


1 Department of Pediatrics, Taipei Veterans General Hospital, 11217 Taipei, Taiwan
2 School of Medicine, National Yang Ming Chiao Tung University, 112304 Taipei, Taiwan
Abstract
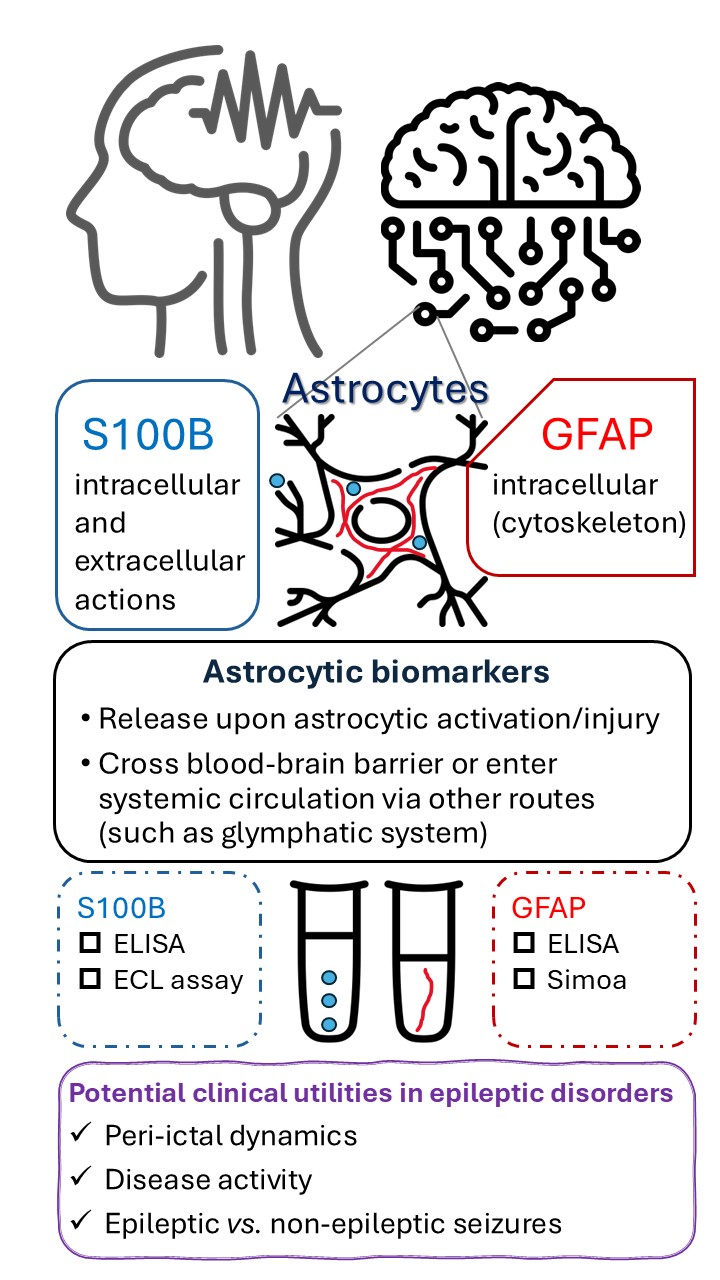
Epilepsy is one of the most common neurological disorders in both children and adults, characterized by significant clinical heterogeneity and dynamic natural course. The pathophysiological roles of astrocytes in epilepsy have been increasingly recognized. Fluid biomarkers derived from astrocytes are actively studied in epileptic disorders, although their use remains limited in clinical practice. This review aims to compile and analyze clinical and experimental findings concerning astrocytic biomarkers in epilepsy and related conditions, with a focus on glial fibrillary acidic protein (GFAP) and S100 calcium-binding protein B (S100B). Herein we examine their roles in assessing seizure burden and temporal dynamics, explore their potential in distinguishing epileptic from psychogenic non-epileptic seizures, and discuss their therapeutic, prognostic, and mechanistic implications in the context of epileptic disorders.
Graphical Abstract
Keywords
- astrocyte
- biomarker
- epilepsy
- glial fibrillary acidic protein
- neuroinflammation
- S100B
- seizure
Epilepsy is one of the most common neurological disorders globally, and the number of people with epilepsy is approximately fifty million worldwide. Epilepsy exhibits significant etiological and clinical heterogeneity, requiring a personalized approach to patient management. Various diagnostic tools are commonly utilized in both clinical practice and research, including electrophysiological studies, structural and functional neuroimaging, genetic testing, and neuropsychological evaluations. This multi-faceted approach has enhanced our understanding of epilepsy’s mechanisms and improved clinical management. However, the disease course varies widely among patients and can be highly dynamic within individuals, while the aforementioned tools are either insufficient or impractical for capturing fluctuations in disease activity over time. Fortunately, advances in biomarker research have led to the identification of several neuronal and glial markers as promising candidates to address this challenge.
Epilepsy is generally conceptualized as resulting from dysregulated neuronal excitability and/or synchrony, which in turn leads to abnormal paroxysmal clinical events. Nonetheless, the roles of other cell types in the central nervous system (CNS) have been increasingly recognized and prioritized in research [1, 2]. Astrocytes, one of the most abundant cell types in the CNS, have taken center stage in the study of epilepsy. Astrocytes can influence neuronal excitability through various mechanisms, including calcium signaling, potassium buffering, and gap junction coupling. Experimental findings suggest that the configuration and functional state of the astrocyte network can influence epileptiform activities [3, 4]. Moreover, diverse ways of astrocyte-specific stimulation were shown to induce excessive neuronal activities [5]. Taken together, these observations indicate that astrocytes may act as both modulators and initiators of epileptic activities, making astrocytes a potential cellular target for therapeutic interventions in epilepsy. As such, this review aims to collate and synthesize recent findings concerning fluid biomarkers that reflect astrocytic activation or injury in the context of epileptic disorders. Specifically, we examine their roles in assessing seizure burden and temporal dynamics, explore their potential in distinguishing epileptic from psychogenic non-epileptic seizures (PNES), and discuss their therapeutic, prognostic, and mechanistic implications in epileptic disorders.
Cerebrospinal fluid (CSF) and blood-based astrocytic biomarkers have been investigated for decades, with a particular focus on glial fibrillary acidic protein (GFAP) and S100 calcium-binding protein B (S100B). The key characteristics of these two biomarkers are listed in Table 1 (Ref. [6, 7, 8, 9, 10, 11]). Together with neuronal markers, these astrocytic markers have been found to be useful in a range of neurological conditions like traumatic brain injury or multiple sclerosis [12, 13, 14, 15, 16]. Although GFAP and S100B are seldom used clinically for epileptic disorders, they have been investigated in the context of epilepsy and related conditions, which is the main theme of this review. Other biomarkers presumably derived from astrocytes, such as chitinase 3–like protein 1 (CHI3L1) and SERPINA3, have been much less investigated in the context of epileptic disorders [17, 18]. In addition to being an acute phase glycoprotein during systemic inflammation, SERPINA3 was also considered a biomarker of reactive astrocytes. However, its cellular origins in the CNS remain controversial, and its role—whether pathogenic or neuroprotective—has yet to be fully elucidated [19]. Therefore, this biomarker will not be discussed further in this review.
| GFAP | S100B | |
| Entrez Gene ID | 2670 | 6285 |
| OMIM | *137780 | *176990 |
| Molecular weight | ~50 kDa | Dimer ~21 kDa (9–14 kDa per monomer) |
| Half-life (in blood) | 24–72 hours [6] | 25–120 minutes [7] |
| Cell or tissue sources other than astrocytes [6] | Renal tubular cells, enteric cells | Oligodendrocytes, Schwann cells, other peripheral tissues (such as melanocytes, skeletal muscles, and adipose tissue) |
| Physiological functions | The main intermediate filament in astrocytes, involved in structural support, motility, signal transduction and cell homeostasis [8] | A calcium-binding protein, involved in cell signaling, cell survival, proliferation, and differentiation [9] |
| Interacting partners | Vimentin; Desmin; HSP90 cochaperone | RAGE; MDM2; p53; and many others [10] |
| Solubility | Soluble in low ionic strength solutions [11] | High solubility (even in 100% sodium persulfate) [10] |
Abbreviations: OMIM, Online Mendelian Inheritance in Man; RAGE, receptor for advanced glycation end-products; GFAP, glial fibrillary acidic protein; S100B, S100 calcium-binding protein B.
S100B is a calcium-binding protein mainly expressed by glial cells, particularly astrocytes. It is also produced at nonneural cells and tissues, such as melanocytes and adipocytes (Table 1). S100B exerts both intracellular and extracellular actions, both of which have clinical relevance [20, 21]. S100B could be detected in CSF and systemic circulation, particularly in pathological conditions, using enzyme-linked immunosorbent assay (ELISA) [22, 23, 24] or electrochemiluminescence (ECL) assay [25, 26, 27, 28]. Some studies have measured serum and CSF S100B concurrently [29], and it has been shown that the S100B levels in CSF and blood were correlated in patients with traumatic brain injury [26]. However, it should be noted that S100B has extracerebral sources, and an increase in serum S100B can occur without concomitant increase in CSF S100B [10]. Serum S100B levels may vary with age [29] and should be considered when interpreting data.
Serum S100B level could be elevated during interictal period in some but not all patients with epilepsy [22, 30]. Calik et al. [28] reported that children with intractable focal epilepsy had significantly higher interictal serum S100B than healthy controls at group level. Similarly, Maiti et al. [31] reported that adults with focal epilepsy had significantly higher serum S100B than healthy controls. Portela et al. [30] reported that significantly higher serum S100B levels were observed in patients with bilateral electroencephalographic (EEG) findings than in patients with unilateral or normal EEG findings. Aksoy et al. [32] recently reported that serum S100B levels were significantly higher in the refractory pediatric epilepsy group (34.13 pg/mL) than in both the control group (28.05 pg/mL) and the nonrefractory pediatric epilepsy group (28.05 pg/mL). In a study involving 174 subjects including 82 patients with status epilepticus (SE), Hanin et al. [33] reported that 84% of patients with serum S100B above 90 pg/mL presented with a SE, and 90% of patients without SE had serum S100B lower than 90 pg/mL. Serum S100B performed better at detecting SE (area under the ROC curve (AUC) 0.748) compared with serum neuron-specific enolase (NSE) (AUC 0.624) in that study. Taken together, the aforementioned observations suggest that serum S100B could be a potential marker of seizure severity. In addition, Hanin et al. [33] found no difference in serum S100B levels with regard to SE etiology. In contrast, Wang et al. [34] reported that serum S100B in children with convulsive SE varied with etiologies, with those with viral encephalitis having significantly higher levels compared to those with primary generalized epilepsies or complex febrile seizures.
Several studies focused on serum and/or CSF S100B in febrile seizures [23, 24, 29], while some studies included children with febrile seizures as a comparison group [34, 35, 36, 37, 38]. Febrile seizures do not appear to elevate serum S100B levels beyond those associated with febrile illnesses alone [39]. Serum S100B was not predictive of the risk of recurrent febrile seizures [29]. These findings may provide reassurance about the relatively benign nature of febrile seizures. Benign convulsions with mild gastroenteritis, another common cause of seizure in young children, have been associated with a higher serum NSE but not S100B compared to febrile seizures [36]. By contrast, some infection-triggered encephalopathy syndromes characterized by the presence of fever and seizures, such as acute encephalopathy with biphasic seizures and late reduced diffusion (AESD), were associated with significantly higher CSF S100B levels [37]. An earlier study reported that CSF S100B at a cutoff value of 100 pg/mL may help distinguish AESD from febrile seizures [38].
GFAP is an intermediate filament protein predominantly expressed in astrocytes. The structural role of GFAP in astrocytes is analogous to that of neurofilaments in neurons, and the utility of these intermediate filament proteins as fluid biomarkers in various neurological disorders is under active investigation in recent years [13, 16].
GFAP could be quantified in diverse ways. Traditionally, ELISA has been used to
measure their serum levels [32, 40]. Single-molecule array (Simoa), a relatively
recent technology that enables ultrasensitive detection, has become increasingly
used [27, 41]. Biosensing platforms, such as graphene field-effect transistor,
are being developed [42, 43], holding promise for future clinical applications.
On the other hand, GFAP has multiple isoforms and various cleavage products,
among which GFAP-
Some studies have measured serum and CSF GFAP concurrently, and it has been shown that the GFAP levels in CSF and blood were correlated in patients with neuroinflammatory disease or other conditions [46, 47]. It is notable that CSF GFAP appears to be less stable after freeze-thaw cycles as compared to serum GFAP [48]. Plasma GFAP may vary with age [41], which should be taken into account in data interpretation.
GFAP level in CSF obtained within 24 hours of seizure was found to be elevated
(
In a recent study reported by Aksoy et al. [32], the GFAP levels in the
pediatric epilepsy groups were significantly higher than those in the control
group (17.9 ng/mL), while no significant difference was found between children
with refractory (25.4 ng/mL) and nonrefractory (26.1 ng/mL) epilepsies. It is
notable that the levels of ubiquitin carboxyl-terminal hydrolase isozyme L1
(UCHL1), a neuronal marker, were not different between groups in that study. In
another study focusing on childhood epilepsy, both serum GFAP and UCHL1 were
significantly higher in those with epilepsy than in controls (GFAP mean
The differentiation between epilepsy and non-epileptic paroxysmal disorders could be clinically challenging at times. The performance of currently available biomarkers (such as prolactin) is less satisfactory in this regard [52, 53], prompting searches for better surrogate markers. Simani et al. [54] reported that serum GFAP was significantly higher following epileptic seizures compared to PNES (Table 2, Ref. [51, 54, 55, 56]). Receiver operating characteristic (ROC) analysis indicated that serum GFAP may be modestly useful in distinguishing epileptic seizures from PNES (AUC = 0.68), with an optimal cutoff value of 2.71 ng/mL (sensitivity 72%, specificity 59%). Similarly, in a larger study involving 104 epilepsy and 22 PNES cases, Dobson et al. [51] found that both plasma GFAP and NfL levels were elevated in patients with epilepsy compared to PNES. Asadollahi and Simani [55] reported that serum S100B was significantly higher in patients with epileptic seizures than PNES, although it is intriguing to note that serum S100B was also significantly higher in PNES patients compared to healthy controls. On the other hand, no significant difference in serum GFAP levels between patients with PNES and healthy controls was noted in another study [54]. It is noteworthy that not all studies found differences in astrocytic markers between epileptic seizures and PNES. For instance, Nass et al. [56] found that serum and CSF GFAP levels were not significantly different between patients with autoimmune-mediated epilepsy, genetic generalized epilepsy, and PNES.
| PNES | Epileptic seizures | ||||||
| Specimen, methodology | n | Mean or median | n | Mean or median | Diagnostic performance | ||
| GFAP (pg/mL) | |||||||
| Simani et al. 2018 [54] | Serum, ELISA | 20 | 2840 |
43 | 3690 |
AUC 0.68 (95% CI 0.56–0.79) | |
| Nass et al. 2021 [56] | Serum, Simoa | 8 | 95.85 | GGE 7 | GGE 71.6 | NA | |
| HS 13 | HS 60 | ||||||
| AIE 13 | AIE 92.5 | ||||||
| CSF, Simoa | 5660 | GGE 5720 | |||||
| HS 7000 | |||||||
| AIE 5880 | |||||||
| Dobson et al. 2024 [51] | Plasma, Simoa | 22 | 61.4 | 104 | 102 | AUC 0.63 | |
| S100B (ng/mL) | |||||||
| Asadollahi et al. [55] | Serum, ELISA | 20 | 23.73 |
43 | 31.58 |
AUC 0.59 | |
Abbreviations: AIE, autoimmune-mediated epilepsy; AUC, area under the ROC curve; ELISA, enzyme-linked immunosorbent assay; GGE, genetic generalized epilepsy; HS, hippocampal sclerosis; NA, not available; PNES, psychogenic non-epileptic seizure; Simoa, single-molecule array.
Blood S100B and GFAP exhibit different temporal dynamics along peri-ictal course, which could be appreciated from a study of pediatric convulsive SE in which several biomarkers were measured at the first, fourth, and tenth day of convulsive SE [34]. Peak serum S100B was noted on the first day, whereas peak serum GFAP was observed on the fourth day after SE. The half-life of S100B in vivo is significantly shorter than that of GFAP (Table 1), which may contribute to the observed differences in their temporal dynamics. Blood levels of S100B and GFAP could rise quickly after a seizure [50, 57]. Calik et al. [57] reported that serum S100B was elevated within 30 minutes after a complex partial seizure in children with temporal lobe epilepsy. Nass et al. [50] reported that serum GFAP level doubled in 15% of adult patients immediately after a tonic-clonic seizure, and it was more sensitive than NfL in terms of biomarker performance. Moreover, postictal changes of GFAP were correlated with leukocytosis in that study, suggesting a link between astrocytic activation and peripheral immune cell mobilization.
S100B and GFAP in peripheral blood may return to baseline a few hours after a seizure [50] or remain elevated for up to 24 hours [58]. Since these astrocytic biomarkers demonstrate dynamic changes along different phases of the peri-ictal period, they could serve as potential state markers in the context of epileptic disorders, although longitudinal sampling may be needed for data interpretation. Given its shorter half-life, S100B may provide better temporal resolution for studying peri-ictal dynamics. However, the clinical utility of a biomarker could be limited if its blood levels fluctuate rapidly, making the optimal time window for specimen collection too narrow.
Because astrocytic biomarkers originate in the CNS, their detection in blood may represent a delayed rather than real-time reflection of disease process. Indeed, changes in serum S100B may lag behind that in CSF [26]. It has been reasoned that the blood-brain barrier (BBB) permeability may influence the detection of S100B in the systemic circulation. In line with this notion, serum S100B may serve as a potential biomarker of BBB integrity [58]. In clinical practice, BBB integrity is often assessed by contrast-enhanced MRI or the CSF/serum albumin ratio (also known as albumin quotient, QA). QA could be elevated in patients with SE or recurrent generalized tonic-clonic seizures [59, 60]. Significant positive correlations between serum S100B and QA were observed in patients with traumatic brain injury or headache [26, 61]. Specificity for abnormal QA was 90% or higher when the cutoff of serum S100B level was set at 27 pg/mL in an earlier study [61]. However, this is less studied in patients with epilepsy. Moreover, whether S100B could be regarded as a biomarker of BBB disruption is still under debate, as some evidence suggests that it may exit the CNS and enter systemic circulation through alternative routes, such as the glymphatic system [10, 25, 62].
Like S100B, the drainage of GFAP from the CNS into blood remains incompletely understood. Multiple pathways, including those involving the arachnoid villi, the glymphatic system, and fluid exchange at the BBB, are likely to coexist [6, 62]. More research is needed to decipher the quantitative relationships between serum levels of these biomarkers and BBB permeability and/or other factors [8].
Findings derived from clinical studies investigating astrocytic markers to date have been largely correlational in nature. The causal relationships between astrocytic marker alterations and other clinical or paraclinical features remain to be elucidated. Understanding the pathobiology of S100B in epileptic disorders is crucial in terms of treatment considerations. Potential therapeutics targeting S100B include arundic acid (ONO-2506), which inhibits S100B synthesis, and nanobodies that modulate the interaction of S100B and RAGE (receptor for advanced glycation end-products) [63]. Arundic acid has been demonstrated to be neuroprotective in several rodent models of seizures, including those induced by hypoxia [64], Li-pilocarpine [65], and genetic lesion [66], respectively. Whether these findings are translatable to epileptic disorders in humans requires further research.
The detection of elevated serum S100B during interictal periods in some patients with epilepsy may indicate ongoing or persistent astrocytic activation, even in the absence of clinical seizures [22]. This could spark interest in exploring whether antiseizure medications affect S100B release. A randomized, open-label trial involving 60 adults with focal epilepsy found that treatment with carbamazepine, but not oxcarbazepine, was associated with significant reduction in serum S100B, although both drugs led to significant improvement in seizure severity [31]. Maiti et al. [31] reasoned that the differential drug effects on serum S100B might be attributed to the modulatory actions of carbamazepine on the signaling in astrocytes. Further research is needed to elucidate the effects of various antiseizure medications on astrocyte biology and related biomarkers.
Emerging evidence shows that S100B from activated or injured astrocytes may act as damage-associated molecular patterns (DAMPs), thereby exacerbating neuroinflammatory responses. This provides rationale for S100B-targeted therapeutics mentioned above [65]. On the other hand, S100B is a zinc-binding protein, and Hagmeyer et al. [67] reported that this property may confer neuroprotection through abolishing excitotoxicity. It is interesting to note that a delayed increase of extracellular S100B (peaking at 24 hours) after kainate-induced excitotoxity was observed in an in vitro rat model (organotypic spinal cord slices) [68]. Kainate treatment induced 40% neuronal death without change in glial cell number in that study, and experimental evidence suggested that the extracellular S100B was released from a readily releasable pool. They also found that the S100B elevation was glutamate receptor-dependent, implicating S100B release as an endogenous protective mechanism against excitotoxicity in the CNS.
GFAP is also involved in the pathophysiology of epilepsy. Protein-protein interaction network analysis identified GFAP, AQP4, and several other astrocyte-specific genes as hub genes in human temporal lobe epilepsy; these genes were upregulated both in human datasets and in the epileptic rat hippocampus [69]. Alexander disease, a genetic leukodystrophy caused by gain-of-function mutations in GFAP, may present with seizures. Gfap-targeted antisense oligonucleotide has been tested successfully in a rat model of Alexander disease, although seizures were not specifically examined in that study [70]. The pathophysiological roles of GFAP in ictogenesis and epileptogenesis deserve further investigations.
Several studies have investigated the predictive potential of glial markers in the context of epileptic disorders. Although serum S100B performed better at detecting SE (AUC 0.748) as compared with NSE (AUC 0.624) [33], it was less useful in predicting the risk of seizure recurrence following refractory SE (AUC 0.592) [71]. Similarly, another study reported limited prognostic value of S100B in predicting disability at 30 days in adults with SE [72]. In adult patients visiting the emergency department following a seizure, plasma S100B also appeared to be a poor predictor (AUC 0.57) of adverse outcomes (such as seizure recurrence, hospitalization, or mortality) in subsequent four weeks [73].
In a prospective observational study of adults with new-onset seizures, serum S100B was higher in patients who were subsequently diagnosed with epilepsy than those with single seizure [27]. In contrast, no significant difference was noted in age-normalized plasma GFAP between patients with single seizure and those who developed epilepsy. These findings suggest that S100B might be a candidate biomarker for identifying individuals at higher risk of developing epilepsy following a first seizure. It is intriguing, however, to note that in patients with acute stroke, a lower blood S100B level was an independent risk factor for subsequent epilepsy (hazard ratio 2.955, 95% CI 1.534–5.491). Further research is needed to clarify the role of S100B in epileptogenesis.
Astrocytic biomarkers could be valuable in assessing the seizure burden and temporal dynamics of epileptic disorders, and aid in the differentiation between epileptic and psychogenic non-epileptic seizures. Investigations involving these biomarkers clearly indicate the significant roles of astrocytes in epilepsy. In view of the clinical and experimental findings reviewed above, further research into astrocyte pathophysiology, associated biomarkers, and their therapeutic implications is warranted in future epilepsy research.
AESD, acute encephalopathy with biphasic seizures and late reduced diffusion; AIE, autoimmune-mediated epilepsy; AUC, area under the ROC curve; BBB, blood-brain barrier; CHI3L1, chitinase 3–like protein 1; CNS, central nervous system; CSF, cerebrospinal fluid; DAMPs, damage-associated molecular patterns; ECL, electrochemiluminescence; ELISA, enzyme-linked immunosorbent assay; GFAP, glial fibrillary acidic protein; NfL, neurofilament light; NSE, neuron-specific enolase; OMIM, Online Mendelian Inheritance in Man; PNES, psychogenic non-epileptic seizure; QA, CSF/serum albumin ratio (albumin quotient); RAGE, receptor for advanced glycation end-products; ROC, receiver operating characteristic; S100B, S100 calcium-binding protein B; SE, status epilepticus; Simoa, single-molecule array; UCHL1, ubiquitin carboxyl-terminal hydrolase isozyme L1.
WSL designed the research. TRH and PHC performed literature research and analyzed the data. TRH and WSL wrote the manuscript. All authors critically reviewed and revised the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This work was partly supported by grants from the Taipei Veterans General Hospital (V114C-171). The Taipei Veterans General Hospital had no role in each of the following: design and conduct of the study; collection, management, analysis and interpretation of the data; preparation, review or approval of the manuscript; and decision to submit the manuscript for publication.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.