
 , Ying Gao 1,*, Qiongqiong Wei 1
, Ying Gao 1,*, Qiongqiong Wei 11 Department of Gynecology and Obstetrics, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, 430022 Wuhan, Hubei, China
†These authors contributed equally.
Abstract
Background: Endometrial infertility accounts for a significant proportion of infertility cases, and single-cell transcriptome data have revealed that hub genes may play an important role during pregnancy. Methods: Based on the endometrial single-cell sequencing data from National Center for Biotechnology Information (NCBI) database, we performed clustering, staging, and functional analyses to screen and validate key genes affecting endometrial infertility. Results: Through bioinformatics analysis, we found that the proportion of ciliated cells peaked from the early to mid secretory phase, ciliary motility decreased in the mid secretory phase, while the hub gene that connexin 43 (GJA1) and secreted phosphoprotein 1 (SPP1) expressed in the endometrium may determine successful pregnancy. In immunohistochemistry validation, GJA1 and SPP1 were significantly highly expressed in the endometrium of a normal pregnancy, compared to recurrent miscarriage. Similarly, GJA1 and SPP1 were expressed higher in the fetal villus of a normal pregnancy as compared to recurrent miscarriage, while no difference was found in the decidua. CellPhoneDB and protein–protein interactions (PPIs) indicated an interaction among notch receptor 1 (NOTCH1), GJA1 and SPP1. Conclusions: GJA1 and SPP1 exhibit higher expression levels in the endometrium and fetal villus of a normal pregnancy as compared to recurrent miscarriage, suggesting that GJA1 and SPP1 may play a pivotal role in endometrial infertility.
Keywords
- single-cell transcriptome
- GJA1
- SPP1
- ciliated cell
- infertility
In recent years, the incidence of infertility has been increasing [1], affecting
8 to 12 percent of couples of childbearing [2]. Hyperprolactinemia, ciliary
dysfunction, cystic fibrosis, infection, ovarian insufficiency, polycystic ovary
syndrome, endometriosis, uterine fibroids, and endometrial polyps may be factors
leading to infertility in women. Among the factors affecting pregnancy,
endometrial infertility plays a pivotal role.
Endometrial infertility refers to the change of endometrial receptivity due to
various reasons, which leads to the failure of embryo implantation. The
receptivity of the endometrium refers to its ability to accept embryo
implantation, which is time-sensitive during the implantation window, usually on
days 22–24 of the menstrual cycle. Embryo implantation involves three processes:
“position”, “adhesion”, and “invasion” [3, 4]. The molecular markers
commonly used to assess endometrial receptivity for a successful pregnancy
include integrin
GJA1 (connexin 43) is a component of gap junctions that allows cell-to-cell communication and regulates proliferation [8], differentiation [9], migration [10], and cell death [11]. Research has shown that GJA1 overexpression contributes to trophoblast differentiation and increases human chorionic gonadotropin (HCG) levels, while low GJA1 expression inhibits trophoblast fusion [12] and disrupts uterine preparation for mouse embryo implantation [13]. Conditional deletion of GJA1 in uterine stromal cells enables disruption of gap junctions and leads to marked impairment of neovascular development within the stromal compartment, resulting in embryonic growth arrest and early pregnancy loss [14, 15]. GJA1 is a marker of oocyte maturation [16, 17, 18] and good embryo quality [19, 20], as well as an important factor in good delivery outcomes [21, 22, 23]. Secreted phosphoprotein 1 (SPP1) is an extracellular matrix glycoprotein that is highly expressed at the maternal-fetal interface and is a critical mediator of embryo implantation [24, 25, 26, 27]. SPP1 is increased in the uterine environment [28] and promotes cell surface integrin binding on the endometrium and trophectoderm, as well as cell adhesion and migration [29, 30, 31, 32]. Therefore, the loss of GJA1 and SPP1 expression can cause the loss of embryo implantation, and may play an important role in endometrial infertility.
Ciliated cells account for a small proportion of endometrial cells, including glandular cilia and surface cilia. In previous studies, although ciliated cells in the fallopian tube were related to egg transport, some ciliated cells were related to cancer development [33], and intrauterine ciliated cells were related to self-cleaning ability [34]. However, a paucity of research exists on their functions. In recent years, with the development of bioinformatics technology, especially the rise of single-cell transcription analysis technology, researchers are able to focus on and explore the target genes in the identification of endometrial infertility.
Institutional Ethical Approval and Informed Consent: all the experimental protocols, including human tissue collection, were approved by the Ethics Committee of Tongji Medical College, Huazhong University of Science and Technology based on the World Medical Association’s Declaration of Helsinki, and the ethical approval number was 0241-01. All tissues were collected with the written consent of the participants. The data analysis process was carried out in Rstudio Version 4.2.1 (Posit, Boston, MA, USA) as detailed in sections 2.2–2.5.
We collected 8 proliferative endometrial samples from women with normal pregnancies, 6 proliferative endometrial samples from patients with recurrent miscarriages as defined elsewhere [35, 36], and 5 mid secretory-phase endometrial samples, each from women with normal pregnancies and patients with recurrent miscarriages. In addition, we collected decidua and villus samples (6–8 weeks (w)) from 9 women with normal pregnancies, and 7 patients with recurrent miscarriages, at the Department of Gynaecology and Obstetrics, Union Hospital, Huazhong University of Science and Technology, from 2021 to 2022. After signing the informed consent with the patient, the endometrium of different periods was collected, cleaned with normal saline three times and sterile gauze, stored in 1.5 mL EP tube and marked, then stored in the –80 °C refrigerator for follow-up experiments
GSE111976 single cell RNA-seq of human endometrium [37] was acquired from National Center for Biotechnology Information (NCBI)’s Gene Expression Omnibus (GEO) [38]. The data were filtered by the percentage of mitochondria. Nonlinear dimensionality reduction was performed using the t-Distribution Stochastic Neighbour Embedding (t-SNE), the FindAllMarkers function was used to identify significant clusters [39], and cell types were annotated based on specific biomarkers [40, 41], DCN, UCA1, FOXJ1, PTPRC, RNASE1, ACTA2, and LYZ was respectively one of biomarkers of Stromal fibroblast, Unciliated Epithelium, Ciliated cell, Lymphocyte, Endothelium, Smooth muscle cell and Macrophage.
We analyzed ciliated cells independently by the subset function and performed a statistical analysis of the proportions of ciliated cells, unciliated epithelium, and stromal fibroblasts according to the staging of the menstrual cycle, and the count function in Rstudio Version 4.2.1 was used to calculate the ratio of the number of cells to the total number of cells. In addition, we found that 2 of the 10 samples had large differences in cell numbers, and we removed the samples numbered 58 and 41. We performed a staging analysis according to the number of days in the menstrual cycle and counted the proportion of ciliated cells by using Rstudio and the function of count.
Differentially expressed genes (DEGs) were identified by EdgeR packages Version
3.38.4 (https://bioconductor.org/packages/release/bioc/html/edgeR.html), which
works on a table of integer read counts, the counts represent the total number of
reads aligning to each gene. EdgeR was concerned with differential expression
analysis and relative changes in expression levels between conditions rather than
with the quantification of expression levels and not directly with estimating
absolute expression levels. The differential expression threshold was set as a
p value
Circular cnetplots and gseaplot2 graphs of significant biological processes
(BPs) of the early and mid secretory stages were constructed by using Rstudio
Version 4.2.1. Then, PPI networks [47] of the early secretory phase and mid
secretory phase were structured using search tool for recurring instances of
neighbouring genes (STRING Version 11.0 (https://string-db.org/)) [48]. A
combined score
All tissues were fixed in 4% paraformaldehyde, deparaffinized after sectioning,
and incubated with 1% peroxidase to block endogenous peroxidase activity.
Endogenous peroxidase activity was blocked by soaking in 1% H
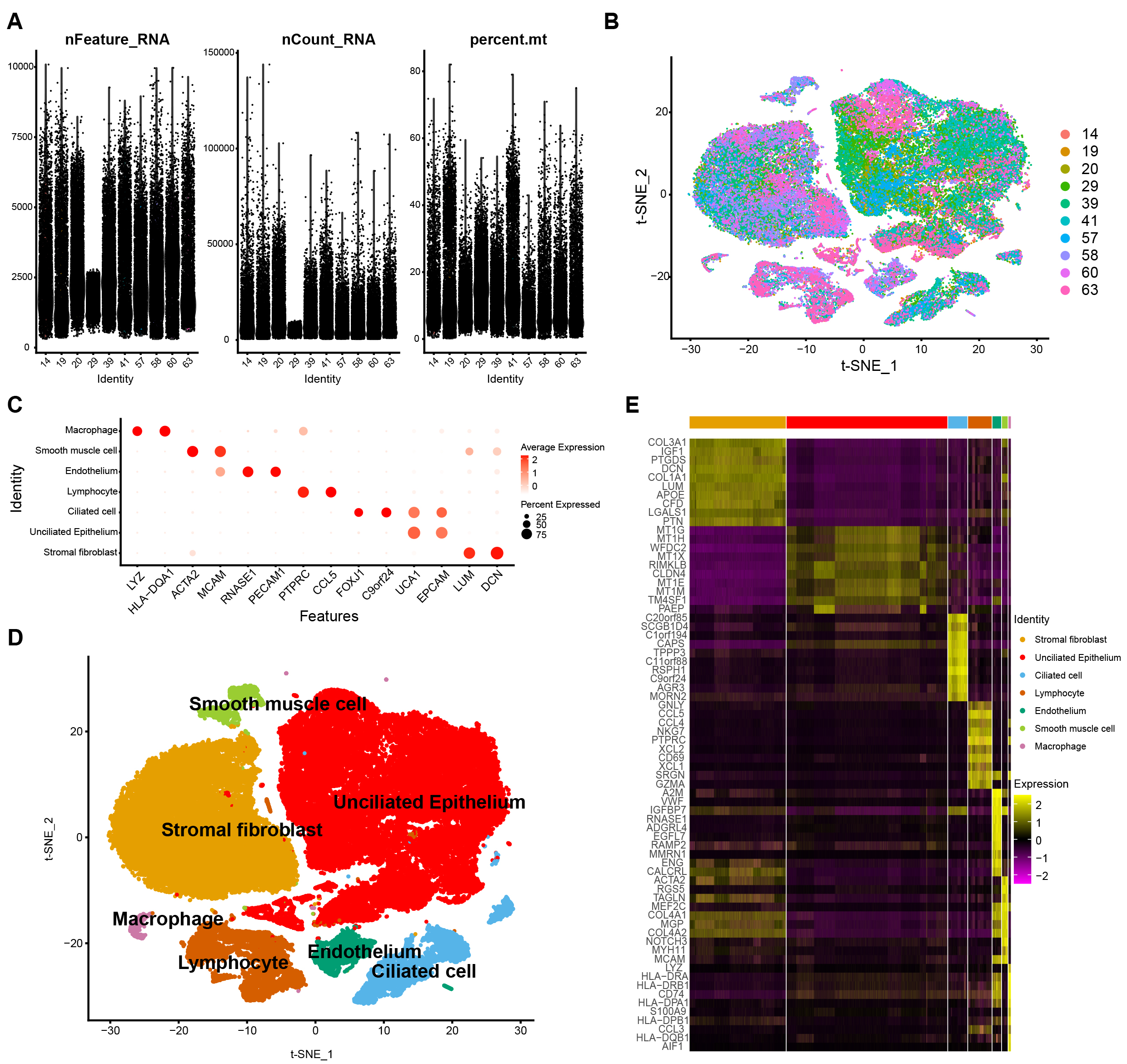
The data were filtered and described in descending order of their characteristics. A violin diagram demonstrates the features, count, and percent of RNA (Fig. 1A) of each sample. The t-SNE diagram demonstrates nonlinear dimensionality reduction clustering (Fig. 1B). Then, after annotating the cells with markers specific to each cell type (Fig. 1C), an annotated clustered t-SNE plot (Fig. 1D) was constructed. Lastly, an heatmap showing the expression signature of the top 10 marker genes for each 7 cell type was created (Fig. 1E).
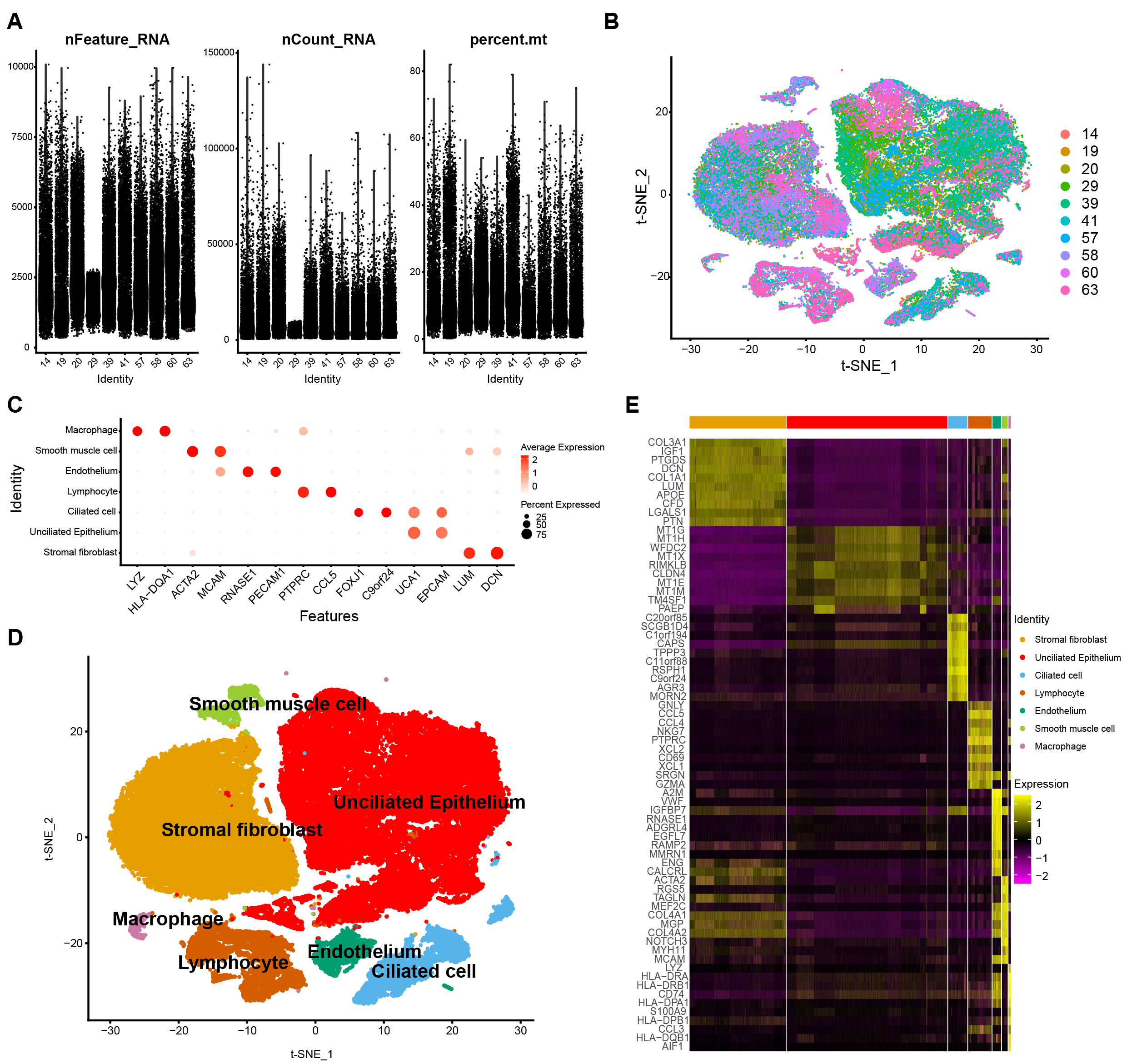 Fig. 1.
Fig. 1.Basic data processing. Violin Diagram (VlnPlot) (A). Integrated distribution t-distributed stochastic neighbor embedding (t-SNE) plot of 10 sample data in GSE111976 dataset, with each color-coded to indicate different samples (B). Dot plot showing average expression for marker genes in each cell types (C). Annotated clustered t-SNE plots for the different marker genes (D). Heatmap showing the expression signature of the top 10 marker genes for each 7 cell type (E).
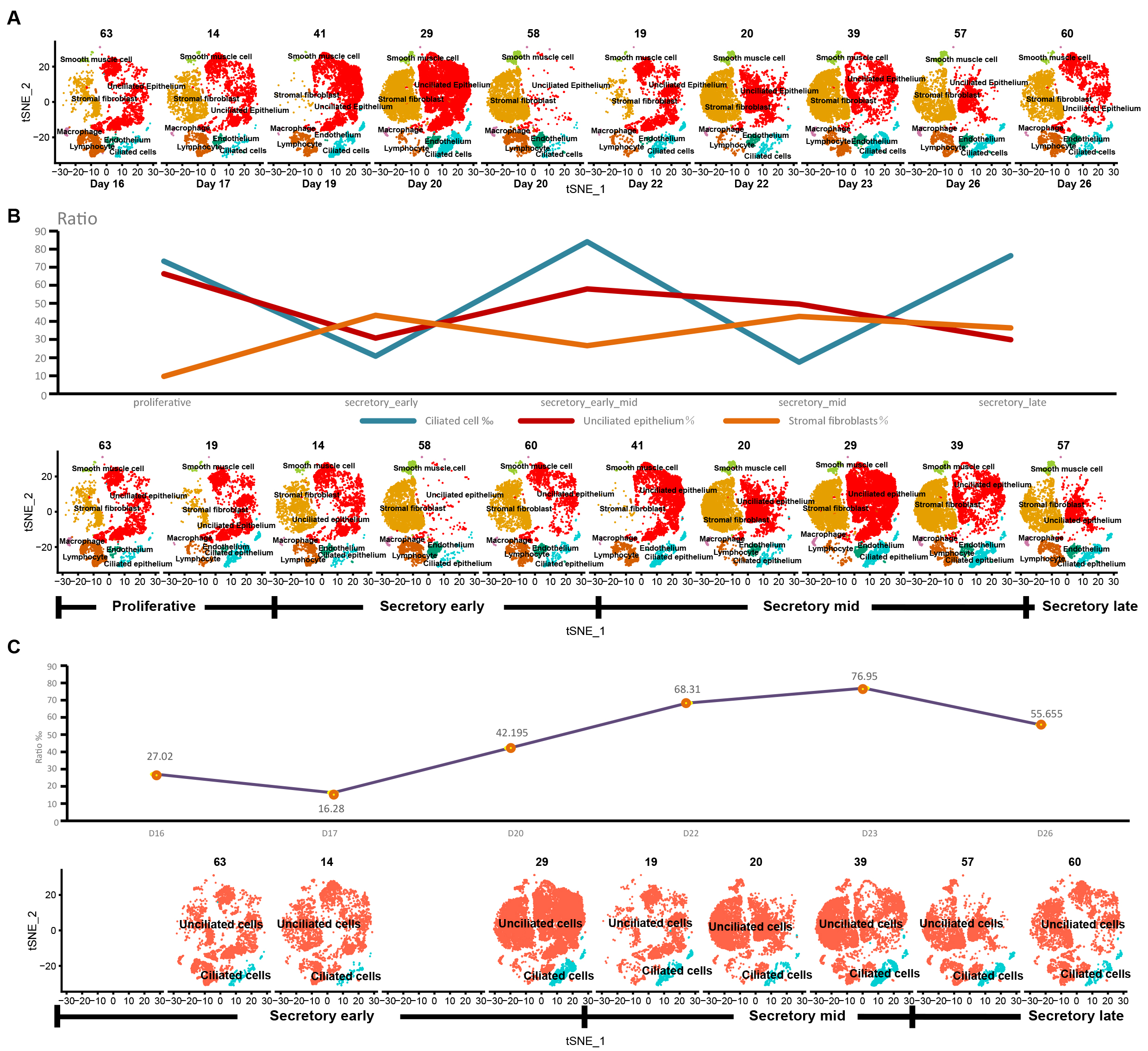
We annotated t-SNE clustering plots for 10 samples of single-cell sequencing data (Fig. 2A) and determined the proportions of ciliated cells, unciliated epithelium, and stromal fibroblasts according to the staging. The proportion of ciliated cells peaked at the early to mid secretory phases (Fig. 2B). After removing 2 samples (58 and 41) with large differences in cell counts and performing new staging of remaining samples according to the days of the menstrual cycle, ciliated cell ratios were plotted, and found to peak in the mid secretory phase, suggesting that ciliated cells may play a key role in the mid secretory phase (Fig. 2C).
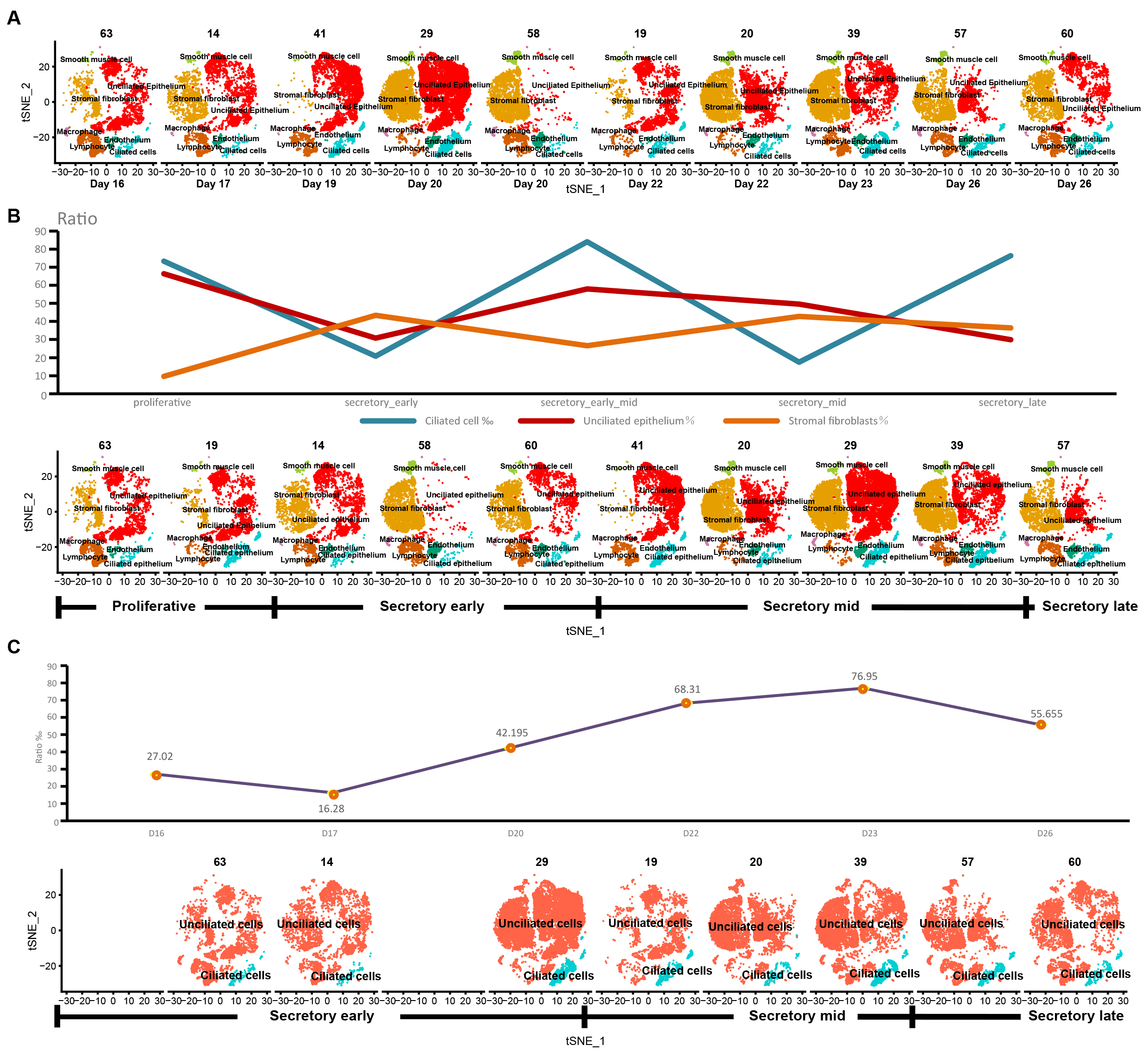 Fig. 2.
Fig. 2.The stage and ciliary cell proportion of the sample. t-SNE plots for 10 samples (A). After analyzing the cell type annotation, we got seven cell types per sample, and calculated the proportions of ciliated cells, unciliated epithelium, and stromal fibroblasts according to the original staging (B). The proportion of ciliated cells according to the new staging after removing samples 58 and 41 (C).
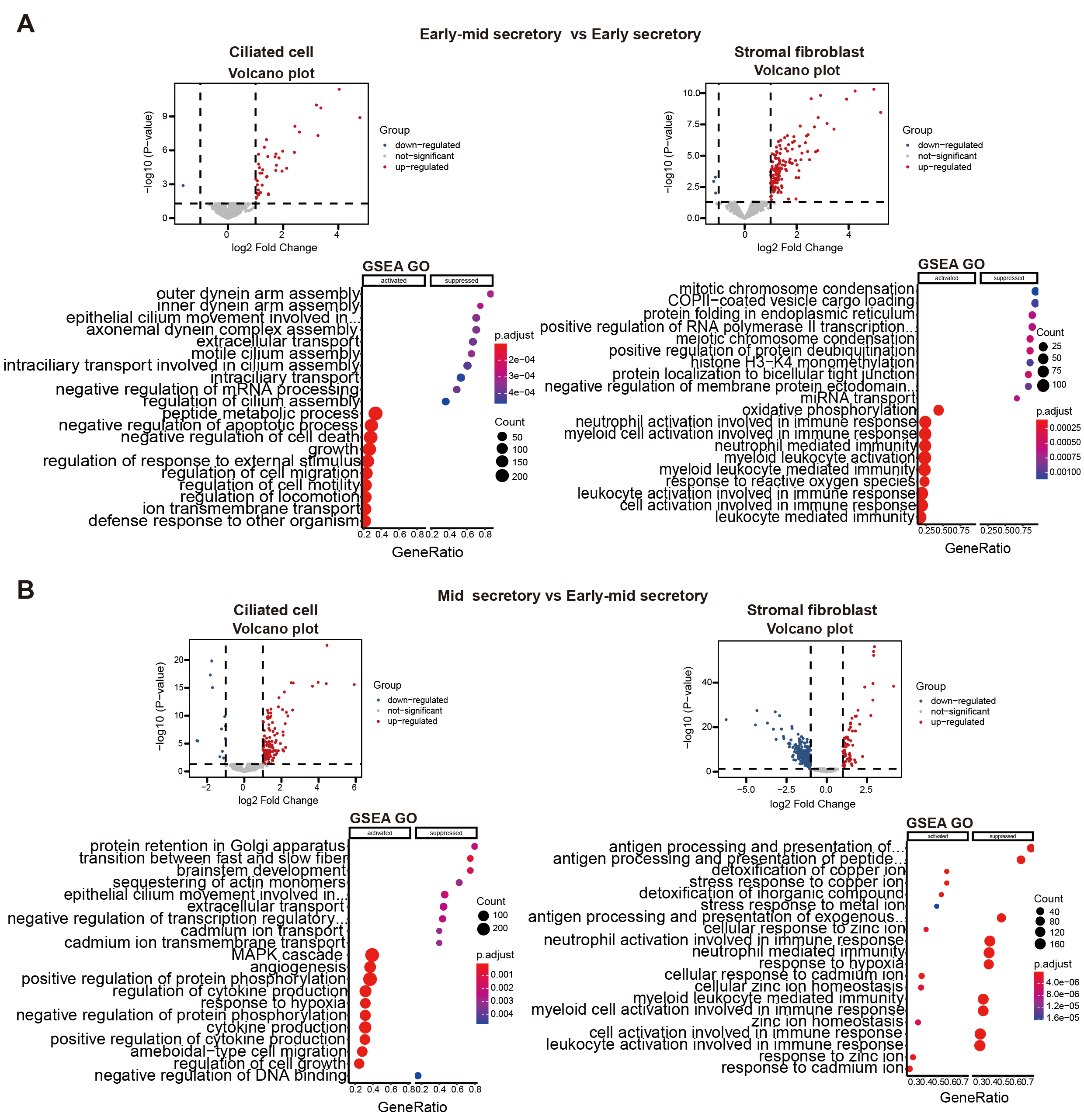
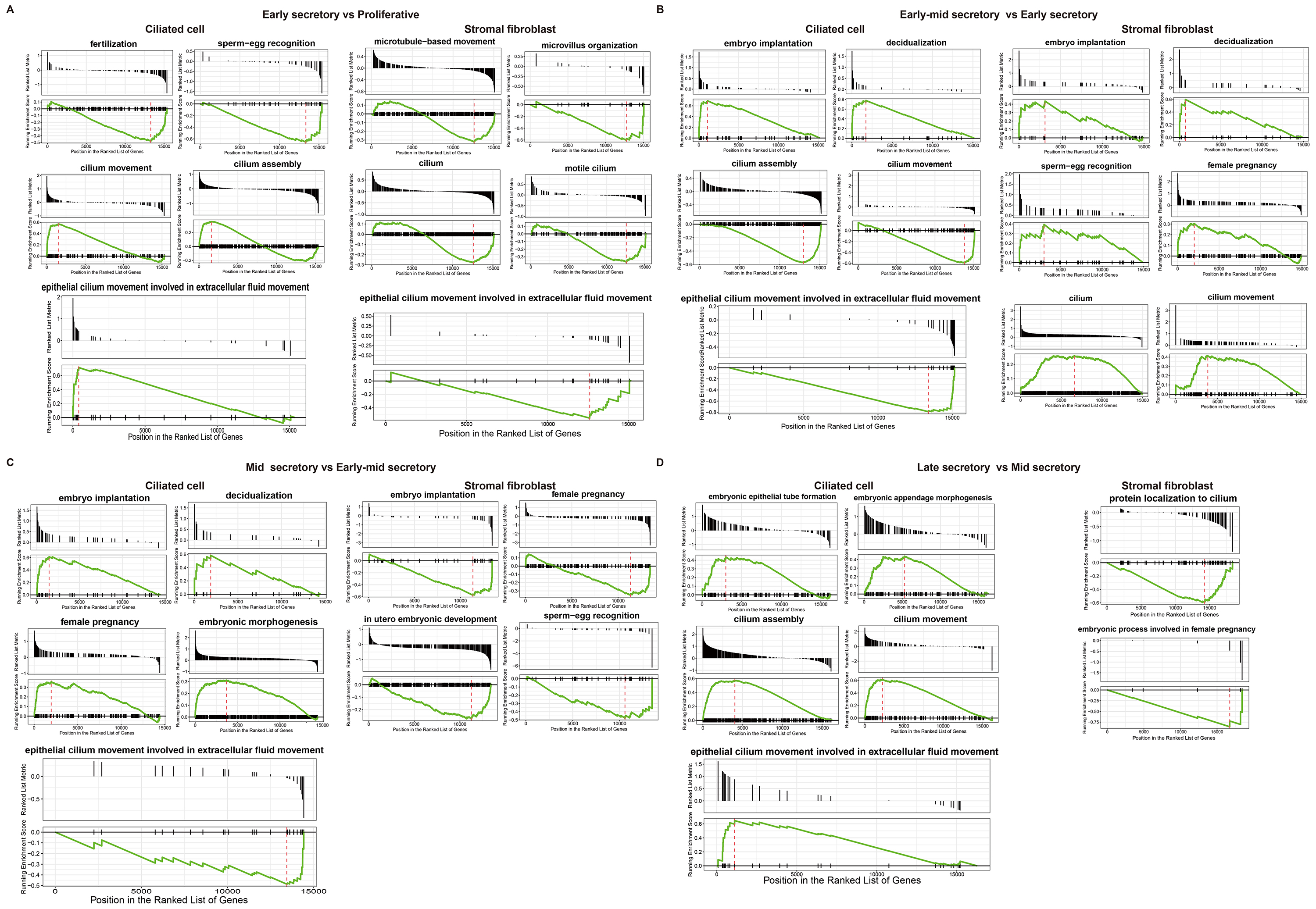
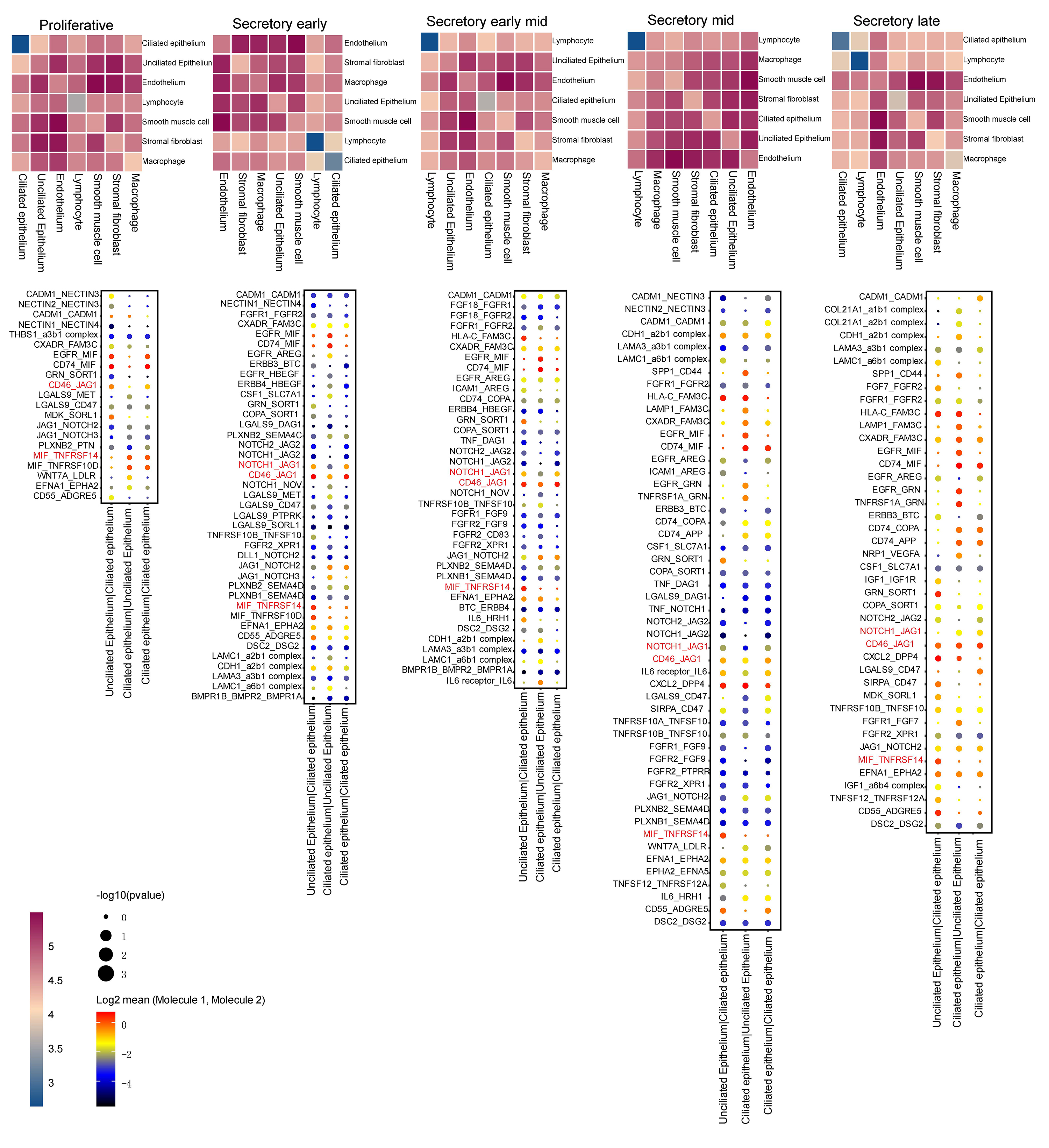
Volcano plots and GSEA GO bubble diagrams were generated by comparing ciliated cells to stromal fibroblasts mainly in the early mid secretory phase to the early secretory phase (Fig. 3A) and the mid secretory phase to the early-mid secretory phase (Fig. 3B). A gseaplot2 of meaningful BP was drawn. Compared to the proliferative phase, in the early secretory phase the fertilization, and sperm-egg recognition processes were downregulated, while ciliated cell assembly and epithelial cilium movement, which was involved in extracellular fluid movement, were upregulated in ciliated cells. In contrast, ciliated cell assembly and cilium movement in fluid were downregulated in stromal fibroblasts (Fig. 4A). In the early mid secretory phase, in contrast to the early secretory phase, embryo implantation and decidualization processes were upregulated, cilium assembly and epithelial cilium movement involved in extracellular fluid movement were downregulated in ciliated cells. In stromal fibroblasts, embryo implantation, decidualization, female pregnancy, cilium movement, and sperm-egg recognition were upregulated (Fig. 4B). In the mid- secretory phase, in contrast to the early mid secretory phase, embryo implantation, decidualization, pregnancy, and embryonic morphogenesis were upregulated, while epithelial cilium movement involved in extracellular fluid movement was downregulated in ciliated cells. All processes were downregulated in stromal fibroblasts (Fig. 4C). In the late secretory phase, in contrast to the mid secretory phase, the function of epithelial cilium movement involved in extracellular fluid movement, cilium movement, and embryonic appendage morphogenesis in ciliated cells were all upregulated, while embryonic processes involved in female pregnancy and protein localization to the cilium were downregulated in stromal fibroblasts (Fig. 4D). CellPhoneDB analysis of ligand/receptor expression revealed interactions between unciliated epithelium and ciliated epithelium (Fig. 5), and NOTCH1_JAG1, CD46_JAG1, and MIF_TNFRSF14 may be the most important ligands/receptors.
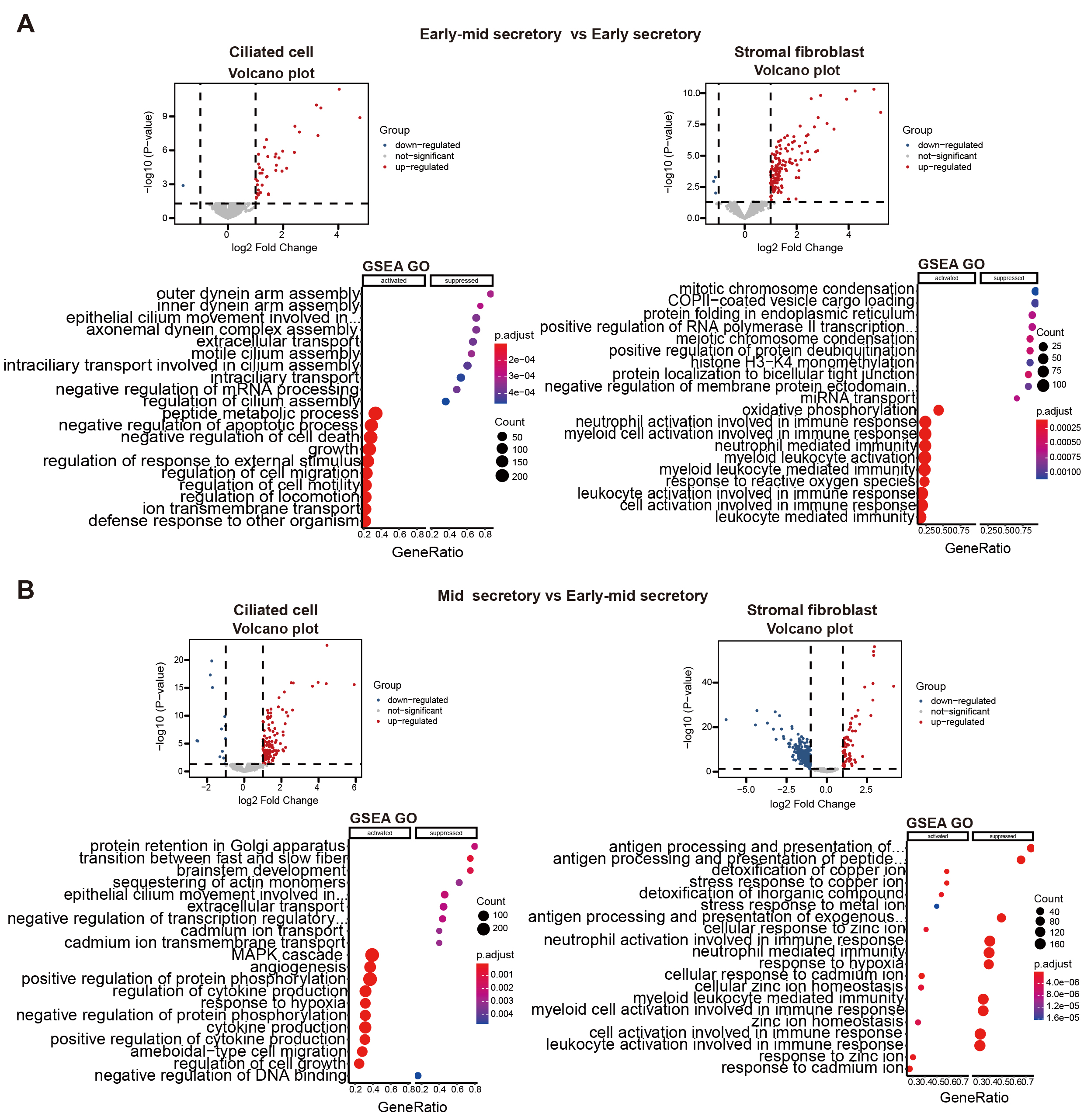 Fig. 3.
Fig. 3.Functional analysis of differential genes. Volcano plots, gene set enrichment analysis (GSEA) gene ontology
(GO) bubble diagrams of early mid secretory phase compared to early secretory phase (A), and mid secretory phase compared to early mid secretory
phase (B). log2 Fold Chang
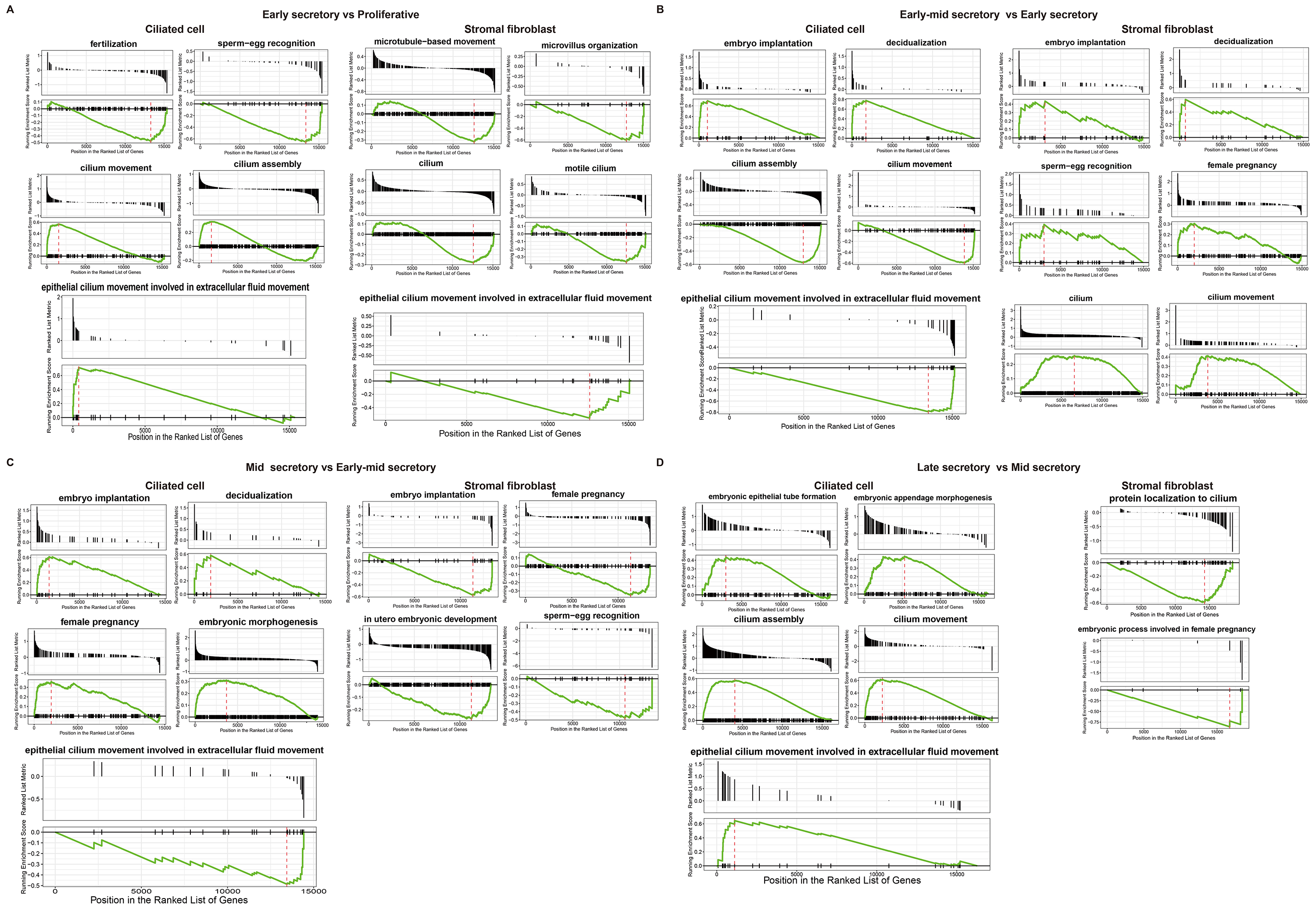 Fig. 4.
Fig. 4.Functional analysis of ciliated cells and stromal fibroblasts at different periods. Gseaplot2 plots of early secretory phase compared to proliferative phase (A). Early mid secretory phase compared to early secretory phase (B). Mid secretory phase compared to early-mid secretory phase (C). Late secretory phase compared to mid secretory phase in both ciliated cells and stromal fibroblasts (D).
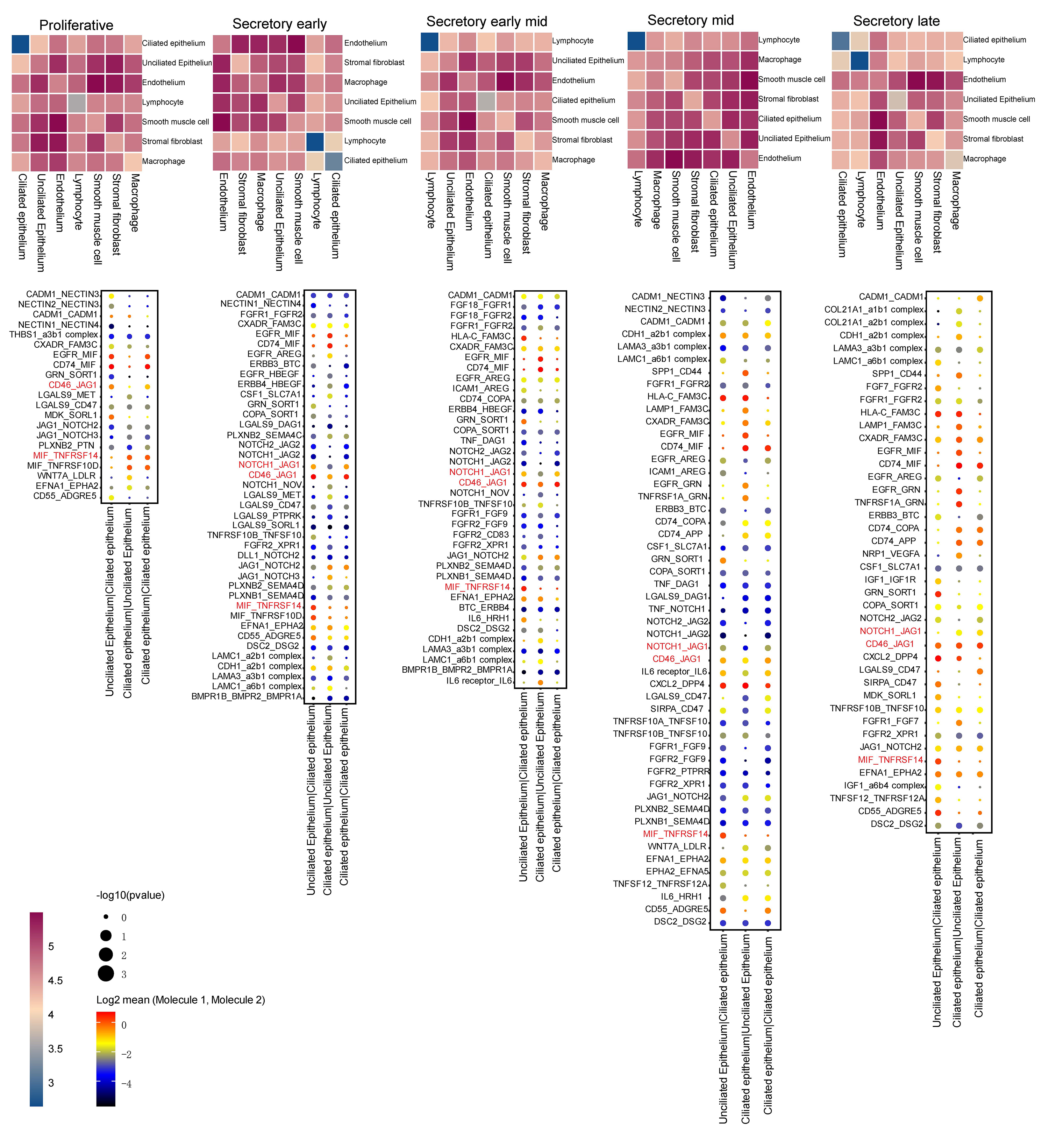 Fig. 5.
Fig. 5.CellphoneDB analysis of ligands/receptors between unciliated epithelium and ciliated epithelium. The darker the red, the more significant the up-regulation.
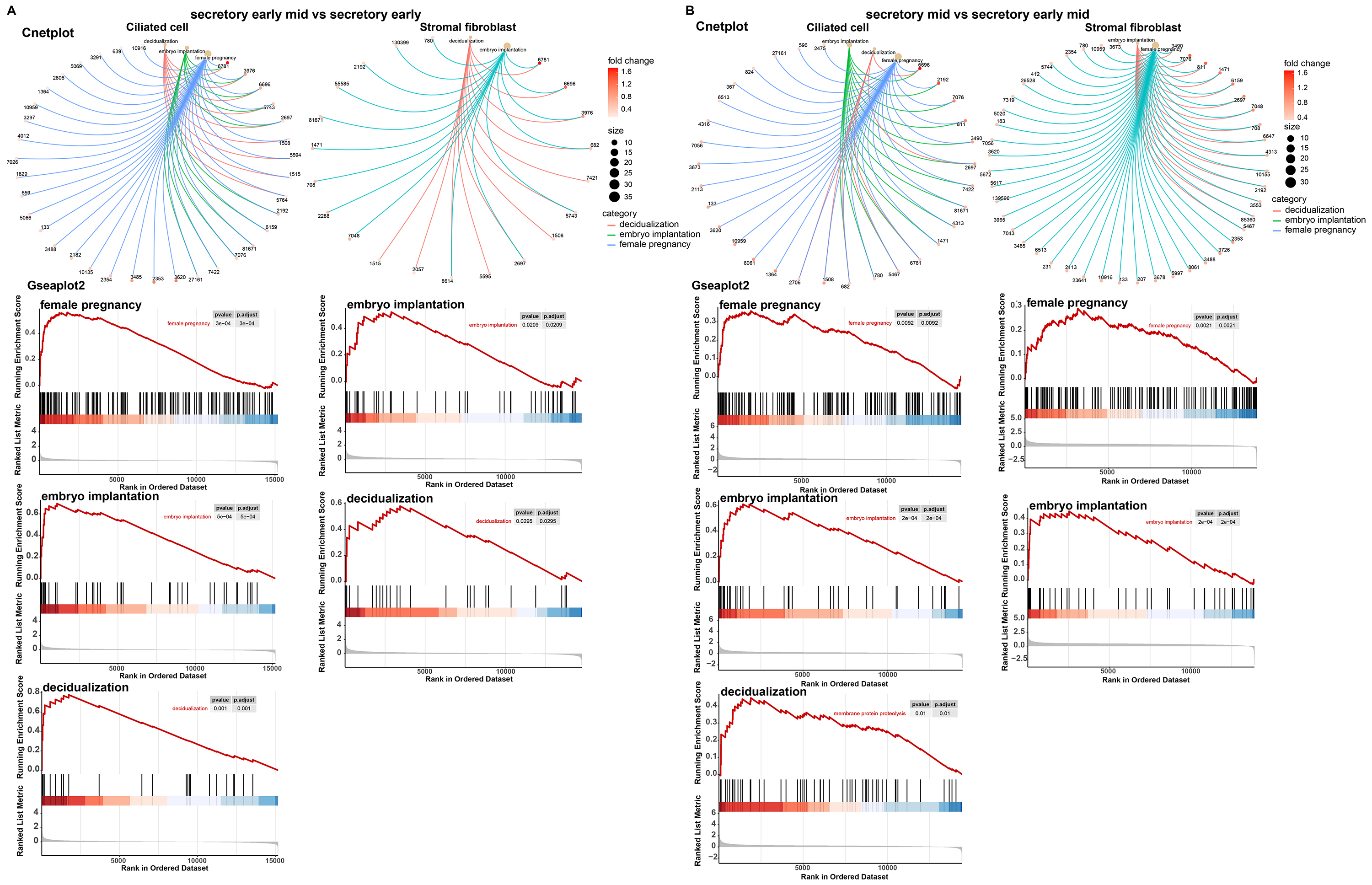
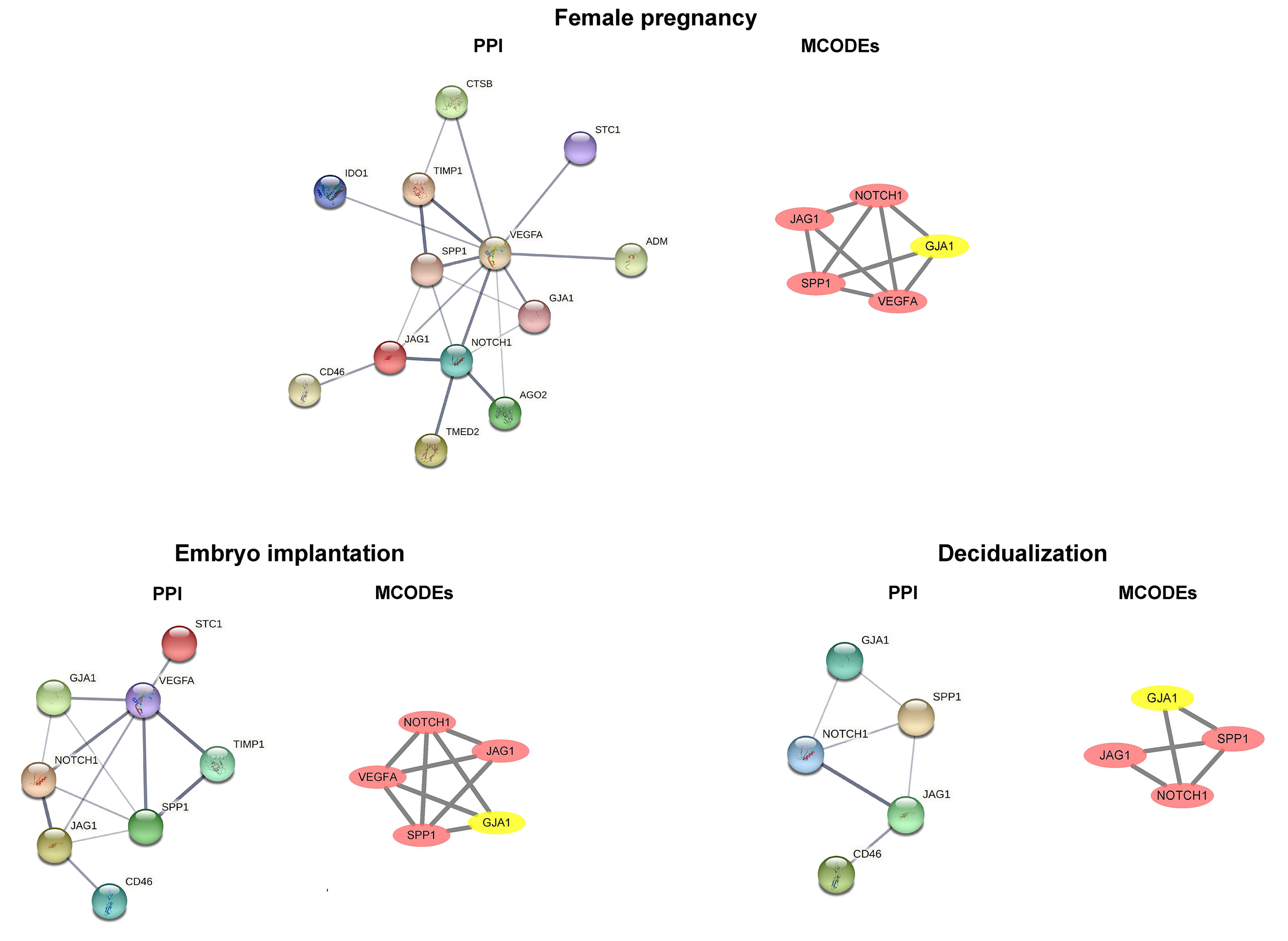
We performed a functional analysis of ciliated cells and stromal fibroblasts and constructed cnetplots and gseaplot2 graphs of the pregnancy, embryo implantation, and decidualization. In comparative analyses of the early mid secretory phase to the early secretory phase (Fig. 6A) and the mid secretory phase to the early mid secretory phase (Fig. 6B), we found that the three BPs were all upregulated. This was followed by mapping the PPI networks and MCODEs for the three biological processes to identify the hub genes, with all seed genes being marked with yellow (Fig. 7). According to our preliminary data, we found that GJA1, SPP1 were common gene of pregnancy, embryo implantation, and decidualization, NOTCH1_JAG1 might be the main communication ligand (Fig. 5). There’s a protein interaction between the GJA1, SPP1, NOTCH1 and JAG1, so they might play key roles in pregnancy.
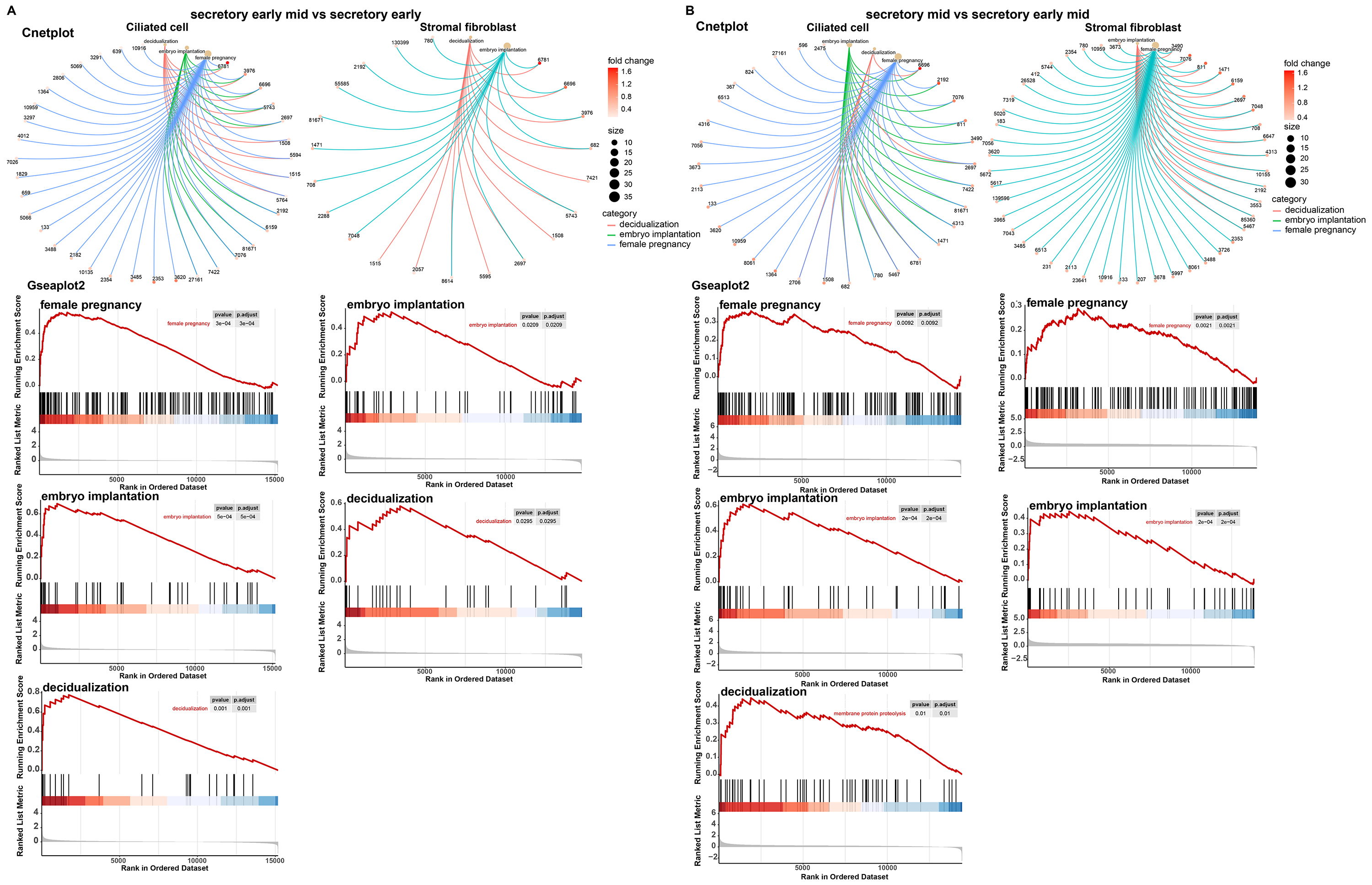 Fig. 6.
Fig. 6.Major biological processes and their involvement in genes. Cnetplot and gseaplot2 of pregnancy, embryo implantation, and decidualization in early mid secretory phase compared to early secretory phase (A). Mid secretory phase compared to early-mid secretory phase in ciliated cells and stromal fibroblasts (B).
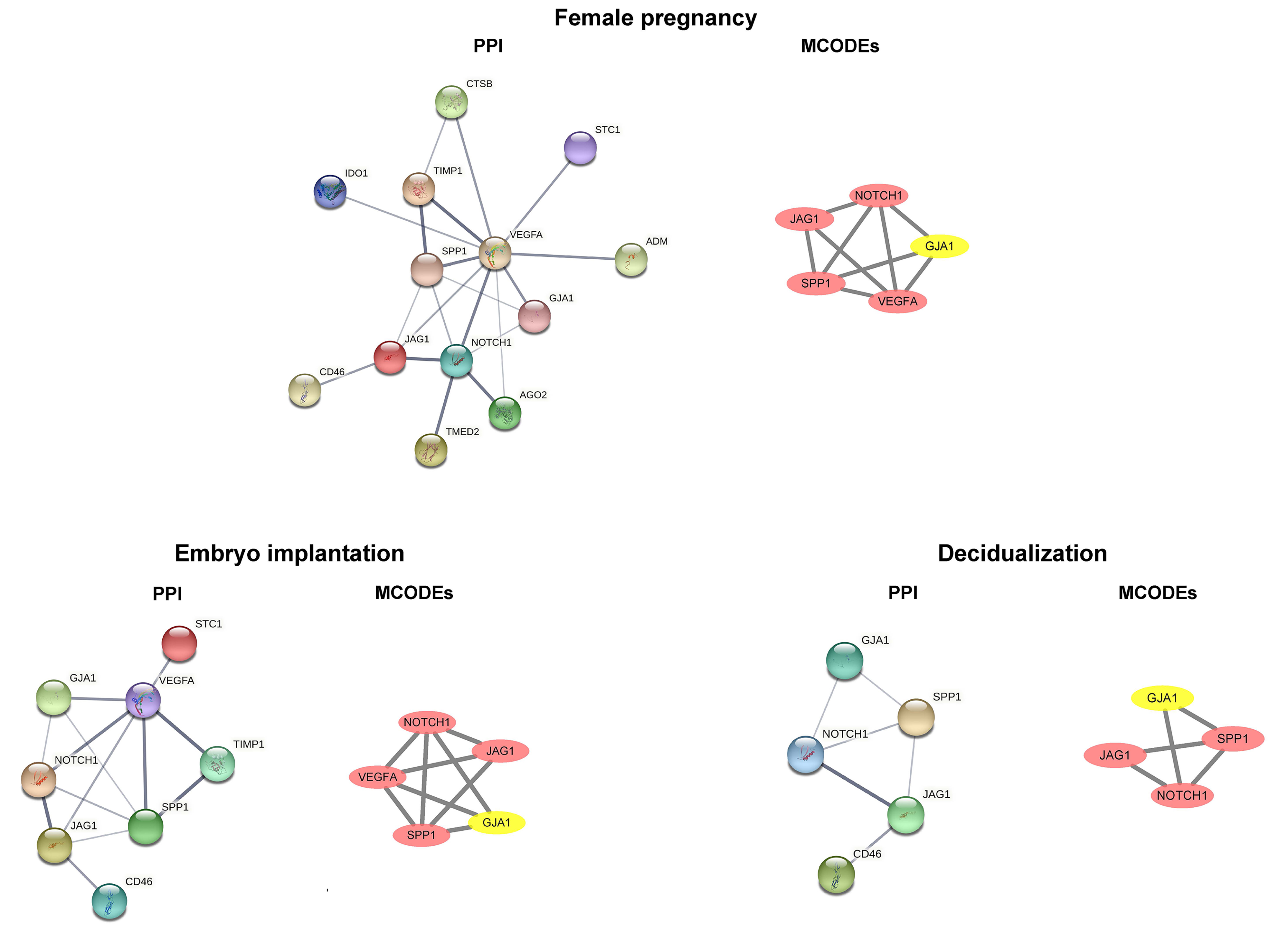 Fig. 7.
Fig. 7.Common DEGs identified by PPI networks and molecular complex detection (MCODEs). DEGs, differentially expressed genes; PPI, protein–protein interaction; GJA1, connexin 43; SPP1, secreted phosphoprotein 1; CTSB, Cathepsin B; STC1, Stanniocalcin‑1; IDO1, Indoleamine 2,3-dioxygenase 1; TIMP1, Tissue inhibitor of metalloproteinases-1; VEGFA, Vascular endothelial growth factor A; ADM, Acinar-to-ductal metaplasia; JAG1, Jagged-1; CD46, Membrane cofactor protein; NOTCH1, Neurogenic locus notch homolog protein 1; AGO2, Argonaute-2; TMED2, Transmembrane emp24 domain-containing protein 2.
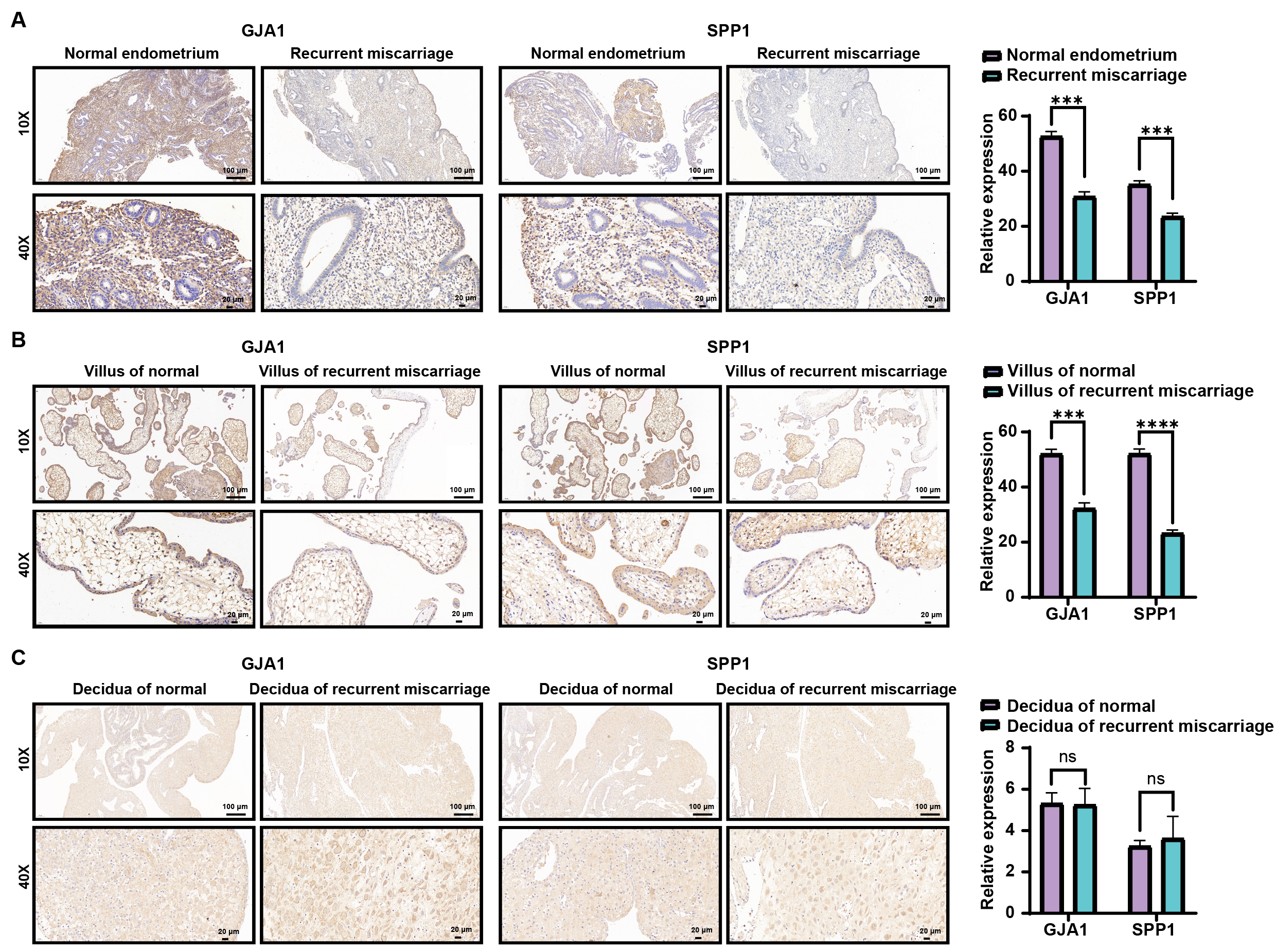
The results of immunohistochemistry showed that the staining intensity of GJA1 and SPP1 were significantly higher in the proliferative endometrium (Fig. 8A) and fetal villus (Fig. 8B) in normal pregnancy as compared to that of recurrent miscarriage, but no significant difference in decidua was noted (Fig. 8C).
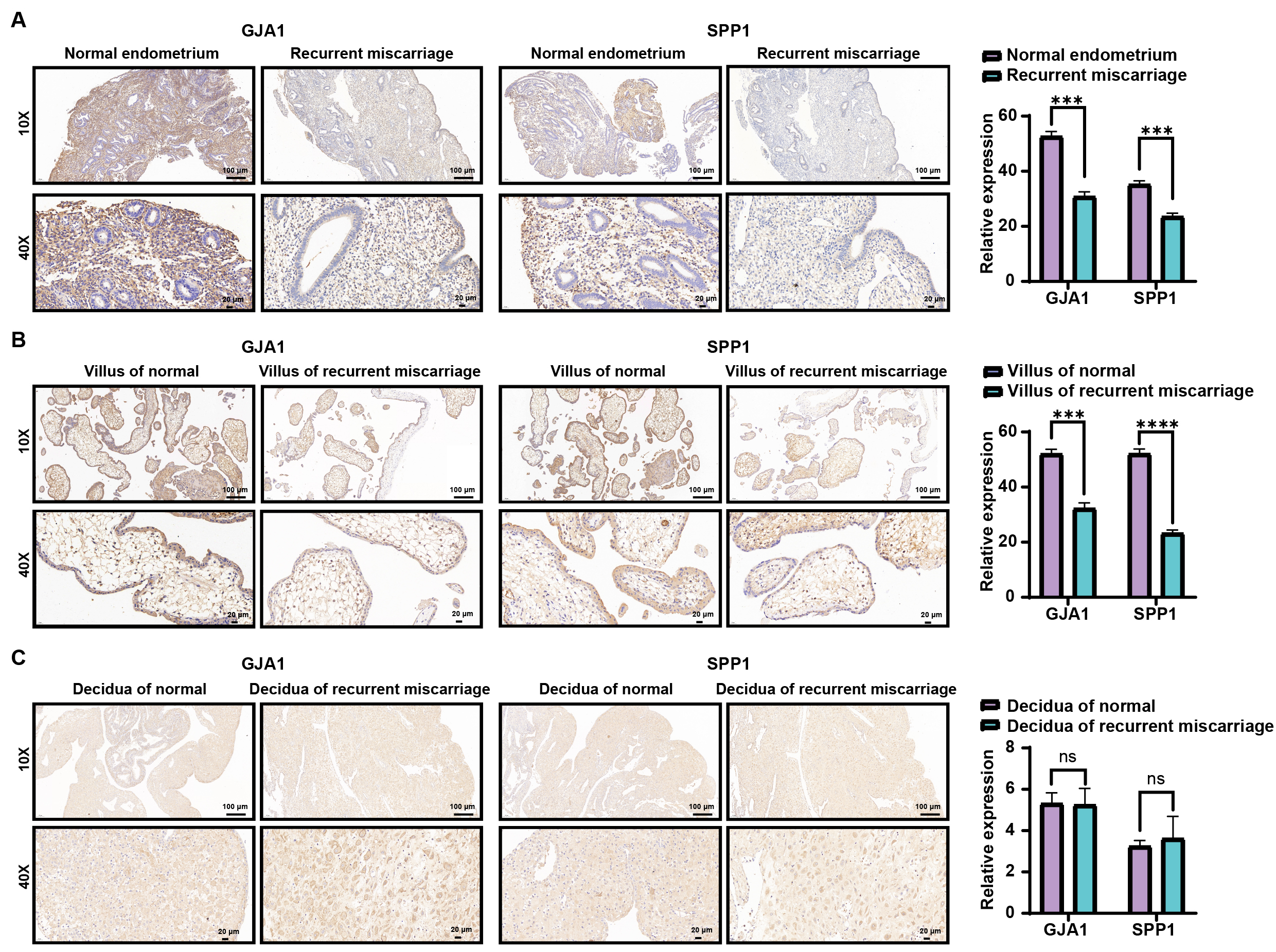 Fig. 8.
Fig. 8.Immunohistochemistry of proliferating endometrium (A), villus
(B), and decidua (C) between normal and recurrent miscarriage. *** represented
p
Surface cilia are most plentiful in the mid and late proliferative phases and the secretory phase, with a peak in the first part of the secretory phase [51]. Their relative lack in the early proliferative phase is best explained by the need for rapidly replicating cells. The reduction in cilia in mature secretory cells may be due to the fact that cilia are buried at the base of the gland, resulting in a loss of surface numbers [34]. In our study, the proportion of cilia cells in each phase of the endometrium was analyzed statistically, and we determined that the proportion peaked in the mid secretory phase (Fig. 2). The increased number of ciliated cells in the middle secretory stage, the movement of ciliated cells, and the proteins expressed by ciliated cells, all may play an important role in the early recognition and communication of embryos entering the implantation site.
In our study, the staining intensity of GJA1 and SPP1 in proliferative
endometrium and fetal villus was higher in the normal pregnancy than that in
recurrent miscarriage, suggesting that GJA1 and SPP1 may play an important role
in preparation of the endometrium before successful implantation. However, no
difference in GJA1 and SPP1 expression in the decidua was identified, which may
be related to our inability to accurately obtain the decidua at the implantation
site. Previous studies have explored the regulation of GJA1 and SPP1. For
example, GJA1 promotes cell adhesion through the PI3K/AKT/NF-
In our bioinformatics analysis, GJA1 and SPP1 were involved in the biological processes of pregnancy, embryo implantation and decidualization. They directly interacted with NOTCH1 and JAG1, as observed in the PPI network (Fig. 7). In the analysis of CellphoneDB (Fig. 5), NOTCH1_JAG1 plays an important role in all stages of the endometrium, and some studies have shown that the NOTCH1 may regulate GJA1 expression [61]. The JAG1/NOTCH1 cascade represents a potential therapeutic target for hepatocellular carcinoma metastasis [62], and we speculate that the expression of GJA1 and SPP1 in the endometrium may occur through the regulation of NOTCH1. However, this hypothesis needs to be supported by more data and verified by further experiments.
The number of ciliated cells may peak in the mid- secretory phase of the endometrium, and they may play an important role in the early recognition process of embryos prior to implantation. In addition, the expression of GJA1 and SPP1 in the endometrium may play an important role in successful implantation and embryonic development. These conclusions require further confirmation by a large number of subsequent experiments.
The data that support the findings of this study are available from the corresponding author. And the datasets GSE111976 was accqured at NCBI’s Gene Expression Omnibus.
ZL, XZ, QT and YG designed the research study. ZL, XZ and QT analyzed the data. CC and QW provided help and advice on the immunohistochemistry experiments. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All subjects gave their informed consent for inclusion before they participated in the study. The study was conducted in accordance with the Declaration of Helsinki, and the protocol was approved by the Ethics Committee of Union Hospital, Tongji Medical College, Huazhong University of Science and Technology (approval number: 0241-01).
Thanks to Yanfang Lu for his help in the data analysis process.
This research received no external funding.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.