
Background: urban forest in coastal cities encounters multiple disturbances of frequent typhoon events caused by global change, under which ecological remediation can help to improve urban environment. We measured and analyzed the growth and ecosystem services of four newly-planted tree species in Zhuhai after Typhoon Hato (2017), aiming to evaluate the efficiency of the ecological remediation. Methods: National Meteorological Information Center of China supplied climate variables. From June 2018 to December 2019, we measured soil physical and chemical properties, above- and below-ground development regarding stem, tree height, and root growth of all the selected tree species. Results: Sl (Sterculia lanceolata Cav.), Ir (Ilex rotunda Thunb), Ss (Schima superba Gardn. et Champ.) could be more wind-resistant from the above-ground morphological perspective. For the below-ground process, Sl was the only tree species with continuous development, while Ir, Ss, and Es (Elaeocarpus sylvestris (Lour.) Poir.) decreased. Furthermore, Sl, Ir, and Ss maintained their investment in deep roots when Es had apparent deep root biomass reduction. The edaphic condition showed notable improvement in chemical properties rather than physical properties, especially for AN (available nitrogen), AK (available potassium), and SOM (soil organic matter). Conclusions: The ecological remediation in Zhuhai after Typhoon Hato (2017) was efficient, and in the future, tree species like Sl with advantages in root development and morphological profile were preferentially recommender for plantation in typhoon-affected areas.
The last decade has witnessed robust evidence that global warming progressively affected human society and natural ecosystems, which also increased the periodicity and intensity of extreme climate events such as heat waves, droughts, tornadoes, and hurricanes [1, 2, 3]. Extreme climate events, altering ecosystem structure and function well outside normal variability, attracted increasing attention as drivers of change in ecological and evolutionary communities [4, 5, 6]. For example, Stocker et al. (2019) [7] applied satellite retrievals of information on the earth’s surface to examine the ecological impacts of droughts on global terrestrial photosynthesis and primary production. Elsner et al. (2019) [8] quantified the magnitude of the power of tornadoes in consideration of diurnal and seasonal influences embedded within natural variations in the US for the recent 20 years. Besides, it was predicted that El Nino Phenomenon associated with sea-level rise might lead to changes in the frequency and intensity of extreme coastal flood events in Latin America [9]. Researches also developed predictive models to predict how to reduce wind damage in Austrian forests by analyzing remote sensing images [10].
Researches have consistently been conducted on the impact of various extreme climate events on human society or ecosystems, however, the efficiency of people’s subsequent ecological remediation was scarcely investigated [11]. Furthermore, it might not be comprehensively and thoroughly answered whether the ecological remediation could enhance our capacity of coping with similar extreme climate events in the future from the perspectives of tree physiological development in China [12]. For example, Zhuhai, a coastal city located in southeastern China, frequently confronts typhoon events every several years. Although the local ecological department invariably implemented ecological remediation after every typhoon left, lack of evaluation of the quality of their remediation coupled with neglect of subsequent adaptation could probably expose the city to typhoon damage again, partly resulting in exponentially increasing economic losses as well as rising numbers of injuries. Therefore, pertinent evaluation and specific recommendations could be positive efforts for those typhoon-suffering areas to enhance their abilities of resisting multiple natural disturbances.
Urban forests and trees, providing multiple ecosystem services such as improving urban air quality, reducing noise, attenuating storm-water flooding, and conserving energy, could be a key component in the adaptation of cities to climate change [13, 14, 15]. In addition, the plantation of urban trees was generally considered to effectively help cities acclimate to extreme climate events, especially for ecological remediation after typhoon events [16]. This was because urban trees played a crucial role in promoting both above- and below-ground processes such as recovering in soil fertility, water conservation, construct and improve environmental conditions, which was most likely damaged or affected by fierce winds and intense rainfalls [17, 18, 19]. Hence, detailed information regarding the development of those urban trees planted for ecological remediation could contribute potently to a profound understanding of the efficiency of remediation and future prediction.
Zhuhai city, suffering severely from Typhoon Hato (2017), implemented imperative ecological remediation via tree species selection and urban tree plantation in June 2018. Nevertheless, the remediation was principally from political perspectives and carried out by the local administrative department, which might lack scientific assessment, hampering a profound understanding of the quality of the remediation progress. Therefore, we selected the remediation district encountering Typhoon Hato as our objected area and established both above- and below-ground measurements for the newly-planted tree species. Additionally, with environmental data, we expected to answer questions as following: (1) whether did the newly-planted trees adapt to the local environment regarding their growth patterns? (2) How did the selected tree species develop their fine root biomass in vertical and horizontal levels? (3) Under the ecological remediation, was the edaphic condition improved in terms of physical and chemical properties? (4) what kind of tree species could be recommended in typhoon-affected areas?
Located in the southeastern coastal areas of China, Zhuhai is frequently
confronted with extreme climate events as typhoons’ disturbance (Fig. 1). On 20th
August 2017, Typhoon Hato generated upon the northwestern Pacific Ocean and
constantly intensified within 72 h. At 12:50 on 23rd August, it landed on
Zhuhai at wind scale 14 and moved forward in the northwest direction. During its
route in Zhuhai lasting over 4 h, more than 25% of urban trees in
quantitative terms in Zhuhai were damaged by the strong wind. After Typhoon Hato
left, imperative ecological remediation was planned and launched according to the
practical condition in each area. In Xiangzhou district, which suffered
enormously from the typhoon disturbance, the local forestry administration
department took several steps to recover and re-build urban greening, including
sweeping typhoon-affected areas, restoring land and soil, and making ecological
restoration plans. In June 2018, 18 indigenous tree species were selected and
over 50,000 saplings were planted. To analyze their growing conditions that
helped to evaluate the quality of the imperative ecological remediation, we
selected four most-planted tree species as our objects within the whole planting
area, i.e., Sterculia lanceolata Cav. (Sl), Ilex
rotunda Thunb (Ir), Schima superba Gardn. et Champ.
(Ss), and Elaeocarpus sylvestris (Lour.) Poir. (Es).
The four tree species are all native tree species with flexible soil adaption. In
order to select the most representative trees of the four tree species, five
sampling plots with the size of 30
 Fig. 1.
Fig. 1.Schematic of Zhuhai in China (blue area with red framework), landing location and route of Typhoon Hato (broad red arrow), and location of our study area in Zhuhai (green area).
National Meteorological Information Center of China (http://data.cma.cn)
supplied climate data, based on which monthly climatic variables including wind
velocity (km h
The primary soil type in Zhuhai is red acidic soil. The soil sampling campaign
was launched in the study area in June 2018 and December 2019 to indicate the
development of soil physical and chemical properties. For each time, five
metallic-cylindrical sampling cores were used to dig out two kilos of soil in the
depth of 30 cm, which were randomly located in the study area same as the
sampling plots. Then all the soil samples were well kept in sampling bags and
transported to our laboratory for further analysis. The soil’s physical
properties were measured according to Li and Shao’s reports [20]. Soil bulk
densities (BD, g cm
where ds is the soil density (mg m
For soil chemical properties, soil PH was determined in 1 : 2.5 soil-water slurry
using a combination glass electrode. Soil organic matter (SOM, g
kg
In June 2018, the saplings of all the tree species were simultaneously planted in the study area, which had unified short and small sizes (height around 25 cm and DBH around 0.5 cm). Therefore, the four tree species were considered to have the completely same and tiny initial conditions. To investigate their above-ground growth, their ground diameters were measured with the help of a vernier caliper (Altraco Inc., Sausalito, California, USA) and their height was measured using a standard tape. The measurement was conducted in December 2018, June 2019, and December 2019 to explore the above-ground development patterns from 0M to 18M.
To clarify the below-ground progress, fine root coring campaigns were launched
for all the selected trees at the same time as the above-ground measurement. A
pre-test coring campaign showed that the range of the root system was similar to
a cylinder, with a diameter of 70 cm and a height of 35 cm. Therefore, for the 16
trees, eight soil cores were collected for every individual tree: four at a
distance of 15 cm from the trunk and the other four at 30 cm. The soil was
sampled down to a depth of 30 cm using a soil auger with a length of 30 cm and a
radius of 3 cm. Each sample was divided into three horizons: soil depths of 0–10
cm (upper layer), 10–20 cm (middle layer), and 20–30 cm (deep layer). Fine
roots (
The software SPSS 22.0 (IBM, State of New York, USA) was used for statistical
analysis. To investigate the difference between means, two-sampled
t-test and analysis of variance (ANOVA) with Tukey’s HSD (honestly
significant difference) test were used. In all the cases, the means were reported
as significant when P
Five physical and eight chemical variables were measured to analyze the soil
revolution under the ecological remediation (Table 1). For physical properties,
no significant difference was detected between June 2017 and December 2019, among
which SMC and BD exhibited slight decreases (from 22.64%
| Soil properties | n | Date | ||
| June 2018 | December 2019 | |||
| Physical properties | SMC (%) | 5 | 22.64 |
19.90 |
| Stp (%) | 5 | 39.80 |
45.72 | |
| Scp (%) | 5 | 35.66 |
39.31 | |
| Sncp (%) | 5 | 4.14 |
6.41 | |
| BD (g cm |
5 | 1.28 |
1.23 | |
| Chemical properties | TN (g kg |
5 | 1.02 |
1.01 |
| TP (g kg |
5 | 0.26 |
0.31 | |
| TK (g kg |
5 | 15.40 |
21.52 | |
| AN (mg kg |
5 | 47.78 |
152.60 | |
| AP (mg kg |
5 | 1.08 |
1.09 | |
| AK (mg kg |
5 | 18.18 |
85.80 | |
| PH | 5 | 4.20 |
5.04 | |
| SOM (g kg |
5 | 10.36 |
18.44 | |
| Soil moisture content (SMC, %), soil total porosity (Stp, %), soil capillary
porosity (Scp, %), soil non-capillary porosity (Sncp, %), bulk density (BD, g
cm | ||||
Under the same initial condition (all the four tree species were short and small
saplings.), the four tree species displayed different growing patterns regarding
tree height and ground diameter (Fig. 2). Es had the most rapid growth
of tree height for the whole period and reached 2.98
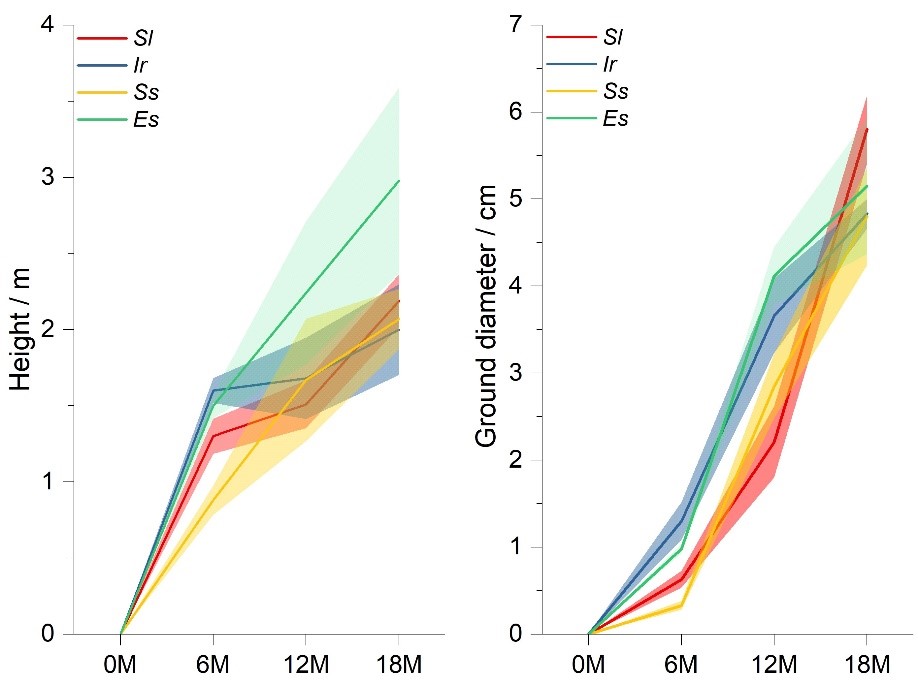 Fig. 2.
Fig. 2.Development of tree height (m) and ground diameter (cm) of Sl (red line), Ir (blue line), Ss (orange line), and Es (green line) from June 2018 (0 months, 0M) to December 2019 (18 months, 18M), around which the light red, blue, orange, and green shading areas are the 95% confidence intervals, respectively.
Four tree species displayed different fine root growth patterns for the whole
period (Fig. 3). During the first six months, they showed no significant
differences (19.64
 Fig. 3.
Fig. 3.Development of total fine root biomass (g m
To clarify the vertical fine root growth for the whole period, we divided them
into three layers, i.e., 0–10 cm (shallow layer), 10–20 cm (medium layer), and
20–30 cm (deep layer) (Fig. 4). From December 2018 to June 2019 (6M to 12M),
fine root from the three different layers of all the four tree species obtained
rapid and dramatic development (P
 Fig. 4.
Fig. 4.Vertical development of fine root biomass (g m
Fine root biomass from 15 cm (nearby root) and 30 cm (outmost root) to trunk
provided us a perspective on the root’s horizontal development (Fig. 5). From 6M
to 12M, Sl, Ir, and Ss had more root growth at 15 cm
than 30 cm, e.g., the nearby root of Ir (229.06
 Fig. 5.
Fig. 5.Horizontal development of fine root biomass (g
m
Numerous studies verified that above-ground growth patterns could reflect trees’ health condition, geographic adaptation, and supplying ecosystem services [22, 23, 24]. In our research, the growth of tree height and ground diameter for the four tree species were measured from 0M to 18M. Although all of them exhibited a general increase, different periodic patterns implied trees’ particular above-ground strategies. Es was distinguished for its height growth among the four tree species, implying a robust above-ground construction coupled with its second-highest value of ground diameter, which was in line with the previous findings [25, 26]. On the contrary, the other three tree species exhibited an inclination to develop ground diameter than height. Their height growth primarily occurred from 0M to 12M and slowed down from 12M to 18M, while the ground diameters increased sharply from 6M to 18M. Such growth pattern indicated a stable above-ground morphological profile, which could effectively provide wind-damage resistance that lower but thicker trees might now easily break down in typhoon or hurricane events [27]. This might also be verified by using convolutional neural networks to estimate the relationships between tree failures due to high winds and tree sizes [28]. As showed in Fig. 2, preferential growth of Sl was distinctly given to ground diameter to effectually make the above-ground tree profile stable, which could be expected to have higher survival rates under typhoon disturbances in the future.
As the primary pathway for water and nutrient uptake, fine roots were a prominent sink for carbon acquired in terrestrial net primary productivity, tightly related to multiple ecosystem services [29, 30, 31]. In our research, we not only measured the total fine root biomass but also the root biomass development in both horizontal and vertical levels, aiming to investigate their absorption capacity of water and nutrient and morphological patterns.
For total fine root biomass, Sl was the only tree species growing within the whole 18 months, showing a persistent below-ground investment. Distinct growth occurred within the first year since plantation, however, the other three tree species showed a biomass reduction from 12M to 18M. Combined with their height and ground diameter growth, it could be explained by their specific allocation strategy between the above- and below-ground processes. Generally, urban trees dynamically adjusted the above- and below-ground investment on the basis of their characteristics and environmental conditions [32, 33], such as enhancing above-ground growth for more photosynthesis or increasing root growth for water and nutrients uptake [34, 35, 36]. Nevertheless, Es’s remarkable decrease of fine root biomass showed a weakened below-ground growth, which could be deeply worried about its adaption in the future. This was because typhoons usually broke down or shaded tree trunks, leading to more severely damaged root systems. In addition, site conditions in typhoon-affected areas were generally fragile and negative that soil quality were highly reduced and ecosystem flow were disturbed, making the fine roots require a lengthy period for recovery [36, 37].
For fine root biomass in the vertical level, the four tree species had
significant growth from 0M to 12M (Fig. 4). Similar decreasing patterns from 12M
to 18M were found as the total fine root biomass, however, several positive
phenomena existed in their deep root layer, i.e., 20–30 cm. As shown in Fig. 6,
Zhuhai usually had more than six months with precipitation lower than 100 mm
every year and its highest temperature from May to October reached over 30
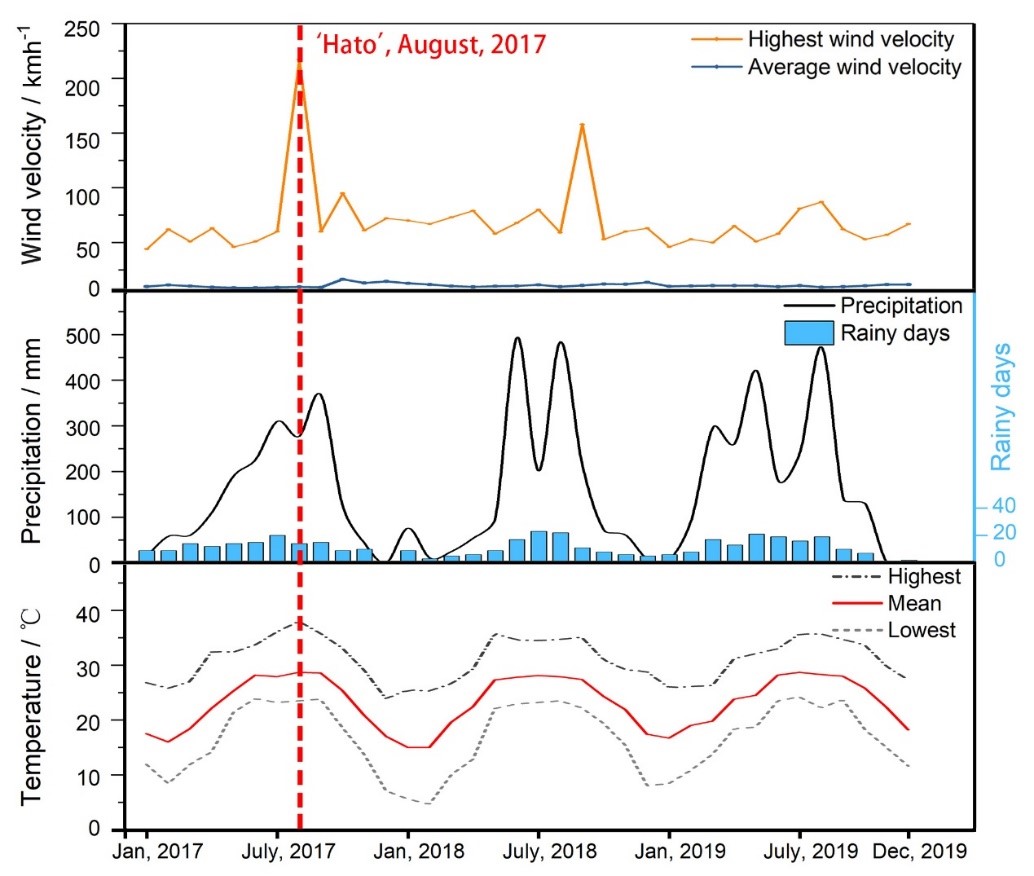 Fig. 6.
Fig. 6.Monthly climate variables including highest wind velocities (km
h
Roots’ growth in horizontal level was closely relevant to its stabilization, which was essential for those trees confronting typhoon disturbance [41, 42]. In general, trees with higher proportion of outmost roots would be more stable than those with stem-centered roots from the morphological perspective [43]. Nevertheless, the outmost roots might be abandoned priorly when the whole roots system faced shrinkage, such as Es from 12M to 18M. From this point of view, it could support that Ss tended to be steadier in a typhoon event among the four tree species as it had relatively balanced investment on both nearby and outmost fine roots within the whole study period, even when fine roots died.
Vegetation was generally regarded to have substantial potential in soil
restoration for cities suffering from extreme climate events [44, 45, 46]. Comparing
the edaphic conditions in June 2017 and December 2018, the impact of the
ecological remediation on local soils could be evaluated in terms of physical and
chemical properties. For physical properties, all the variables showed no
significant differences that SMC and BD slightly decreased and
Stp, Scp, and Sncp increased to a small extent,
implying the ecological remediation rarely influenced the edaphic physical
structure. Regarding chemical properties, all the variables displayed escalation,
notably for AN, AK, and SOM (P
Accounting for the complicated interaction between the restricted urban space (compacted soil, complex microclimate, multiple human activities) and the fragmentized environment under the extreme climate events’ disturbance [49, 50, 51, 52], the efficiency of ecological remediation was essential and urgent from different perspectives. Urban trees could mitigate environmental degradation and provide multiple ecosystem services, specifically for cities encountering extreme climate events, which played a central role in ecological remediation [13]. Their above- and below-ground processes were closely related to their survival rates, morphological growth, geographic adaption, water absorption, and edaphic improvement, which primarily reflected the quality of the ecological remediation [53]. In this study, the selected four tree species showed no deaths but growth, together with which edaphic chemical variables exhibited several improvements. It could be inferred that the ecological remediation in Zhuhai after Typhoon Hato (2017) was effective. However, it still could be improved according to specific growth conditions of different tree species. For example, Ir, Ss, and Es reduced fine root biomass from 12M to 18M, which could be paid more attention to, such as specific fertilization to promote their root growth in this period. We observed that Sl had a relatively thicker tree trunk accompanying steady development of total and deep fine root biomass. This could be inferred that Sl had morphological and physiological resistance to typhoon events, which could be recommended for future plantation in Zhuhai or be preferentially considered for future ecological remediation to obtain promoted ecosystem services.
After Typhoon Hato (2017) intruded on Zhuhai city and severely damaged its urban environment, the local administrative department planned and launched imperative ecological remediation via planting plenty of trees. Aiming to evaluate the efficiency and make relevant suggestions, we measured the above- and below-ground growth of four tree species (Sl, Ir, Ss, Es) together with the edaphic evolution in terms of physical and chemical properties.
For the above-ground process, Es had advantageous growth with the highest value of tree height and second-highest value of ground diameter. The other three tree species exhibited inclinations to develop ground diameter than height, which were probably more wind-resistant from the morphological perspective. For the below-ground process, Sl was the only tree species that obtained continuous development, while Ir, Ss, and Es decreased from 12M to 18M. Besides, Sl, Ir, and Ss maintained their investment in deep roots when Es had significant deep root biomass reduction. In the horizontal level, Ss had the highest proportion of outmost roots among the four tree species.
After 18 months since the ecological remediation started, the eight physical variables showed minor changes, implying the ecological remediation rarely influenced the edaphic physical structure. However, all the variables displayed escalation, notably for AN, AK, and SOM, which could be attributed primarily to fine roots’ participation in edaphic chemical evolution and promote the release of microelement.
In conclusion, the ecological remediation effectively improved the environmental quality via the plantation of various trees in Zhuhai. In the future, the selection of planted trees could be more reliable through a comprehensive analysis of the above- and below-ground processes. Those tree species like Sl with advantages in root development and morphological profile are preferentially recommender for plantation in typhoon-affected areas.
CZ: conception and design; acquisition of data; analysis and interpretation of data; drafting the manuscript; making figures and tables. WQ, LS and DX: acquisition of data; making figures and tables; acquisition of funding. QZ: Acquisition of funding; Project administration.
Not applicable.
We greatly appreciate Dajun Guo, Xiu Chen, Qifei Pan, Weiyi Huang, Yuqi Li, Qing Yang, Qiuxia Pan, and Rongchang Nie for their assistance in the collection and calculation of fine root biomass, tree measurement, and soil coring campaign.
This research was funded by the Forestry Science and Technology Innovational Specific Project of Guangdong Province, grant number (2018KJCX029) and (2020KJCX006), and by the Forestry Science and Technology Program of Guangdong Province, grant number (2019-21).
The authors declare no conflict of interest.