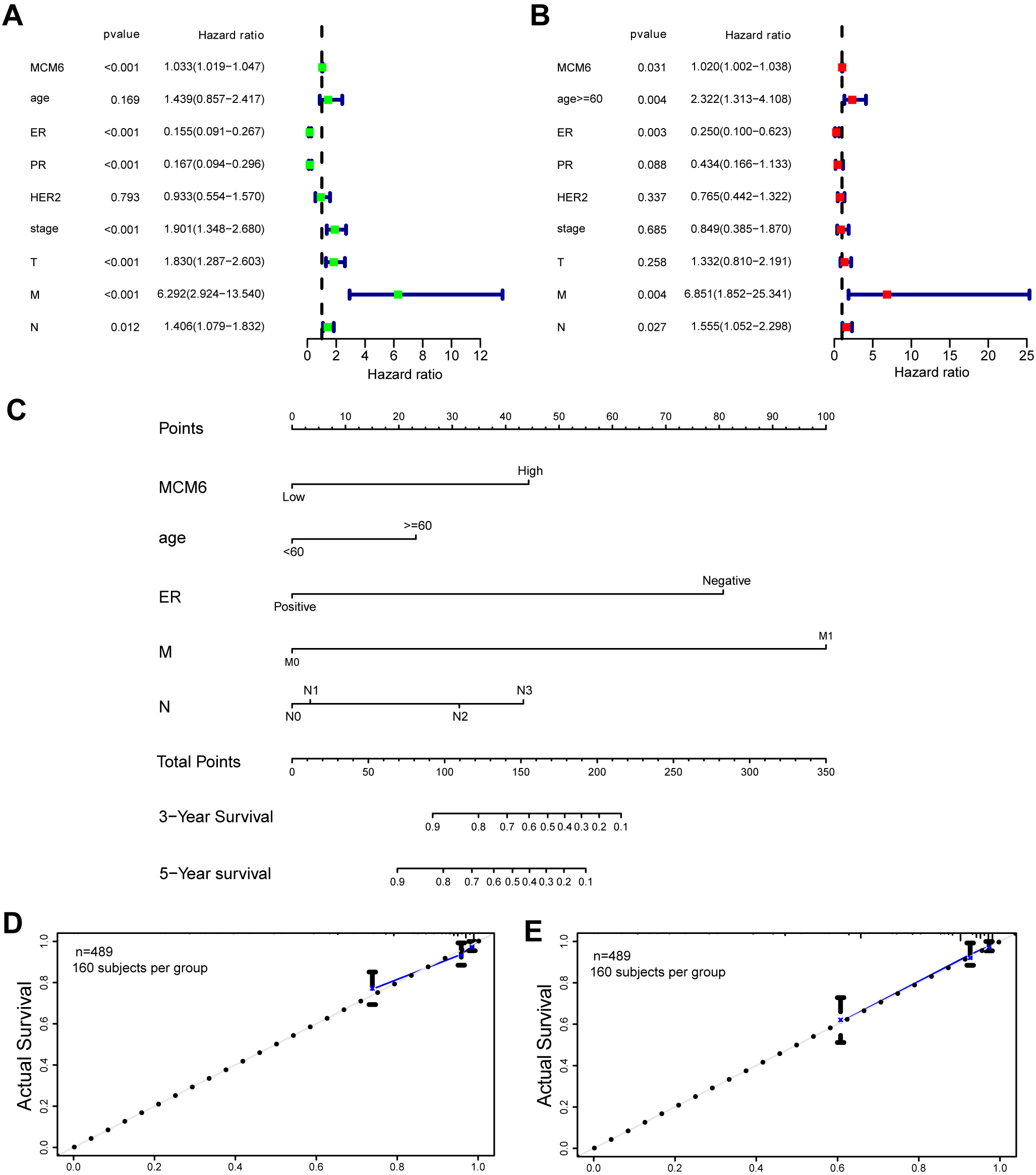 Fig. 5.
Fig. 5.Univariate and multivariable Cox regression analysis and construction of nomograms related to patient prognosis. (A) The prognosis of MCM6 was analyzed by univariate Cox regression. (B) Multivariate Cox regression analysis of the prognosis of MCM6. (C) The nomogram combined with the risk signature and clinicopathological factors. (D,E) Calibration plots for the prediction of prolonged survival of three and five years.
GSEA was performed between subgroups with different levels of MCM6 expression using the whole gene network based on the TCGA dataset with a view to explore the biological pathways via which MCM6 works in diseased tissues. The biological processes involved mainly included DNA repair, different pathways of IL6-JAK-STAT3, MTORC1, PI3K-AKT-MTOR, MYC, etc. (Fig. 6A). KEGG analysis demonstrated that overexpression of MCM6 was mainly involved in the process of mismatch repair, the regulation of ubiquitination proteolysis, ERBB signaling pathway and p53 signaling pathway. In addition, MCM6 is enriched in immune signaling pathways and activates related immune responses, such as B cell and T cell receptor pathways, and cytotoxic effects of natural killer cells (Fig. 6B).
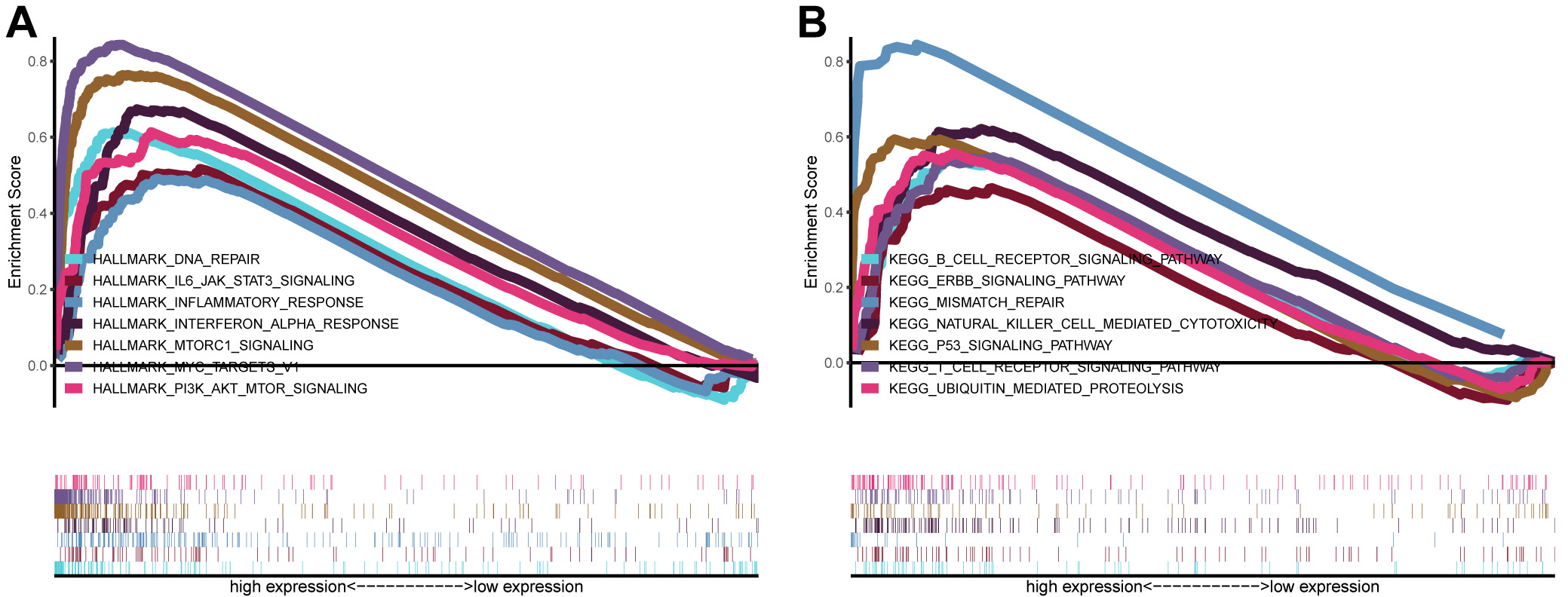 Fig. 6.
Fig. 6.Analysis of signaling pathways by using GSEA. (A) GSEA plots showing the NES for HALLMARK, which had close relations to the overexpression of MTM6 in breast carcinoma and involved signaling pathways, including DNA repair, signaling types of IL6 JAK STAT3, MTORC1 and PI3K AKT MTOR, inflammatory and interferon alpha responses, and MYC targets V1. (B) NES for KEGG, which has a close association with the overexpression of MTM6, including B-cell receptor, ERBB, p53, T-cell receptor signaling pathways, mismatch repair, natural killer cell-mediated cytotoxicity, and ubiquitin-mediated proteolysis.
The impact of MCM6 expression on chemosensitivity in patients with breast
carcinoma were assessed via the pRRophetic algorithm and the half-maximal
inhibitory concentration available in the Genomics of Drug Sensitivity in Cancer
(GDSC) database. The results demonstrated that 134 chemotherapeutic drugs were
identified to be significantly correlated with MCM6 expression. In the group
overexpressing MCM6, we found that there were 14 chemotherapy drugs with
significant effects (Fig. 7A–N), especially PD-0332991 (p = 3.7
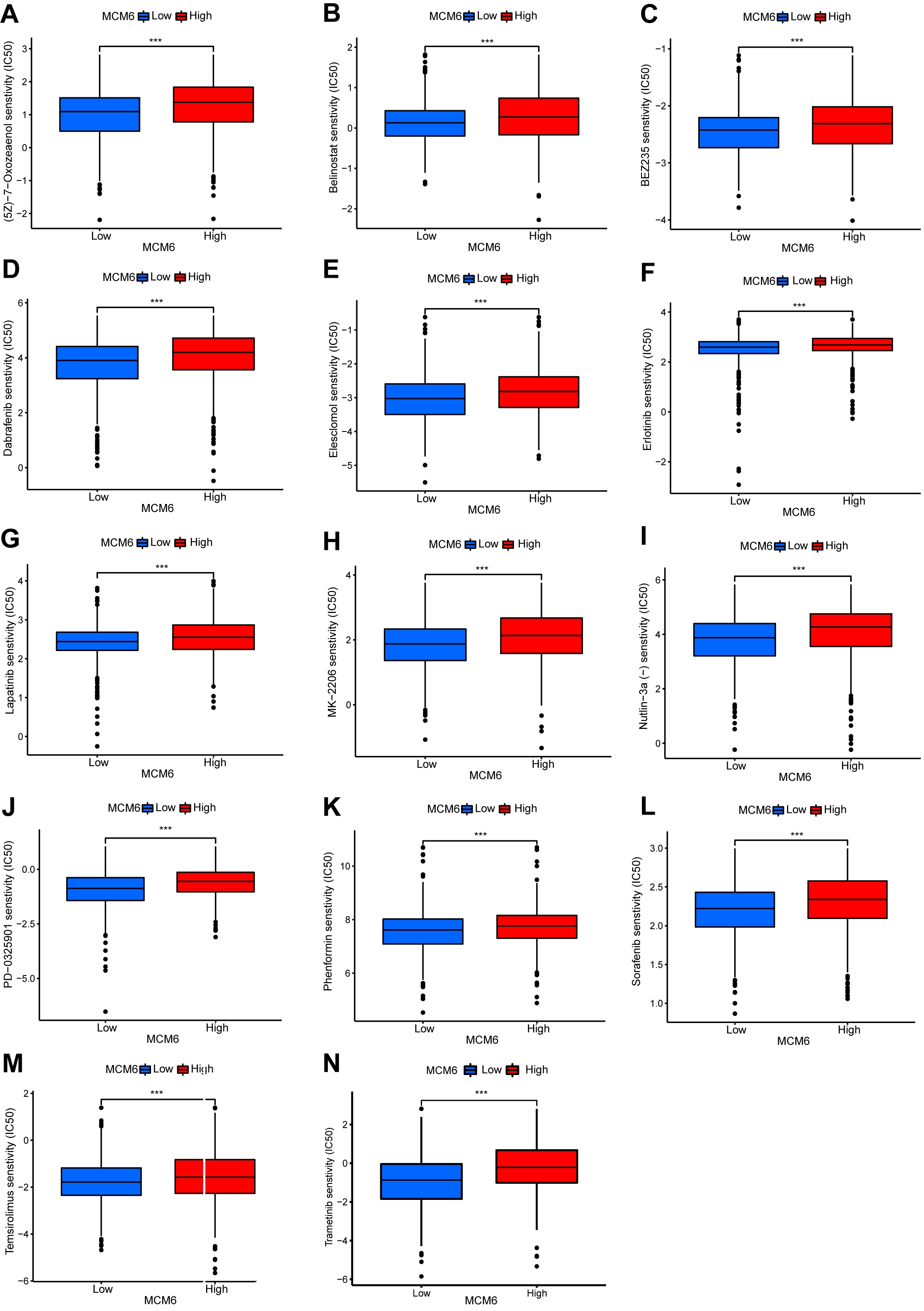 Fig. 7.
Fig. 7.MCM6 expression correlates with the chemosensitivity of breast
cancer. Box plots show the chemosensitivity of (5Z) –7–oxozeaenol (A),
belinostat (B), BEZ235 (C), dabrafenib (D), elesclomol (E), erlotinib (F),
lapatinib (G), MK–2206 (H), Nutlin–3a (I), PD-0325901 (J), phenformin (K),
sorafenib (L), temsirolimus (M) and trametinib (N) in high- and low-expression of
MCM6 in breast cancer. ***p
The expression correlation between MCM6 and genes related to immune detection
points was analyzed based on the TCGA BRCA database. Our research outcomes
indicated a positive correlation between MCM6 and the expression of multiple
immune checkpoints and a negative correlation with that of BTNL9 (Fig. 8A).
Analysis of the relationship between MCM6 and immunity, including 24 kinds of
immune cells, showed a significantly negative correlation with eight immune cells
(iDC, CD8 T cell, Th17 cells, eosinophils, mast cells, NK CD56 bright cells, NK
cells, pDC) but a significantly positive correlation with fifteen immune cells
(cells of Th2, aDC, Th1, Treg, T helper, NK CD56dim, Marcophages, B, Tcm, DC,
Tgd, T, Term, Neutrophils, TFH) (Fig. 8B). In addition, MCM6 expression was
positively correlated with TMB (R = 0.33, p
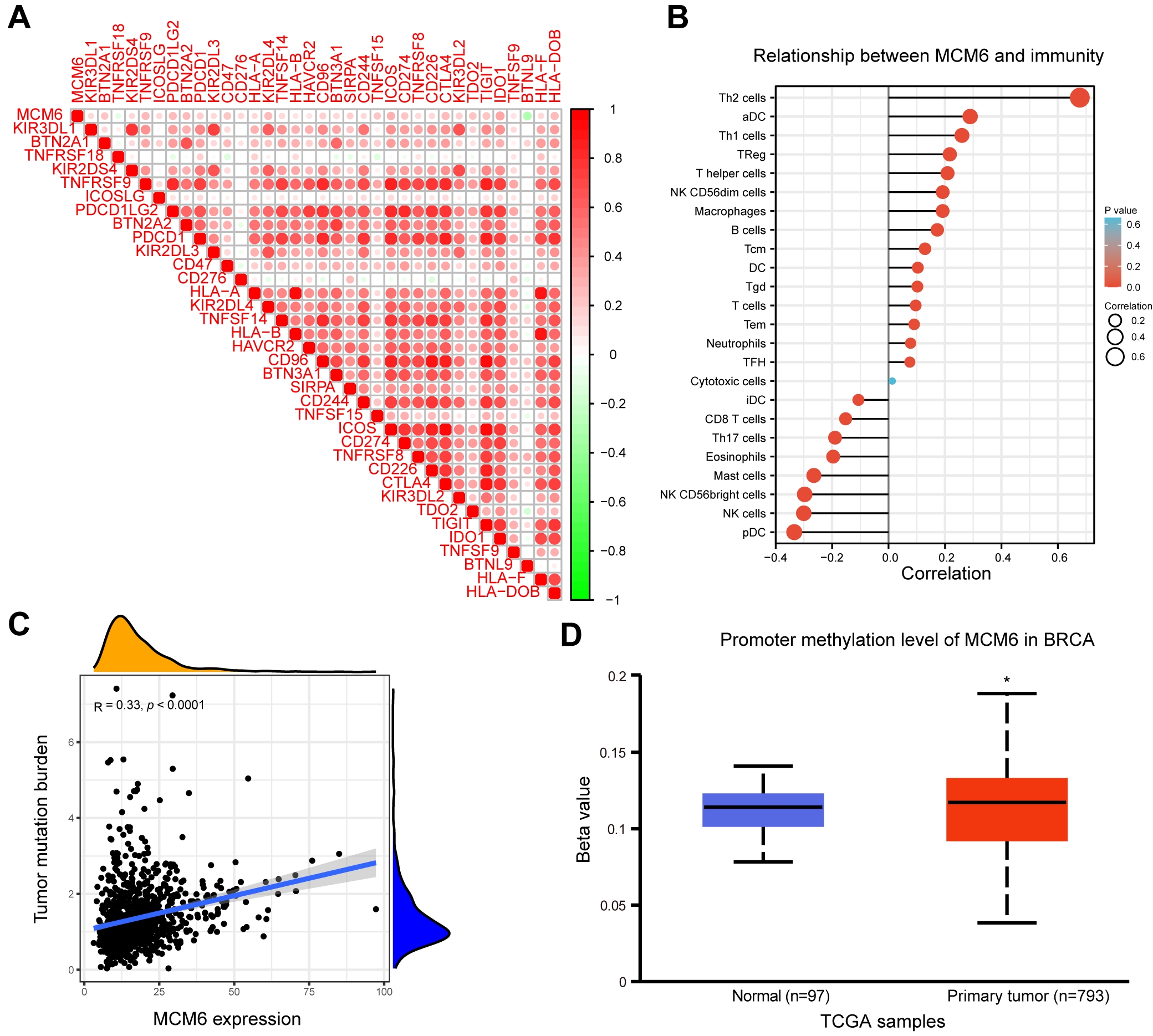 Fig. 8.
Fig. 8.Correlation analysis of MCM6 and immune checkpoint-related gene expression, immune cells, TMB and methylation. (A) TCGA BRCA database was used in correlational analysis of MCM6 and the expression of immune checkpoint-related genes, and the correlation coefficient threshold was set at 0.2. (B) Correlation analysis of MCM6 expression and immune cell infiltration. (C) Correlation analysis of tumor mutation burden (TMB) with MCM6 expression. (D) TCGA Breast Invasive Carcinoma (BRCA) database was used to analyze the promoter methylation level of MCM6 between normal tissues and diseased tissues.
To make a clarification of the biological role of MCM6 in the progression and prognosis of breast carcinoma, the lentivirus system was used to conduct stable MCM6 knockdown in MDA-MB-231 and MCF-7 cancerous cells (shMCM6), by which the mRNA and protein expression of MCM6 was considerably down-regulated (Fig. 9A,B). MCM6 down-regulation prevents the growth of cancerous cells (Fig. 9C), lead to a higher percentage of apoptotic cells (Fig. 9D). MCM6 silencing caused cancerous cells to reproduce in large numbers in G1 phase and reduced cancerous cells in S phase (Fig. 9E). MCM6 knockdown also reduced the migrating and invasive power of diseased cells (Fig. 9F,G). To test whether MCM6 promoted breast tumor growth in vivo, a subcutaneous transplantation tumor model of BALB/c nude mice was constructed with MDA-MB-231 breast cancer cells (Fig. 10A). The volume and weight of the xenografts of MDA-MB-231 cells with MCM6 knockdown were lighter than those of the control group (Fig. 10B,C). The quantity of ki-67-positive cells in the xenograft tumor with MCM6 knockdown was decreased compared with that in the control group, consistent with MCM6 expression (Fig. 10D). These results suggested that MCM6 knockdown can successfully retrain the growth of cancerous cells both in vivo and in vitro.
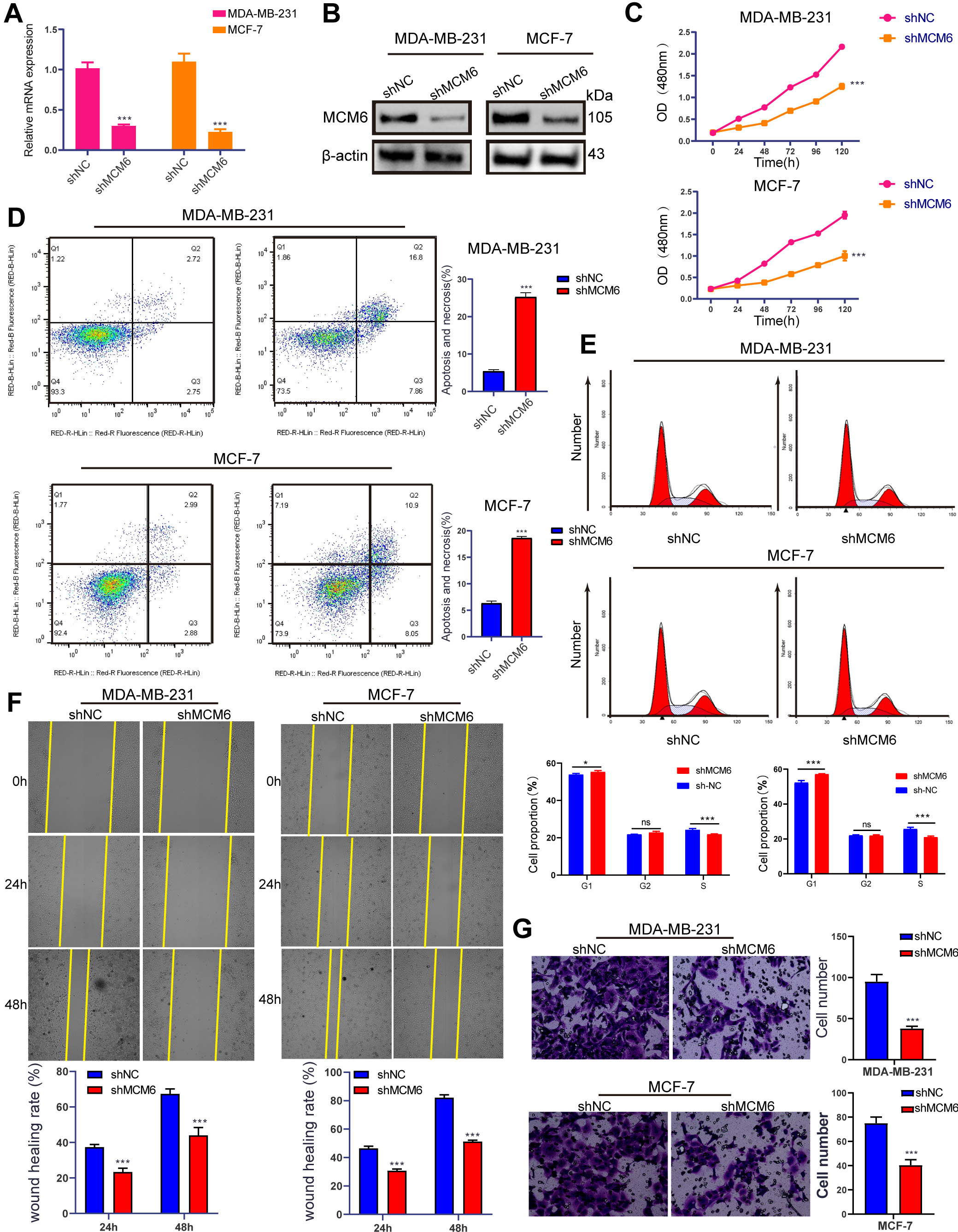 Fig. 9.
Fig. 9.MCM6 promotes breast cancer cell growth, migration, and invasion
and inhibits apoptosis in vitro. (A,B) qRT‒PCR and WB showing MCM6
expression in stably silenced in MDA-MB-231 and MCF-7 cells.
(C) CCK8 experiment results show that inhibition of MCM6
significantly inhibits the proliferation of MDA-MB-231 and MCF-7 cells. (D)
Inhibition of MCM6 significantly promotes the
apoptosis rate of MDA-MB-231 and MCF-7cells. (E) Knockdown MCM6
leads to a large number of G1 stage cancer cells to multiply, and S stage cancer
cells to reduce. (F) Wounding healing assay showed that knockdown of MCM6
significantly slowed down the wound healing rate of MDA-MB-231 and MCF-7cells.
(G) Invasion chamber experiment confirmed that knockdown of MCM6 significantly
inhibited the invasion rate of MDA-MB-231 and MCF-7cells. Statistics are
presented as the means
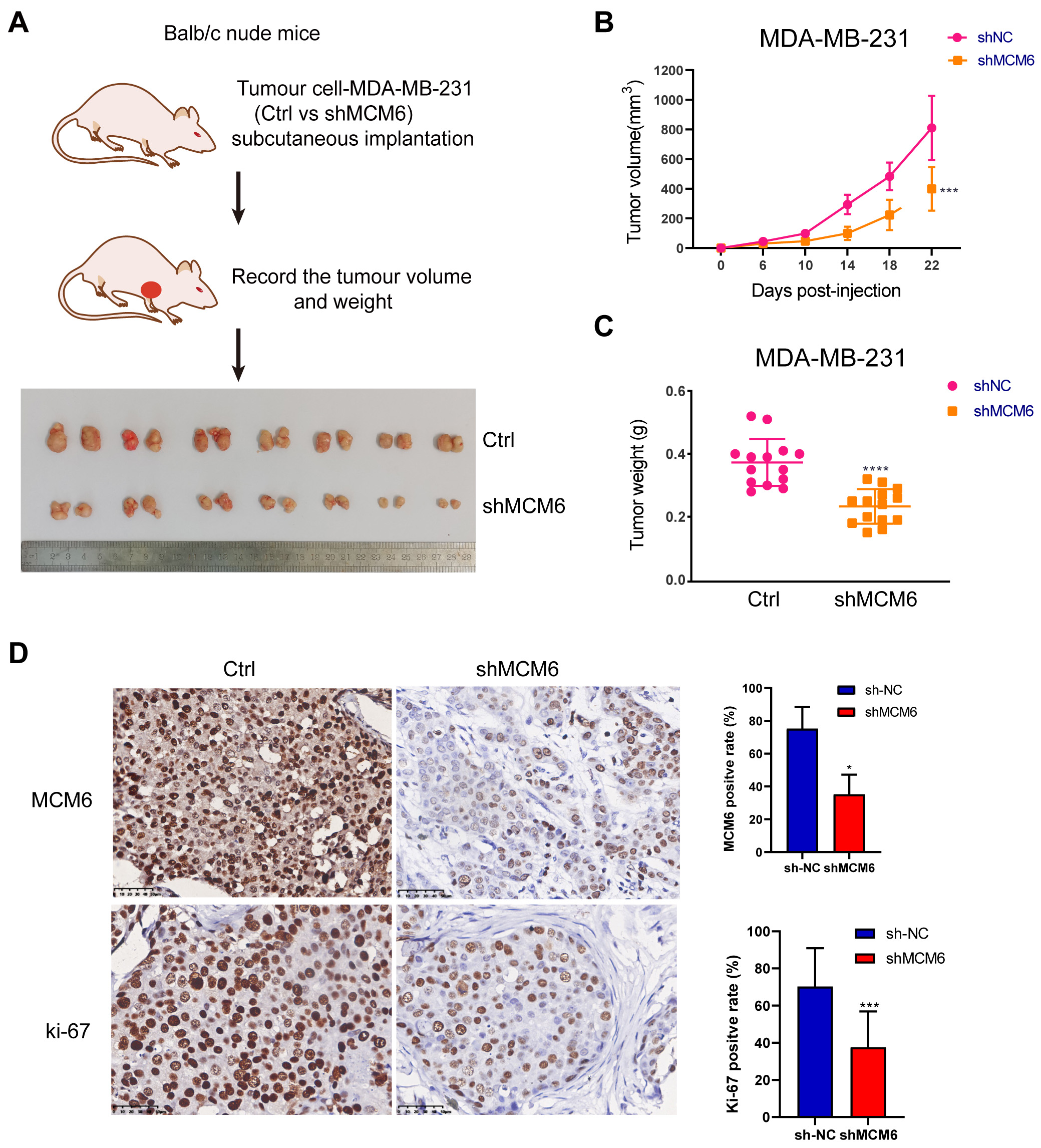 Fig. 10.
Fig. 10.CKAP4 promotes breast cancer cell growth in vivo.
(A) Schematic diagram of the nude mouse xenograft model and
all the tumors. (B,C) Knock down MCM6 significantly inhibits the growth of
transplanted tumor in nude mice. (D) The immunohistochemical results showed that
the expression of MCM6 was significantly decreased in the knockdown group, and
the positive rate of KI67 in the knockdown group was significantly lower than
that in the control group. Statistics are presented as the means
Breast cancer prevails in the world’s female population, with increasing incidence and a trend toward younger age year by year [2]. In recent years, targeted therapy, including trastuzumab, pertuzumab, lapatinib and lenatinib for HER2 treatment; bevacizumab, an antiangiogenic drug; PARP inhibitors iniparib, veliparib and olaparib for BRCA1/2 mutation, etc., has significantly prolonged life expectancy and quality of life in patients [26, 27, 28]. Although the mentioned drugs have achieved good clinical efficacy, they are only targeted at patients with specific molecular changes. There are still many breast cancer patients with poor efficacy because of drug resistance and other problems. Breast cancer is a very heterogeneous tumor, and its occurrence and development involve many gene changes. Hence, it is imminent to explore new biological markers and potential targets of breast cancer to provide a new basis for prognostic effect management and the development of targeted medication.
This research suggested that MCM6 expression was increased in a variety of tumors, including breast cancer, combined with TCGA database, clinical samples and breast cancer cells. Our conclusions are supported by Issac et al. [16]. Furthermore, MCM6 expression in luminal B, basal and HER2 breast cancer was significantly increased, and that in invasive lobular carcinoma was higher than that in invasive ductal carcinoma, which indicated that the role and prognostic value of MCM6 in different types of breast cancer may be different and that the mechanism needs to be further investigated.
In recent years, studies on biomarkers, prognostic markers and prognostic models of breast cancer have attracted increasing attention and can provide a good basis for the prognosis of breast cancer. We constructed a nomogram prediction model that included five factors (age, expression of ER, M stage, N stage and expression of MCM6) by multivariate Cox regression analysis, and the research results showed that low expression of MCM6 may indicate longer survival time for patients with breast cancer in our prognostic model. Relevant studies reported that excessive expression of MCM6 was related to poor prognostic effect. High expression of MCM6 mRNA was significantly associated with shorter survival time of patients with breast cancer [29]. Low expression of MCM6 mRNA is associated with increased recurrence-free survival in breast cancer patients [16]. The expression level of MCM6 in high-grade tumors is significantly increased and correlated with mitotic count, which is an effective reference index for tumor grading [30]. These results agreed closely with the outcomes in the model we constructed and suggested that overexpression was independent as a prognostic factor for breast carcinoma.
An increasing number of studies have shown that the role of MCM6 in tumor progression is extensive [14]. MCM6 silencing leads to arrest of S to G1 phase progression by reducing CDK2, CDK2, cyclin A, cyclin B4, cyclin D1 and cyclin E in hepatocellular carcinoma [31]. MCM6 promotes the metastasis of hepatocellular carcinoma through the MEK/ERK pathway [21]. CDK5RAP3 reduces MCM6 nuclear translocation, thereby reducing gastric cancer proliferation [32]. Liu et al. [33] found that SIX1 is an oncoprotein in cervical cancer that promotes proliferation by promoting the expression of MCM6. Boulay et al. [34] found that ARF1 promotes breast cancer proliferation by enhancing the expression of MCM6 and up-regulating the activity of pRB/E2F1. Nevertheless, the function of MCM6 in breast carcinoma remains unclear. The results of this study showed that MCM6 promoted breast cancer cell proliferation in vitro and in vivo, and promoted breast cancer migration and invasion in vitro. However, the biological pathway by which MCM6 plays a role in breast cancer remains unknown. By using GSEA, the study clarified the fact that overexpression of MCM6 promotes tumor progression mainly through the JAK-STAT3, mTOR myc, PI3K-AKT ERBB and p53 signaling pathways, which are closely related to tumorigenesis. Liu et al. [21] reported that MCM6 activates the MEK-ERK signaling pathway to promote the metastasis of liver cancer. We found that MCM6 overexpression is also related to the regulation of ubiquitination proteolysis, and whether MCM6 is involved in these signaling pathways has not been reported. Notably, MCM6 is involved in mismatch repair, which is consistent with the findings of Sherman et al. [35] reported that MCM6 was an important regulatory factor in the DNA replication system through complex formation with other MCM6 proteins to regulate DNA replication of the cell cycle in the carcinogenic process. In addition, our results indicated that the increased MCM6 expression had a bearing on cytotoxic effects of B-cell and T-cell receptor pathways and natural killer cells. Recent studies have shown that MCM6 functions to obstruct immune progression by reducing the immune response in microglial cells [14]. Therefore, MCM6 may be the key mediator of the immune response. We analyzed the relationship between MCM6 expression and immune checkpoint-related genes and immune cells. The results demonstrated that MCM6 displayed positive relevance with many immune checkpoint-related genes and negative correlation with BTNL9 in breast cancer, which has not been reported before and is worthy of further exploration. Whether MCM6 can affect the function of these immune checkpoints in breast cancer needs further study.
Given that MCM6 is key to the progression of breast carcinoma, we further explored the correlation between MCM6 and breast cancer-related chemotherapy drugs for therapy. Two potential drugs, PD-0332991 and trametinib, which are more sensitive when MCM6 is highly expressed, were screened out from breast cancer-related chemotherapy drugs. PD-0332991 is an inhibitor of CDK4 and CDK6 with oral activity and has antiproliferative activity on cancer cells, inducing their cell cycle arrest [36]. In addition, PD-0332991 guided the apoptosis of diffuse large B-cell lymphoma cells in a dose-dependent manner [37]. Studies have shown that PD-0332991 can be used for hormone (ER/PR)-positive and HER2-negative breast cancer [38]. Trametinib is an effective reversibility inhibitor of MEK 1/2 that restrains cell proliferation by blocking the MAPK pathway [39]. The FDA has approved trametinib combined with dabrafenib for patients with metastatic melanoma who have the BRAF V600E/K mutation [40]. Our results indicated that keener sensitivity to PD-0332991 and trametinib can be found in patients with overexpression of MCM6, which can be used as a monitoring indicator of drug sensitivity in breast cancer. However, our screening results still need to be verified by a large number of clinical cases and related experiments.
In conclusion, our research shows that overexpression of MCM6 can be found in patients with breast carcinoma and used as a poor prognostic factor for the disease. MCM6 can promote the proliferation of cancerous cells in vitro and in vivo. Based on database and bioinformatics analyses, MCM6 plays a crucial role in tumor immunity, tumor mutation burden and methylation through a variety of biological pathways and is related to drug sensitivity (Fig. 11). MCM6 can be used as a poor prognostic marker of breast cancer and a potential target for immunotherapy and targeted therapy. However, in our experiments, we have not further verified the biological pathways of MCM6 in breast cancer and the role and mechanism of MCM6 in tumor mutation burden and tumor immunity, and further studies are needed. In the future, comprehensive individualized treatment with chemotherapy combined with targeted therapy and immunotherapy will become the main treatment option for breast cancer patients. Our preliminary study showed that the high expression of MCM6 in breast cancer increases the sensitivity of related drugs. Targeting MCM6 and combining with related sensitive drugs is a promising research direction. Therefore, our study demonstrates that MCM6 has a potential oncogenic role in breast cancer and provides a new direction for targeted therapy and immunotherapy in breast cancer patients.
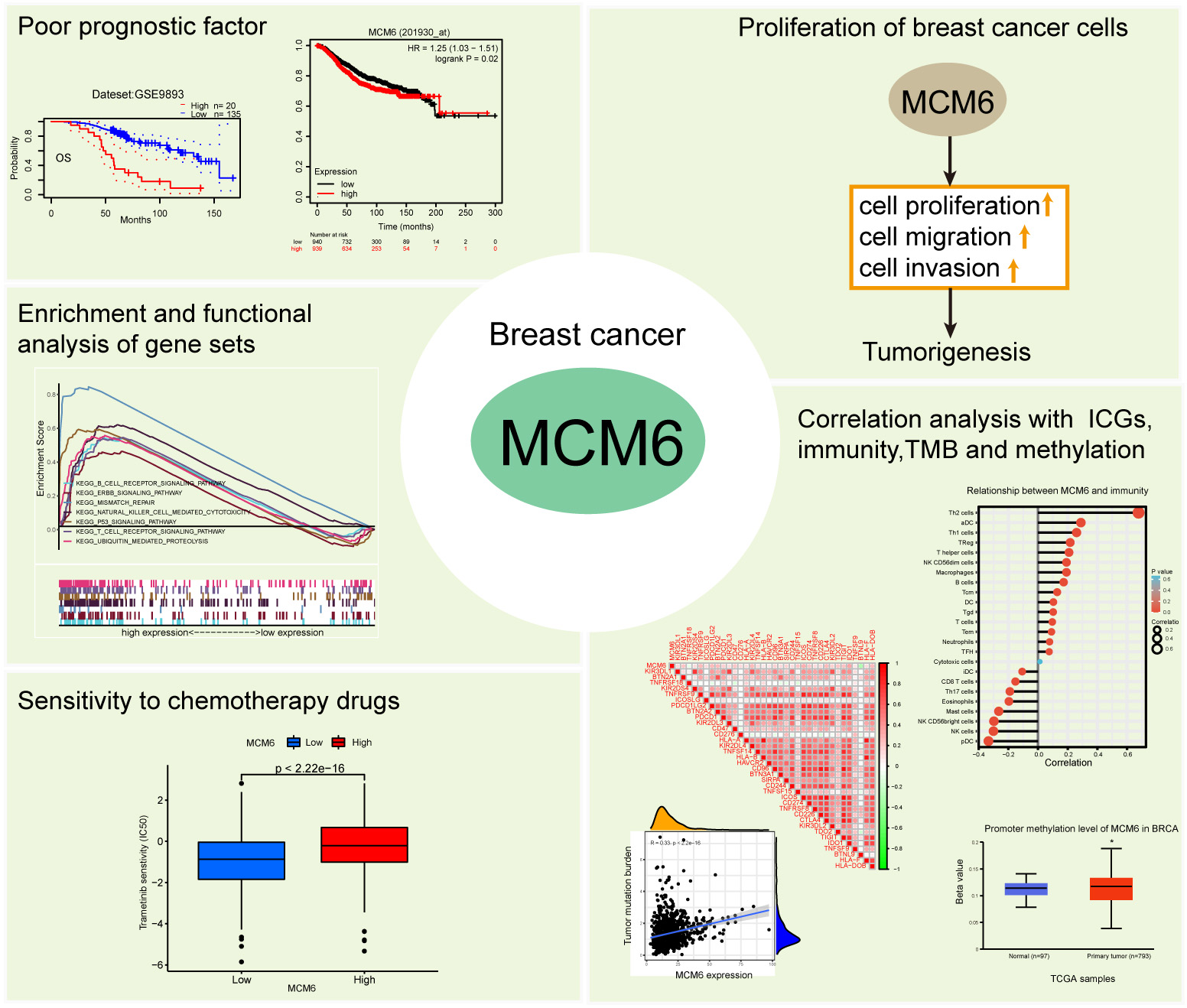 Fig. 11.
Fig. 11.MCM6 is a poor prognostic biomarker and promotes progression in breast cancer. Overexpression of MCM6 is poor prognostic biomarker in breast cancer. MCM6 can promote the proliferation of cancerous cells in vitro and in vivo. Based on database and bioinformatics analyses, MCM6 plays a crucial role in tumor immunity, tumor mutation burden and methylation through a variety of biological pathways and is related to drug sensitivity.
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
ZL and GP designed the study. PW, DJ, and LL analyzed the data. ZL, YW, and YY performed the experiments. ZL, PW, and GP contributed to the writing of the manuscript. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work
This study was supported by the Ethics Committee of the First Affiliated Hospital of Kunming Medical University (kmmu20220767). Specimens were handled in accordance with legal and ethical regulations.
Not applicable.
This study was supported by National Natural Science Foundation of China (82260512), Basic Research Program of Yunnan Province (202201AS070076, 202201AT070280), Joint Projects of Applied Basic Research of Kunming Medical University and Yunnan Province Department of science and Technology (202101AY070001-016), Science and Technology Innovation team of Education Department of Yunnan Province (K1322121), and High-level personnel training program of Yunnan Province (RLMY20200016).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.