





1 Behavioral Neuroscience Branch, National Institute on Drug Abuse–Intramural Research Program, National Institutes of Health, DHHS, Baltimore, MD 21224, USA
Abstract
Peripheral vasoconstriction is a centrally mediated physiological effect known to play an important role in regulating body temperature by adjusting heat exchange with the external environment. However, peripheral vasoconstriction as a component of sympathetic activation also occurs following exposure to various salient stimuli and during motivated behavior at stable environmental temperatures. This review aims to consider available evidence suggesting a significant contribution of this peripheral effect to physiological increases in both brain temperature and entry of oxygen and glucose into the brain’s extracellular space. While these effects are triggered by neuronal activation, constriction of blood vessels in the skin and most internal organs results in redistribution of blood from the peripheral to central domains, thus dilating cerebral vessels, increasing global cerebral blood flow, and enhancing the intra-brain entry of oxygen and glucose from arterial blood. This powerful influence appears to determine the long duration of physiological increases in both brain temperature and brain levels of glucose and oxygen and their basic similarity across different brain structures. This work underscores the tight interrelationship between the brain and periphery and a significant contribution of cardiovascular effects in providing the enhanced inflow of oxygen and glucose into brain tissue to prevent metabolic deficit during functional neural activation.
Keywords
- Brain temperature
- Brain oxygen
- Brain glucose
- Neural activation
- Peripheral vasoconstriction
- Central vasodilation
- Hyperoxia
- Hyperglycemia
- Hypoxia
- Rats
Peripheral vasoconstriction is a centrally mediated physiological effect known to play an important role in regulating body temperature [1, 2, 3, 4, 5]. Peripheral vasoconstriction is the narrowing of blood vessels resulting from the contraction of muscular walls in large arteries and the small arterioles of skin and most internal organs. This effect is opposite to vasodilation, the widening of blood vessels resulting from the relaxation of smooth muscle cells in vessel walls, particularly in large veins, large arteries, and smaller arterioles of skin and some internal organs. Changes in the tone of peripheral blood vessels result in changes in vascular resistance, consequently affecting tissue blood flow and arterial blood pressure [2, 3, 5]. Following exposure to low ambient temperatures, vasoconstriction decreases heat dissipation from skin surfaces, thus preventing life-threatening hypothermia. Peripheral vasoconstriction is also an important factor for body heat accumulation during fever—a transient temperature increase during bacterial or viral infection. Its increase is also important for controlling hemorrhages and reducing blood loss during traumatic damage. While the role of peripheral vasoconstriction is well established in thermal physiology, the goal of this paper is to discuss the large body of experimental evidence that suggests a more global role of this autonomic effect in shaping adaptive changes of brain activity and brain homeostatic parameters. The data discussed in this work were obtained in different animal species, primarily in rats, and special focus was on thermorecording and neurochemical studies conducted in our laboratory during the last ten years.
Temperature is a basic homeostatic parameter, with temperature stability being
an essential condition for a normal organism’s functioning. Since all
physicochemical processes in the brain are temperature-dependent, temperature
stability is especially important for the brain. While thermophysiology is
focused primarily on the mechanisms that underlie stability of body temperature
following decreases and increases in external (ambient) temperature, brain and
body core temperature in awake, freely moving organisms at stable environmental
temperatures is not stable and fluctuates within 2–4
Since most direct thermorecording studies were conducted in rats, it is still
unclear whether such large temperature fluctuations occur in other animals and
humans, which have more sophisticated mechanisms regulating body temperature. It
is likely because temperature responses induced by arousing stimuli in cats [8]
and monkeys [9] share basic similarities with those observed in rats. Like rats,
body temperature recorded in humans during intense physical exercise [10] and
marathon run [11] can reach 39.5–40.5
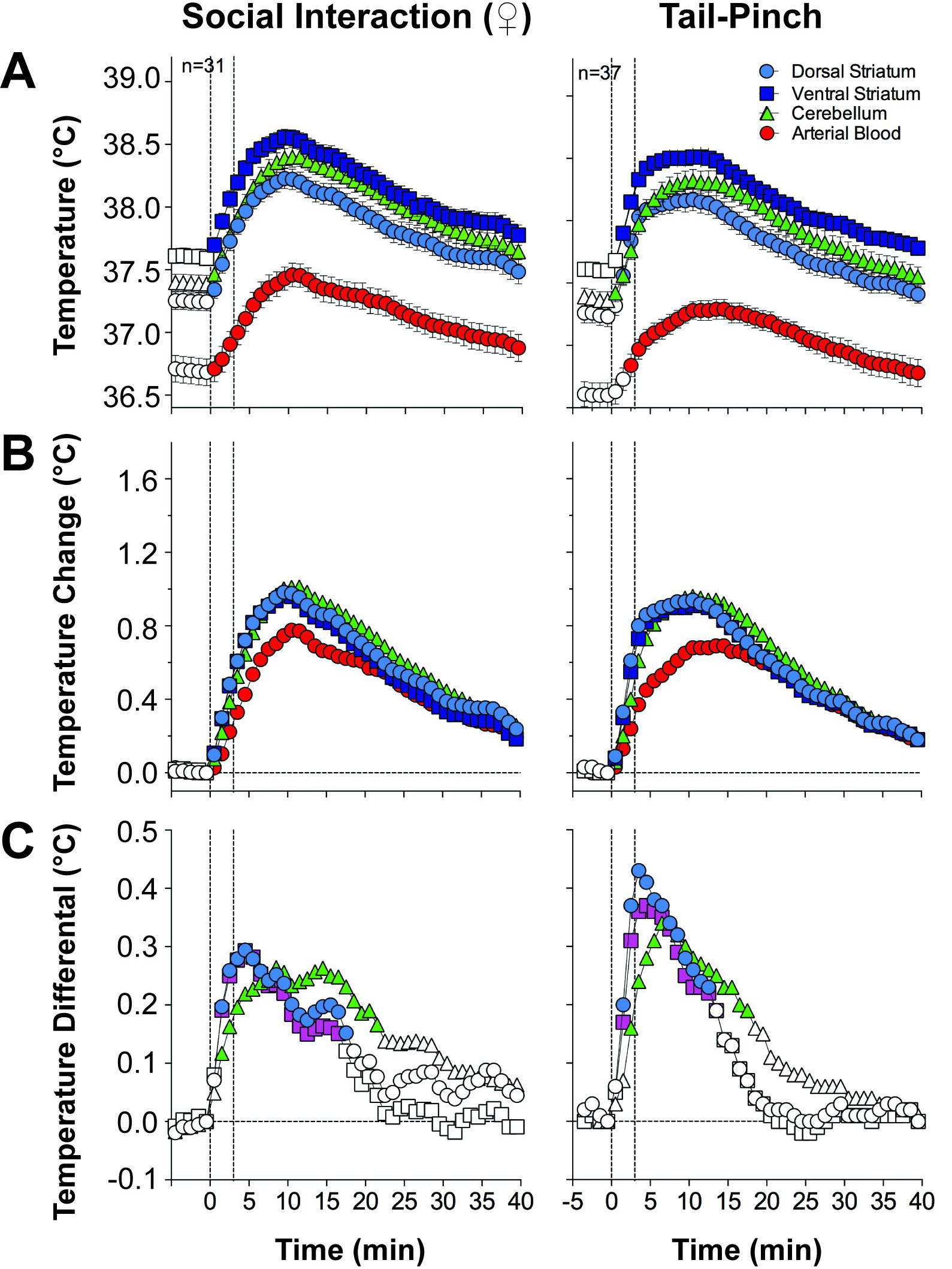
Although physiological hyperthermia is a well-known phenomenon, its functional significance and underlying mechanisms are still controversial. Since brain tissue is highly vascularized and receives a significant portion of total arterial blood inflow, it is often speculated that brain hyperthermia results from the inflow of warmer arterial blood from the body core [14, 15, 16]. However, direct monitoring of arterial blood temperature in conjunction with brain temperature in freely moving animals does not support this mechanism [8, 9, 17, 18, 19]. In our early studies, we implanted miniature thermocouple sensors in the abdominal aorta and a number of brain structures. We examined temperature responses to several mild arousing stimuli in freely moving rats [20]. These experiments revealed that arterial blood temperature is consistently lower than in any brain structure, and changes induced by arousing stimuli are consistently slower than those in brain structures (Fig. 1, Ref. [20]). Due to more rapid temperature increases in the brain, the brain-blood differential significantly increased, suggesting the brain’s heat-producing metabolic activity as the initial source of brain hyperthermia. Importantly, temperature responses detected in different distantly located structures (i.e., nucleus accumbens, or NAc, dorsal striatum and cerebellum) had similar patterns despite differences in basal temperatures.
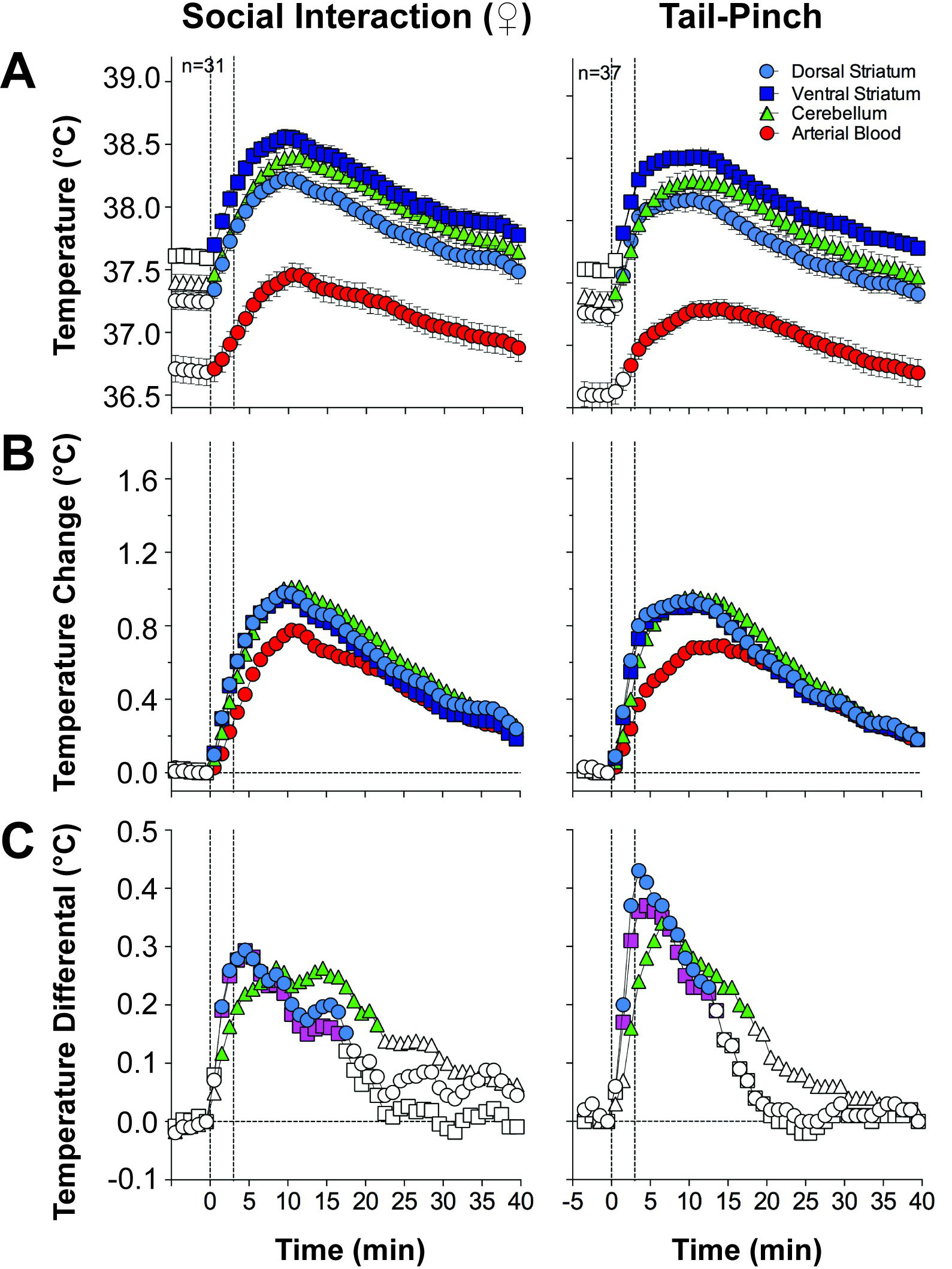 Fig. 1.
Fig. 1.Changes in temperature in several brain structures and arterial blood induced by tail-pinch and male-female social interaction in freely moving rats. Top graphs (A) show changes in absolute temperature. Middle graphs (B) show relative temperature changes. Bottom graphs (C) show temperature differences between brain sites and arterial blood. Filled symbols show values significantly different from the pre-stimulus baseline. Original data were presented in [20].
In contrast to general beliefs that temperature is a slow-changing parameter, high-speed monitoring revealed that brain temperature increases induced by arousing stimuli are rapid, starting with the onset latencies of about 5–10 s and reaching significance at 20–30 s (Fig. 2, Ref. [20]). In contrast, increases in arterial blood temperature were consistently slower, showing statistical significance at 50–80 s after the stimulus onset.
Hence, (a) the source of heat production responsible for functional brain hyperthermia is in brain tissue, implicating increased brain metabolism as its cause; (b) inflow of arterial blood is the primary factor for cooling brain tissue and removal of metabolism-produced heat to the body and the external environment; (c) the pattern of temperature responses in different brain structures is surprisingly similar, implicating a global factor affecting the brain as a whole.
The brain’s metabolic activity is an evident source of enhanced heat production. However, heat could also be accumulated in brain tissue because of its decreased dissipation from brain tissue. To clarify this two-component mechanism, we used three-point thermorecording, with sensors chronically implanted in a brain structure, temporal muscle, and subcutaneous (SC) space [21].
As shown in these experiments, temperature increases in brain structures were more rapid and stronger than in temporal muscle (Fig. 3, Ref. [21]). Since temporal muscle is not involved in locomotion and has similar brain blood inflow from the carotid artery, the brain-muscle differential removes the influence of arterial blood temperature, thus providing a measure of intra-brain heat production. In contrast to the relatively long-duration of temperature increases in both the brain structure (here NAc) and temporal muscle, the brain-muscle differential increased rapidly but transiently with a tendency towards rebound-like decreases. In contrast to monophasic temperature increases in brain structures and muscle, biphasic down-up changes were found in the SC space—a densely vascularized location. Skin temperature also depends on two variables: the vessel tone and the temperature of incoming arterial blood. By calculating the skin-muscle differential, the influence from the latter factor is eliminated, providing an accurate measure of skin vascular response. This parameter decreased rapidly, stronger, and for a longer time than brain-muscle differential, suggesting robust skin vasoconstriction as an important contributor to functional brain hyperthermia. Like changes in brain-muscle differential, the skin-muscle differential also showed rebound-like increases, suggesting post-constrictive dilation of skin vessels.
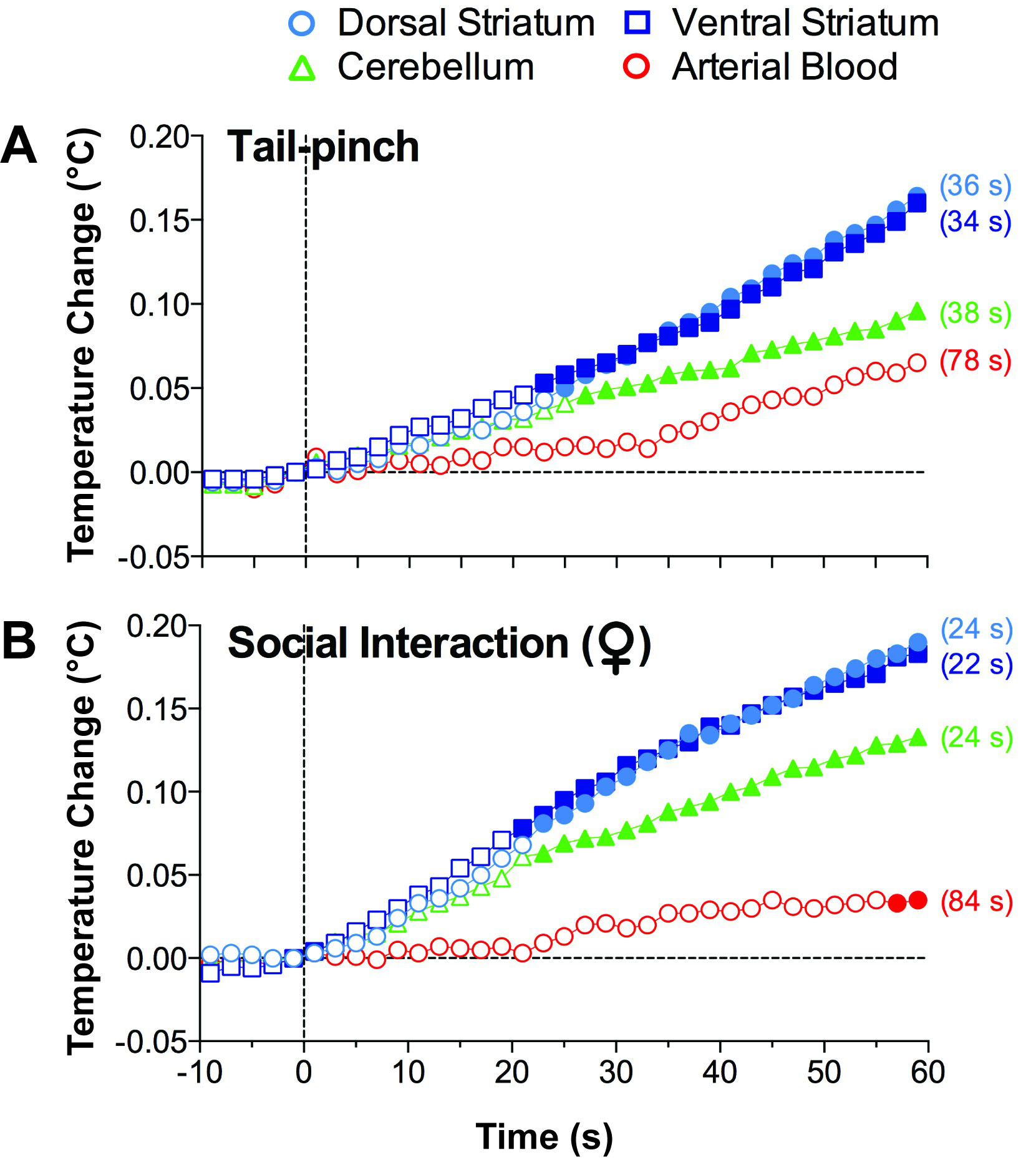 Fig. 2.
Fig. 2.Rapid temperature changes in several brain structures and arterial blood induced by tail-pinch (A) and male-female social interaction (B) in freely moving rats. Filled symbols show values significantly different from the pre-event baseline. Values in brackets show latencies to increase determined as the first value significantly different from the pre-event baseline. Original data were presented in [20].
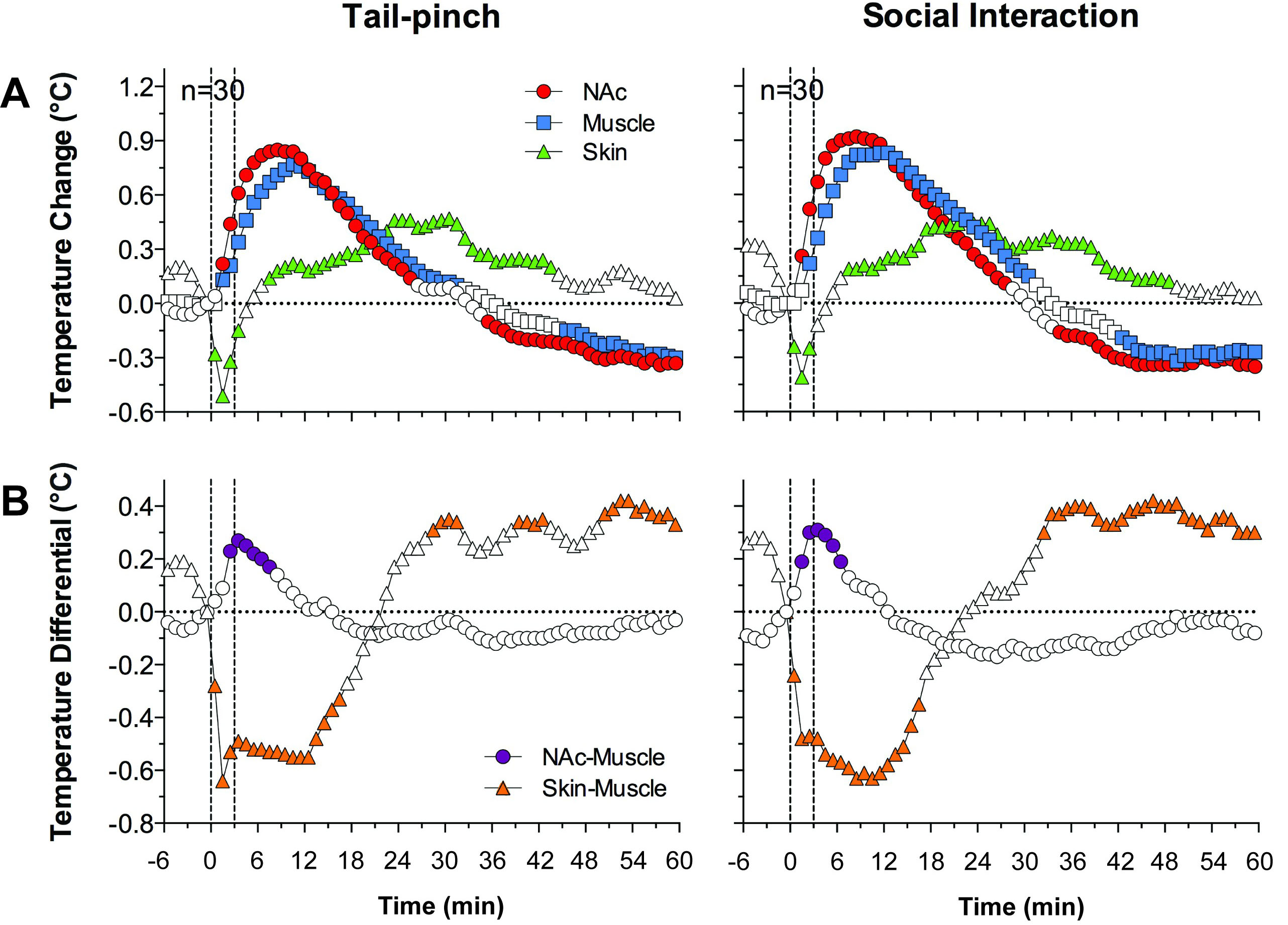 Fig. 3.
Fig. 3.Changes in temperature in the NAc, muscle, and subcutaneous space induced by tail-pinch and male-male social interaction in freely moving rats. Top graphs (A) show relative temperature changes, and bottom graphs (B) show NAc-muscle and Skin-muscle temperature differentials. Filled symbols show values significantly different from the pre-event baseline. Original data were published in [21].
Hence, physiological brain hyperthermia results from the joint contribution of two factors: intra-brain heat production due to metabolic neural activation and inhibition of heat loss due to peripheral vasoconstriction. Peripheral vasoconstriction is a more dynamic and prolonged effect that may determine the long duration of brain hyperthermia and its similar pattern occurring in different brain structures.
Along with its known role in temperature regulation, peripheral vasoconstriction was considered as a component of Walter Cannon’s fight-or-flight response and the initial manifestation of Hans Selye’s general adaptation syndrome (stress). It is also viewed as a measure of emotional excitation of both negative and positive valence, a component of sympathetic activation, and the peripheral manifestation of general arousal as a non-specific adaptive response elicited by novel and potentially dangerous stimuli. Despite attribution of this peripheral effect to different physiological and psychological constructs, this is an effect that is triggered by neural activation to redistribute blood supply from organs that are less important for survival (skin, digestive tract, kidney, etc.) to organs critical for survival (brain). With increased cerebral blood flow (CBF) to the brain, such blood redistribution appears to play an important adaptive role in providing more oxygen and glucose to prevent their potential decreases from enhanced metabolic consumption.
Both oxygen and glucose arrive at the brain from arterial blood. Therefore, the primary focus of numerous studies was on different aspects of brain blood supply (tone of cerebral vessels and CBF) associated with functional neural activation. Although these studies revealed a basic association of increased CBF with neural activation [22, 23, 24, 25], most techniques used to assess CBF are indirect, have a low temporal resolution, and cannot be used in freely moving animals, especially rats. The development of electrochemical techniques that provide high substrate sensitivity, selectivity, and second-scale temporal resolution made it possible to examine real-time fluctuations of oxygen and glucose in selective brain areas and different body locations in awake, freely moving rats [26]. While assessments of CBF and the tone of cerebral vessels are important for understanding how oxygen and glucose arrive in brain capillaries, it is critically important to know the real-time dynamics of these substances in the brain’s extracellular space and their relationships with oxygen changes in peripheral tissues.
Our electrochemical measurements revealed that brain oxygen and glucose levels significantly increased following arousing stimulation and showed a significant correlation within the entire duration of responses and their initial components [26] (Fig. 4, Ref. [6]). For both substances, the strongest concentration rise occurred during the first minutes of stimulation. Although glucose and oxygen enter brain tissue from arterial blood based on gradient-dependent mechanisms, the rapidity of these changes argues against this mechanism and points at neuronal activation with subsequent cerebral vasodilation (neurovascular coupling) as the initial trigger for increases in the brain’s extracellular space. While direct neural effects on brain capillaries for a number of vasoactive substances released by neurons and glia (i.e., NO, CO2, adenosine, etc.) are usually viewed as the primary mechanism underlying cerebral vasodilation and increased CBF [24, 25, 27], these effects could also occur as a result of redistribution of arterial blood from the skin and most internal organs to the brain due to peripheral vasoconstriction.
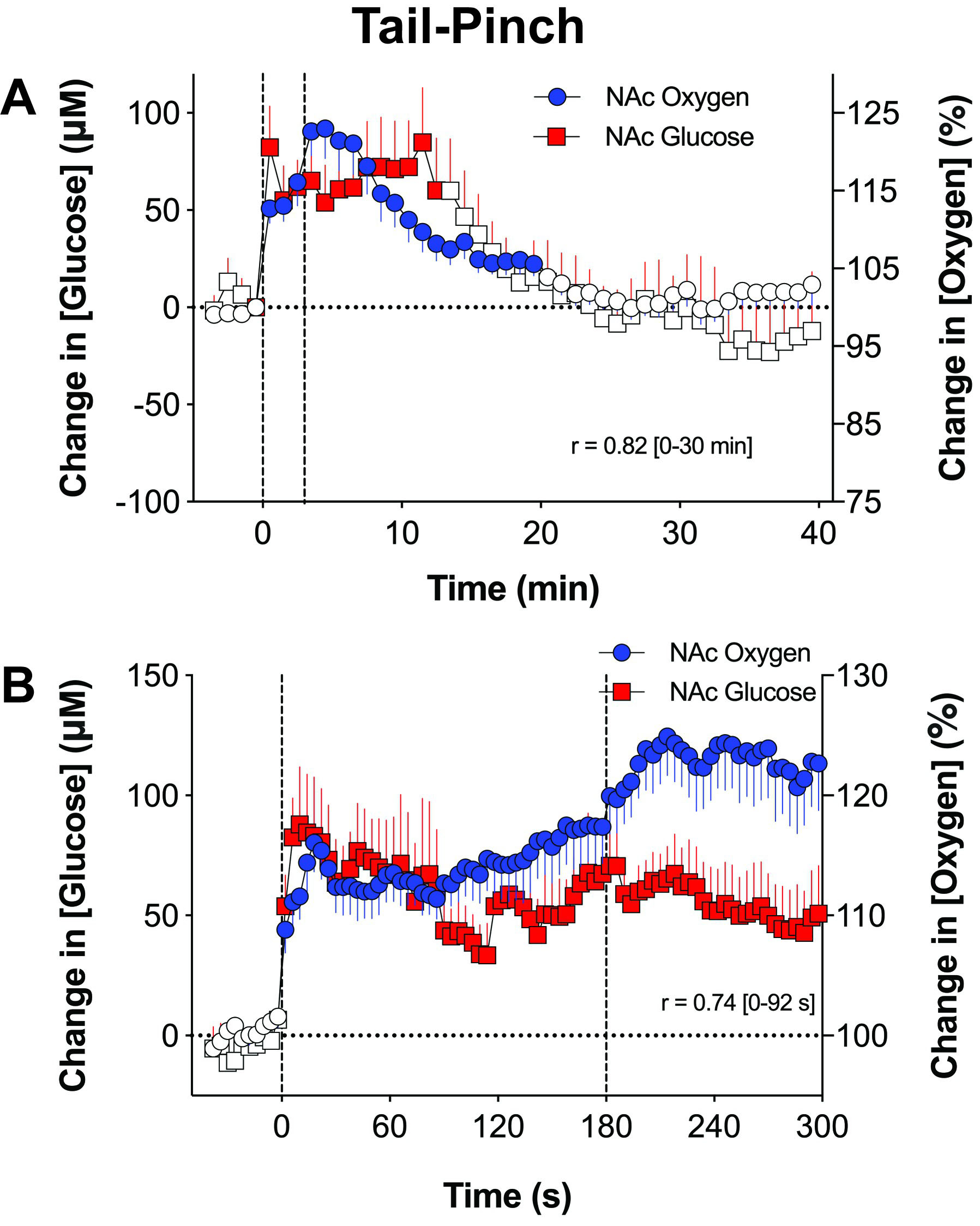 Fig. 4.
Fig. 4.Mean changes in NAc oxygen and glucose induced by tail-pinch in awake, freely moving rats. Data were assessed with low (A) and rapid (B) time-course resolution. Filled symbols show values significantly different from the pre-event baseline. Changes in oxygen and glucose assessed at slow and rapid time resolution significantly correlate (r = 0.82 and 0.74). Original data were presented in [6].
To provide more direct proof for the mechanisms responsible for physiological increases in brain oxygen and glucose, we recently developed a two-sensor recoding technology where one oxygen sensor was implanted in the brain structure (NAc) and another in the SC space [28]. Using this approach, we found that oxygen changes in the NAc and SC space induced by both tail-pinch and social interaction occurred in opposite directions, rapidly increasing in the brain and decreasing in the periphery (Fig. 5, Ref. [28]). Moreover, these changes are very tightly correlated (r = –0.76 and –0.92 for tail-pinch and social interaction, respectively). High-speed resolution analysis revealed that the change in oxygen levels first appears in the brain and, after a certain latency, in the SC space. Therefore, the entire brain oxygen response combines the initial direct neural effect and subsequent cerebral vasodilation, resulting in strong and prolonged skin vasoconstriction. Due to this latter effect, the brain oxygen response is relatively long, likely exceeding a much shorter neuronal activation, which appears to be responsible for the initial oxygen change.
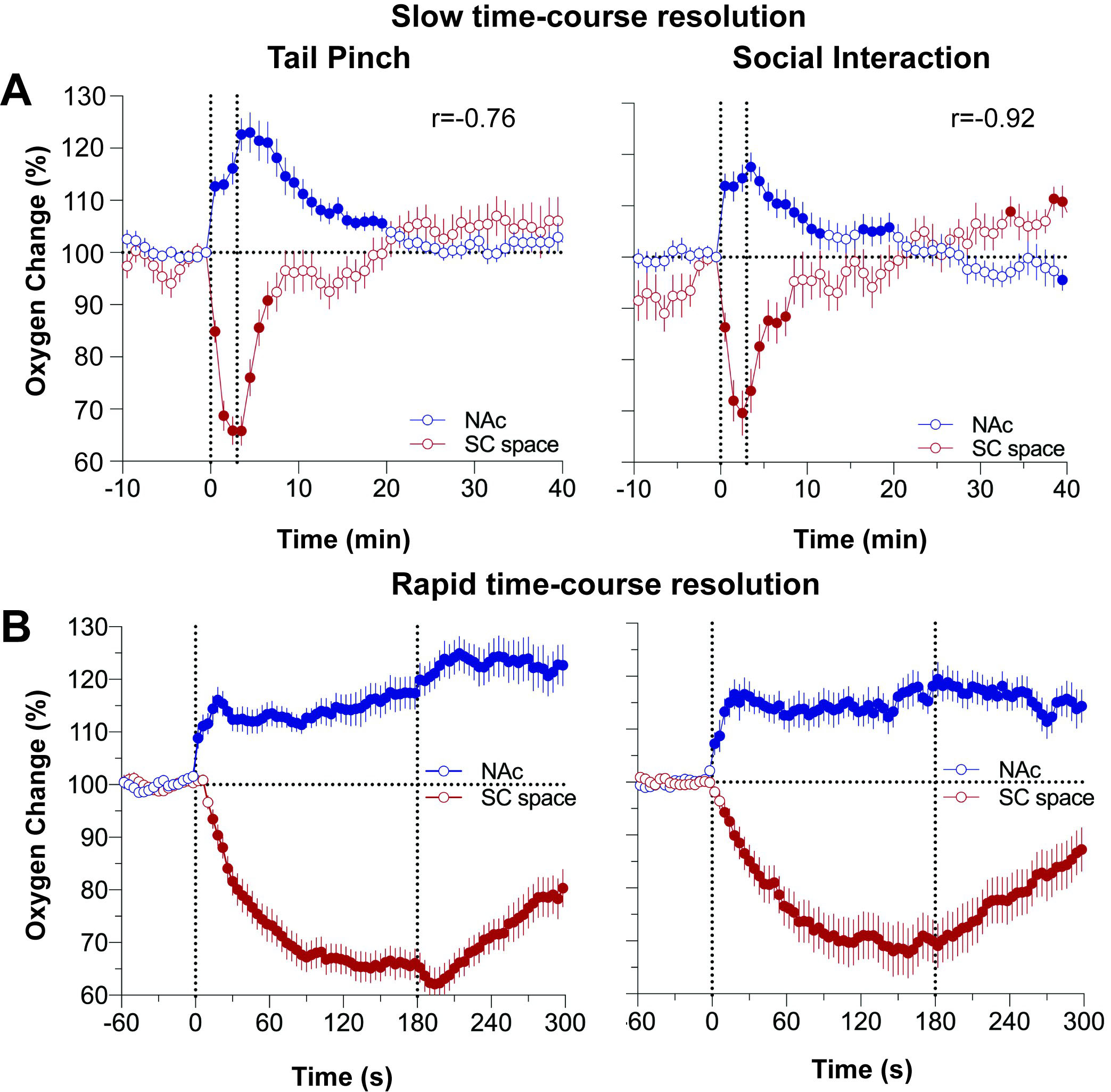 Fig. 5.
Fig. 5.Changes in oxygen levels in the NAc and subcutaneous space induced by tail-pinch and social interaction in awake, freely-moving rats. Top graphs (A) show changes assessed at slow, 1-min time resolution, and bottom graphs (B) show the results of rapid-resolution analyses. Changes in oxygen in the NAc and subcutaneous space elicited by both arousing stimuli show a significant opposite correlation (r = –0.76 and 0.92 for tail-pinch and male-to-male social interaction. Original data were published in [28].
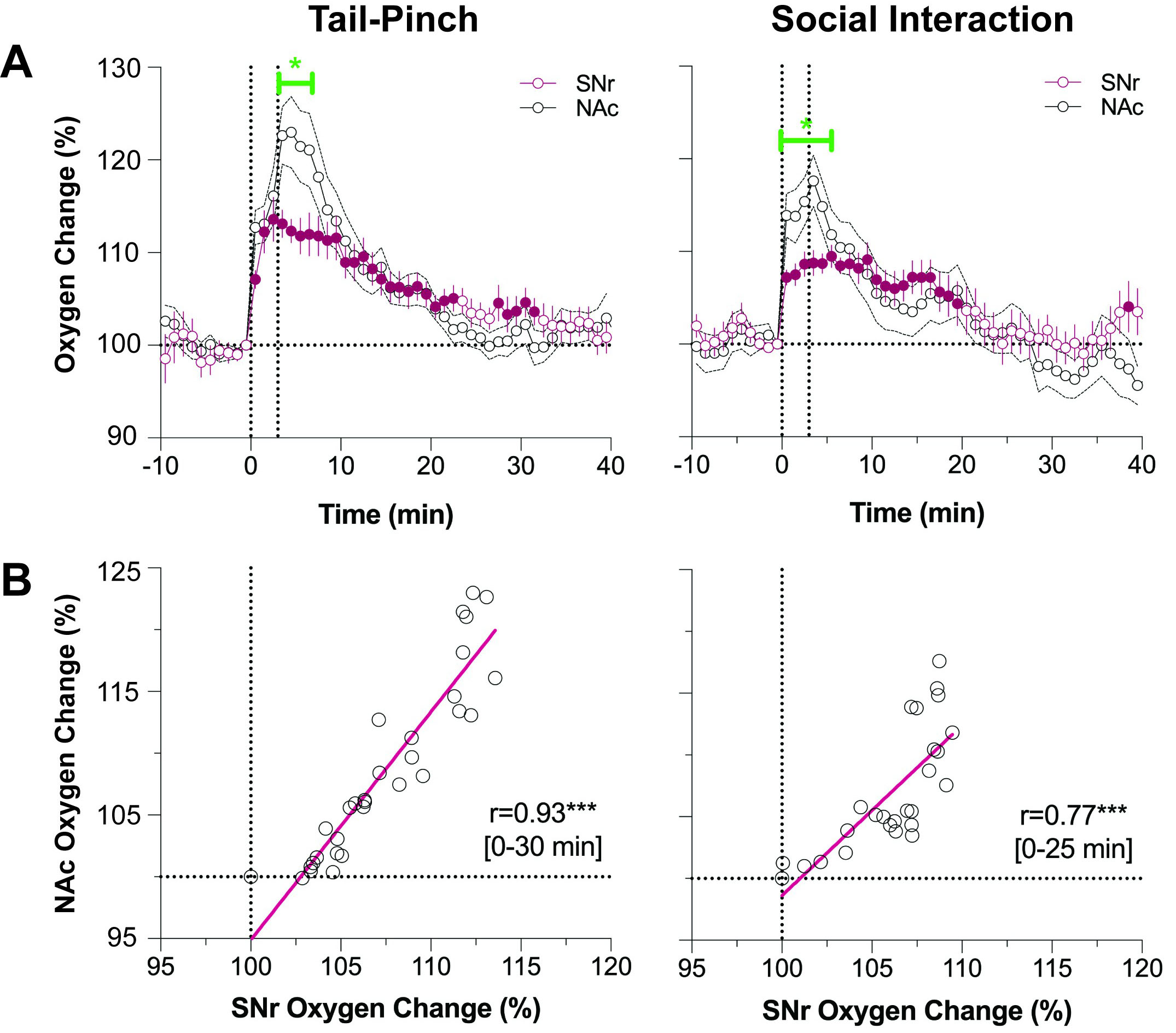
Powerful peripheral vasoconstriction that results in enhanced blood inflow to the entire brain tissue (global CBF) can also explain why oxygen responses in different brain structures have a similar time course. Usually, the specific changes in local CBF or oxygen levels in individual brain structures are viewed as reflections of specific changes in neuronal activity, which should be different in different brain structures. To test whether brain oxygen responses are structure-specific or have a global pattern, we conducted a similar experiment where the brain electrode was implanted in the substantia nigra, pars reticulata (SNr). Both basal and stimulus-induced changes in neuronal activity in this structure profoundly differ from those in the NAc. While most accumbal neurons are silent or have slow sporadic activity showing phasic excitations associated with sensory stimulation and initiation of movements [29, 30, 31, 32], SNr neurons are autoactive and maintain a relatively high rate of impulse activity showing transient inhibitions or activations elicited by sensory stimuli [33, 34, 35, 36]. Therefore, by comparing oxygen responses in both structures, we can assess the role of neuronal activity in mediating brain hyperoxia-induced by arousing stimuli.
This comparison revealed that both tail-pinch and social interaction induced rapid oxygen increases, similar in their time-course in the NAc and SNr [28] (Fig. 6, Ref. [37]). These changes for both stimuli are also tightly correlated. However, changes in the NAc were larger in magnitude, but these differences resulted from a stronger oxygen increase at the first or second minute of stimulation. Both concentration curves descended similarly to the baseline. This higher initial oxygen jump seen in the NAc is consistent with the high excitability of accumbal neurons and phasic increases in their impulse activity following sensory stimulation.
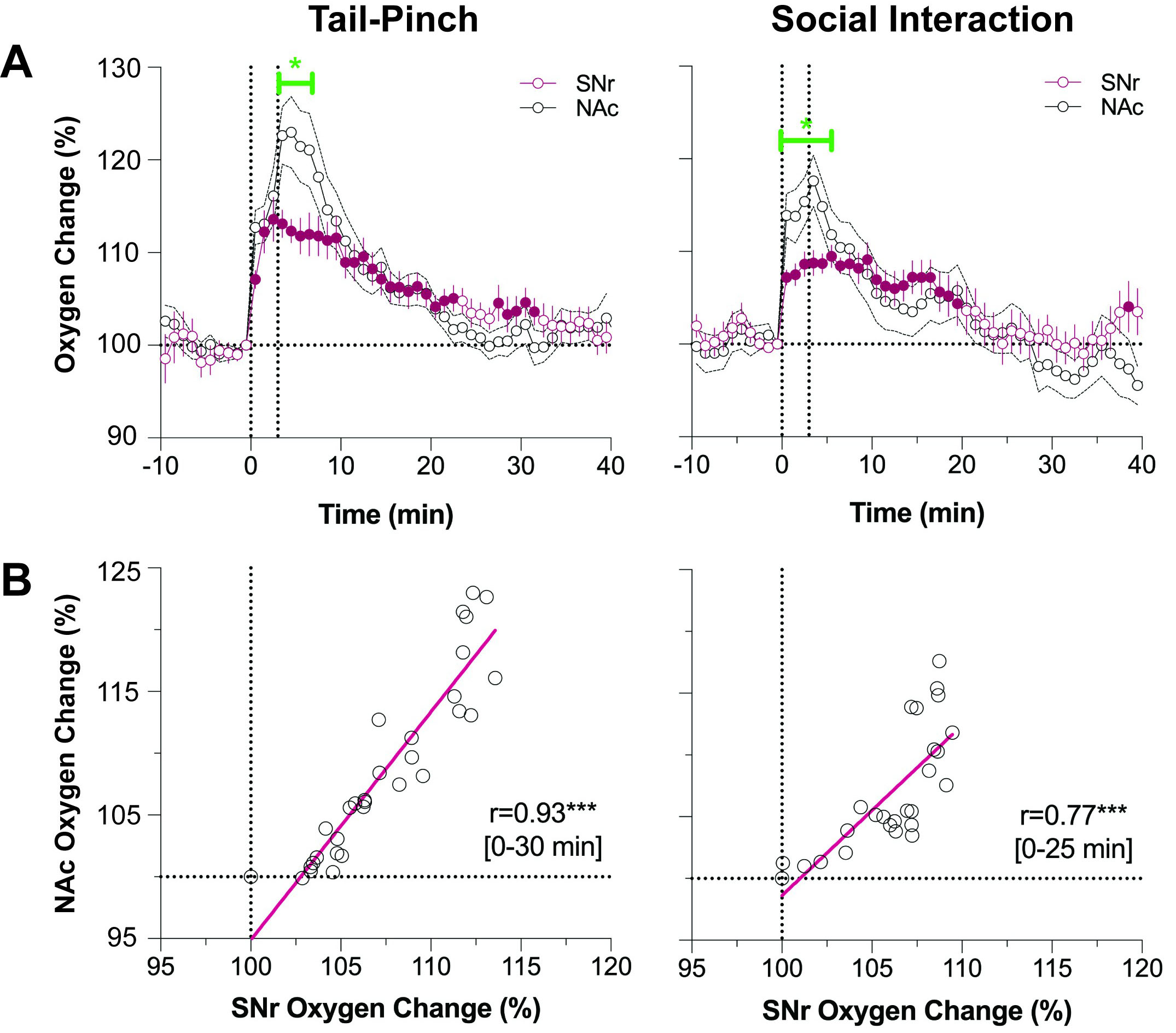 Fig. 6.
Fig. 6.Differences in oxygen responses in the NAc and SNr induced by tail-pinch and social interaction in awake, freely moving rats. Top graphs (A) show changes in relative changes in oxygen response, and bottom graphs (B) show their correlative relationships. Original data were published in [37].
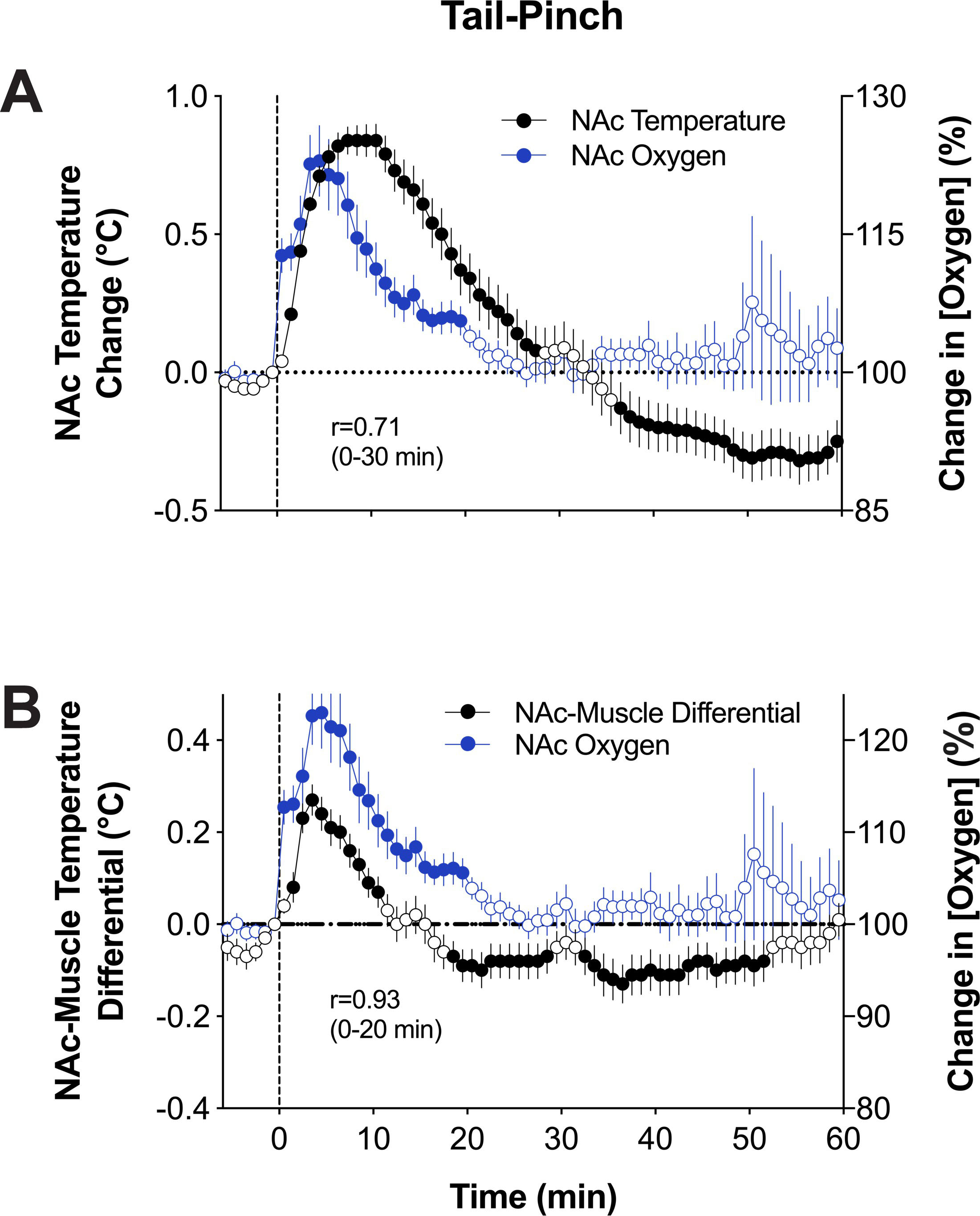
The proper inflow of oxygen from arterial blood is essential for brain metabolic
activity, but the relationships between its rapid change in brain tissue and
cerebral metabolism remain unclear. Since multi-site thermorecording provides an
approach to assess changes in metabolic activity based on heat production, we
examined how these temperature parameters are related to changes in NAc oxygen
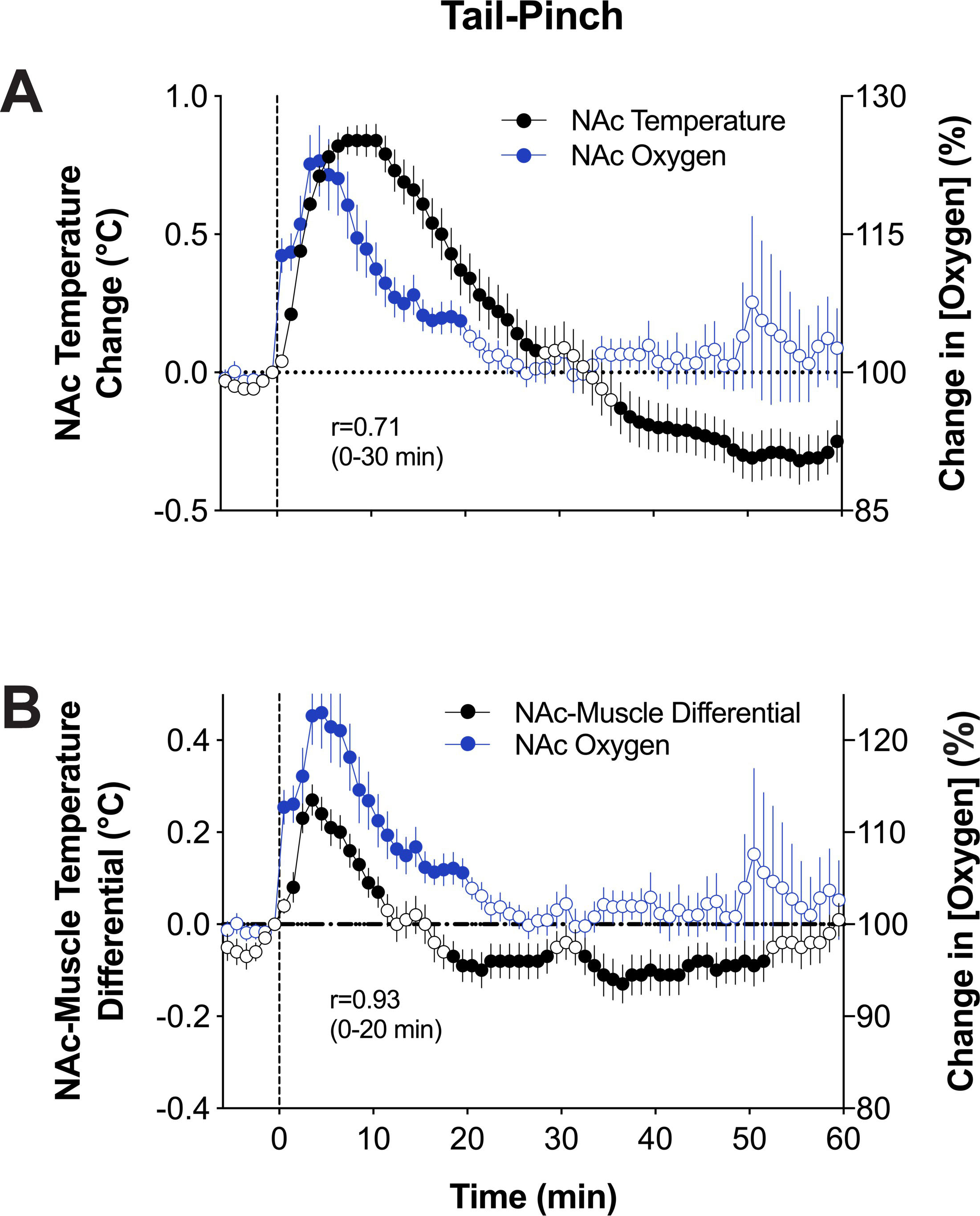
[37]. This study revealed that NAc oxygen increases induced by tail-pinch showed
a similar pattern to brain temperature, which increased more slowly than oxygen
(Fig. 7, Ref. [28, 37]). Moreover, these two parameters were tightly correlated (r = 0.71,
p
Norepinephrine (NE) can be viewed as a neurotransmitter, neuromodulator and hormone. In the CNS, it is released by axon terminals of NE-containing neurons, affecting the activity of multiple neurons and modulating their responsiveness to other neurochemical inputs [38, 39]. NE release in target areas of NE-containing neurons is an essential part of functional neural activation elicited by various arousing stimuli and during various types of motivated behaviors [40, 41, 42]. In the periphery, NE is also released by the terminals of sympathetic adrenergic nerves that innervate most of the body’s organs [43, 44, 45]. In this case, NE acts primarily on adrenoceptors expressed on vascular smooth muscles, thus affecting their tone. NE is also released by the adrenal medulla cells directly in the bloodstream, thus affecting at lower concentrations on distantly located adrenoceptors to provide more tonic effects on the tone of blood vessels. Peripheral NE release is also triggered by neuronal activation and is a critical component of sympathetic activation.
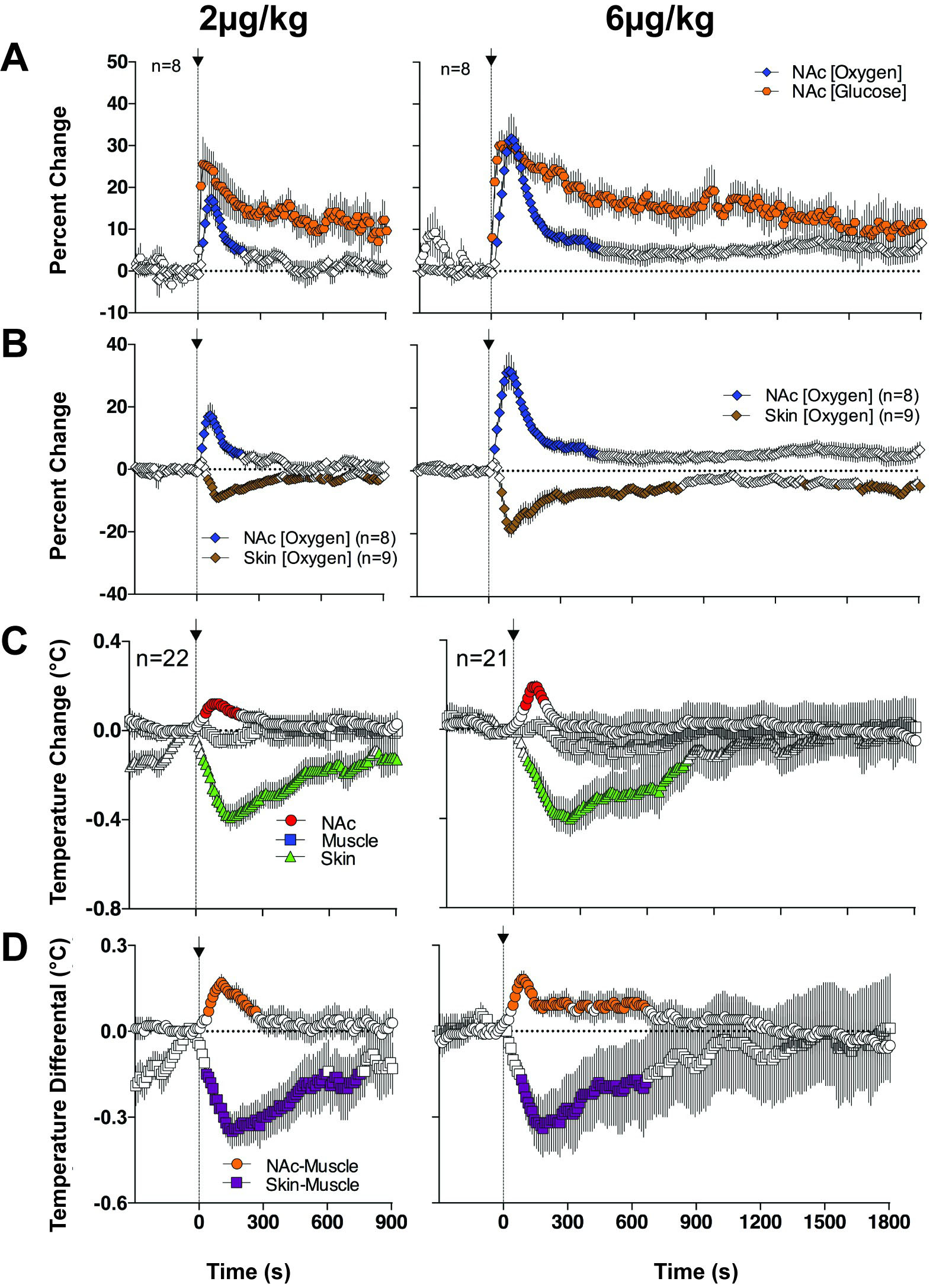
By interacting with
To determine whether NE increases CFB and entry of oxygen and glucose in brain
tissue, we examined changes in NAc levels following intravenous (iv) delivery of
NE at low, physiologically relevant doses (2 and 6
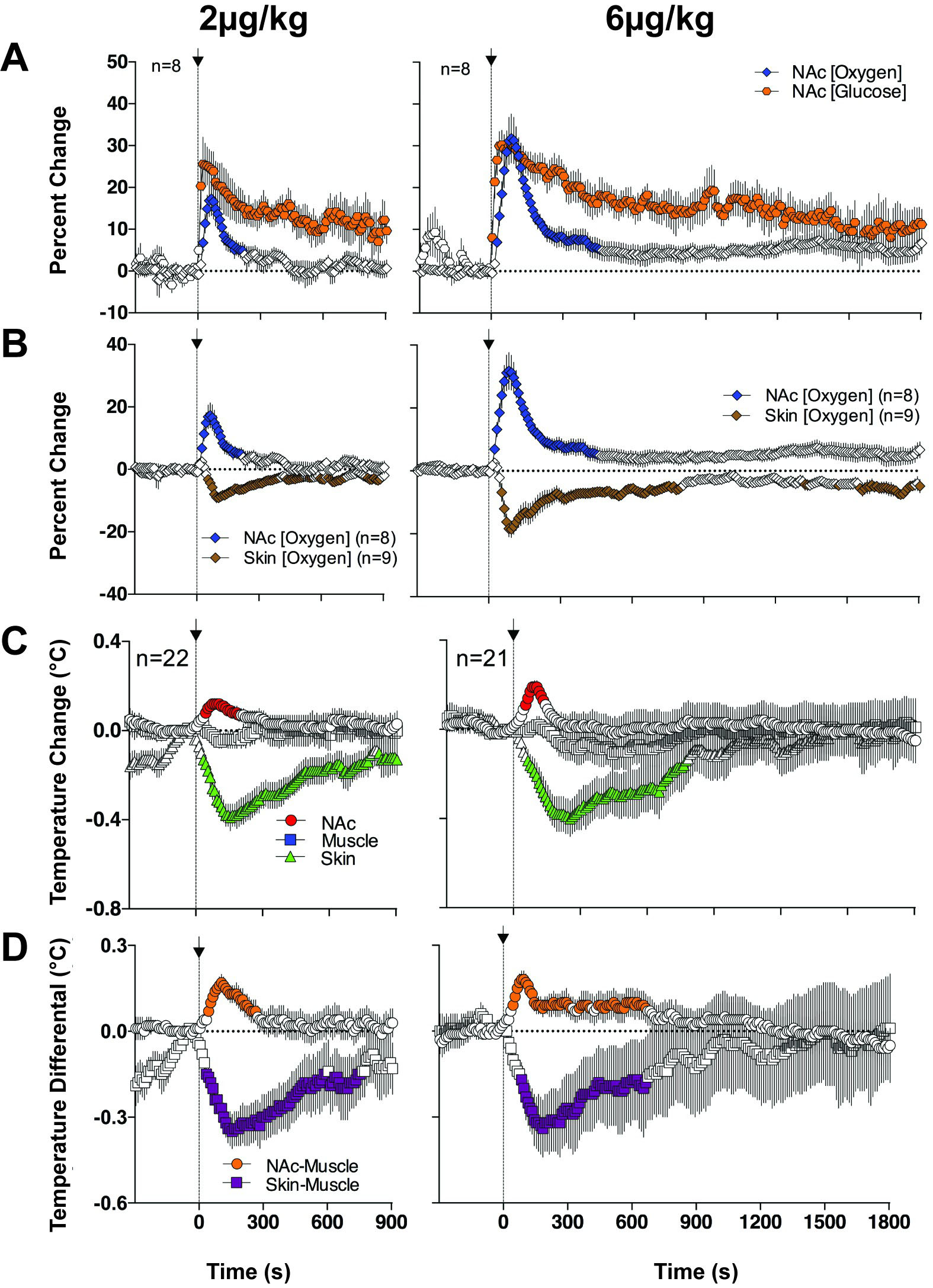 Fig. 8.
Fig. 8.Changes in NAc oxygen and glucose and temperature parameters induced by iv administration of NE at two low doses in awake, freely moving rats. (A) shows changes in NAc oxygen and glucose. (B) shows relationships between oxygen changes in the NAc and SC space. (C) shows temperature changes in NAc, temporal muscle and SC space. (D) shows changes in NAc-muscle and Skin-Muscle temperature differentials. Filled symbols show values significantly different from baseline. Original data were published in [50].
Unexpectedly, we found that iv NE increases brain temperature and brain-muscle differential suggesting metabolic neural activation. Since heat production is an ultimate consequence of neural activation [53, 54, 55], this finding may suggest that BBB-impermeable NE by some actions in the periphery creates an ascending sensory signal transmitted to the CNS via visceral sensory nerves. As shown in Fig. 8, NE-induced increases in glucose and oxygen were more rapid than increases in brain temperature and brain-muscle differentials. This is not surprising because temperature parameters, by their physical nature, show slower dynamics and follow neuronal activation as a rapid electrophysiological event [54].
While the exact mechanism of how peripherally acting NE induces metabolic neural activation remains unknown, it is possible to speculate that this effect is triggered by NE-induced changes in vascular tone and cardiac activity. Peripheral blood vessels are densely innervated by afferents of sensory nerves, which are sensitive to mechanical, pressure and temperature stimuli and chemical properties of blood [56, 57]. Powerful vasoconstriction and a sharp rise of arterial blood pressure induced by iv NE can activate these receptors [58, 59], thus creating ascending sensory signals from the periphery. This signal is transmitted to the CNS by sensory nerves and triggers a chain of neurovascular events, finally resulting in enhanced entry of oxygen and glucose in brain tissue. While this mechanism remains hypothetical, iv NE induces cortical EEG desynchronization [60], an index of neuronal activation. This central effect is secondary to a rapid rise in arterial blood pressure induced by this drug [61, 62].
Increases in NAc oxygen and glucose induced by iv NE may also result from a rapid increase in arterial blood pressure with subsequent increases in perfusion pressure in cerebral vessels. While this mechanism cannot be excluded, its significant contribution seems unlikely because of the tight regulation of arterial blood pressure due to the baroreflex mechanism. Because of this mechanism, physiological fluctuations in arterial blood pressure are highly phasic, and its increases are followed by rebound-like decreases [63].
While it is well known that neural activation is the initial trigger for multiple physiological and behavioral effects elicited by arousing stimuli, in this review, we present evidence that peripheral vasoconstriction, a universal, centrally triggered physiological effect, is a significant contributor to changes in neural parameters by redistributing arterial blood inflow from the body surfaces and most peripheral organs to the brain as the most critical organ for organism’s survival. This mechanism appears to be critical for proper delivery to the brain of oxygen and glucose, which are essential for brain metabolic activity. While the direct action of brain neurons, glial cells, and products of their metabolic activity are considered major factors in mediating cerebral vasodilation and increased CBF, these effects also occur due to the redistribution of arterial blood from the periphery to the brain. Due to the robust vascularization of brain tissue, these changes appear non-specific and occur globally in the brain. Through this mechanism, the brain can, within certain limits, prevent possible deficits of oxygen and glucose consumed during metabolic activation.
BBB, blood-brain barrier; CBF, cerebral blood flow; CNS, central nervous system; iv, intravenous; NAc, nucleus accumbens; NE, norepinephrine; SC, subcutaneous; SNr, substantia nigra, pars reticulata.
Not applicable.
The author wants to thank Shruthi A. Thomas for editorial work with this manuscript and valuable suggestions on its improvement. I thank Carlos M. Curay for help in creating high-quality figures.
This research was funded by the Intramural Research Program of the NIH, NIDA, grant number 1ZIADA000566-10.
The author declares no conflict of interest.