
 , Rajitha Rajeshwar Tatikonda 1,2, Raji Reddy Chada 2,3,*
, Rajitha Rajeshwar Tatikonda 1,2, Raji Reddy Chada 2,3,* , Srinivasa Reddy Bonam 1,2,*
, Srinivasa Reddy Bonam 1,2,*1 Vaccine Immunology Laboratory, Department of Applied Biology, CSIR-Indian Institute of Chemical Technology, 500007 Hyderabad, Telangana, India
2 Academy of Scientific and Innovative Research (AcSIR), 201002 Ghaziabad, India
3 Department of Organic Synthesis & Process Chemistry, CSIR-Indian Institute of Chemical Technology, 500007 Hyderabad, Telangana, India
Abstract
Immunomodulators play a central role in the treatment of cancer and immune-mediated disorders. Small-molecule immunomodulators are particularly attractive due to their structural diversity, pharmacological versatility, and potential for oral administration. This study aimed to investigate the immunomodulatory potential of newly synthesized aza-spirocyclic derivatives.
A series of novel aza-spirocyclic compounds was evaluated for cytotoxicity in RAW264.7 murine macrophages. Immunomodulatory activity was assessed by quantifying nitric oxide (NO) production and profiling pro- and anti-inflammatory cytokines in macrophages and murine splenocytes. In vitro anti-inflammatory effects were further examined using a lipopolysaccharide (LPS)-induced inflammation model. Mechanistic insights were explored using molecular docking and immuno-informatics analyses.
All compounds were non-toxic at the tested concentrations. Most derivatives failed to induce NO production, indicating an absence of danger-associated molecular pattern (DAMP)-like activity. Several compounds significantly reduced the pro-inflammatory cytokine interleukin-6 (IL-6) while increasing the anti-inflammatory cytokine IL-10 in vitro. In the LPS-induced inflammation model, four compounds notably suppressed IL-6 and enhanced IL-10 expression. The binding affinities obtained from docking, along with in vitro validation of these compounds on Janus kinases (JAKs) signaling, suggested that compounds AS8 and AS10 modulated inflammatory signaling pathways.
The synthesized aza-spirocyclic derivatives exhibit promising anti-inflammatory profiles, characterized by IL-6 suppression and IL-10 enhancement. These findings position aza-spirocyclic scaffolds as candidates for the development of next-generation small-molecule immunomodulators.
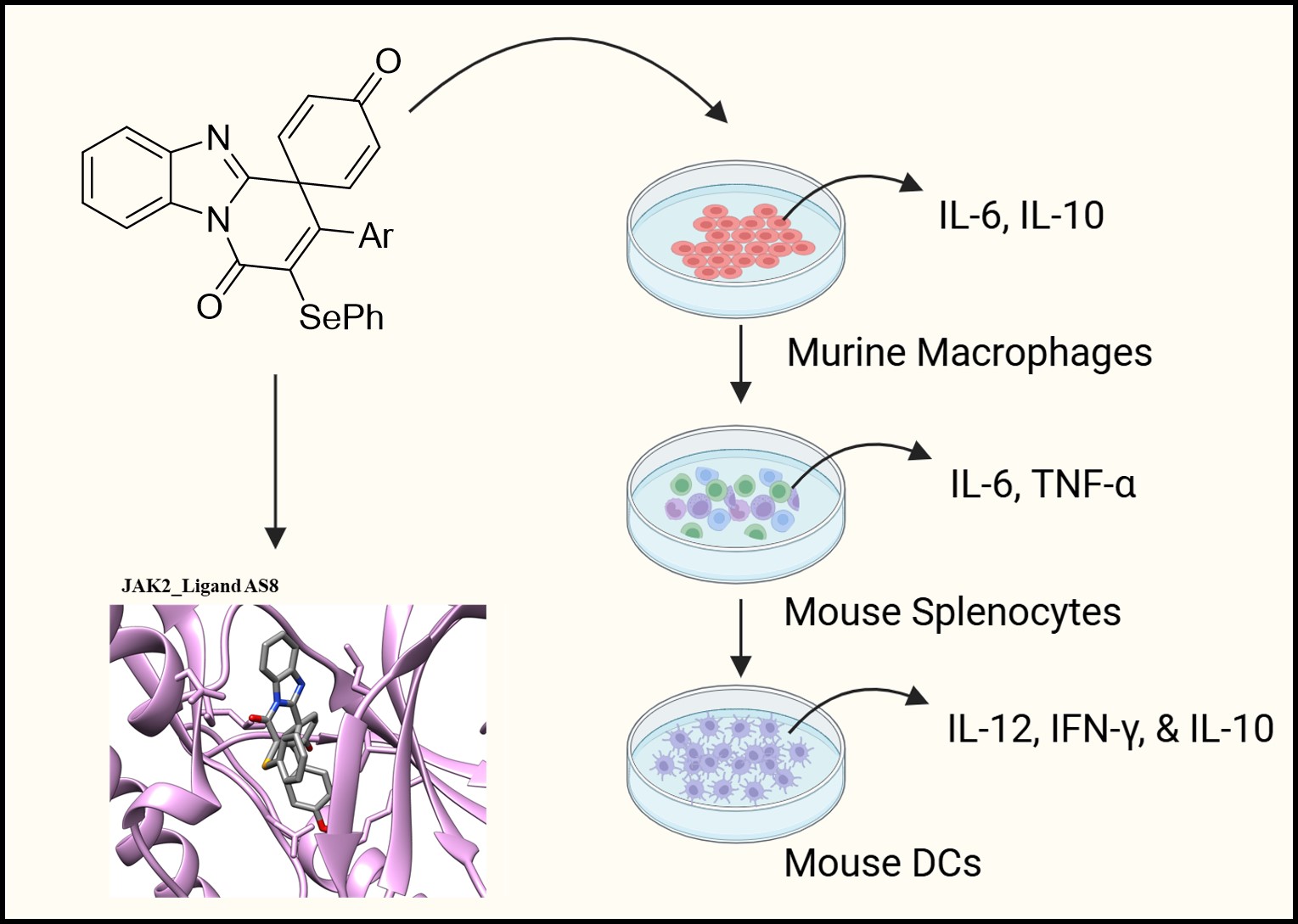
Graphical Abstract
Keywords
- aza compounds
- immunologic
- anti-inflammatory agents
- cytokines
- janus kinase inhibitors
Immunomodulators are prophylactic or therapeutic agents, derived from natural or
synthetic, that regulate immune system activity either by enhancing immune
responses (immunostimulation) or suppressing them (immunosuppression). The
development of novel small-molecule immunomodulators holds considerable promise
for the treatment of autoimmune disorders, chronic inflammatory diseases, and
cancer, as well as for use as vaccine adjuvants. Among the chemical scaffolds
explored, derivatives of aza-spirocycles have drawn ample interest because of
their broad spectrum of biological activity and physical properties [1].
Moreover, spirocyclic frameworks have emerged as a valuable design strategy in
bioactive small molecules, enabling enhanced molecular complexity, improved
physicochemical properties, and refined modulation of biological activity.
Aza-spirocycles represent an important class of heterocyclic frameworks, widely
recognized for their presence in numerous pharmacologically active compounds [2].
The immunomodulatory potential of aza-spirocyclic compounds can be attributed to
their capacity to interact with key molecular targets involved in immune
regulation, such as cytokine signaling pathways, transcription factors (e.g.,
NF-
The aza-spirocyclic compounds were synthesized using the recently developed ipso-annulation reactions [4, 5]. In the first case, radical-promoted ipso-annulation using aryl sulfonates or AgSCN or AgSCF3 in the presence of ceric ammonium nitrate (CAN) as an oxidant, provided thio-functionalized (SO2Ar or SCN or SCF3) spirocycles in good yields [4]. Next, selenylated aza-spirocycles were prepared via electrophilic ipso-annulation using diphenyl diselenide/FeCl3 reaction conditions. Accordingly, spirocycles AS1 to AS6 were obtained from the reaction of N-benzylacrylamides induced by S-centered radicals (SCN/SCF3/SO2Ar) in the presence of CAN as an oxidant. Next, the chalcogenated (SCN/SCF3/SePh) benzimidazo-fused azaspiro[5,5]undecatri-enones AS7 to AS12 were synthesized by the reaction of N-propiolyl-2-arylbenzimidazoles involving both the radical-based and electrophilic reactions. All the general synthetic strategies (Scheme S1 & S2) are provided in the Supplementary Information along with the analytical data (1H NMR, 13C NMR & mass) along with the HPLC data for the representative compounds.
The cell line was obtained from the National Centre for Cell Science (NCCS, Pune, India). All cell lines supplied by NCCS were authenticated using Short Tandem Repeat (STR) profiling and tested for mycoplasma contamination [6]. RAW264.7 cells were maintained in Dulbecco’s Modified Eagle Medium (DMEM) (AT241-1L, Grand Island, NY, USA) medium supplemented with 10% fetal bovine serum (FBS) (Gibco-16170078) and 1% Penicillin-Streptomycin (Pen-Strep) (Gibco-15140-122, Thermo Fisher Scientific, Grand Island, NY, USA) solution under standard conditions of 37 °C and 5% CO2 for 24 hours. Cells were cultured in T75 flasks containing 10 mL of media. Then, cells were counted using a haematocytometer. Ten thousand cells per well were cultured in 96-well plates for 24 hours under optimum conditions. Also, compounds were serially diluted in culture media starting with 100 µM up to 0.8 µM concentration. Then, cells were treated with decreasing concentrations of the compounds and incubated for 24 hours at 37 °C and 5% CO2. Control wells contained only untreated cells. Following 24 hours of incubation, 50 µL of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) (Sigma, M5655, Milwaukee, WI, USA) solution (5 mg/mL concentration in PBS) was added into each well and incubated for 4 hours at 37 °C and 5% CO2 for 24 hours. Later, ~100 µL of media was aspirated and discarded, and formazan crystals were dissolved in 50 µL of dimethyl sulfoxide (DMSO) and incubated for 15 minutes. Absorbance (O.D.) was then measured at 570 nm using a microplate reader (MEGALLAN, Tecan Life Sciences, Männedorf, Switzerland). Percentage cell viability was then calculated based on the compound treatment’s absorbance relative to the cell control’s absorbance [7].
RAW264.7 cells were maintained as mentioned in the above section. Cells were
counted using a haematocytometer, approximately 0.5
Animal experiments were approved by the Institutional Animal Ethics Committee
(IEAC) of the Council of Scientific & Industrial Research, Indian Institute of
Chemical Technology (CSIR-IICT). Three C57BL/6 mice were acquired from the
internal animal house facility (CSIR-IICT, Hyderabad) and used for isolating
spleens for immunological study (IAEC approval number: IICT-IAEC-069-2025). Mice
(6–8 weeks old, female) were sacrificed with cervical dislocation, and the
spleen was dissected carefully. Splenocytes were isolated by using Ammonium-Chloride-Potassium
(ACK) lysis buffer [prepared in the lab]. Later, cells
were counted, and approximately 1
Secreted cytokines were measured by sandwich enzyme-linked immunosorbent assay
(ELISA). For this, RAW264.7 cells and Splenocytes (0.5
Molecular docking was performed using AutoDock Vina (Center for Computational Structural Biology, Scripps Research, La Jolla, CA, USA; https://vina.scripps.edu/) [11, 12], which employs a scoring function that combines empirical and knowledge-based terms to estimate the binding affinity between ligands and the protein. It utilizes a gradient-based optimization algorithm to efficiently explore the ligand’s binding conformations. The 2D structures of the 12 compounds were obtained using ChemDraw (Revvity Signals Software, Inc., Waltham, MA, USA), and their 3D structures were generated with OpenBabel (http://openbabel.org). The experimental crystal structures of JAK1 kinase (PDB: 6N7A) [13], an isoquinoline derivative inhibitor with JAK2 kinase (PDB: 2B7A) [14], and JAK3 (PDB: 5LWM) [15] in complex with a pyrazolopyridine inhibitor and TYK2 (PDB: 6AAM) in complex with peficitinib were considered as starting structures for docking. The protein structures were processed by modeling missing residues, removing all heteroatoms and co-crystal ligands, and converting them to the required PDBQT format using Mgltools. All compounds were also converted to PDBQT format. The binding site was defined by obtaining the center of mass of the respective co-crystal ligand for each JAK isoform. The exhaustiveness parameter was set to the default value of 8. The docking results were visualized with UCSF Chimera, and interaction analysis was performed using BIOVIA Discovery Studio (2024, Dassault Systèmes BIOVIA, San Diego, CA, USA).
The mouse macrophages (RAW264.7) cells were seeded in a 12-well plate at a
density of 1
Data were analysed using GraphPad Prism 8 (GraphPad Software, LLC, San Diego, CA, USA). Comparison of two groups was performed by Student’s t-test, an unpaired, non-parametric test, and the Mann-Whitney test.
The azaspirocyclic framework is a structural motif found in various natural and synthetic molecules known for their significant pharmacological activities [16]. In addition, thio- and seleno-functionalized molecules are valuable in medicinal chemistry because sulfur and selenium introduce distinctive electronic and redox properties that considerably modify the pharmaceutical properties. These groups can improve lipophilicity, metabolic stability, and target interactions, while offering useful bioisosteric options for tuning potency and selectivity [17]. In light of the favorable properties associated with this class of molecules, we have developed an efficient protocol to access a series of thio-functionalized azaspiro[4,5]decatrienones via the dearomative ipso-annulation of unactivated N-benzyl acrylamides, induced by S-centered radicals (SCN/SCF3/SO2Ar) in the presence of CAN as an oxidant [4]. Later, we reported an ipso-carbocyclization of N-propiolyl-2-arylbenzimidazoles leading to chalcogenated (SCN/SCF3/SePh) benzimidazo-fused azaspiro[5,5]undecatrienones in good yields, involving both radical-based and electrophilic pathways [4, 5]. Having developed these two ipso-annulation strategies to synthesize both the azaspiro[4,5]decatrienone and azaspiro[5,5]undecatrienone frameworks bearing chalcogenated (S/Se) functionalities, we became interested in investigating their immunomodulatory effects. In particular, the benzimidazole-fused azaspiro[5,5]undecatrienones incorporate two high-value pharmacophoric motifs within a single molecular framework. These motifs are widely represented in bioactive small molecules allied with diverse pharmacological activities, thereby underscoring the potential relevance of these hybrid structures. In addition, the chalcogen-functionalized azaspirocycles obtained through our synthetic strategy provide promising structural templates for medicinal chemistry optimization. Their unique combination of rigidity, three-dimensionality, and heteroatom incorporation positions them as attractive candidates for future drug-discovery and lead-generation efforts (Supplementary Table 1).
The cytotoxic potential of the spirocyclic compounds (Supplementary Table 1) was evaluated in murine macrophages using a colorimetric assay based on the reduction of 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl-tetrazolium bromide (MTT) by mitochondrial dehydrogenases, leading to the formation of blue formazan crystals. All tested compounds preserved cell viability above 90% across concentrations up to 100 µM. None of the compounds exhibited measurable cytotoxicity within this range, and the half-maximal inhibitory concentration (IC50) values were estimated to be greater than 1000 µM (Supplementary Table 2), indicating a favorable safety profile for in vitro studies.
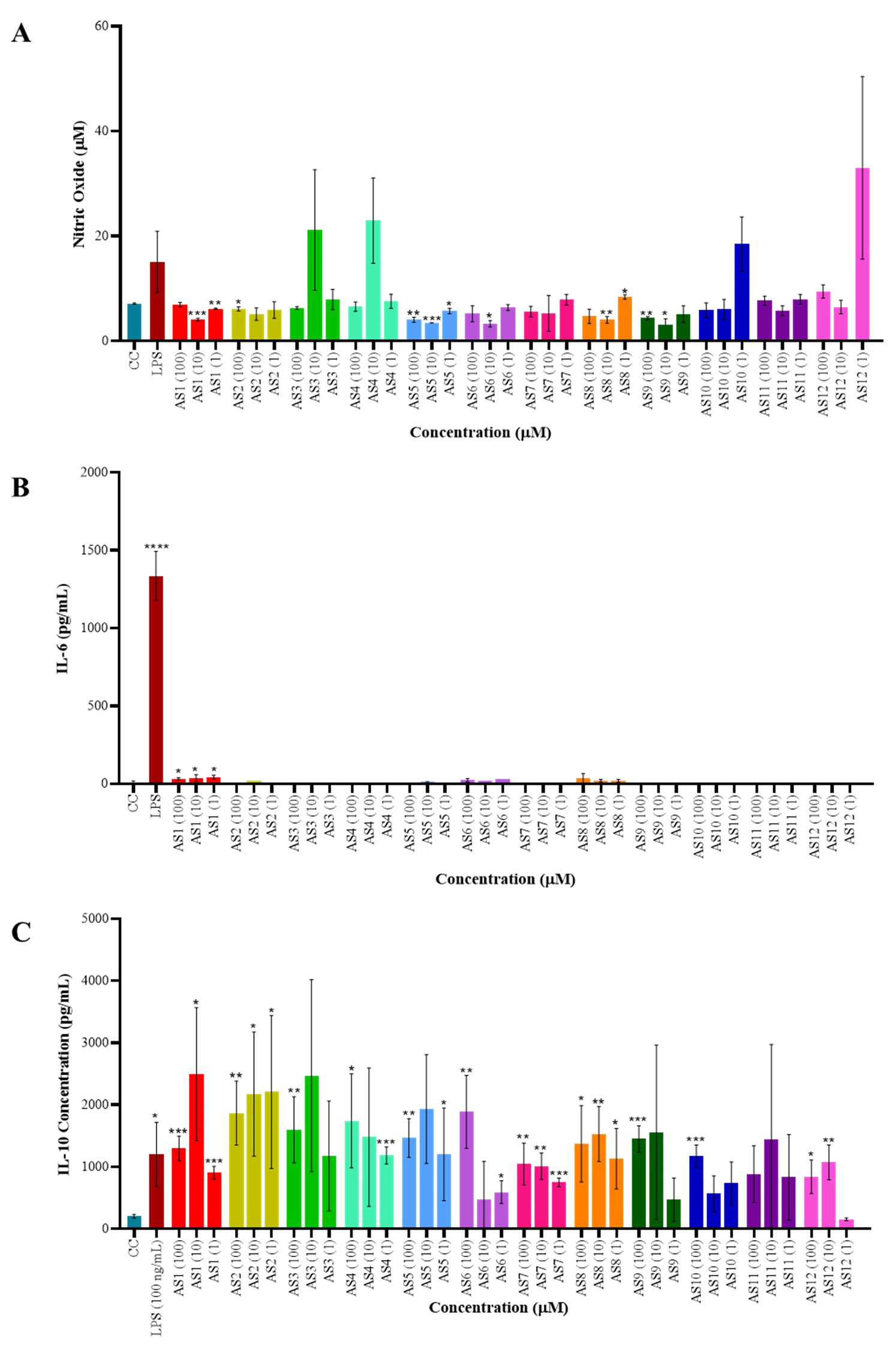
The impact of spirocyclic compounds on reactive nitrogen species (RNS) production was assessed using a nitric oxide (NO) assay in RAW264.7 macrophages. None of the tested compounds induced NO production, however, a few of the compounds showed slightly higher NO, although it was not significant. These findings indicate that the spirocyclic compounds do not function as danger-associated molecular pattern (DAMP) or pathogen-associated molecular pattern (PAMP) inducers (Fig. 1A). To further investigate their immunomodulatory potential, culture supernatants from treated macrophages were analyzed for inflammatory cytokine secretion, specifically interleukin-6 (IL-6) and interleukin-10 (IL-10) (Fig. 1B,C), using a sandwich ELISA. IL-6 production remained largely unchanged across treatments, with the exception of compound 1, which elicited a modest increase compared to the cell control. In contrast, IL-10 secretion was significantly elevated in a dose-dependent manner following treatment with the spirocyclic compounds (AS1, AS2, AS3, AS4, AS5, AS6, AS8, AS9, and AS10), suggesting an anti-inflammatory skewing of the compound on macrophage response. Also, LPS induced the significant IL-10 secretion, which may be due to a response of anti-inflammatory macrophages that usually appear at the late stage of LPS stimulation (after 24 hours) [18].
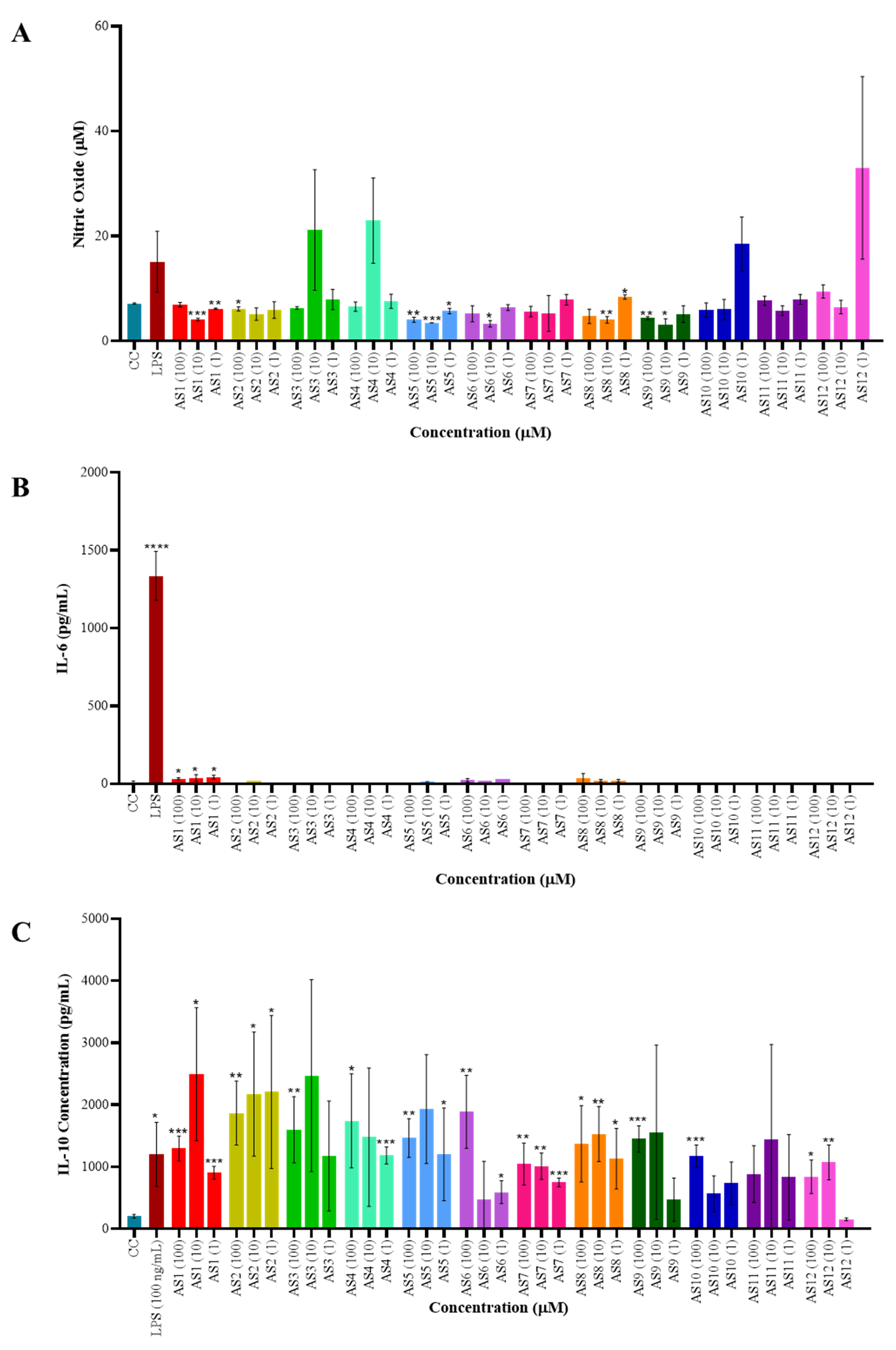 Fig. 1.
Fig. 1.
Immunomodulatory activity of spirocyclic
compounds—cytokine production analysis. (A) Nitric oxide (NO) production was
assessed using the Griess reagent assay. RAW264.7 macrophages (0.5
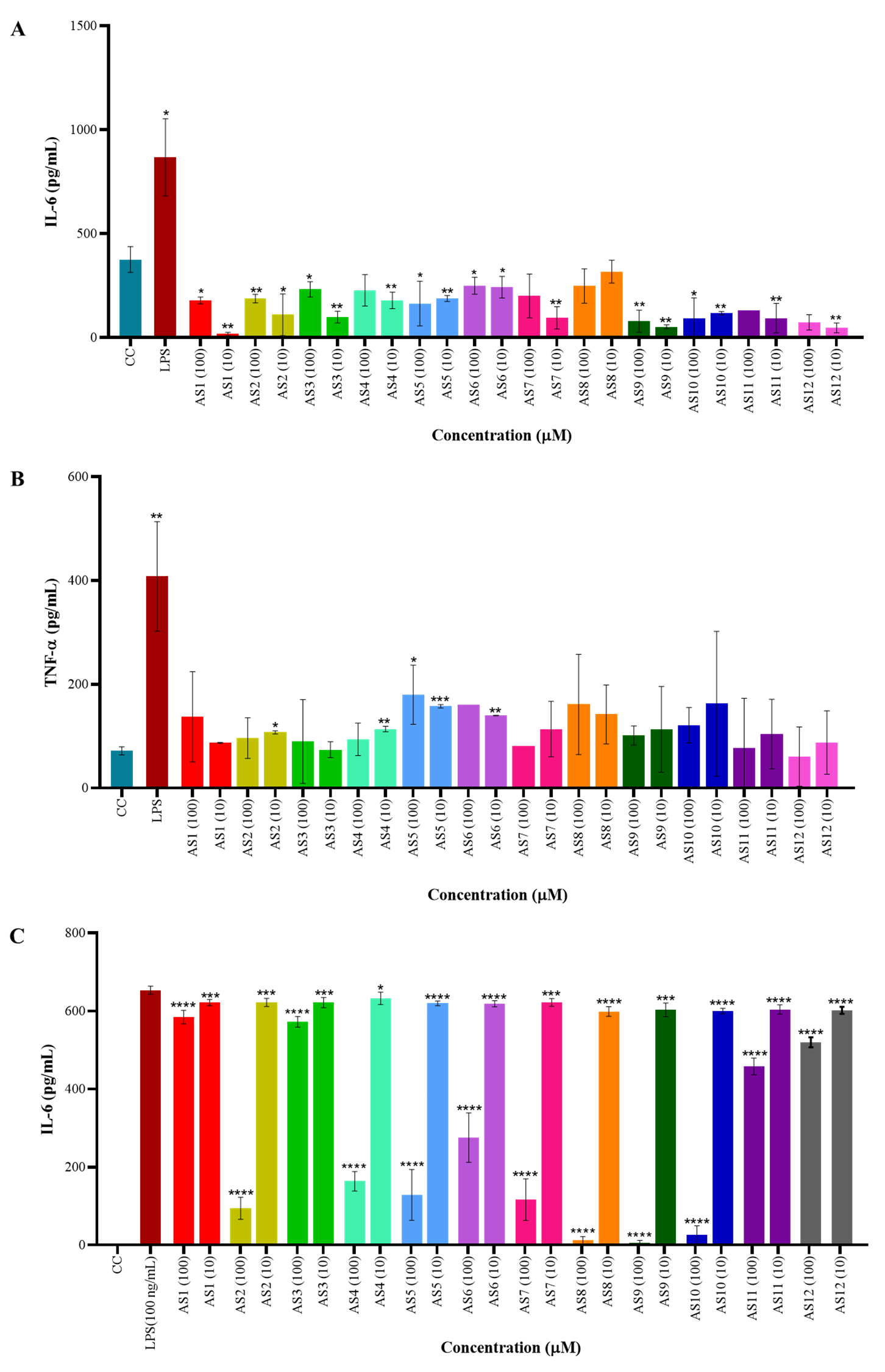
The impact of spirocyclic compounds on cytokine secretion by murine splenocytes
was assessed by quantifying IL-6 and tumor necrosis factor-alpha (TNF-
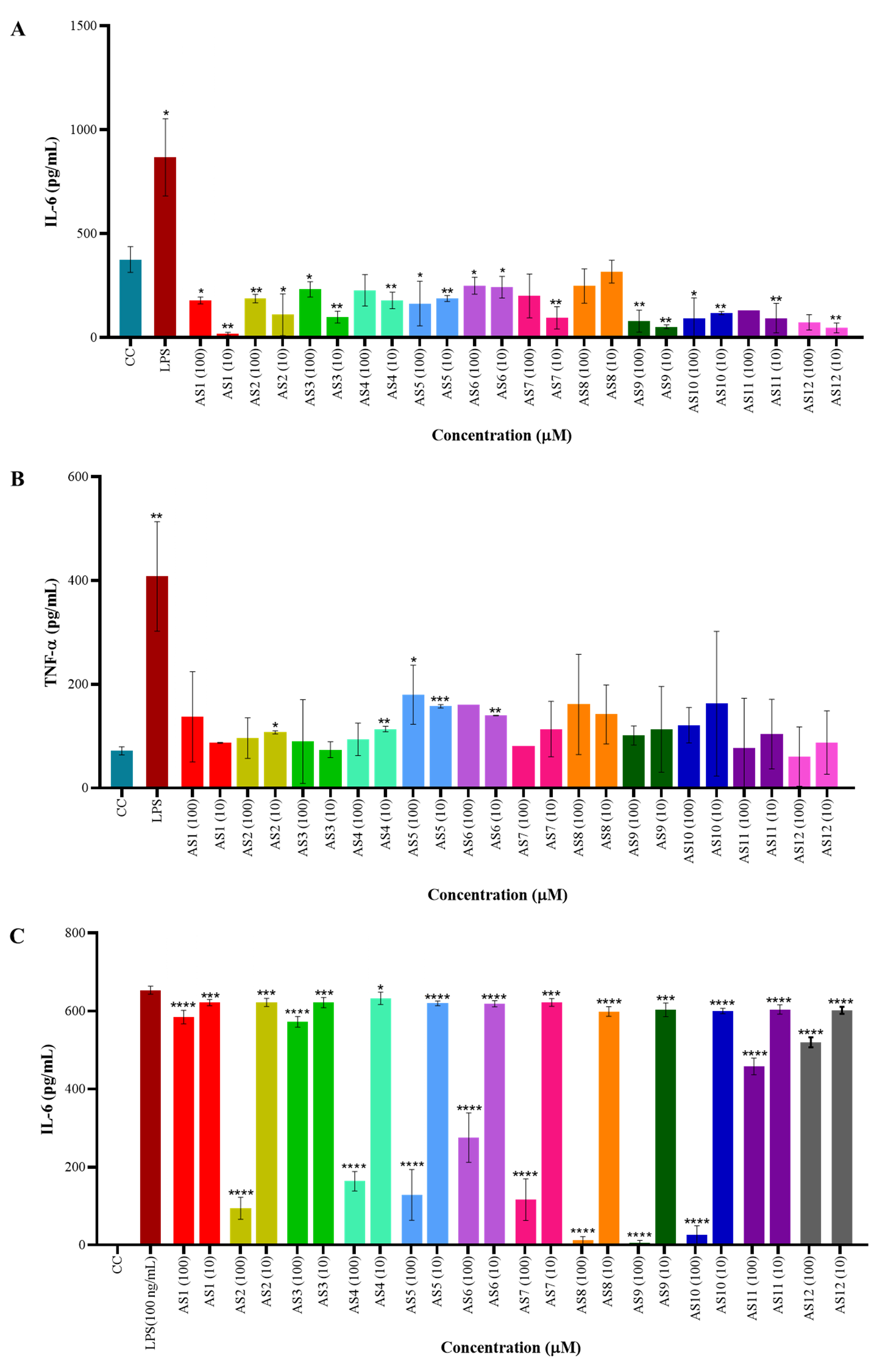 Fig. 2.
Fig. 2.
Anti-inflammatory activity of spirocyclic compounds on murine
splenocytes and RAW264.7 macrophages. (A,B) Quantification of pro-inflammatory
cytokines IL-6 and tumor necrosis factor-alpha (TNF-
The compounds induced significant IL-10 production in RAW264.7 macrophages,
while some also inhibited IL-6 production (
Molecular docking was performed on all 12 compounds with JAK1, JAK2,
JAK3 and TYK2 proteins to elucidate their binding interactions. The top ten
scoring molecules for each protein were identified, and the common candidates
with the potential to bind strongly to all JAK isoforms were shortlisted. These
included compounds AS1, AS2, AS7, AS8,
AS10, AS11, and AS12. Supplementary Table 3
provides the docking scores of these seven molecules with all JAK isoforms. It is
evident that these compounds bind to JAK proteins with varying affinity. The
binding scores for these top seven molecules ranged from –6.9 kcal/mol to –9.3
kcal/mol. Specifically, it is observed that all these molecules exhibited binding
scores
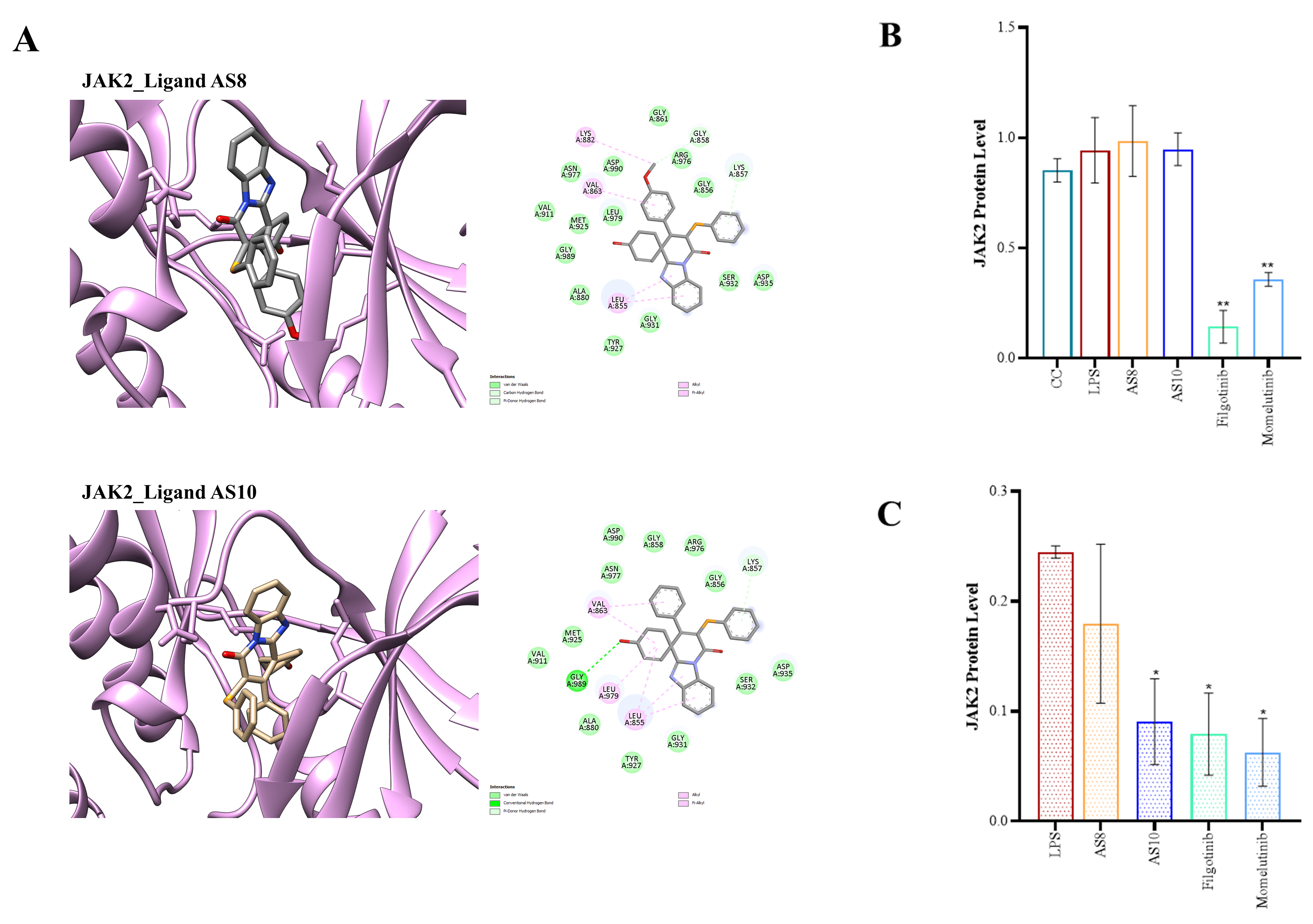
The interaction analysis reveals that while both hydrogen, hydrophobic and pi interactions are present, the binding is majorly driven by hydrophobic interactions. A full summary of each of the compound AS1, AS2, AS8, and AS10 interactions with all four isoforms is provided in the Supplementary Table 4. Moreover, Fig. 3A shows the binding pose for compounds AS8 and AS10 with the JAK2 protein, along with a 2D interaction map [20].
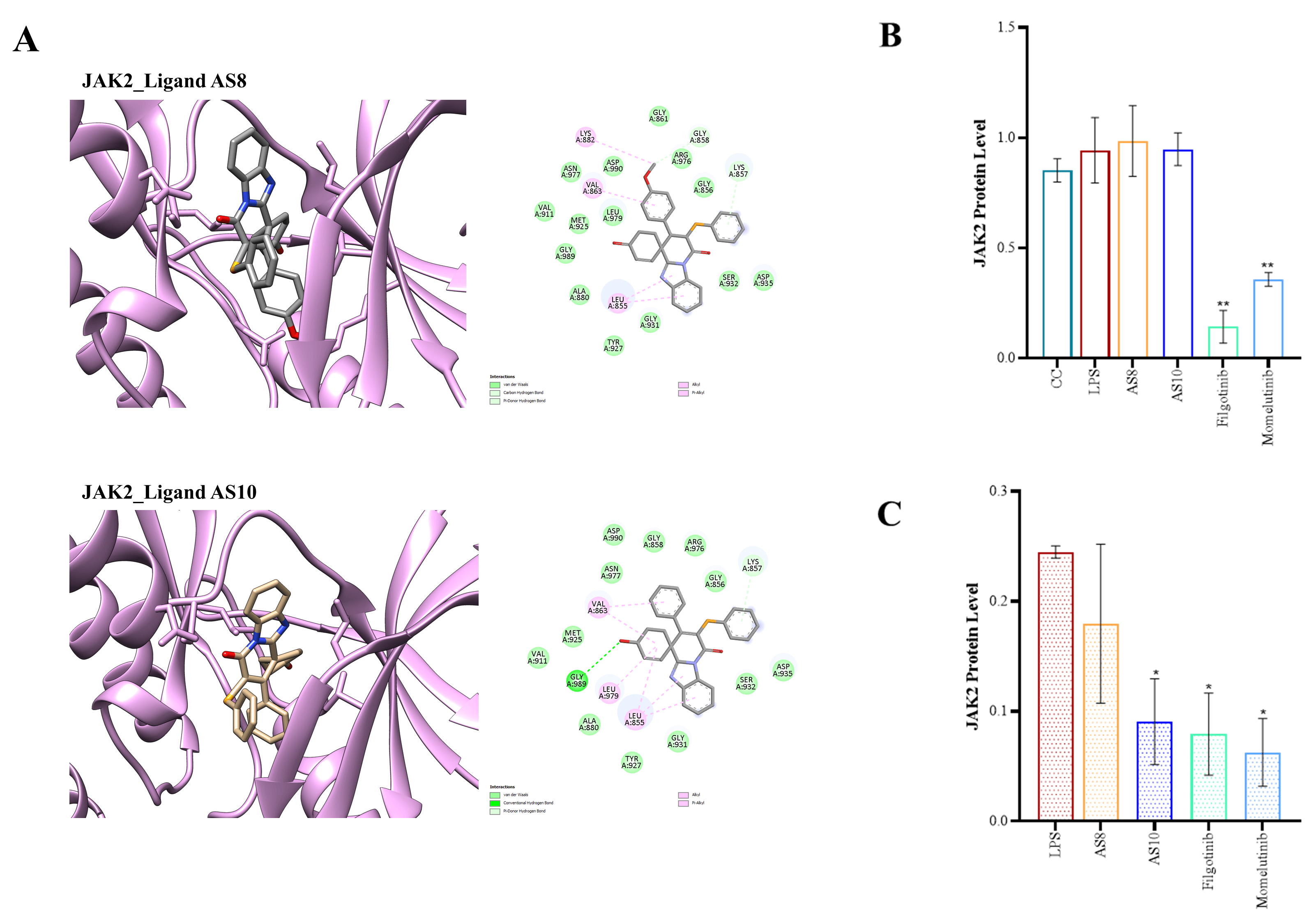 Fig. 3.
Fig. 3.
In silico molecular docking and in vitro
evaluation of the interaction between JAK kinase and the spirocyclic compounds.
(A) The binding poses of compounds AS8 and AS10 in JAK2 proteins, along with
corresponding 2D interaction maps. (B,C) Bar graph representation after
quantification of JAK2 protein expression level in the murine macrophage cell
line (All western blots, including uncropped, are available in the
Supplementary Information). 0.5
In silico docking, supported by prior inhibitory activity data,
indicated that AS1, AS2, AS8, and AS10 are
potential inhibitors of the JAK/STAT pathway. To validate these findings, the
expression of key genes and proteins involved in this pathway was evaluated
following compound treatment. No significant changes were observed at the mRNA
level (Supplementary Fig. 1). However, a pronounced reduction in total
JAK2 protein expression was observed upon treatment with AS8 and
AS10 (Fig. 3B,C and Supplementary Fig. 2). However,
the phosphorylation status of JAK2 following compound treatment remains to be
evaluated. In summary, AS8 and AS10 showed no cytotoxicity and
significantly increased IL-10 while suppressing LPS-induced IL-6 in macrophages
and splenocytes, indicating potent anti-inflammatory activity. Both compounds
exhibited strong binding affinity to JAK isoforms (
Previously, the ipso-annulation of benzimidazole-fused aza-spirocyclic compounds has been reported, demonstrating an effective strategy to enhance the structural and functional diversity of these molecules. Moreover, the azaspirocyclic framework is well represented among bioactive compounds. For example, Annosqualine contains a spirocycloheptadienone motif associated with notable cytotoxic activity, while Atiprimod, an orally bioavailable azaspiro compound, has advanced into clinical evaluation for cancer therapy. These precedents underscore the pharmacological relevance of azaspirocyclic scaffolds and further support their exploration as promising immunomodulatory chemotypes [4]. Due to the intriguing structural features and biological properties, this study proposed to identify immunomodulatory effects of those aza-spirocyclic compounds. We screened 12 aza-spirocyclic compounds, representing novel scaffolds designed and synthesized (Supplementary Table 1), with the expectation of exhibiting immunomodulatory properties. These immunotherapeutic compounds may have significant opportunities for exploring the vast possibilities targeting the afore-mentioned pathways and discovering small molecules as immunotherapeutics and/or vaccine adjuvants for cancer. Several spirocyclic analogues have been investigated for anti-inflammatory activity, yet only a few have progressed to clinical evaluation for autoimmune or inflammatory diseases. Beyond their anti-inflammatory potential, spirocyclic scaffolds have also been associated with antioxidant activity, highlighting their utility in the development of drug candidates for metabolic, neurodegenerative, and oncological conditions, including diabetes, Alzheimer’s disease, and cancer [21].
Initially, the cytotoxicity of the synthesized compounds was assessed using an
MTT assay in RAW264.7 macrophages. None of the compounds exhibited cytotoxic
effects at concentrations exceeding 100 µM (Supplementary Table
2). To further evaluate their immunomodulatory potential, a series of assays,
including NO quantification and cytokine profiling, were performed using both
murine macrophages and murine splenocytes. No significant induction of NO and
IL-6 was observed following treatment of RAW264.7 macrophages with any of the
compounds. In contrast, a marked increase in IL-10 production was detected in
response to the AS1, AS2, AS3, AS4,
AS5, AS6, AS8, AS9, and AS10
compounds. Similar trends were observed in murine splenocytes, which likewise
showed no elevation or inhibition of pro-inflammatory cytokines (IL-6 and
TNF-
Moreover, the regulation of PD-L1 expression is known to be via
activation of the JAK/STAT pathway by cytokines released by immune cells. Also,
previously shown that spirocyclic salts depict inhibition of the immune and
eventually JAK2/STAT3 pathway [3, 22]. The specific JAK-STAT combinations
determine the cytokine signaling pathways that mediate various immunological
functions. It has been observed that JAK1 and JAK3 are critical for the signaling
pathways of several interleukins, including IL-2, IL-4, IL-7, IL-9, IL-15, and
IL-21 [3]. Conversely, JAK2 modulates IL-6 and gp130 cytokine signaling [3].
Additionally, in silico molecular docking revealed that four compounds;
AS1, AS2, AS8, and AS10; exhibited strong
binding affinity toward JAK proteins. To further validate these interactions,
gene and protein expression analyses were performed. While no significant changes
were observed at the mRNA level, AS8 and AS10 demonstrated
inhibition of JAK2 at the total protein level. Furthermore, to exclude potential
off-target effects, MAPK protein expression was evaluated and showed no
significant changes (Supplementary Fig. 3). In contrast, we observed
slightly elevated levels of IL-12p70, INF-
The present study has several limitations. While spirocyclic architectures are
progressively getting popular in drug discovery for their three-dimensional
shape, rigidity, and potential to improve pharmacological profiles, they possess
significant limitations related to synthetic complexity, structural stability,
and physicochemical properties. The immunological assessment was confined to a
limited set of cytokines (IL-6, TNF-
The newly synthesized aza-spirocyclic compounds showed excellent cytocompatibility and clear immunomodulatory potential. A subset of five compounds increased IL-10 while suppressing LPS-induced IL-6, indicating strong anti-inflammatory and immunosuppressive activity. Combined with their predicted affinity for JAK proteins, these findings suggest that aza-spirocyclic scaffolds may modulate JAK/STAT-related pathways. Among the series of compounds, AS8 and AS10 represent promising anti-inflammatory candidates, demonstrating favorable cytokine modulation and interaction with inflammatory signaling pathways. While selenium-containing derivatives showed notable activity, further structure-activity relationship studies are required to delineate the specific contribution of selenium. These compounds provide a useful framework for future optimization of immunomodulatory agents.
All data associated with this study are included in this publication. The synthesized molecules are available at the R.R.C. The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
SRB and RRC designed and supervised the study. RRC synthesized and characterized the compounds. AP performed the biology research and analyzed the data. RRT performed the molecular docking and immuno-informatics analysis. SRB and RRC performed the analysis and wrote the manuscript with the input from all the authors. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All animal experiments were conducted in accordance with the guidelines of the Institutional Animal Ethics Committee (IAEC), CSIR-IICT, Hyderabad, India, and were approved under protocol number IICT-IAEC-069-2025. This study adhered to the animal welfare guidelines of the Committee for Control and Supervision of Experiments on Animals (CCSEA), Government of India.
SRB would like to thank the director, CSIR-Indian Institute of Chemical Technology, Hyderabad, for providing infrastructure, lab space, and support for an institutional research grant (MLP9033). CSIR-IICT manuscript communication number: IICT/Pubs./2025/443. AP would like to thank Poornima Penta for guiding in ELISA experiment and Srilekha Paidighantam for helping during splenocyte harvesting. S.R.B would like to thank Anusandhan National Research Foundation (ANRF) for the award of Prime Minister Early Career Research Grant: ANRF/ECRG/2024/000505/LS. S.R.B. thanks Ignite Life Science Foundation (https://ignitelsf.in/) for the award of the grant: Acorn-AMR3/RNATech/2024/01. R.R.T acknowledges DST for providing the INSPIRE Faculty Fellowship and research grant (IFA-23-PH-308).
This project was supported by the CSIR, Grant No. CSPS24/RDSF/IICT/IHP24/01. AP was supported by the ICMR-JRF fellowship of the Department of Health Research, New Delhi, India (Award Number-3/1/3/JRF-2023/HRD - 046(145819)).
The authors declare no conflicts of interest. Given his role as the Guest Editor, Srinivasa Reddy Bonam had no involvement in the peer-review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process for this article was delegated to Changqing Su.
ChatGPT GO was used to correct grammar and improve the language of the manuscript. After using this tool, the authors reviewed and edited the content as needed and take full responsibility for the content of the publication.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBL48867.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.