
 , Vishnu D. Rajput 2,*
, Vishnu D. Rajput 2,* , Divya Pandey 3, Ragini Sharma 4, Karen Ghazaryan 1, Tatiana Minkina 2
, Divya Pandey 3, Ragini Sharma 4, Karen Ghazaryan 1, Tatiana Minkina 21 Faculty of Biology, Yerevan State University, 0025 Yerevan, Armenia
2 Academy of Biology and Biotechnology, Southern Federal University, 344058 Rostov-on-Don, Russia
3 Department of Fruit Science, Dr YS Parmar University of Horticulture and Forestry, 173230 Solan, Himachal Pradesh, India
4 Department of Zoology, Panjab Agriculture University, 141027 Ludhiana, Punjab, India
Abstract
Deficits in the mineral Zn are responsible for a sizable proportion of the world’s disease burden and child mortality. With the increasing success rate of biofortification in major crops, the development of a genotype with enhanced Zn bioavailability will be an efficient and sustainable solution to nutrient deficiency-related problems. Due to the complex chemistry of the human system, the absorption of Zn from cereals is lower. This complexity is alleviated by phytate, a major phosphorus-storing compound in cereal and legume seeds, which negatively affects Zn binding. The results of recent studies on the distribution of elements and micronutrient speciation in seeds provide strong evidence for the presence of distinct Zn pools. This observation is supported by data from biofortified transgenic plant research. Several studies identify nicotinamide, a metal chelator, as a pivotal molecule. The loading of Zn into grains has been reported to increase with nicotinamide levels, which is a crucial finding. Intestinal Zn absorption can be greatly improved by nicotinamide. Furthermore, bioavailability tests suggest that the use of nano Zn-enabled devices could be an effective strategy to enable plant biofortification, which may significantly boost the Zn content in various cereal crops. This review comprehensively evaluated the scientific publications indexed in WoS, Scopus, and various other reliable databases and explored insights into how nano-enabled technology could be a solution for enhancing Zn content in cereal crops for combating malnutrition in humans.
Keywords
- zinc
- nutrient deficiency
- nano zinc-enabled
- nicotianamine
- cereal crops
Human health depends on high-quality, nutritious food [1], which is negatively impacted by food insecurity, dietary imbalance, the consumption of food grains with low nutritional value, a lack of dietary diversity, and more [2, 3, 4]. Food and nutritional insecurity may hinder dietary quality, greatly increasing the risk of both undernutrition and obesity [5]. Micronutrient uptake and dietary diversity declined due to the rise in cereal and cash crop cultivation in intensive cropping systems [6, 7]. This is especially true in the rural areas of low-income nations because of the prevalence of cereals in the diet. As food and nutrition security is still a far-off goal for many developing countries, finding a way to ensure food and nutrition security is becoming an important task for the scientific community and policymakers.
In contrast to the goal of achieving “Zero Hunger” by 2030, if the current trend continues, the number of people affected by hunger will surpass approx 840 million by 2030 [8]. Moreover, the income gap significantly affects food patterns; as compared to high-income countries, lower-income countries rely more on staple foods and less on fresh vegetables and meat.
Hidden hunger, also known as malnutrition, is a serious condition caused by an inadequate intake of micronutrients in the diet. Cereal-based diets are trendy, especially in countries with low incomes. Therefore, enriching them with micronutrients could increase microelement consumption. Bio-fortified staple grains (such as wheat, rice, and maize) with zinc possess the potential to greatly impact the fight against hidden hunger around the world. Nearly 2 billion people across the globe are suffering from a deficiency of zinc [9]. Zinc deficiency is associated with many major health disorders, namely homeostasis and immune function dysregulation, oxidative stress, apoptosis, and aging. During COVID-19 infections, zinc supplements have been a major source of immunity boosters [10]. Low levels of rhizospheric zinc availability are directly linked to dietary zinc deficiency [11]. Therefore, it is crucial for low-income and developing nations to increase the amount of bioavailable zinc in their staple foods [12].
Zinc is one of the most important micronutrients for plants; it is essential for their growth and development, and is required for cell membrane stability, cytochrome and chlorophyll production, photosynthesis, and cell transparency [13]. Moreover, it is necessary for lipid synthesis, carbohydrate, protein and nucleic acid metabolism, gene expression regulation, and phytohormone activity [14]. Zinc is the only necessary trace element found in all six classes of enzymes (oxidoreductases, hydrolases, transferases, isomerases, lyases, and ligases) [15]. Biofortifying crops is an environmentally friendly strategy to combat human diseases and improve diets. The potential of nanotechnology in agriculture is being pursued because of the increasing food requirements to sustain the fast-growing population throughout the world. Experts estimate that the global population will rise to around 9.6 billion by 2050 [16]. Precision agriculture, which uses nanotechnology, fills the gaps left by nutrient leaching and crop fortification [17]. Farmers are employing this technology in the nano-regime to simultaneously increase the quality and yield of their harvests.
Recently, there has been much discussion about using nanotechnology to strengthen crops in human societies [18]. Plants can be nano-biofortified in several ways, including seed priming, soil and foliar application, and growing plants in media rich in candidate nutrients [19, 20, 21, 22]. Zn-based nano-fertilizers are more effective than traditional methods in facilitating Zn delivery to plants [23, 24]. Zn nanoparticles (NPs) are particles with size ranges between 1 nm to 100 nm with unique mechanical, optical, electrical, thermal, chemical, and magnetic properties different from those of conventional and regular Zn particles [25]. NPs enter the plant through the roots or leaves, and move to other parts of the plant, impacting their positive, negative, or neutral effects on plant growth and development [24]. The ability of plants to produce high-quality proteins, sugars, and lipids is enhanced by the presence of Zn-NPs [26]. Previous studies have shown that the application of ZnO-NPs positively affects crops’ agronomic and physiological properties. Application of a 100 mg/L concentration of ZnO-NPs via foliar spray increased photosynthetic activity in young Cucumis sativus plants [27]. When 500 mg/kg of ZnO-NPs and bulk ZnO were applied to Portulaca oleracea, flavonoid and Zn content were increased by 1000% and 581%, respectively, compared to the controls [28]. According to Durgude et al. [29], the application of ZnO nanocomposites at a concentration of 30 ppm enhances zinc biofortification in rice. This review aims to draw attention to the connections between nano-biofortification and human health in the era of food security and the potential application of Zn-/ZnO-NPs-based agricultural products to raise the nutrient content of crops.
Zinc influences glycogen synthesis and gluconeogenesis, two pathways involved in
carbohydrate metabolism [30]. In most cases, zinc does not affect plant
respiration. Zinc deficiency impacts the net photosynthetic rate by 50%–70%,
depending on the plant species and the degree of nutritional deficiency. Zn is
important element for carbonic anhydrase enzyme activity and Zn deficiency may be
reduced the activity of carbonic anhydrase enzyme activity that reduced
efficient photosynthesis [31]. Dicotyledon carbonic anhydrase is a large molecule
and contains more zinc than monocotyledon carbonic anhydrase (such as in
cereals). Carbonic anhydrase activity in C
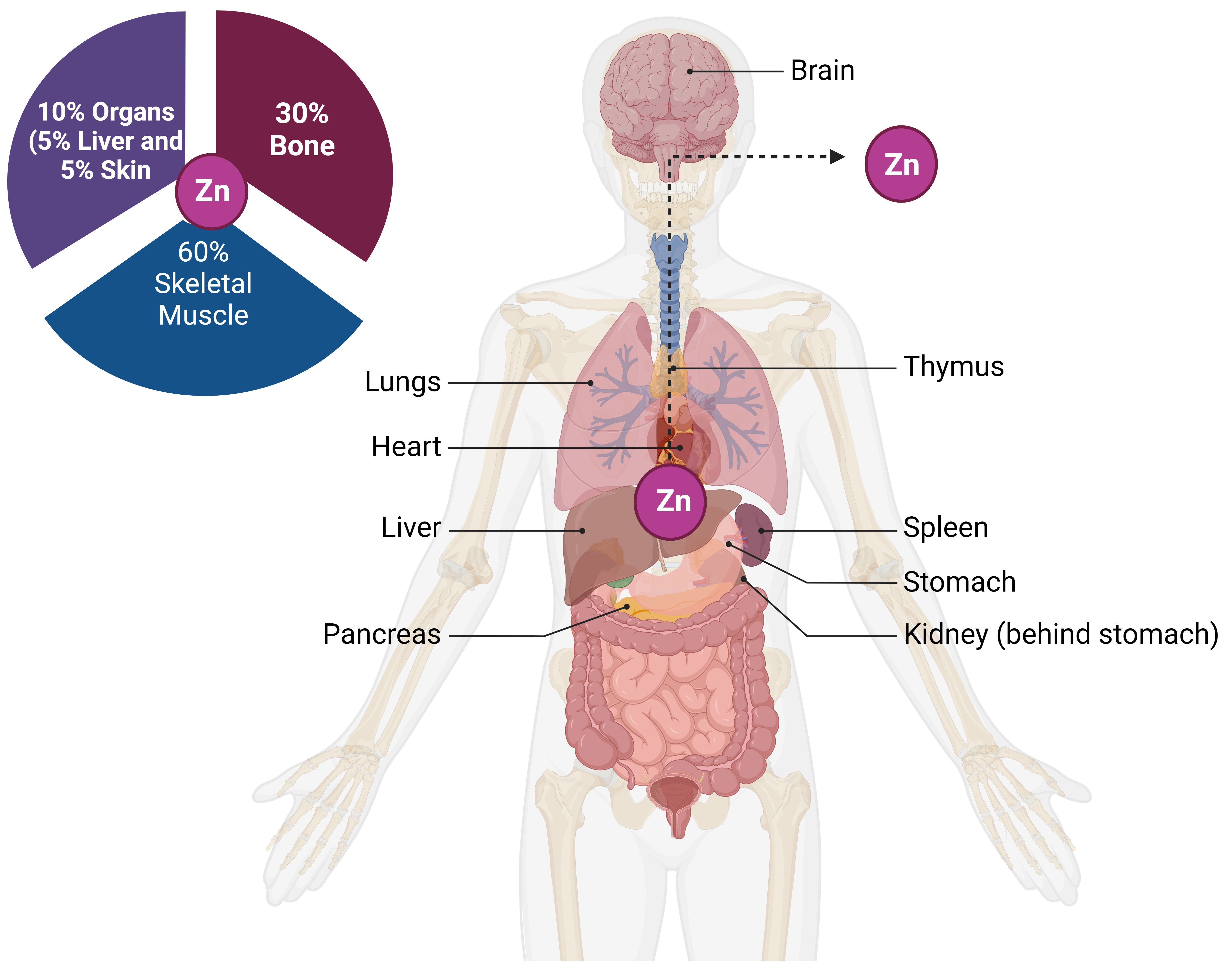
Research into the human genome using omics techniques found that zinc binds to around 10% of all human proteins [15, 38, 39]. This remarkable discovery has dramatically uncovered the importance of Zn in molecules that are involved in cellular processes. Several enzymes, transcription factors, and other proteins rely on Zn for proper functioning [40]. Zn-finger domains, LIM domains, and zinc finger domains are examples of regions in these proteins that might facilitate zinc interaction. The skeletal muscles and bones are the primary Zn reservoirs in the body (Fig. 1), but unable to hold more minerals than the organism requires. For the proper functioning of cells, it is necessary to consume Zn daily [41]. Even though zinc is generally safe and has low toxicity, its lack or overabundance can cause serious health problems [15]. Too much zinc could be toxic, resulting in symptoms like nausea, vomiting, fever, and headaches, while too little zinc can cause eye and skin lesions, hair loss, immune dysfunction, taste abnormalities, and growth retardation [42]. Supplementing with zinc alleviates zinc deficiency symptoms, demonstrating the importance of zinc as a trace mineral and the significance of zinc homeostasis as a physiological process [43]. Zinc is necessary for healthy development and differentiation, immune system function, neurological processes, and protein synthesis. There are reports of health benefits from supplementing with zinc and zinc complexes containing other compounds [44].
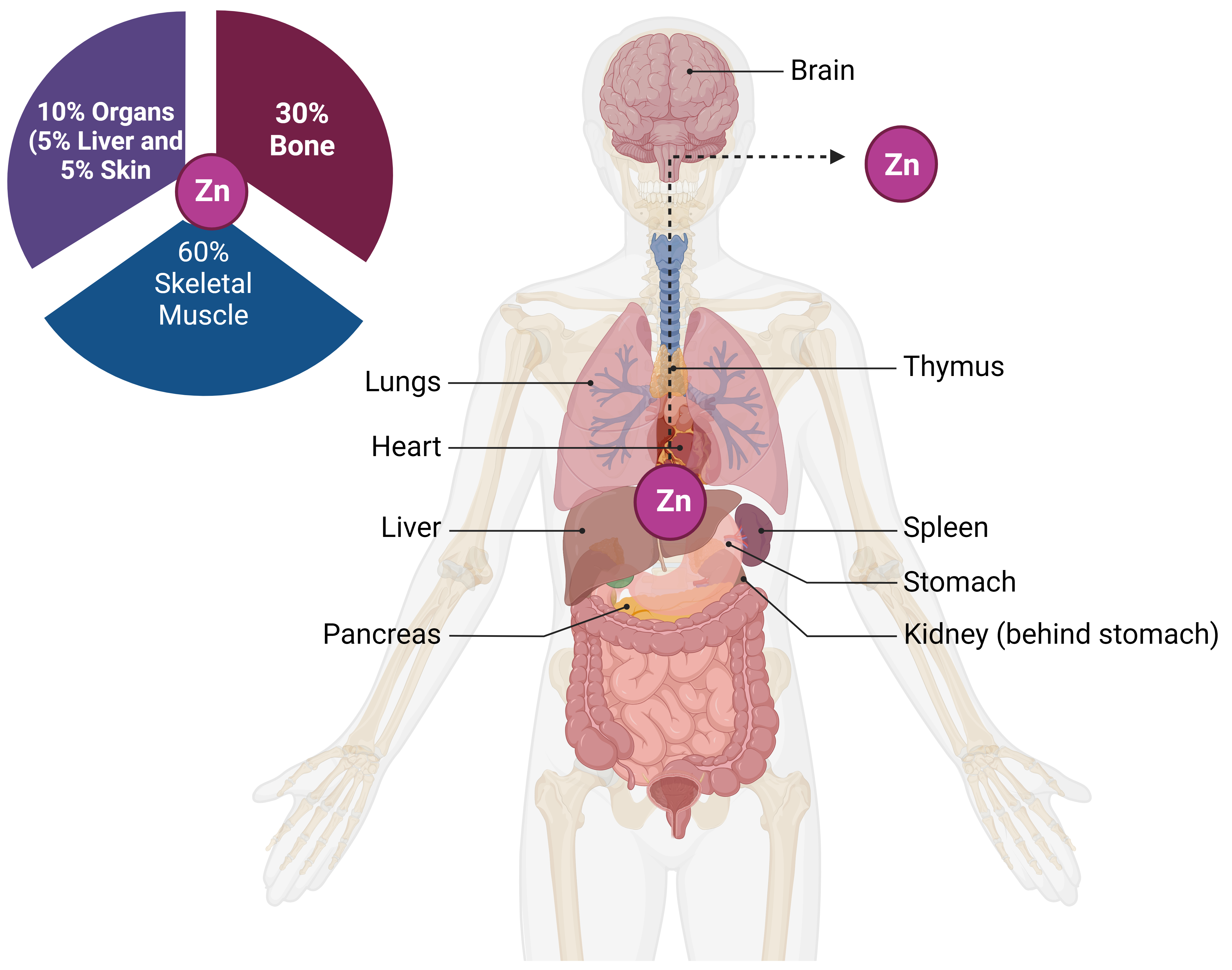 Fig. 1.
Fig. 1.Human body zinc reserves and distribution. Zinc absorption from food occurs in the small intestine and then travels throughout the body. The skeletal system, specifically the bones, and muscles, serves as a major Zn reservoir tissue.
More and more physiological roles for Zn in cellular responses have been uncovered recently. Zinc serves multiple purposes within the cell, including as a neuromodulator during synaptic transmissions and as an intracellular signal transducer. Zinc homeostasis is partly maintained by various zinc transporters, which are also essential for normal cellular processes [45, 46, 47].
An estimated 9% of the eukaryotic and 6% of the prokaryotic proteomes are
composed of zinc-binding proteins [48]. Enzymes such as carbonate dehydratase,
aldehyde dehydrogenase, Zn/Cu superoxide dismutase (SOD), and Zn-finger
DNA-binding proteins contain zinc in plant species [49, 50]. Zn-Cu-SOD protects
membrane proteins and lipids from oxidation by quenching the radicals produced by
the regular metabolism of the body. This vitamin is crucial for seed development
because it increases cytochrome production [51, 52]. Multiple physiological
processes in plants require zinc, including the production of tryptophan, a
precursor of indole acetic acid, as well as the transmission of inputs via
mitogen-activated protein kinases [53]. The control of auxin, the rejuvenation of
photosystem II, the stabilization of CO
To increase the likelihood of plants absorbing the added nutrients, fertilizers are fortified with plant-necessary nutrients [58]. Plants show nutrient deficiency symptoms in many ways, including unusual growth; however, in some cases, even plants grown in micronutrient-rich soil may show deficiency symptoms due to the inability of the roots to uptake and transport the nutrients because of their small pore size [59]. To sustainably feed the world’s growing population, it is necessary to discover methods to enhance plant nutrition and, by extension, the quality of the food they produce. The use of chemical fertilizers has dramatically increased crop yields and has been a long-standing practice [58]. However, they cause an imbalance of soil minerals and disturb the structure and nutrient status of soil, and the overall ecosystem, which together has long-term devastating effects [60]. According to recent research, graphene has emerged as an effective material that can carry plant nutrients [58]. It has minimal adverse effects on the environment and can increase crop production by slowly and deliberately releasing plant nutrients [58]. Biofortification of crops is a long-term strategy to address human health issues and reduce hunger. In recent years (Table 1, Ref. [8, 19, 20, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75]), there has been much discussion about using nanotechnology to strengthen crops in human societies [18].
| Scientific name | Concentration of ZnO-NPs | Characterization of ZnO-NPs | Application Methods | Main finding | References |
| Phaseolus vulgaris | 10–40 mg/L | 20 nm | Foliar Spraying | ZnO-NPs at 30 ppm produced the highest seed yield. | [72] |
| Triticum aestivum L. | 20–1000 mg/L | In soil | In comparison to ZnSO4, ZnO-NPs increased Zn in grain while not affecting Zn in leaf. | [20] | |
| Triticum aestivum L. | 25–100 mg/L | 20–30 nm | Seed primming | ZnO-NPs increased photosynthetic pigments and crop yield. | [19] |
| Phaseolus vulgaris L. | 4% ZnO-NPs | NA | Seed primming | Increased yield and Zn contents in the grain of red beans. | [8] |
| Moringa oleifera L. | 10 mg/L ZnO-NPs | 16.49 nm | Seed primming | Improving germination, photosynthesis pigment contents, antioxidant enzyme activity, and Zn nutrient contents. | [64] |
| Oryza sativa L. | 25 ppm | 20–30 nm | Seed primming | Pre-sowing treatment with 25 ppm ZnO-NPs increases rice yield and grain Zn nutritive value. | [65] |
| Triticum aestivum L. | 10 mg/L | NA | Seed primming | Improved seed germination, vigor index, fresh biomass, water uptake, and photosynthetic efficiency. | [66] |
| Oryza sativa L. | 10 µmol | 10 nm | Seed primming | Increase seed vigor, seedling emergence, and DSR field plant growth and productivity. | [67] |
| Amaranth L. | 10 ppm | 15–30 nm | Foliar | Foliar application of biosynthesized ZnO-NPs enhanced fertilization that increases yield, zinc content, and zinc use efficiency. | [68] |
| Brassica oleracea L. | 100 µM | 25 nm | Foliar | ZnO-NPs applied foliar to cauliflower improve zinc biofortification. | [69] |
| Solanum lycopersicum L. | 100 ppm | NA | Foliar | ZnO-NPs improved tomato growth, physiological traits, yield, and quality. | [70] |
| Vigna radiate L. | 20 ppm | NA | Foliar | ZnO-NP positively affects growth, photosynthetic pigments, protein content, activity, and enzymatic and non-enzymatic antioxidants in mung bean seedlings. | [71] |
| Glycine maxL. | 2 mg/kg | 18 nm | In soil/pot culture | ZnO-NPs increased grain yield and P uptake by 14% by regulating nutrient availability. | [73] |
| Sorghum bicolor L. | 1, 3, and 5 mg/L | 18 nm | In soil/pot culture | ZnO-NPs increased nitrogen, potassium, and zinc contents of grain under drought stress to help in biofortification under abiotic stress. | [63] |
| Solanum melongena L. | 50–100 mg/kg | Foliar Spraying | ZnO-NPs increased crop production under drought. | [61] | |
| Sesamum indicium L. | 3–10 mg/L | 10 nm, Biogenic | In soil/pot culture | Biogenic ZnO-NPs improved sesame germination and growth. | [74] |
| Eriobutria japonica L. | 12 mg/mL | Petriplate culture | Antimicrobial activity with fortification use. | [75] | |
| Triticum aestivum L. | 40, 80, 120 ppm | Soil culture | At a dose of 80 ppm, ZnO-NPs caused the greatest increases in height, seed weight, yield, and biomass. | [62] |
NA: Not available; NPs: Nanoparticle; DSR: Direct Seeded Rice.
Techniques like seed priming, soil and foliar application, and growing plants in nutrient-rich media can all be used to nano-biofortify the crops [61]. Several factors should be considered when nano-biofortifying crops, including:
(i) Identifying the most effective nanomaterials;
(ii) Calculating the ideal dosage without causing any harm to plant physiology or growth;
(iii) Analyzing the impact of NPs on the absorption, accumulation, and transport of other mineral elements.
One of the crucial micronutrients for plant growth is zinc, which serves as a cofactor for many vital enzymes. A worldwide shortage of zinc is a major cause of concern. Agricultural output in West Asia has taken a severe hit in recent years due to climate change impacts like dust storms, widespread salinization of the land, and significant droughts. Many studies have shown that providing plants with zinc helps them to manage environmental stresses [76]. When zinc is applied externally either as a foliar spray on leaves or as fertilizer in the soil, the bioavailability of Zn in the edible part of the plant also increases. Zinc enrichment through foliar fertilization (Fig. 2) is by far found to be an effective and environment-friendly method to elevate crop yields. Substances applied to leaves can enter the plant either through the cuticle or the stomatal pathway. The Zn content of rice can be significantly increased by applying zinc foliar fertilizer during the flowering stage [77]. However, the foliar spray is not successful every time. Due to heavy rains, the foliar spray may wash off the leaves and be leached into the soil, thus reducing the effectiveness of foliar input [78]. Zinc fertilizer increases the Zn content of harvested grains and improves crop yields. Zinc fortification of Oryza sativa (rice) has been the subject of numerous studies, which consequently deduced that adding zinc to the soil improves crop yield when used with other fertilizers [78]. However, the presence of fixed Zn ions, humus, and clay minerals, along with aluminum and iron oxides, renders Zn fertilization less effective in soil [79]. Thus, unutilized Zn fertilizer that builds up in farmland soil may harm agricultural ecosystems. As a result, it is essential to discover a Zn fertilizer that can replace traditional Zn inputs and have high positive performance with reduced negative environmental impacts [20]. The severe zinc deficiency in crops was first documented in 1937. However, the first instance of human Zn deficiency was documented in the 1960s [80]. A more significant section of the world has experienced acute Zn deficiency after the green revolution due to newly introduced high-yielding varieties within the system over the past 5 decades. These new crop varieties often had lower yields and were Zn-inefficient compared to older, locally adapted (landrace) varieties. To show their maximum yield potential, these varieties require more N, P, and K (nitrogen, phosphorus, and potassium, respectively) fertilizers; in some cases, the pH of the soil must be raised through liming [81]. In general, problems caused by Zn deficiency have worsened with higher phosphorus concentrations and high pH levels. Those varieties, however, are no longer high yielding. Instead, they now have yield plateaus in areas where they were introduced to serve the purpose [81]. A hidden Zn deficiency was later found. For instance, during the early 1960s, Turkey was able to increase its annual wheat grain production by 10 million metric tons (Mt) while using the same amount of arable land. This was made possible by the introduction of new wheat varieties and an increase in the intensity with which wheat was grown. However, Zn deficiency became increasingly common in some regions, such as central Anatolia. Furthermore, in the 1990s, a research program funded by NATO determined Zn deficiency as the root cause of low yields. Wheat yields increased by a factor of 6–8 with the use of Zn fertilizer in an area where they had previously been as low as 0.25 t/ha. Overall, with the addition of 10 kg of zinc per hectare, the yield of Triticum aestivum L. (bread wheat) increased by 43% (range 5–550%) [82].
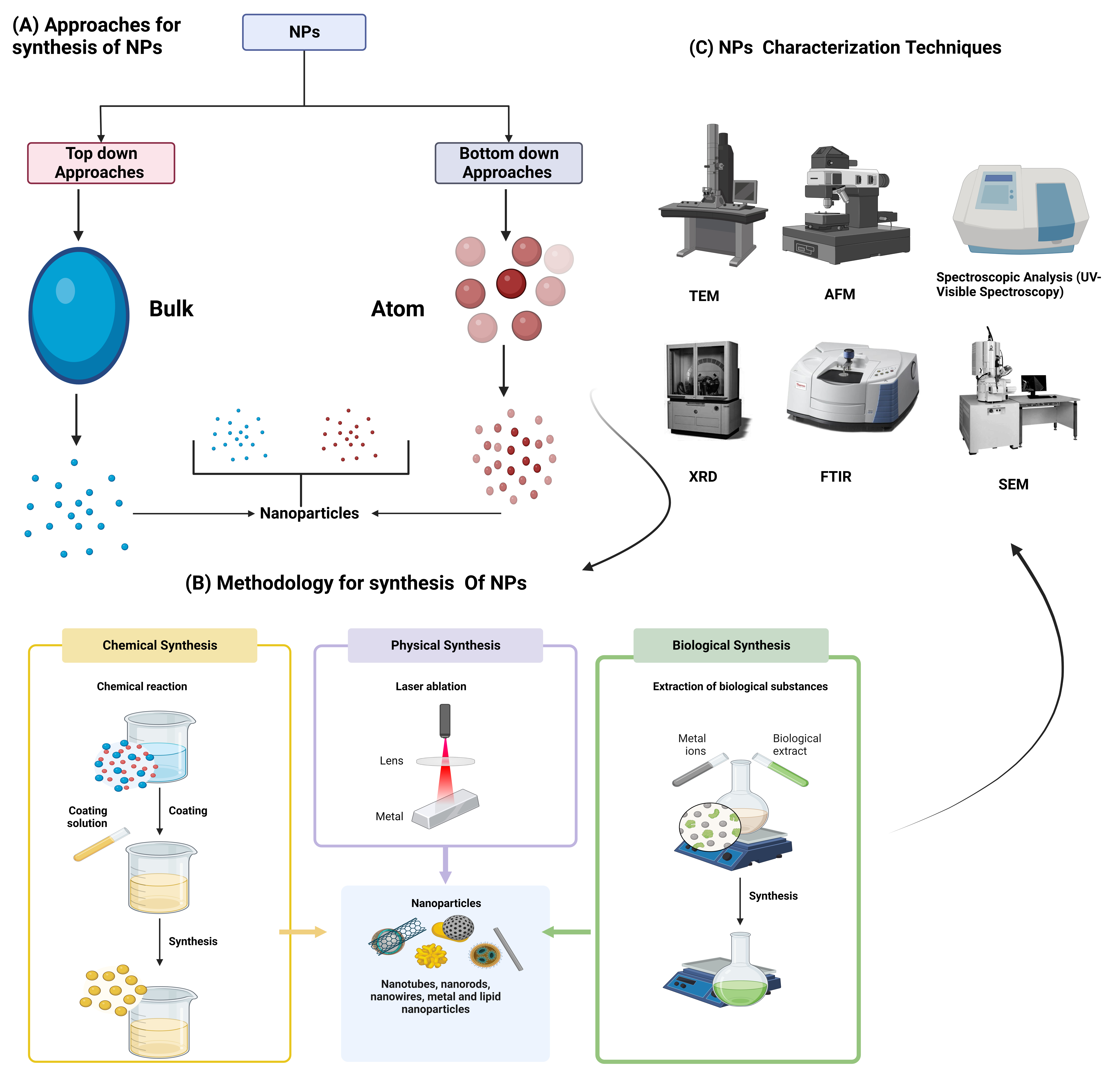
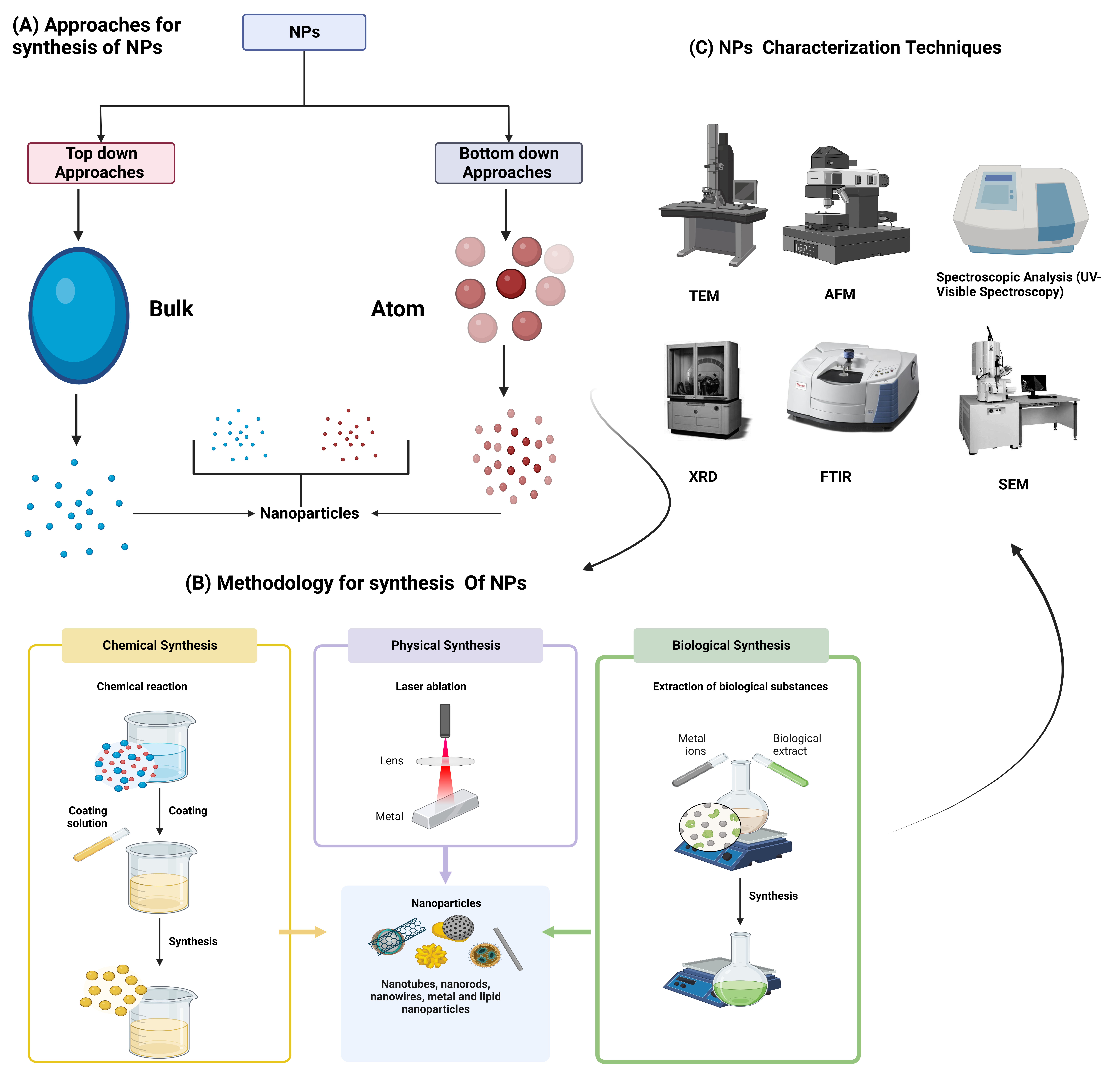 Fig. 2.
Fig. 2.Diagrammatic representation of (A) approaches (B) methodology and (C) characterization techniques involved in the synthesis of ZnO-NPs for crop fortification.
There are five different forms of zinc that plants can absorb: (a) free and
complex ions within soil solution; (b) non-specifically adsorbed cations; (c)
ions occluded mainly in soil carbonates and Al oxide; (d) living organisms along
with other biological residues; (e) lattice structures of primary and secondary
minerals [83]. Roots primarily uptake zinc as a divalent cation (Zn
Soil solutions generally have lower concentrations of Zn (4–270
µg/L) than the average concentration of approximately 64 mg Zn/kg.
Solubility is found to have a strong negative correlation with the pH of the
soil, with concentrations as high as 7137 µg/L being found in highly
acidic soils [89]. The primary factors that determine how much zinc is available
to plants are the ‘total Zn’ content, pH, oxidation/reduction levels, calcite
(CaCO
The fact that most plants have zinc concentrations ranging from 30–100 g
Zn
Human health depends on the quality of food consumed, and plants continue to be our primary source of nourishment [79]. The diet of communities consists of staples that are typically high in calories and are consumed regularly in large quantities [92, 93]. Therefore, staple foods are strongly associated with the nutrition status of their consumers, particularly among people living in rural and poor areas, where supplementary nutrients are scarce. In these regions, foods often lack the necessary micronutrients, leading to severe consequences and this is now an increasing problem around the world [94]. Half of the children go without the nutrition they need, causing them to be more susceptible to illness and reducing their cognitive potential. Iron and zinc deficiencies are responsible for significant infant mortality and worldwide health issues [95]. Dietary diversification, drug therapy, and industrial fortification are just a few of the proposed approaches to resolving nutrient deficiencies [96]. The long-term solution advocated is food diversity, but the people on the brink of malnutrition cannot afford it. Except for iodized salt, industrially fortified foods have not been particularly effective. Enhancing the nutritional value of a crop before, during, and after harvest is known as biofortification [13]. Biofortification is different from other interventions that address micronutrient deficiencies because it is cheap and accessible to all people [97]. Many people rely on these fortified crops as their staple diet. Thus, it does not incur additional expenses and is unaffected by social habits [96]. Vegetables and other non-staple foods are expensive despite their high vitamin and mineral content [13]. Many low-income people spend most of their food budget on staples, providing them with minimal energy and leaving them with bare minimums for more expensive foods like fruits, vegetables, and meat [98].
Agronomic and breeding techniques are among the biofortification strategies frequently utilized for enhancing the nutritional status of crops [98]. Fertilizers, both organic and inorganic, along with biofertilizers, are highlighted as part of agronomic interventions. Organic matter is preferred because of its significantly lower mineral content and prolonged nutrient release time, while inorganic fertilizers that are generally available in sizes greater than 100 nm are prone to volatilization and leaching. Many efforts to biofortify crops by enhancing their ability to absorb nutrients have been made but with limited success. Therefore, it is essential to employ nanotechnology to address some of these issues.
Top-down and bottom-up approaches are two of the most common classical methods for producing NPs with desirable shapes and sizes (Fig. 2A) [99]. Despite using distinct synthesis principles, these approaches yield NPs with the desired properties [99, 100]. Top-down methods involve breaking up large materials into smaller pieces, reducing them to eventually form NPs [99, 100]. These NPs were created via various methods, such as photolithographic processes, grinding, sputtering, and milling [101]. There are benefits and drawbacks to using each method to maximize the output efficiency of NPs. The top-down approach is feasible and can potentially produce a sizable number of NPs. The disadvantages of top-down methods include the imperfection on the surfaces of NPs and the risk of damage [102]. As the optical, physical, and chemical functions of NPs are dependent upon the surface architecture of NPs, top-down synthesis of NPs is only applicable in a few cases [103]. Another prominent technique for NPs production at the moment is the bottom-up method, which entails the blending or assembly of individual atoms, molecules, or clusters to create a wide variety of NPs [104]. NPs can be produced through self-assembling monomer and polymer molecules, nano structural precipitation, laser pyrolysis, flame spraying synthesis, sol-gel processing, and bio-assisted synthesis techniques [105]. Therefore, bottom-up is a versatile technique for creating nanoclusters that can be applied to various settings.
NPs are synthesized in a variety of physical methods including plasma arcing, pyrolysis with sprays, ball milling, electron irradiation, sputter accumulation, lithography methods, layer-by-layer growth, thermal evaporation, pulsed laser desorption, or the diffusion flame method (Fig. 2B) [106]. These methods employ a tube furnace and atmospheric pressure to create metal NPs via evaporation and condensation. Nanomaterials can be manufactured using various physical techniques, such as deposition, sputtering, ball milling, and plasma-based processes [105]. The production rate of metal NPs using most of these methods is extremely low. For example, 50% of nanomaterials are produced by ball milling [107]. The particle size distribution is large because 6–8% of the sputtered material is smaller than 100 nanometers. Furthermore, high amounts of power are required for laser ablation and plasma techniques. Most physical processes are too costly to be practical owing to the low throughput, uneven size distribution, and excessive energy requirements [107].
ZnO-NPs can be physically synthesized through high-energy ball milling, melt mixing, physical vapor deposition, laser ablation, sputter deposition, electric arc deposition, and ion implantation [108]. ZnO-NPs are produced at excessively high rates during most physical/mechanical processes and are typically used in manufacturing. Since its inception in 1961, the high-energy ball milling method has become the most popular and widely used process for the synthesis of various type of NPs [108]. Ball mills are used to grind materials into extremely fine powder by subjecting them to high-velocity collisions between the mill’s steel balls [109]. Burmeister and colleagues reported high-energy ball milling processes as efficient, cost-effective, and simple techniques for preparing ZnO nanostructures. ZnO nanopowder particles were identified to have a crystallite size of 15 nm, a particle size of approximately 60 nm, and a lattice strain of 0.67% using X-Ray diffraction (XRD) and field emission scanning electron microscopy (FESEM) [110]. Additionally, Salah et al. [109] employed a similar approach with ZnO for antimicrobial activity. Particles can be removed from the surface of a solid or a liquid using a laser ablation technique. Ismail and coworkers found that the average diameter of ZnO spheres is 35 nm. In double-distilled water, they used pulsed laser ablation [111]. The energy absorbed by the laser causes the material to heat up and evaporate at low reflux, but may turn into plasma at high reflux. Vapor-solid-liquid (VLS), physical-vapor-deposition (PVD), and chemical-vapor-deposition (CVD) are other common and well-studied techniques [112]. Metals are deposited onto the surfaces using physical vapor deposition (PVD) techniques. PVD utilizes two different types of processing methods: evaporation and sputtering. The term “sputtering” describes the process by which particles are ejected from a surface after being struck by higher-energy particles. The sputtering process utilizes ions provided by plasma [113].
The chemical synthesis of NPs is performed using a bottom-up strategy (Fig. 2B) [114]. One such process is nucleation which reduces metal cations to metal monomers using water-soluble cations as a precursor [115]. Where a cluster of reduced metal atoms has assembled, the growth of particles is automatically halted by the reducing agent [116]. Consequently, the particles grew to a thermally stable size [117]. Atoms and smaller molecules interact to create nanomaterials. Sol-gel synthesis, chemical vapor deposition (CVD), pyrolysis, microemulsion, hydrothermal, polyol synthesis, and plasma-enhanced chemical vapor deposition are examples of different chemical synthesis methods [115]. Metal NPs (MNPs) are synthesized in a chemical solution during chemical preparation; different chemical reactions and chemical compositions are used for this purpose. For instance, the most widely used method for the preparation of MNPs is the chemical reduction of metal ions inside reversed micelles in a nonpolar solvent [118]. The reversed micelles provide a safe environment for the chemical reduction of a metal slat that has been dissolved in water, and the ratio of water to the volume of reversed micelles influences the particle size.
Zinc salts are typically precipitated and coprecipitated upon reaction with a
reducing agent (typically inorganic alkalis). To obtain the desired morphology
and characteristics of the NPs, the resulting soluble or insoluble precipitate is
washed and calcined at various temperatures. By adjusting the reaction
conditions, the shape and size of the ZnO can be controlled [119]. Colloidal
techniques rely on basic chemistry. In the literature, however, Faraday’s
synthesis of gold nanoparticle colloids via the reduction of HAuC
The spray pyrolysis method, in which the zinc salt precursor is heated in a flame to produce aerosol droplets, is widely used. Dehydration causes the droplets to become smaller and disperse throughout the gas. The final steps are the decomposition and sintering of the necessary material [121]. Another technique is inert gas condensation, which can be divided into physical vapor deposition (without catalytic interaction) and chemical vapor deposition (with catalytic interaction). These techniques primarily involve resistive heat evaporating a zinc source inside a chamber. Various heat sources like electron, laser, or radio frequency beams are also used. The vapors are compelled to move from the heating chamber into the cooler, inert-gas chamber, where they are gathered for further consolidation [108]. The aggregation and coalescence of ZnO NPs is the main drawback of this method. The one-step levitational gas condensation method for ZnO that Uhm and colleagues reported is another significant technique. The apparatus is comprised of a levitation and evaporation chamber, a 2.5 kW induction generator, and an oxygen concentration control unit. The obtained NPs, which had a diameter of 30 nm, displayed excellent morphology [122].
Biological methods of NP synthesis are becoming increasingly popular in the field of nanotechnology [123]. Eco-friendly materials for the biological synthesis of NPs include plant extracts [124], bacteria [125], and fungus [126] (Fig. 2B). Since no toxic chemicals are used in this synthesis of Ag-NPs, it is therefore environmentally friendly and suitable for use in the pharmaceutical and biological industries. Green synthesis is superior to traditional methods because it is cost-effective, safer for the environment, and can be easily scaled up to meet high production needs [127].
Using plants and plant extracts as machinery for synthesizing metal NPs is attractive because it avoids the use of potentially dangerous materials and the hassle of culturing and processing [127]. However, plant extracts are more favorable due to the simplicity and low cost of the methodology [11, 127, 128, 129, 130]. Plant extracts have been studied more than other biological catalysts for nanoparticle synthesis. Plants can produce the NPs through in vivo and in vitro synthesis processes [11]. Plants synthesize such NPs by absorbing metals from the soil and water, hyperaccumulating them, and then reducing them to nanoparticle form. Phytoremediation and phytomining both make heavy use of these methods [131]. Nonetheless, silver and gold NPs used on plants have garnered the most attention [108]. Plant- and plant extract-based approaches for the synthesis of ZnO are the recent focus, e.g., Carom-Trachyspermum ammi seed extract [132], Nyctanthes arbor-tristis flower extract [133], Ulva lactuca seaweed extract [134], Muraya koenigii seed extract [135], Calotropis procera leaf extract [136], Artocarpus heterophyllus leaf extract [137], Moringa oleifera [138], Carica papaya leaf extract [139], Nephelium lappaceum L. fruit peel extract [133], Moringa oleifera leaf extract [140], Catharanthus roseus leaf extract [141], Camellia sinensis leaf extract [142], Citrus aurantifolia fruit extract [143], Oryza sativa rice extract [144], Passiflora caerulea L. leaf extract [145], sucrose (as a capping agent) [146], whey (as a chelating agent) [147], and Citrus sinensis fruit peel extract [148].
Microbes are considered to be green factories for producing NPs. Bioremediation, biomineralization, bioleaching, and biocorrosion are some biological applications that exploit metal-microbe interactions [149]. Despite its limitations, it is one of the most eco-friendly and long-lasting methods currently available. Metal and metal oxide synthesis, especially ZnO, use prokaryotic and eukaryotic organisms. Furthermore, the synthesis can take place either inside or outside the cell.
In nature, fungi serve as both decomposers and parasites. Fungal biomass is incubated with a zinc salt solution for a specified period in the dark during intracellular synthesis, while fungal filtrates are treated with the precursor solution and synthesis is evaluated during extracellular synthesis [150]. Using fungi as a source of ZnO has been the subject of several studies. To study fungi in rhizosphere soil, Jain et al. [151] isolated 19 cultures. Extracellular synthesis of ZnO was found to be most promising in Aspergillus aeneus isolate NJP12 when grown at room temperature. Protein coating on the resulting NPs provided stability [151]. Jacob and his colleagues used Aspergillus niger fungal filtrate in a different experiment to develop ZnO. The average size of the synthetically produced NPs was 39.4–114.6 nm [152]. By using Aspergillus terreus filtrate, Baskar, and colleagues were able to prepare ZnO extracellularly. ZnO particles synthesized in this study ranged in size from 54.8 nm to 82.6 nm [153].
Numerous studies have been conducted on the use of bacteria as a green synthesizer for ZnO; for instance, the bacterial strain Aeromonas hydrophila formed spherical- and oval-shaped NPs with an average diameter of 57.72 nm [154]. Basic combinations of cellular biochemistry, metal ion transportation into and out of cells, microbes are resistance to toxic metal mechanismand able to activated metal-binding sites, accumulation of metal ions within cells, and metal oxide nucleation have been proposed as NPs synthesis through microbial routes [155]. ZnO with a diameter of 5–15 nm was reported to be produced by Lactobacillus sporogenes. The authors proposed a synthesis mechanism predicated on the reduction of metals by oxidoreductases that were triggered by the constituents of the nutritional media and changes in pH [156]. ZnO particles can also be synthesized by actinomycetes, which share characteristics with both fungi and bacteria among prokaryotes. For instance, the aqueous extract of the brown marine macroalgae Sargassum muticum is being studied as a potential component of an environmentally friendly and economically viable method. The resulting NPs were hexagonal wurtzite with a diameter of 30–57 nm [157].
NPs can be described by their size, shape, surface area, and distance from each other [158]. The consistency of these values is crucial in many contexts. Standard methods for determining the nature of NPs include Fourier transform infrared spectroscopy, powder X-ray diffraction, energy-dispersive spectroscopy, atomic force microscopy, UV-visible spectrophotometry, scanning electron microscopy (SEM), transmission electron microscopy (TEM), and dynamic light scattering (Fig. 2C) [159]. NPs of ZnO-NPs are typically characterized using wavelengths between 350–380 nm [160]. Dynamic light scattering (DLS) showed that it could be used to characterize the surface charge and size distribution of particles suspended in a liquid solution [161]. Standard techniques for morphological characterization on the nanometer to micrometer scale include SEM and TEM [127, 162]. However, the resolution of the TEM is one thousand times that of the SEM. Fourier-transform infrared spectroscopy (FTIR) spectroscopy is an effective method for elucidating the chemical composition of surfaces [130, 131, 163, 164]. Surface chemical residues and organic functional groups of plant biomolecules (such as hydroxyls and carbonyls) can be identified by FTIR. Nanoparticle crystal structure, characterization, and phase identification can be established with XRD [165]. This method involves penetrating the nanomaterial with X-rays and comparing the resulting diffraction pattern to reference patterns to determine its internal structure. The elemental composition of the metal nanoparticle is determined using energy-dispersive spectroscopy [165].
The effectiveness of Zn fertilizer applied to the soil depends on the pH of the
soil; Zn is more accessible in acidic soils. Specifically, each increment of pH
has a one-hundred-fold negative effect on soil solubility [166]. There is
evidence that liming acidic soils can decrease Zn availability [167]. The
formation of insoluble calcium zincate, a pH increase caused by CaCO
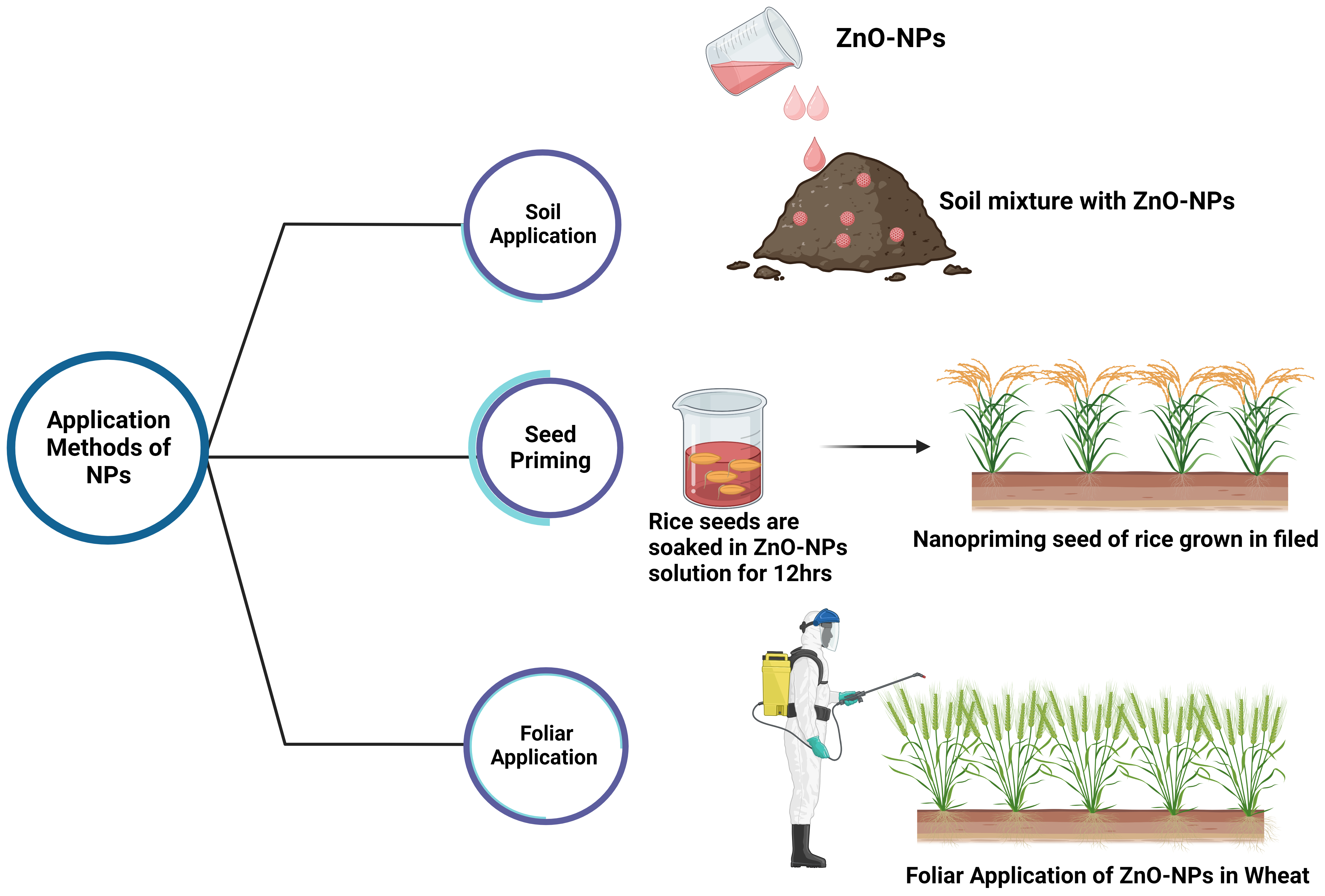
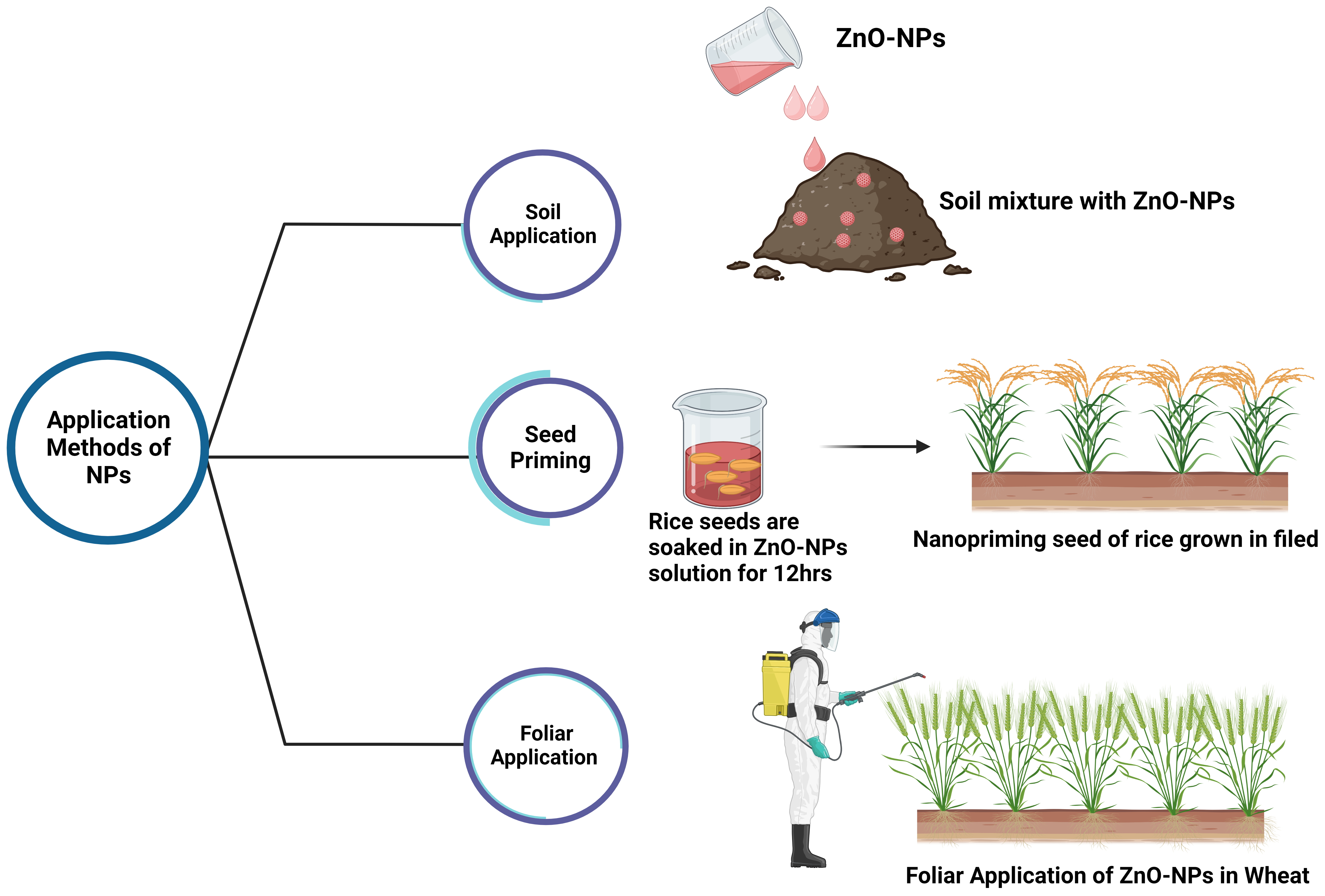 Fig. 3.
Fig. 3.Application methods of ZnO-NPs for fortification of crops.
Seed priming is a pre-planting process that causes a physical change in the seed that increases its germination rate [172]. NPs-based pre-soaking treatments of seeds, known as seed nanopriming, induce nanopore formation in the shoot, which promotes the uptake of water (Fig. 3). Additionally, it activates antioxidant mechanisms against reactive oxygen species (ROS) in the seeds, which form hydroxyl radicals that loosen the cell walls and act as catalysts for the rapid hydrolysis of starch and uptake of nutrients [173, 174]. Seeds with a high zinc concentration have been shown to boost seedling health and crop stand (Fig. 3) [175]. Plants grown in nutrient-poor soil produce nutrient-deficient seeds, reducing the overall vigor, growth, and yield of subsequent generations of plants [175]. Since plant roots absorb Zn via diffusion, a lack of moisture negatively impacts Zn availability [167]. Therefore, the effectiveness of Zn applied to the soil may vary depending on the soil irrigation. The outcome of Zn nutrition can be highly variable under rainfed conditions, where soil moisture content entirely depends on rainfall. Application of pre-soaked ZnO-NPs from Zn-rich seeds increases plant growth and yield under Zn-deficient conditions, particularly in rainfed conditions (Table 1) [175].
Seed priming with 4% ZnO-NPs can improve the red bean’s morphological and phenological traits, yield and yield components, third trifoliate leaf, maturity, grain filling period, plant height, and number of seeds per pod [8]. Nanopriming moringa (Moringa oleifera L.) with 10 mg/L ZnO-NPs can increase the percentage of germination, length, dry weight of the plumule and radicle, concentration of photosynthetic pigments, phenolic compounds, flavonoids, vitamin C, glutathione (GSH), Zn content and antioxidant activity, including ascorbate peroxidase (APX), catalase (CAT), and glutathione (PAL) [64]. Pre-sowing treatment with 25 ppm of ZnO-NPs can enhance the agronomic profile of rice by increasing yield traits and grain Zn nutritive value [65]. The priming of wheat seeds with ZnO-NPs at a concentration of 10 mg/L significantly and positively influences seed germination performance, vigor index, fresh biomass, water uptake, and photosynthetic efficiency [66]. Pretreating direct-seeded rice with 10 mol ZnO-NPs increases seed vigor, promotes early seedling emergence, and boosts plant growth, grain nutrients, and productivity in Direct Seeded Rice (DSR) [67]. However, under poor resources and stressed conditions, nanopriming seeds may play an essential role in overcoming the Zn deficiency problem.
The effectiveness of foliar Zn application is highly dependent on the type of fertilizer used, crop characteristics (particularly leaf characteristics), and the genetic potential of the crop (Table 1) [176]. The foliar application method has many benefits, including a reduced need for fertilizer, eliminating Zn fixation, and preventing antagonist nutrient interference with Zn uptake (Fig. 3, Table 1) [167]. When applied to the leaves, Zn travels through the phloem to reach the developing grains [169]. Foliar application of Zn has been proven to be more effective than soil application at raising grain Zn concentration in crops like wheat (Fig. 3). [177]. When applied, foliar applications exert a greater or lesser effect on increasing grain Zn concentration [178]. Foliar applications of 10 ppm biogenic ZnO-NPs significantly contribute to nutrient recovery, increased yield, and productivity compared to nutrient inputs with higher Zn contents in amaranth [68]. Applying 100 µM ZnO-NPs via foliar spraying can potentially improve zinc biofortification in cauliflower [69]. Tomatoes treated with a foliar spray of ZnO-NPs with a concentration of 100 ppm show improvements in growth parameters, physiological traits, yield attributes, yield, and quality traits [70]. Research has revealed that a concentration of 20 ppm of ZnO NP in mung bean seedlings produces the best results concerning growth, photosynthetic pigments, protein content, activity, and the amount of both enzymatic and non-enzymatic antioxidants present [71].
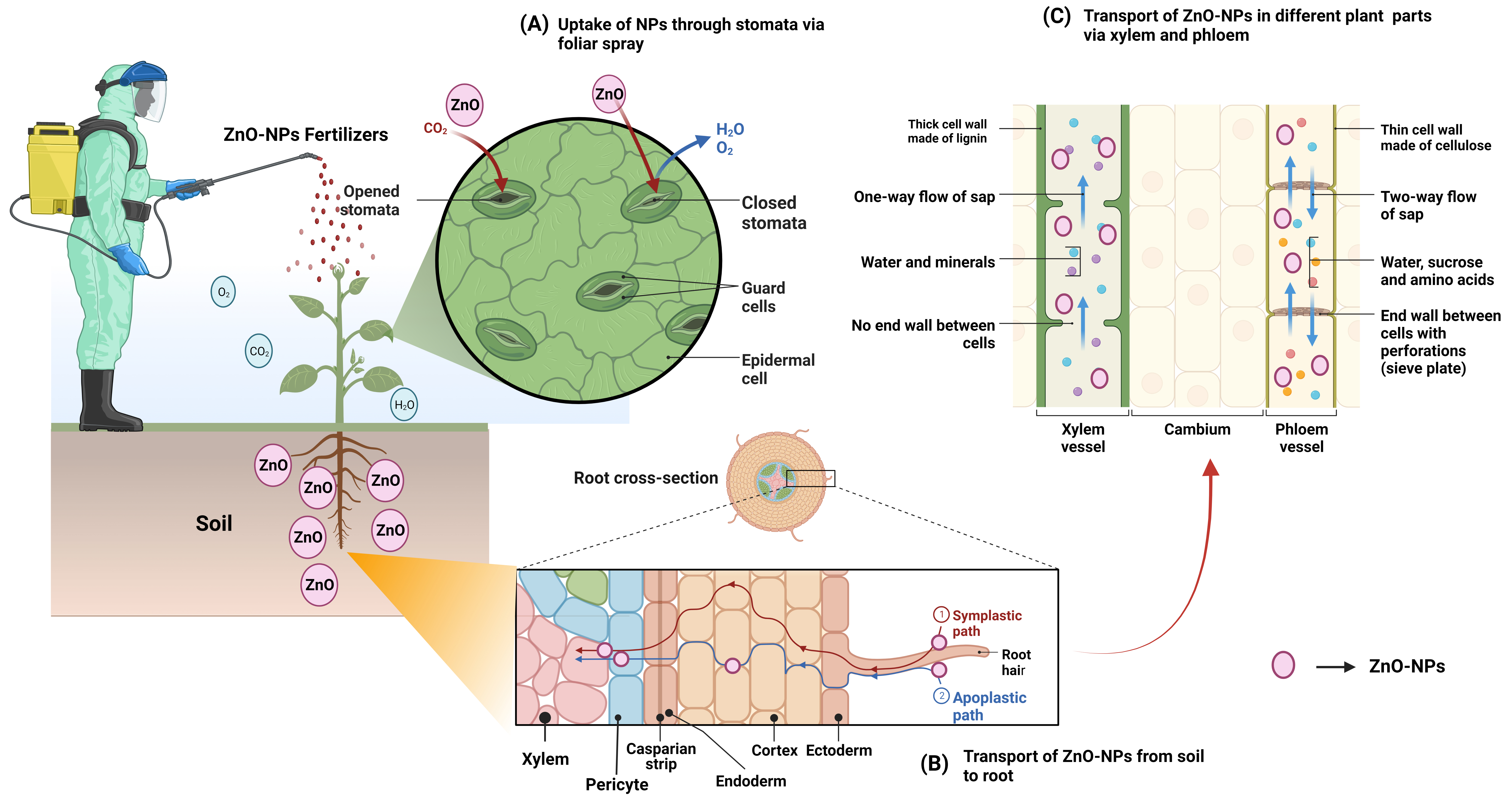
Bio-transformations in the soil determine the bioavailability and toxicity of NPs. In response to root contact, NPs translocate to the aerial parts of the plants, where they accumulate in cells and organelles (Fig. 4). Plant roots are responsible for the initial stage of bioaccumulation, which is the uptake of NPs from the soil [179]. Numerous studies have analyzed various NPs and proposed that NPs accumulate in plants through crystal phase dissolution, biotransformation, and bioaccumulation, which play a role in the spread of absorbed compounds from the roots to the rest of the plant. Likewise, size plays a role in the accumulation of NPs, toxicity, and the rate at which they are transported into plant cells [180]. This is because the size of NPs depends on how they enter into cells (i.e., plasmodesmata) or plant cell organelles. Regarding the reactivity with cells or plant structures, the surface area, shape, and agglomeration of NPs are interconnected [181]. The area and morphology of the NP surface are crucial parameters in localizing the point of contact between NPs and plant cells. Due to the negative charge of the cell wall, the NPs can only cling to the plant cells via their electrical charges. Hydrophobicity on the plant surface was the third most important factor in uptake and translocation after charge and particle size [182]. NP absorption, translocation, and aggregation in plants can be better understood by observing the fundamental structures of the nanomaterials [183]. Standardized laboratory experiments assessing NPs in plant tissues at various levels are required to pinpoint the precise effect of NPs supported by their physical-chemical properties. Therefore, a thorough investigation is necessary to fully understand and explain the processes of absorption, translocation, and accumulation, and knowledge of the composition of NPs [184]. The distribution and localization of NPs within plants and their cellular organelles require careful tracking and monitoring. Osmotic pressure, capillary forces, and translocation through root epidermal cells have been reported as pathways for the tiny NPs (diameters ranging from 3 to 5 nm) to access plant roots [185]. Due to the small pore size, root epidermal cells are only partially permeable, preventing large NPs to enter the root (Fig. 4).
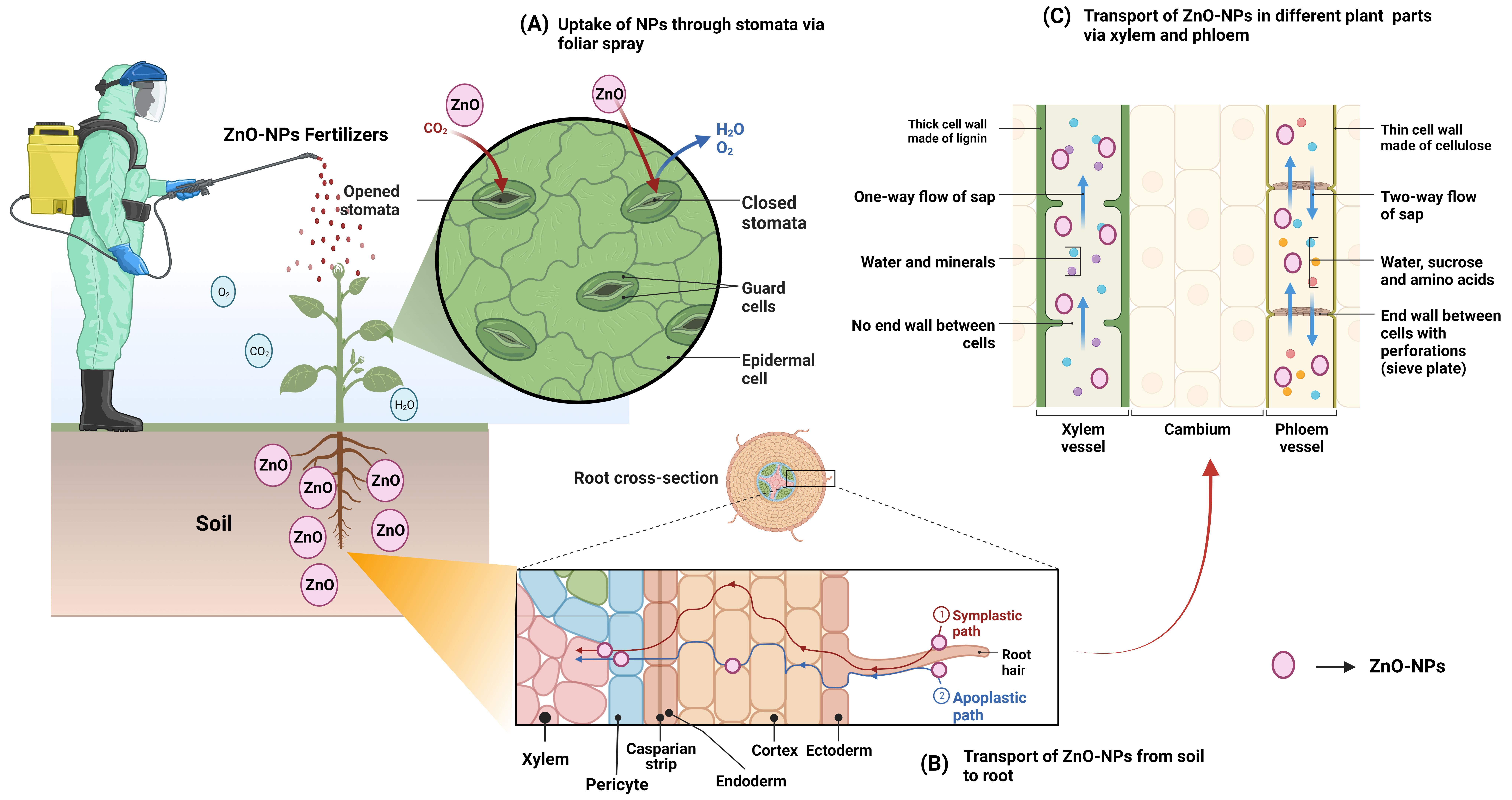 Fig. 4.
Fig. 4.Mode of actions of ZnO-NPs based nanofertilizers depicted diagrammatically (A) plant system; (B) from soil to root and (C) transport through xylem and phloem in different parts of plants via symplastic and apoplastic pathway.
The uptake of some NPs is aided by the formation of new pores in the epidermal cell wall [186]. The xylem can grow vertically after NPs travel apoplastically to the central vascular cylinder through cell walls and extracellular spaces. However, NPs must symplastically cross the casparian strip barrier to reach the central vascular cylinder (Fig. 4). This is accomplished by binding to carrier proteins in the membrane of endodermal cells, which triggers endocytosis, pore formation, and transport. When NPs are taken up into the cytoplasm, they can be transferred to neighboring cells via plasmodesmata [187]. The casparian strip is a collection point for NPs that cannot internalize, however, NPs that reach the xylem are transported to the shoots, where they are returned to the roots via the phloem (Fig. 4) [102]. Additionally, NPs have been detected in plant nuclei, cytoplasm, and epidermal cell walls [188]. Nutrient absorption can be altered by NPs that are not absorbed in the root surface of the soil aggregate [189]. Seeds can absorb NPs directly through the parenchyma and into the intercellular spaces, where they are diffused to the cotyledons [190]. When leaves are applied with NPs, the particles are absorbed through the stomata or the cuticles. The cuticle is the first line of defense in protecting a leaf from foreign particles, the cuticle is the first line of defense, keeping NPs smaller than 5 nm out. NPs gain access to the plant vascular system via stomata and are subsequently transported to the organs of photosynthesis and transpiration via apoplastic and symplastic pathways, respectively [189]. NPs (10–50 nm) are transported to new locations via the cytoplasm of neighboring cells (symplastic route). Henceforth, bigger NPs (in the range of 50 to 200 nm) are shuttled around in the intercellular spaces of the body (apoplastic route).
Internalized NPs travel through the phloem sieve tubes with the sugar solution. An effective sap sink is found in the roots, young leaves, stems, fruits, and grains of plants, thus NPs travel unrestrictedly via phloem transport and accumulate to varying degrees [183]. As a nonselective path with least resistance, the apoplastic pathway is well known. The apoplastic route is commonly accepted as the most efficient route for translocating many water nutrients and non-essential metal complexes [189]. The capacity of NPs for adsorption following foliar application was found to be primarily influenced by application method, particle size, concentration, and environmental conditions [181]. Several factors that affect the trapping of NPs on the leaf surface include morphology, chemical makeup, trichrome, waxes, and exudates on the leaf [191]. ZnO-NPss were examined using synchrotron X-ray absorption spectroscopy to determine their transformation after contacting different plants [192]. The various forms of ZnO-NPs, such as Zn-phosphate, accumulate most heavily in the roots and shoots of Zea mays when exposed to hydroponic conditions. It might be a consequence of increased rhizosphere ionic Zn dissolution, plant uptake, or translocation [193]. Soil-grown wheat crops exhibited similar Zn accumulation and speciation [194]. The translocation of ZnO NPs from the soil into glycine max was observed [195].
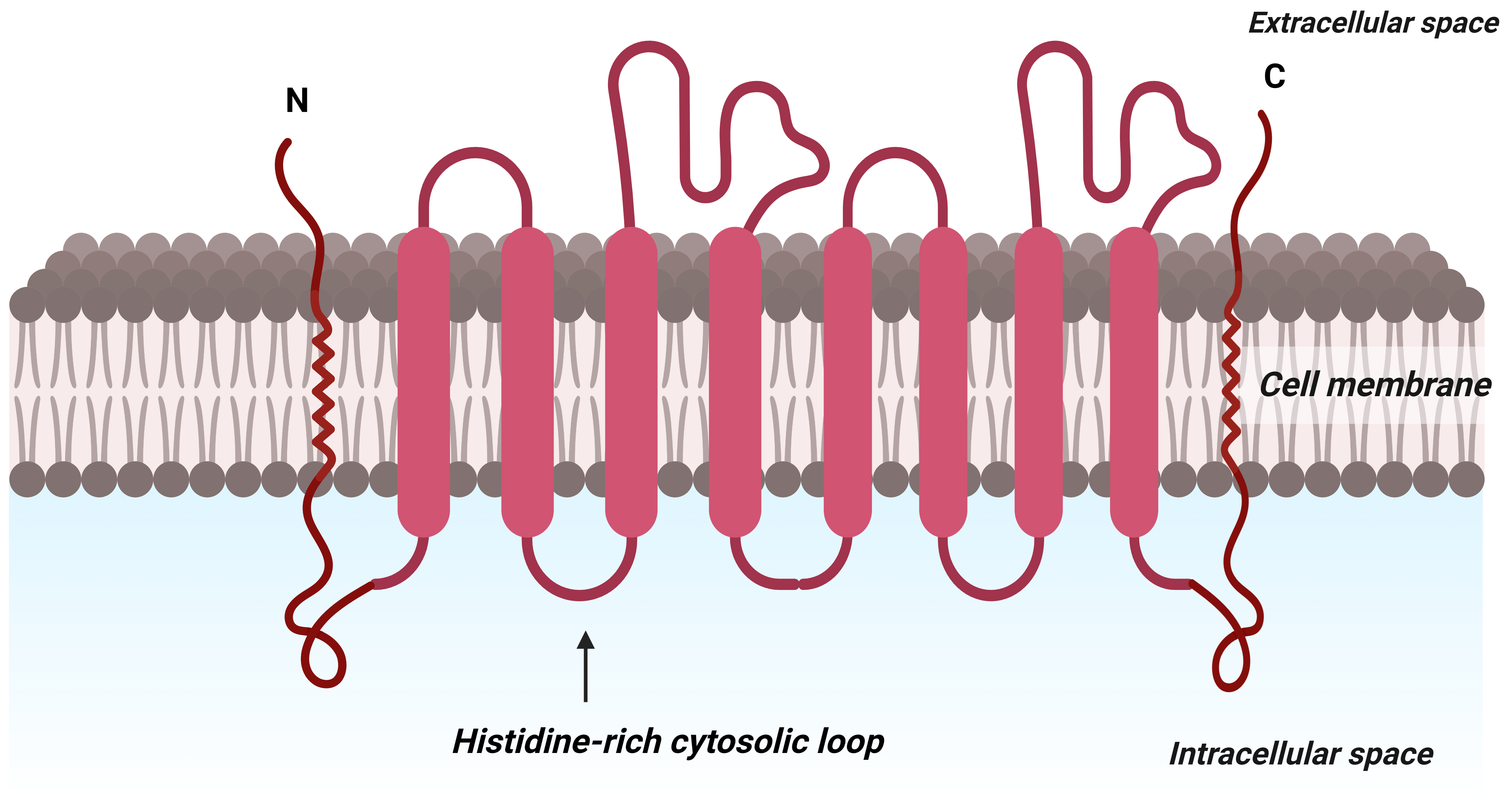
Micronutrient transporters in plants are responsible for the uptake of Zn from
the soil. ZRT-IRT-like proteins directly absorb the Zn
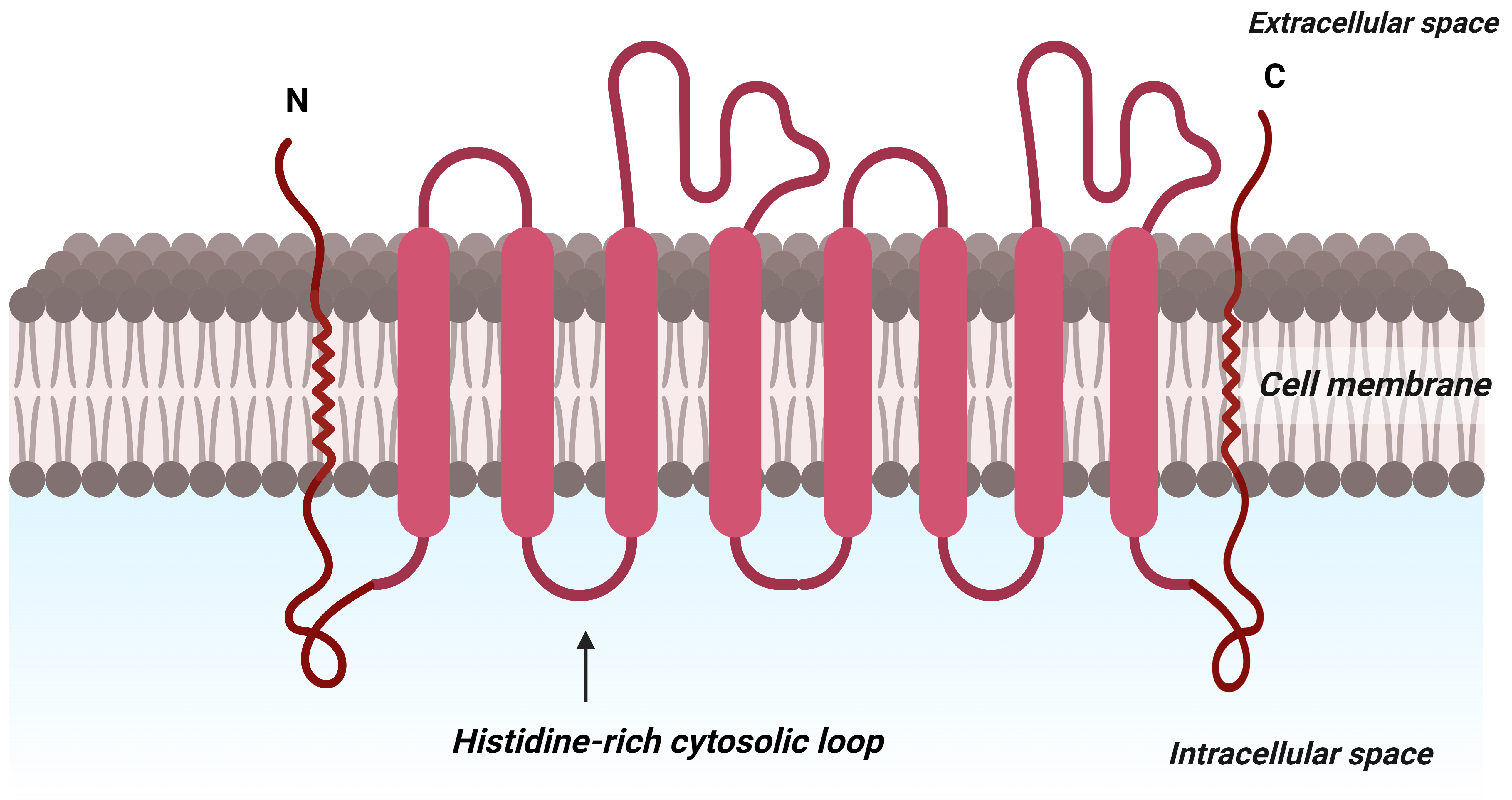 Fig. 5.
Fig. 5.Predicted diagrammatic structure depicting the protein structure of membranes of ZIP family of micronutrient transporter that aids in transporting zinc at the molecular level in plants. C: C terminus; N: N terminus.
Gene expression studies in yeast and experiments on roots show that the proteins encoded by ZIP1, ZIP3, and ZIP4 possess a high affinity for zinc and may also aid in transporting Cd and Cu into the plant. While ZIP1 and ZIP3 are expressed solely in the roots during Zn deficiency, ZIP4 is expressed both in the roots and the shoots. Plants typically utilize various members of multigene families to facilitate the ZnO-NPs metal transport along with ZIPs, e.g., YSL1, FER1, FER2, and PHT4 (Fig. 4) [196]. ZIPs transporters are involved in the transport of external ZnO-NPs, continuously supplying Zn to different parts of plants during Zn deficiency [197].
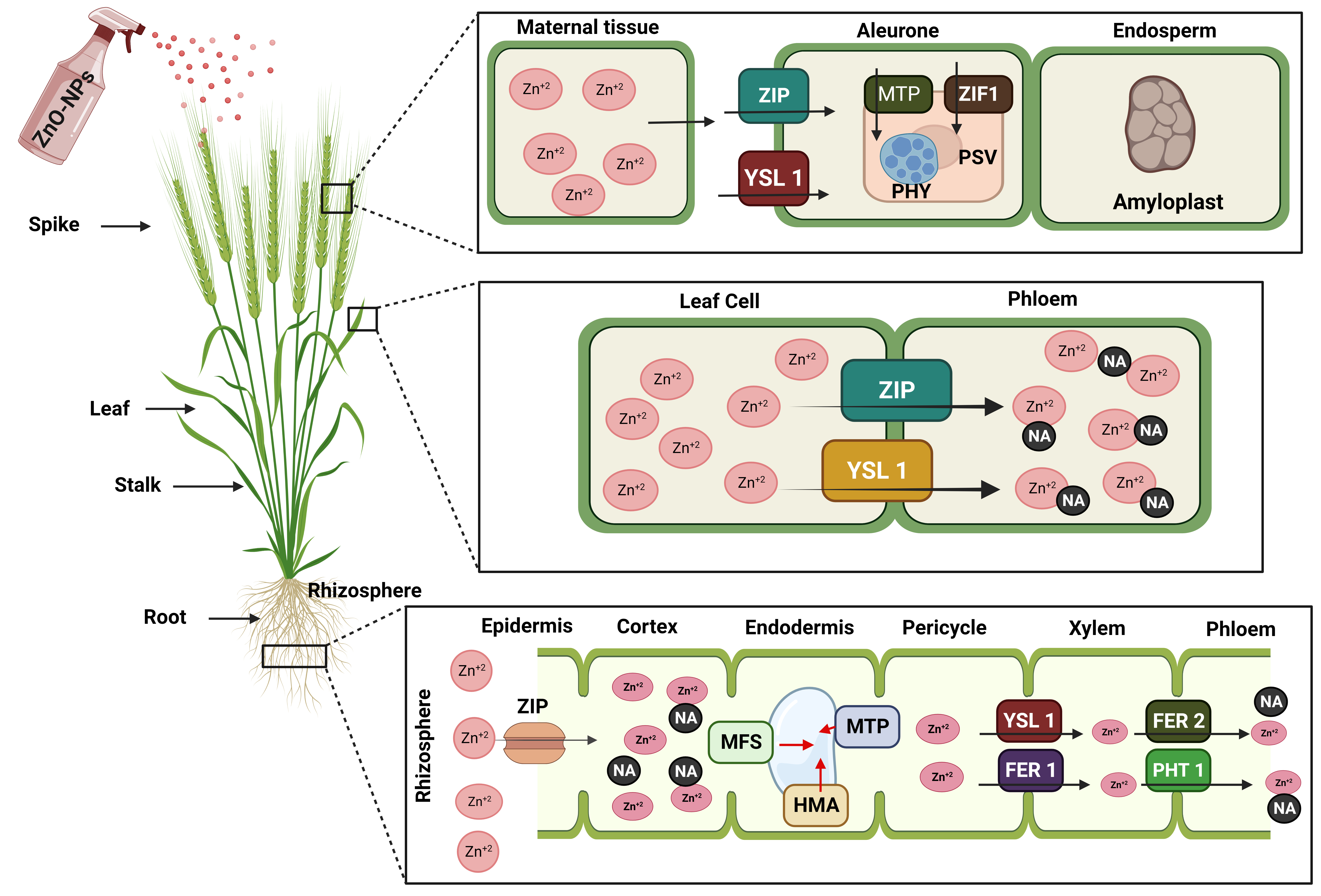
Overall Zn transport from the roots to the shoot is affected by the transport of Zn into the vacuole [198], and metal chelators like nicotianamine (NA) are important for this movement (Fig. 6) [199]. In ZnO-NPs, the zinc ions are transported in the xylem either as a free cation or as a complex with organic acids like citrate [200]. ZIP family proteins are involved in the remobilization of sugars from the leaves during grain filling, as well as the transfer of sugars from the root or basal part of the shoot to the phloem (Fig. 6). Phloem is the only route for nutrient transport in some cereals due to the discontinuous nature of the xylem [201]. Zn is carried through the phloem in complexes with NA or small proteins. Several members of the ZIP and metal tolerance protein (MTP) families have been proposed as transporters from maternal tissue into the endosperm cavity, aleurone, and embryo (Fig. 6) [202].
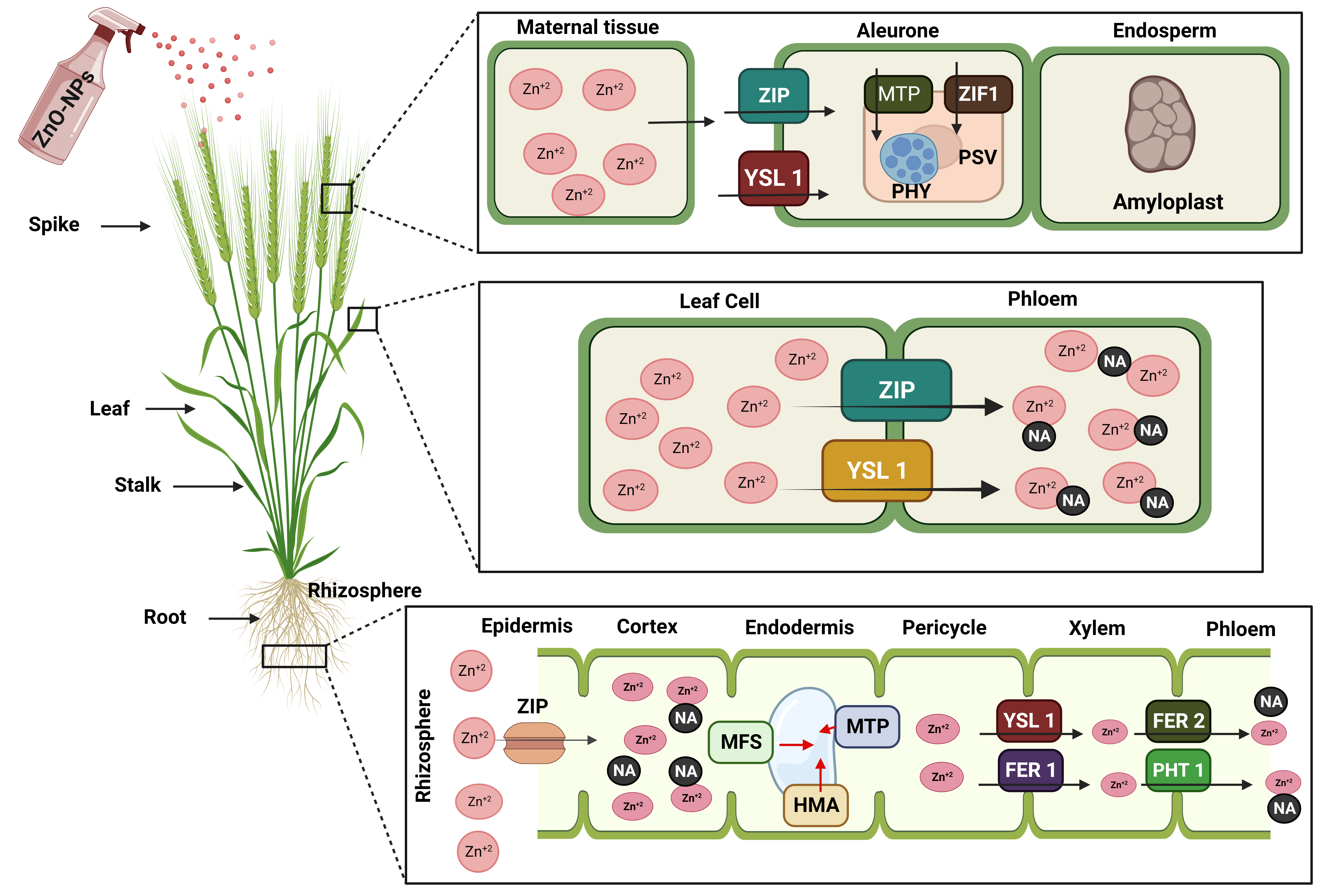 Fig. 6.
Fig. 6.Diagrammatic representation depicting the molecular basis of translocation of ZnO-NPs into Zn supplement in wheat plants. Translocation of Zn from roots to grain via shoots (stalk) assisted by ZIP (ZRT-IRT like protein), YSL (Yellow stripe like transporter), MFS (Major facilitator super family transporter), MTP (Metal tolerance protein), HMA (Heavy metal ATPase), FER (Ferritins transporter), PHT 1 (Phosphate transporter 1), NA (Nicotianamine), PHY (Phytate), and PSV (Protein storage vacuoles).
Nanomaterials are unique materials due to their small size, large surface
area-to-volume ratio, and remarkable optical properties. Owing to these
properties, nano-fertilizers are advantageous, facilitating advancements in plant
growth, nutrient security, and a wide range of farming methods (Fig. 7). There
have been a lot of developments in the field of nanotechnology applications
research recently, and many sectors have started implementing this cutting-edge
technology into their processes to create products. With the increasing human
population, it is essential to expand agricultural output. Fertilizers have been
useful in increasing food production; nanotechnology has enabled the
manufacturing of new and better fertilizer varieties, nano-fertilizers.
Nano-fertilizers are readily taken up by the ground, improving soil quality and
consequently plant growth. Although nanotechnology has many potential
applications, most research and development efforts have focused on electronics,
optical devices, water purification, and healthcare. Owing to the vigorous nature
of nano-size materials and the fact that conventional fertilizers lack the
elements necessary for crop development and nutritional composition, there is a
great scope for developing materials that can carry nano-fertilizers, which can
be utilized as an eco-friendly solution to nutrient-related problems [203]. The
use of Zn-NPs as a way to improve plant uptake is a very recent approach. The
effectiveness of Zn fertilizers is also regulated by particle size. The number of
particles per unit weight and surface area both increase as particle size
decreases, which in turn speeds up the dissolution of fertilizers like ZnO that
are poorly soluble in water. The increased zinc content of shoots in response to
ZnO-NP supplementation has demonstrated the effectiveness of applying Zn-NPs to
increase the zinc content of crops. While fine ZnSO
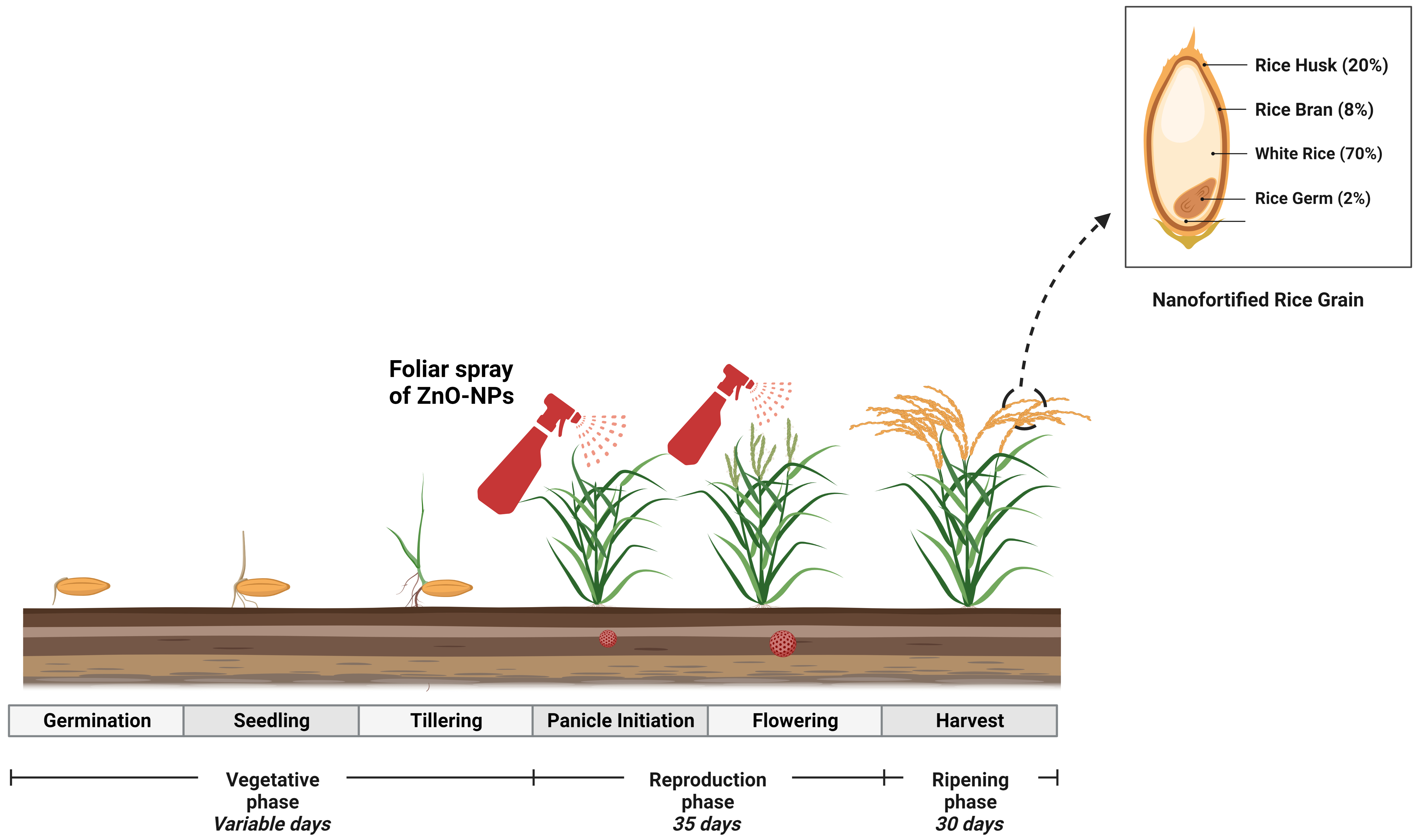
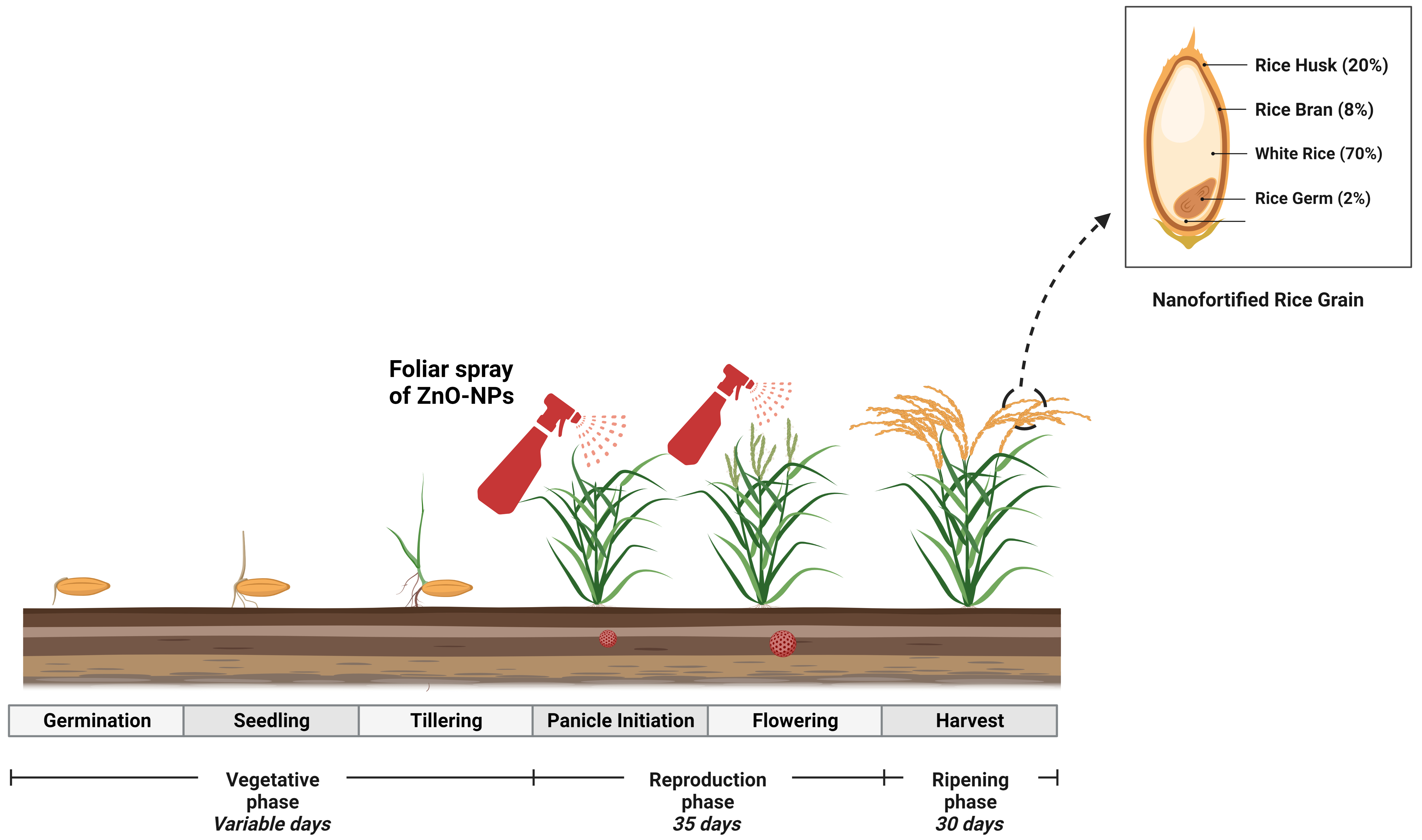 Fig. 7.
Fig. 7.Diagrammatic representation use of ZnO-NP-based fertilizer into rice plants through foliar spray that increased Zn contents into rice grain.
New studies suggest that nanotechnology may one day completely alter our agricultural methods [204]. This opens up the possibility of using agrochemicals with a sophisticated delivery structure that is risk-free, target-specific, and simple to implement. Compared to even the most cutting-edge polymeric-type conventional fertilizers, nano-fertilizers have higher surface-to-volume ratios resulting in increased efficiency. Furthermore, their composition may allow for a slow release, allowing the plants to absorb more nutrients. Therefore, this technology provides a foundation for developing novel and sustainable nutrient delivery systems that utilize the nonporous surfaces of plant parts (Fig. 4B,C). Encapsulated NPs, nano-clays, and zeolites improve fertilizer efficiency and reinstate soil fertility and plant health without causing environmental pollution and agroecological disturbance [205].
Potential ingredients in nano-fertilizers include NPs of zinc oxide, silica,
iron, and titanium dioxide; ZnS/Zn, Cd, Se core-shell quantum dots (QDs); InP/ZnS
core-shell QDs; Mn/ZnSe QDs; gold nanorods; Al
Nano-fertilizers have many advantages over traditional fertilizers, including increasing crop resilience to drought and disease while simultaneously reducing chemical fertilizer use by three times. However, the intensity of plant uptake of NPs from the soil is strongly influenced by particle sizes and shapes. The NPs may need to undergo a chain reaction, including oxidation and recombination before plants can take them up to use as a source of the micronutrients needed to thrive. The nano-size of the nutrients enabled fortification with plants, rendering the micronutrients an attractive prospect. The ability of plants to multiply by storing such nutrients addresses the shortcoming caused by the lack of those nutrients. In addition, specific nutrient deficiencies in plants could be corrected through the use of nano-fertilizers that have been engineered for this purpose. Different arrangements of the atoms on the nanomaterial surface can be used in a variety of ways to produce the desired effects. For effective crop production and the lessening of environmental vulnerabilities related to agricultural activities, zeolites, which are mineral materials, are used to protect the soil by conserving nutrients available in the soil [207]. The newly developed natural zeolite is a nano-fertilizer composed of more than fifty different mineral types of basic alkali and alkaline earth aluminosilicates. This zeolite has been used in the production of maize due to its accessibility and low cost [208]. Being a smart carrier and regulator of principal mineral fertilizers, it also supplies a small quantity of minerals to plants. Zeolite’s capacity to transport nitrogen and potassium implies that less fertilizer can be applied while achieving the same results [209]. The combination of zeolite compounds and humus materials has been shown to increase crop yields. Researchers have successfully developed nanocomposites of zeolite containing nitrogen, phosphorus, and potassium. Plants have been able to absorb various nutrients, including amino acids, mannose, calcium, iron, and zinc, which have all played a significant role in promoting their growth.Due to the limited capacity of zeolite to uptake and store anions, biopolymers are added to improve its performance in this regard [207].
Zinc oxides and sulfates are the most prevalent zinc compounds used to fortify
fertilizers. It would be impossible to overstate the importance of zinc to living
organisms. Zinc is a component of the majority of enzymes that is necessary for
hormone and chlorophyll regulation, and essential for carbohydrate metabolism. In
the form of NPs, ZnO is absorbed, metabolized, and accumulated in plant systems
with relative ease. Zinc is a micronutrient, thus only small amounts are
required. Furthermore, excessive ZnO-NPs concentrations can stunt plant growth
and cause abnormalities in germination, root development, and early-stage
seedling biomass [210]. However, the growth and development of plants may be
stunted if they lack zinc. Due to its insolubility in water, ZnO is combined with
zinc sulfate to create fertilizer. In alkaline soils, crop yields are lower
because plants are more susceptible to zinc deficiency [211]. There is a need for
a large quantity of these micronutrients, making it challenging for application
to the soil in a uniform manner. Unlike zinc, zinc oxide, and zinc sulfate are
only moderately effective when used alone as micronutrients [212]. These are
efficiently utilized along with micronutrients, which act as carriers. While
polyphosphate, ammonia, and anhydrous ammonium nitrate could dissolve 2.0% of
zinc, orthophosphate facilitates the absorption of only 0.05% of zinc; the
carriers play an important role in the solubilization and absorption of the
micronutrients, especially in the case of aqueous solutions [212]. The mineral
composition, ionic strength, and organic matter of soil are critical to nutrient
uptake and bioavailability. Solubility and absorption of zinc are affected by the
presence of calcareous soils, which has an elevated pH and calcium carbonate
(CaCO
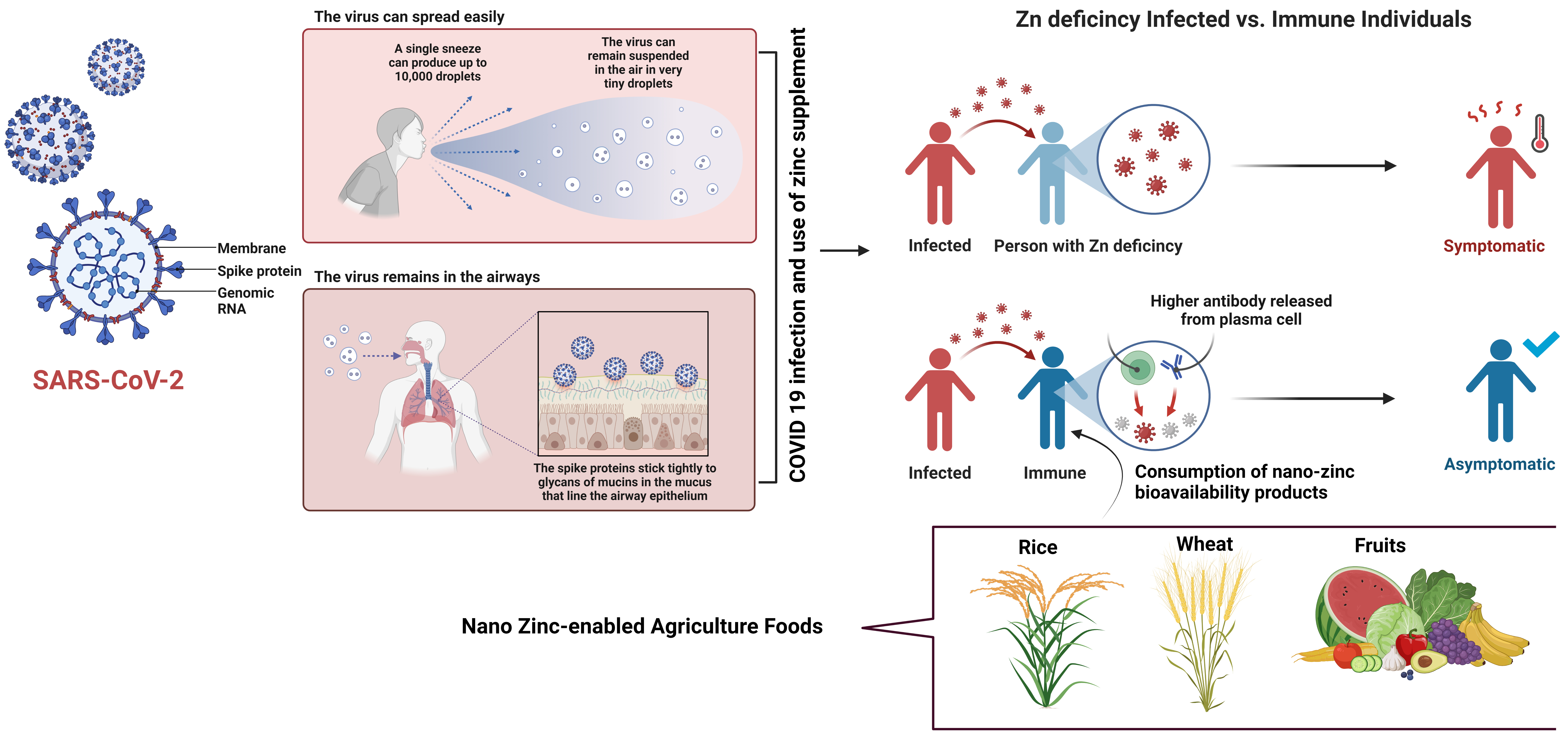
The COVID-19-led severe acute respiratory syndrome is a global pandemic [214]. As a precaution, freedom of movement has been restricted in most parts of the globe. The Centers for Disease Control and Prevention (CDC), National Institutes of Health (NIH), and World Health Organization (WHO) have developed therapeutic guidelines for infected people [215]. However, no universally accessible preventative measure has yet been established besides handwashing and vigilance surrounding commonly encountered oronasal symptoms and fever. The availability of a biologically active substance for use as a preventative measure during a pandemic is crucial. The ideal solution would be affordable, accessible, and low-risk for the majority of the population. The mineral zinc has been shown to inhibit viral replication and stimulate the immune system. Clinical studies are warranted to determine whether zinc consumption (within the recommended upper safety limits) can be used as a prophylactic measure to protect against the development of COVID-19. A low zinc status can affect vaccination responses even after vaccination [10]. Zinc is a vital mineral that facilitates the functions of over a hundred enzymes, and is required for proper immune system operation, synthesis of DNA and RNA, and proper cell integration. Because the body is unable to synthesize or store zinc, it is an important nutrient. Zinc needs to be consumed at a rate of 10 to 15 mg per day by an adult Indian to meet the RDI [216]. A higher incidence of COVID-19 has been linked to immune suppression (Fig. 8). Among these factors is eating a healthy diet, which contributes to a healthy immune system. The antioxidant nutrient zinc has an extensive impact on the immune system. Zinc plays an important role in the maturation and operation of neutrophils and natural killer cells, which mediate innate immunity. Zinc deficiency also has an effect on macrophages, the cells responsible for phagocytosing invading pathogens [216]. There is a correlation between zinc deficiency and the development and activity of T and B lymphocytes. In addition to its anti-inflammatory properties, zinc may also act as an antioxidant. COVID-19, like SARS-COV-2, is caused by an RNA virus. RNA viruses store their genetic information in ribonucleic acid (RNA). The central enzymes of the RNA-synthesizing machinery of RNA viruses are RNA-dependent RNA polymerase (RdRp) enzymes, therefore, RdRps are highly prioritized for antiviral drug development [18, 217].
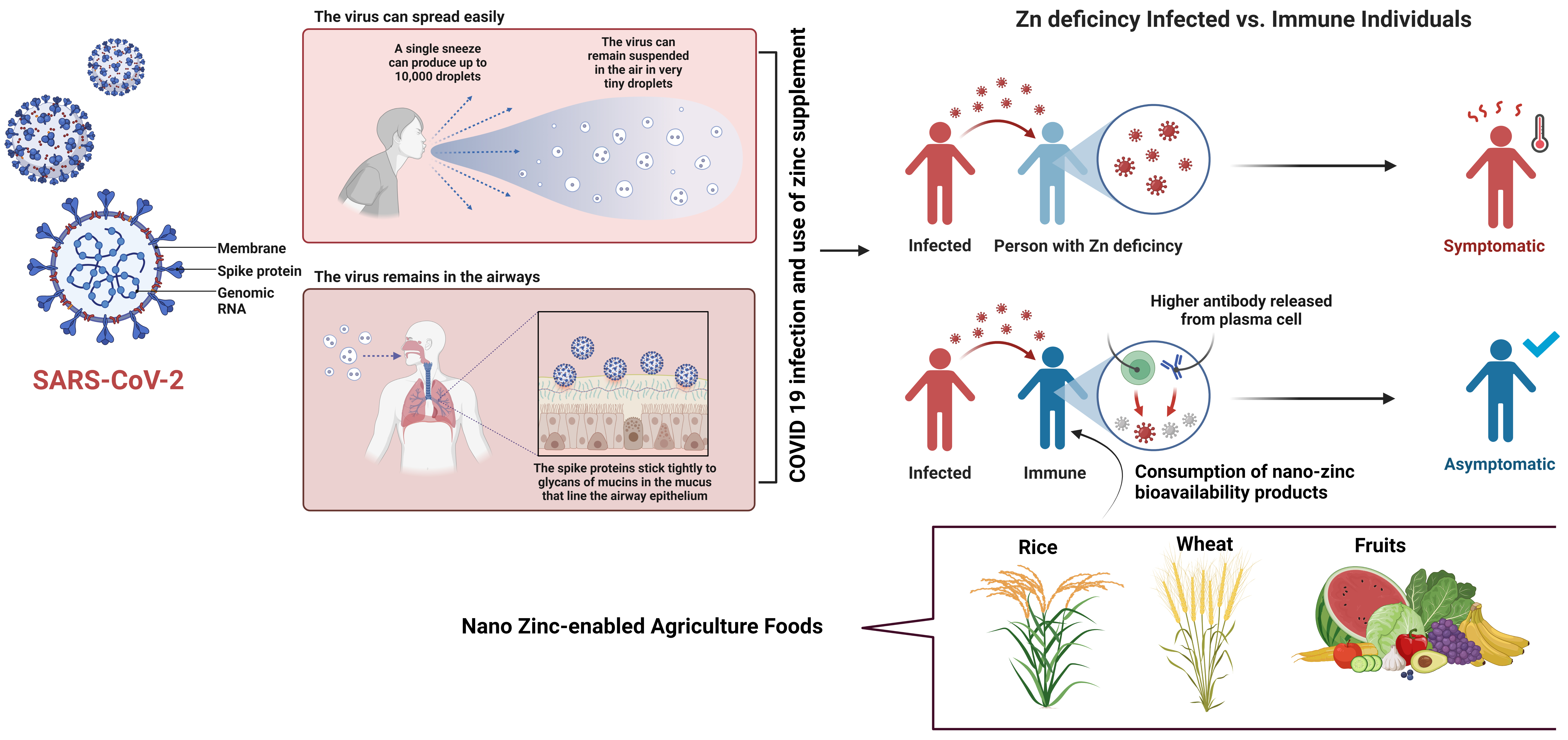 Fig. 8.
Fig. 8.Diagrammatic representation of COVID-19 SARS-CoV-2 virus infection mode, spreading process, and impact of nano zinc enabled agriculture food to boost the immune system.
A recent study shows that intracellular Zn
Zinc deficiency endangers crop production and food safety due to its decreasing accessibility in arable soils worldwide [218]. Low zinc contents, high pH, lower reduction potential, sustained flooding, rhizosphere microbial communities, high organic matter, high iron/manganese oxide contents, high calcium carbonate and bicarbonate contents, high sodium and phosphorus availability, high exchangeable magnesium/calcium ratio, and numerous other factors all negatively impact the available zinc in soils for plant growth [219]. Zn deficiency disrupts protein metabolism because it leads to the breakdown of ribosomes [220]. Low zinc levels inhibit RNA and protein synthesis because zinc is an essential component of several enzymes required for protein synthesis, along with glutamate ribonuclease, ATPase, and dehydrogenase [221]. Zn deficiency reduces the production of protein via reducing the protein synthesis process which varies among plant species. Yellowing immature leaves, reduced leaf size, and wilted stems are the most noticeable symptoms of zinc deficiency in plants under severe Zn deficiency stress; nevertheless, older leaves frequently exhibit leaf bronzing, restricted growth, rolling, and wilting [222]. Phyto-siderophore and organic acid ion generation, arbuscular mycorrhiza symbiosis, and improved resistance to oxidative stress caused by zinc shortage are a few of the defenses plants have evolved to survive [223]. These plant modifications influence the transcription of Zn-required processes and boost zinc uptake and transport mediated by zinc transporters [224, 225]. The complete acceptance and propensity of the agricultural sector to implement nanotechnologies are limited by rising concerns about bioavailability, the toxicity of NPs, and an improper regulatory framework [226]. It is important to note that NPs are a double-edged sword, and improper application of them can have detrimental effects on plant growth by inhibiting the production of proteins and pigments in plants [227]. Plants react differently to high concentrations of NPs at the morphological, physiochemical, and molecular levels [228]. Soil bio/geo transformations of NPs are a key factor in determining their bioavailability and toxicity [226]. In-depth analyses of the variables that influence the phytotoxicity of NPs have been conducted previously [229]. Studies have shown that various factors, such as the type of nano-material, test plants used, NP concentration, NP size distribution, NP morphology, and NP surface charge, play a role in determining the phytotoxicity of NPs. Furthermore, these studies have reported that the mediated effect of NPs on plant growth and development is concentration dependent, with the use of high concentrations of these NPs leading to problematic aftermath for plants, animals, and humans [230].
Plants and the human body require Zn, a micronutrient, in small amounts. However, this trace amount has a major role in the proper functioning of the plant and the human system, as Zn is the cofactor for many enzymes. As the concentration of Zn is low in the grain, it negatively impacts the yield potential and thus impacts food security. To overcome this, the crops are treated with the correct amount of ZnO-NPs fertilizer with careful application timings to increase Zn concentrations within the seeds efficiently. To ensure that everyone has access to zinc-enriched diets, the biofortification of grains and other crops like pulses are great ideas for the human population worldwide, more specifically for those in rural areas. The nano-biofortification strategy is a tool with great potential for combating malnutrition. This method, especially in its biological nano-form, shares some of the benefits of nano-fertilizers, such as efficiency and reduced usage. Nano-biofortification is still in its early stages and requires additional testing before being implemented worldwide.
The article contains no such material that may be unlawful, defamatory, or which would, if published, in any way whatsoever, violate the terms and conditions as laid down in the agreement.
AS, VDR and KG designed the review idea study. DP and RS performed the editing of the article. AS and VDR make all the figures. AS, VDR, KG and TM analyzed the data. AS, VDR and KG interpreted the data, and AS, VDR and KG wrote the manuscript. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
We acknowledge the late Dr. Kirti Singh, a well-renowned agriculture scientist, for their invaluable contributions and pioneering research in the field of agriculture science. Their dedication and passion for advancing agriculture science have left a lasting impact on the agricultural community and beyond. We extend our deepest gratitude for their significant achievements and will continue to be inspired by their work as we pursue further advancements in agriculture research. May their legacy continue to inspire future generations of scientists.
VDR and TM are supported by the Strategic Academic Leadership Program of the Southern Federal University (“Priority 2030”) and the Ministry of Science and Higher Education of the Russian Federation (no. FENW-2023-0008). KG are supported by internal grant of Yerevan State University.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.