



 , Chaoyi Wang 1,2,†, Yundong Xu 1, Bingchen Li 3, Jianzhou Ye 4,*
, Chaoyi Wang 1,2,†, Yundong Xu 1, Bingchen Li 3, Jianzhou Ye 4,*
1 The First School of Clinical Medicine, Yunnan University of Chinese Medicine, 650011 Kunming, Yunnan, China
2 Now with Gynecology Department, Kunming Traditional Chinese Medicine Hospital, 650000 Kunming, Yunnan, China
3 School of Public Health, Southeast University, 211189 Nanjing, Jiangsu, China
4 Department of Dermatology, The First Affiliated Hospital of Yunnan University of Traditional Chinese Medicine, 650032 Kunming, Yunnan, China
†These authors contributed equally.
Abstract
The C-X-C chemokine receptors (CXCR) chemokine receptor family, consisting of seven primary members (CXCR1–CXCR7), is crucial in regulating immune cell recruitment, angiogenesis, cellular proliferation, and maintaining skin homeostasis and immune functions. This review evaluates the expression and roles of CXCR receptors across a range of skin cells, including keratinocytes, fibroblasts, endothelial cells, melanocytes, and various immune cells such as T cells, dendritic cells, and macrophages. Aberrations in CXCR signaling have been associated with a variety of skin disorders, such as psoriasis, atopic dermatitis, acne, and skin cancers. Despite significant advancements in the field, several critical questions persist. These include the differential effects of CXCR signaling in distinct skin pathologies and the intricate interactions between CXCR receptors and their ligands within diverse skin microenvironments. Moreover, the therapeutic targeting of the CXCR family remains unresolved, necessitating further research into its long-term efficacy and possible adverse effects. Future investigations should prioritize these critical issues to develop more effective therapeutic strategies for managing skin diseases, ultimately improving patient outcomes.
Keywords
- CXCR receptors
- skin diseases
- immune response
- inflammation
- psoriasis
- atopic dermatitis
- therapeutic targets
The C-X-C chemokine receptors (CXCR) chemokine receptor family, consisting of seven primary members (CXCR1–CXCR7), represents a critical subset of chemokine receptors, essential for immune cell recruitment, angiogenesis, and cellular proliferation [1]. As part of the G protein-coupled receptor superfamily (GPCR), these receptors are characterized by seven transmembrane domains, an extracellular N-terminal, and an intracellular C-terminal, facilitating diverse intracellular signaling cascades. Predominantly expressed on various immune cells, CXCR receptors engage with CXC chemokines to regulate inflammatory responses and immune cell function.
The distinct roles of each CXCR member in health and disease have been extensively documented. For instance, CXCR1 and CXCR2, which share approximately 76% sequence homology, are primarily expressed on neutrophils, where they activate G-proteins and signaling pathways, including phosphoinositide 3-kinase (PI3K) and mitogen-activated protein kinase (MAPK), to enhance chemotaxis and reactive oxygen species production [2]. CXCR3, with its isoforms CXCR3-A, CXCR3-B, and CXCR3-alt, orchestrates immune cell migration and modulates tumor cell behavior via interaction with ligands C-X-C motif ligand (CXCL)9, CXCL10, and CXCL11 [3]. CXCR4, in conjunction with its unique ligand CXCL12, plays a pivotal role in hematopoiesis, angiogenesis, and tumor progression, mediated through multiple signaling pathways [4]. Other family members, including CXCR5, CXCR6, and CXCR7, contribute to immune regulation, cellular migration, and angiogenesis, influencing a range of inflammatory and autoimmune conditions [5, 6, 7].
Skin diseases, which include inflammatory conditions like psoriasis and atopic dermatitis (AD), chronic conditions such as acne, and various skin cancers, often involve complex interactions between immune cells, skin cells, and environmental factors. These interactions lead to symptoms such as redness, itching, and lesions. Unraveling the underlying mechanisms driving these diseases is essential for the development of effective treatments.
In the context of dermatological health, CXCR receptors are fundamental for maintaining skin homeostasis and mediating responses to pathogens and injury. Skin homeostasis, encompassing barrier function and tissue integrity, is maintained through coordinated mechanisms like cell proliferation, apoptosis, differentiation, and immune surveillance. The regulation of keratinocytes, fibroblasts, and immune cells by CXCR signaling pathways is critical to this balance. While CXCR receptors facilitate pathogen clearance and tissue repair by modulating immune cell dynamics, such as T cells and macrophages, dysregulation of these pathways can disrupt homeostasis, contributing to chronic inflammation and other skin disorders [8, 9]. Elucidating the precise roles of CXCR receptors in these conditions not only enhances our understanding of disease mechanisms but also provides a basis for the development of targeted therapeutic strategies.
This review aims to offer a comprehensive overview of the CXCR family, focusing on their expression and functions in various skin cells and their involvement in skin diseases. By elucidating the complex roles of CXCR receptors in skin pathology, this review aims to highlight potential therapeutic targets and inform strategies for managing skin disorders, thereby improving patient outcomes.
The CXCR chemokine receptor family is a prominent subset of chemokine receptors, consisting of seven primary members (CXCR1–CXCR7). These receptors are expressed on various immune cells and mediate their functions through interactions with CXCL chemokines, driving key biological processes such as inflammatory cell recruitment, angiogenesis, and cellular proliferation (Fig. 1) [10].
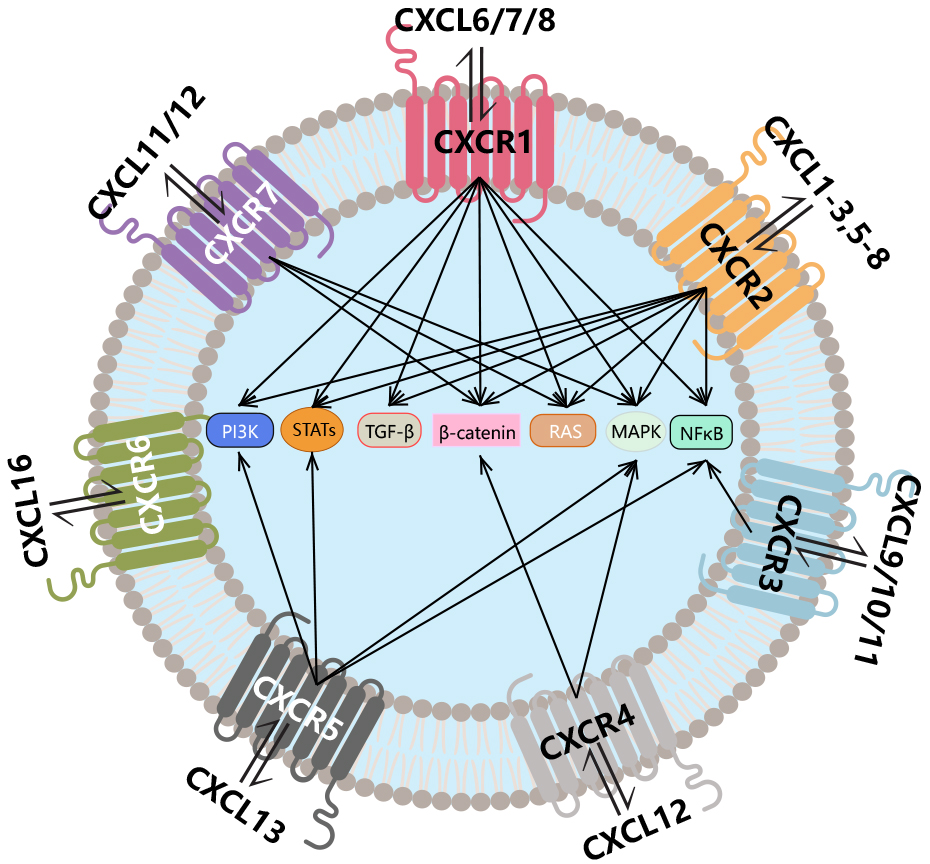 Fig. 1.
Fig. 1.
Introduction to the CXCR family. The CXCR family consists of
seven members (CXCR1–7) that function by binding to CXCL chemokines, triggering
the activation of downstream signaling pathways. CXCR, C-X-C chemokine receptors;
CXCL, C-X-C motif ligand; PI3K, phosphoinositide 3-kinase; STAT, signal
transducers and activators of transcription; TGF-
CXCR1 and CXCR2, located on chromosome 2q35 with approximately 76% sequence homology, span 3.5 kilobases (kb) and 3.3 kb, respectively. Both receptors interact with ELR-positive C-X-C motif chemokines, including CXCL1–3 and CXCL5–8, initiating downstream signaling through G protein-coupled cascades. These pathways include the activation of PI3K-AKT and MAPK pathways (extracellular signal-regulated kinase (ERK)1/2, p38, and JNK), which promote cellular chemotaxis, adhesion, reactive oxygen species (ROS) generation, and migration. For example, CXCL8 recruits neutrophils to tumor lesions through CXCR1/2, subsequently triggering the formation of neutrophil extracellular traps (NETs) [2]. In atherosclerotic plaques, CXCR2-expressing monocyte-macrophages are recruited by macrophage migration inhibitory factor (MIF) and contribute to plaque development [11]. Additionally, CXCR1/2 can promote endothelial cell permeability by transactivating vascular endothelial growth factor receptor 2 (VEGFR2) [12].
CXCR3, located on chromosome Xq13.1 and spanning approximately 8.3 kb, exists in three splice variants: CXCR3-A, CXCR3-B, and the truncated CXCR3-alt. These isoforms interact with ligands CXCL9, CXCL10, and CXCL11. CXCR3-A is predominantly expressed on lymphocytes, where it orchestrates immune cell migration and maturation through its binding to CXCL9–11. The CXCR3-A isoform has been closely linked to tumor cell behavior, promoting epithelial-mesenchymal transition (EMT), cell migration, and invasion across various cancers [3]. CXCR3-B, primarily found on endothelial cells, significantly inhibits endothelial cell DNA synthesis and induces apoptosis, thereby exerting anti-angiogenic effects. This isoform has been associated with favorable prognoses in certain cancers due to its inhibitory role in angiogenesis [13]. CXCR3-alt, which binds exclusively to CXCL11, is believed to cooperate with interferon-inducible T-cell alpha chemoattractant (I-TAC) in mediating immune responses [14].
CXCR4, a member of the GPCR family located on chromosome 2q21 and spanning
approximately 13 kb, is ubiquitously expressed across various cell types. The
CXCR4/CXCL12 axis is critical for hematopoiesis, angiogenesis, and tumor
progression, operating through several signaling pathways, including rat sarcoma
virus oncogene homolog (RAS)-MAPK-MEK1/2, ERK1/2, and nuclear factor kappa B
(NF-
CXCR5, expressed on B cells, follicular helper T cells (Tfh), and
antigen-presenting dendritic cells (DCs), is another GPCR member [5] located on
chromosome 11q23.3, with a gene span of approximately 11 kb. The interaction
between CXCL13 and CXCR5 plays a pivotal role in B cell maturation and
differentiation, aiding the production of natural antibodies [5]. CXCL13 binding
to CXCR5 triggers the conversion of Guanosine diphosphate (GDP) to Guanosine
triphosphate (GTP), dissociating the trimeric complex into G
CXCR6, also known as CD186, Bonzo, STRL, and TYMSTR, interacts with CXCL16 to
activate the PI3K-Akt/PKB pathway, NF-
CXCR7, or ACKR3 (atypical chemokine receptor 3), is implicated in regulating cell migration and angiogenesis. Positioned on chromosome 2q37.3 with a gene span of approximately 4.6 kb, CXCR7 binds to ligands such as macrophage MIF, CXCL11 (I-TAC), and CXCL12 (SDF-1) [7]. Like CXCR4, CXCR7 operates through GPCR signaling pathways, including MAPK and RAS [7].
The skin constitutes the body’s primary defense against external threats, preventing pathogen invasion and safeguarding underlying tissues. It consists of key structural cells—keratinocytes, fibroblasts, endothelial cells, and melanocytes—along with a variety of immune cells, including T cells, dendritic cells, macrophages, and mast cells, collectively forming the skin’s immune system to counteract microbial attacks (Fig. 2). Keratinocytes, as the cornerstone of the physical barrier, facilitate immune cell recruitment through the secretion of chemokines. Fibroblasts play an essential role in wound healing by regulating the extracellular matrix, while endothelial cells are integral to the maintenance of skin microcirculation. Melanocytes contribute to ultraviolet (UV) protection by producing melanin. Immune cells directly recognize and eliminate pathogens, acting as critical defenders. The dynamic interactions among these cells are essential for maintaining skin integrity, ensuring tissue homeostasis, and fortifying the immune barrier. These functions hold significant pathophysiological relevance and provide a foundation for advancing research into the mechanisms underlying various skin disorders.
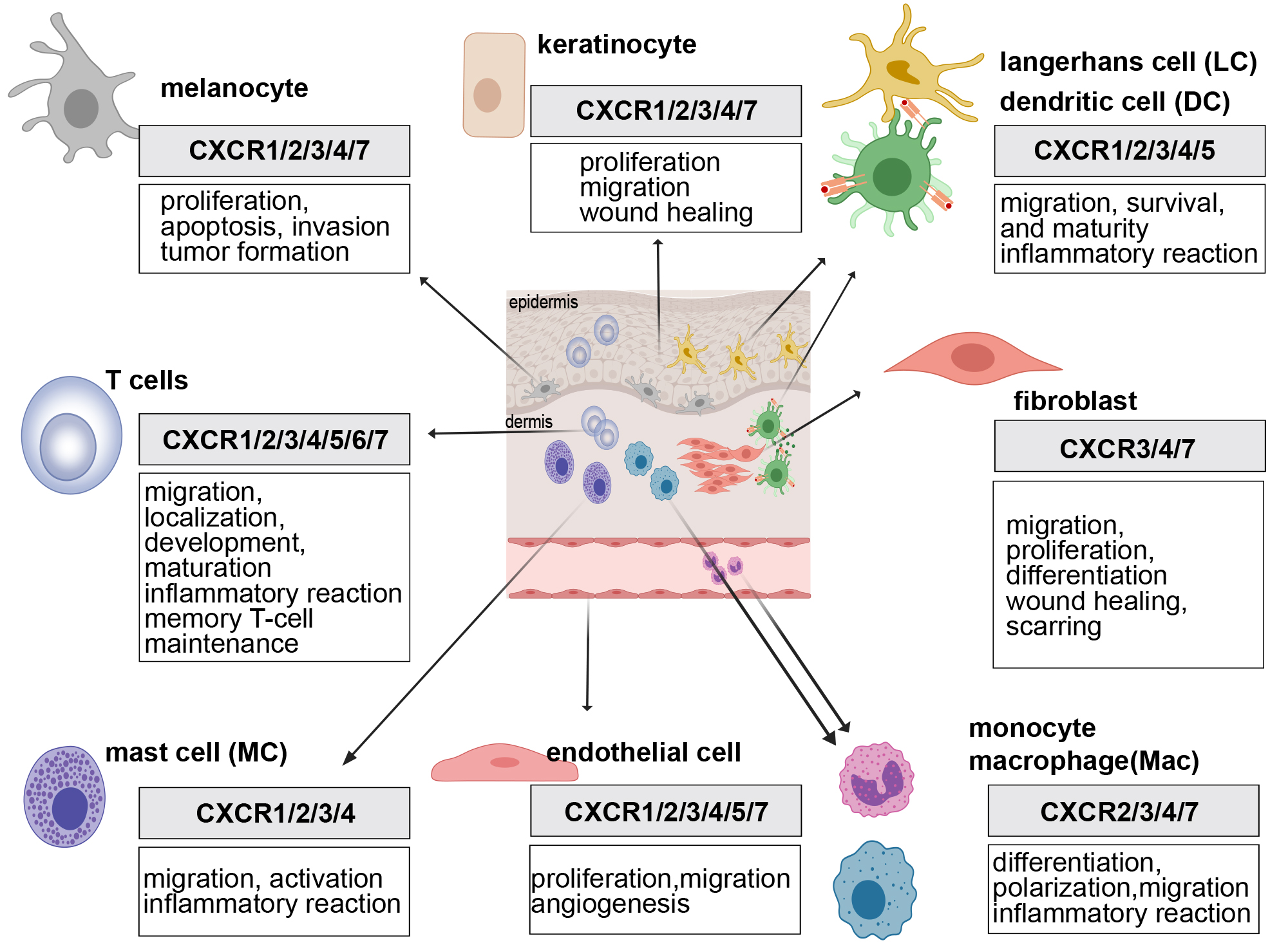 Fig. 2.
Fig. 2.
Roles of the CXCR family in various skin cells. The expression of CXCR family members in different skin cells facilitates key processes such as inflammatory cell recruitment, angiogenesis, and cellular proliferation. Created with MedPeer (https://image.medpeer.cn/show/index/home).
The expression of CXCR1, CXCR2, CXCR3, CXCR4, and CXCR7 in keratinocytes has been well documented. Both resting and activated keratinocytes express CXCR1 [17], while CXCR2 is primarily upregulated under pathological conditions [18, 19]. CXCR2 plays a key role in skin healing, being expressed in the epidermis of human skin explant cultures and regulated by inflammatory mediators such as interleukin (IL)-13 [20]. CXCR3 signaling promotes keratinocyte migration, with CXCR3-deficient mice exhibiting pronounced scar proliferation [21]. The CXCR4/CXCL12 axis is essential for the migration of oral keratinocytes during oral ulcer repair [22]. CXCR7, predominantly expressed in the basal layer, participates in wound healing through binding with its ligand CXCL12/SDF-1 [23].
CXCR4 expression in fibroblasts is critical for tissue injury response and repair [24]. Overexpression of CXCR4 has been associated with pathological conditions such as tumor progression and fibrosis-related diseases [25, 26]. CXCR7, which shares functional similarities with CXCR4, regulates the CXCL12/CXCR4 axis in fibroblasts. Recent study show that CXCR7 (ACKR3) is selectively expressed on senescent fibroblasts, not proliferating cells, suggesting that targeting CXCR7 may selectively eliminate senescent fibroblasts [27]. In fibroblasts, CXCR3 expression mediates immune regulation, with recent evidence showing that CXCR3 expression in localized scleroderma fibroblasts drives inflammatory fibrosis [28]. Additionally, CXCL10-mediated CXCR3 signaling in cervical cancer cells promotes fibroblast Programmed Death-Ligand 1 (PD-L1) expression, influencing tumor immunity [29].
In endothelial cells, CXCR receptors primarily regulate cell proliferation and angiogenesis. CXCR1 and CXCR2 mediate endothelial cell migration, with their downregulation leading to reduced motility and invasiveness [30]. Expression of CXCR1 and CXCR2 in endothelial cells is linked to cytoskeletal reorganization and capillary-like structure formation, essential for tissue repair and neovascularization [30]. CXCR3 is predominantly expressed in medium to large vessels, with lower expression in microvessels [31], where it plays a central role in vascular remodeling and immune cell recruitment [32]. CXCR4, one of the most extensively expressed CXCR receptors in endothelial cells, mediates angiogenesis, inflammation, vascular repair, and disease progression through its interaction with CXCL12. Research on CXCR5 in endothelial cells primarily focuses on lymphatic vessels, where it affects the structure and function of lymphoid tissues, particularly in B and T cell interactions [33]. CXCR5 expression promotes endothelial cell localization in lymph nodes, facilitating immune cell interactions [34]. CXCR7 activation in endothelial cells is critical for vascular formation, repair, and maintenance, working synergistically with CXCR4 [35].
In melanoma, CXCR1 is expressed across all stages and grades, while CXCR2 expression is confined to more aggressive forms, influencing tumor growth and invasiveness [36]. CXCR3 is involved in apoptosis, tumor metastasis, and immune responses, with CXCL10-induced CXCR3 activation promoting apoptosis in human melanocytes [37]. CXCR3 expression enhances melanoma invasiveness, promoting metastasis from the primary site and facilitating endothelial invasion [38]. Increased CXCR3 expression in senescent melanocytes suggests its role in autocrine signaling, influencing cellular aging and apoptosis [39]. Research on CXCR4 in melanoma has positioned it as a potential therapeutic target, with its activation enhancing migration, invasion, and proliferation of melanoma cells [40]. The role of CXCR7 (ACKR3) in melanocytes remains less explored, but its interaction with CXCL12 may regulate melanoma cell migration, invasion, and metastasis.
T cells express a range of CXCR that govern their migration, localization, and involvement in immune responses. Although T cells do not typically express CXCR1 and CXCR2, alterations in local cytokine profiles within autoimmune diseases and tumor microenvironments induce the upregulation of these receptors. Specifically, CXCR1 and CXCR2 significantly enhance T cell migration and persistence, thereby contributing to ongoing tumor immunity [41]. CXCR3 is predominantly expressed on Th1-type CD4+ T cells and effector CD8+ T cells, facilitating T cell migration to peripheral tissues and lymphoid regions, and promoting interactions with antigen-presenting cells [42]. CXCR4 is essential for T cell migration, localization, and development, playing a critical role in T cell differentiation within the thymus and their subsequent migration to lymphoid tissues, sites of inflammation, and other tissues [43]. Additionally, CXCR4 acts as a co-receptor for HIV, enhancing T cell susceptibility to infection and contributing to immunodeficiency [43]. CXCR5 is primarily expressed on Tfh, guiding their migration and localization within the follicular regions of lymph nodes, which is essential for B cell activation and antibody production [44]. CXCR5 may also be involved in the maintenance and functional integrity of CD4+ memory T cells, particularly during long-term immune responses [45]. CXCR6 directs T cells, especially CD8+ T cells, to sites of inflammation and infection, thus playing a pivotal role in immune responses [46]. Recent studies suggest that CXCR6 promotes CD8+ T cell survival and their transition to long-term residency [47, 48]. In Alzheimer’s research, CXCR6 knockout mice exhibit a reduced population of tissue-resident T cells [49]. CXCR7 primarily functions in concert with CXCR4, regulating T cell development, maturation, migration, and localization. In the tumor microenvironment, the interaction between CXCR7 and CXCR4 mediates T cell polarization toward regulatory T cells (Tregs), influencing T cell infiltration, function, and ultimately, tumor growth and immune evasion [50].
Dendritic cells are key elements of the skin’s immune system, responsible for capturing, processing, and presenting antigens, thereby initiating and regulating immune responses. Within the epidermis, dendritic cells predominantly exist as Langerhans cells, dispersed throughout the tissue. CXCR1 plays a significant role in dendritic cell-mediated inflammation, with CXCR1-expressing DCs being implicated in the induction of experimental autoimmune encephalomyelitis (EAE) and acute respiratory distress syndrome (ARDS) [51]. Isolated dendritic cells may also express CXCR2, potentially contributing to their migratory behavior [52]. CXCR3 is highly expressed on dendritic cells within lymphoid tissues, particularly adjacent to germinal centers and paracortical areas, which are rich in B cells, suggesting a possible role for CXCR3 in these B cell-rich regions [31]. During inflammation and immune surveillance, CXCR3 mediates dendritic cell migration and localization. In the tumor microenvironment, CXCR3-mediated recruitment of dendritic cells has been shown to enhance the efficacy of PD-L1 inhibitors [53]. CXCR4 promotes the trafficking of dendritic cells into lymph nodes [54]. CXCR5 facilitates the interaction between dendritic cells and B cells, chemotactically guiding dendritic cells to B cell areas in lymph nodes, thereby fostering effective immune interactions [55]. Furthermore, CXCR5+ dendritic cells contribute to the regulation of T Helper 2 (TH2) cell differentiation, which is pivotal for initiating and modulating immune responses [56]. While direct evidence of dendritic cells expressing CXCR6 remains limited, study indicates that dendritic cells can produce CXCL16, attracting CXCR6+ CD8+ T cells, thereby promoting CD8+ T cell survival and mediating tumor immunity through IL-15 production [57]. The direct expression of CXCR7 on dendritic cells remains largely unexplored; however, given the complexity and significance of chemokine receptors in immune modulation, further investigation is essential to elucidate the expression and functional roles of CXCR7 and other CXCR receptors on dendritic cells.
Macrophages are instrumental in pathogen clearance, necrotic tissue removal, and
inflammatory factor release, functioning as antigen-presenting cells that
interact with various immune cell types. CXCR2 serves as a pivotal target in
inflammation, as evidenced by CXCR2 knockout mice, which exhibit exaggerated
acute inflammatory responses and notable macrophage proliferation [58]. CXCR3
regulates tissue repair by modulating macrophage expression of Arg1, VEGFa, and
TNF-
Mast cells are central to skin immune responses, playing substantial roles in inflammation and allergic reactions. These cells express a variety of receptors, including chemokine receptors, to regulate their function. CXCR1 and CXCR2 are commonly expressed on mast cells, with their expression levels increasing following allergic activation, which may facilitate mast cell migration and activation [62]. CXCR3 has been implicated in mast cell migration and activation in several allergic diseases, with CXCR3+ mast cells being recruited to the nasal and tracheal regions, thereby contributing to allergic rhinitis and asthma [63]. CXCR4 also mediates mast cell migration and activation, influencing the release of histamine, cytokines, and other inflammatory mediators, thereby playing a pivotal role in allergic responses [64].
Skin diseases encompass a broad spectrum of conditions affecting the skin, and commonly including inflammatory disorders (such as AD, psoriasis, and acne) [65], infectious diseases (caused by bacteria, viruses, and fungi) [66], autoimmune disorders (such as systemic lupus erythematosus [SLE] and erythema multiforme) [67], and pigmentary disorders (such as vitiligo) [68]. Within the realm of skin diseases, the involvement of CXCR receptors and their corresponding ligands is pivotal, as these molecules directly modulate immune responses and inflammatory processes within the skin (Fig. 3).
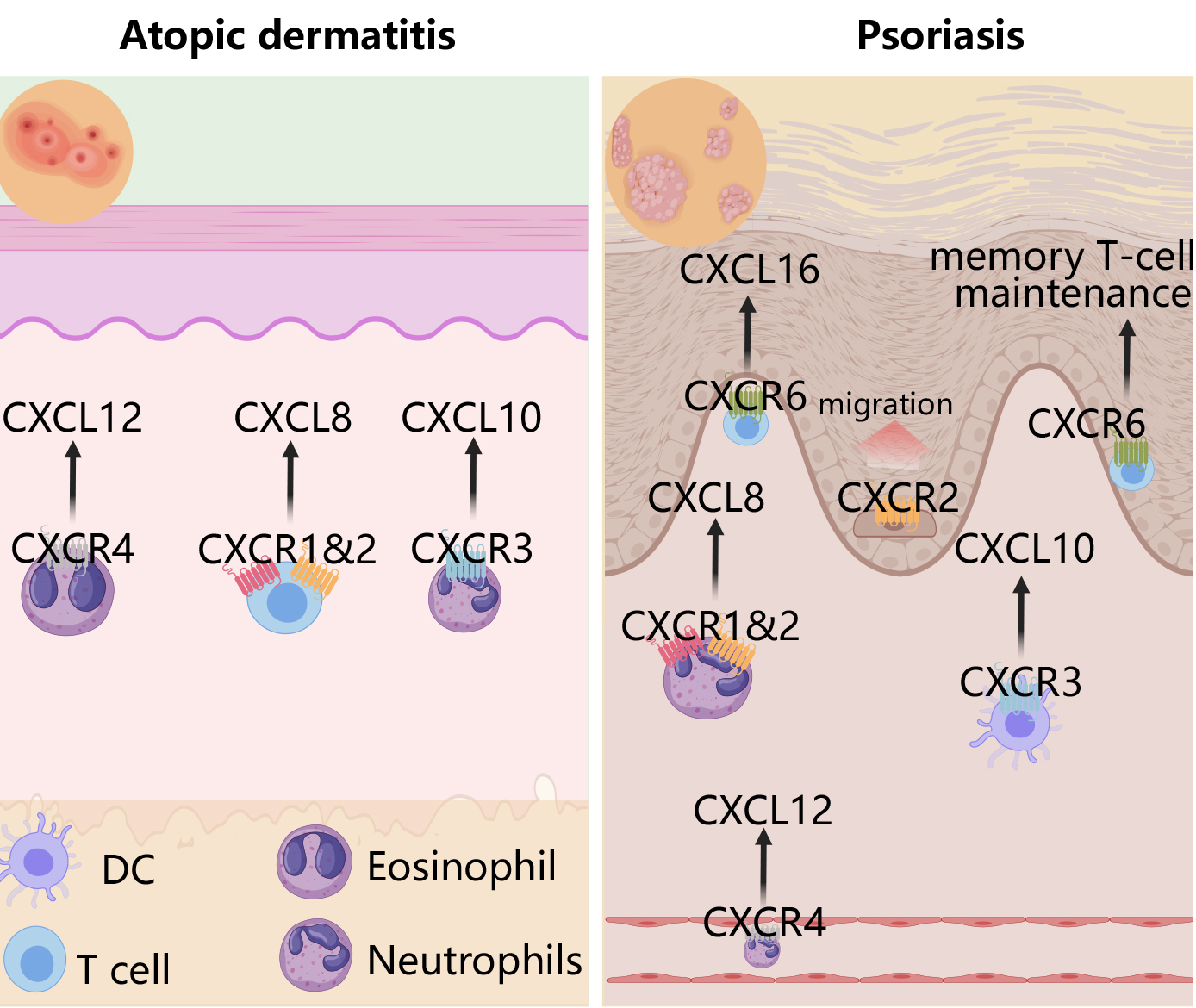 Fig. 3.
Fig. 3.
Roles of the CXCR family in psoriasis and atopic dermatitis. In atopic dermatitis, the CXCL12-CXCR4 axis recruits eosinophils, CXCL8-CXCR1/2 facilitates T cell recruitment, and CXCL10-CXCR3 attracts neutrophils. In psoriasis, CXCL12-CXCR4 promotes neutrophil infiltration into psoriatic lesions, CXCL8-CXCR1/2 amplifies psoriasis-related inflammation by recruiting neutrophils, CXCL10-CXCR3 attracts dendritic cells, and CXCL16-CXCR6 orchestrates T cell migration and retention of tissue-resident memory T cells. Created with MedPeer (https://image.medpeer.cn/show/index/home).
AD is a chronic dermatological disorder that typically manifests in infancy but may persist into adulthood or emerge later in life. The condition is marked by skin inflammation, erythema, pruritus, dryness, and recurrent flare-ups. CXCR receptors play a pivotal role in the pathogenesis of AD. A study utilizing an MC903-treated mouse model revealed that MC903 induces the expression of several genes associated with AD, including various chemokine receptors such as Cxcl1, Cxcl2, Cxcl3, and Cxcl5, suggesting that cytokine-driven pathways may contribute to the immune responses responsible for the characteristic itching and inflammation observed in AD [9].
CXCR2 has emerged as a potential therapeutic target for AD (eczema); treatment with a humanized monoclonal antibody against CXCR2 demonstrated therapeutic efficacy in models of both AD and rheumatoid arthritis (RA) [69]. CXCR3 may facilitate the activation of sensory neurons by neutrophils through its interaction with CXCL10, contributing to pruritus, and CXCR3 inhibitors have been shown to alleviate itching. Additionally, IL-14, a key cytokine involved in the resolution of AD, upregulates CXCR3 in transgenic mouse models, highlighting the receptor’s potential role in AD pathogenesis [70]. CXCR4 expression on eosinophils and T cells mediates inflammation in AD, with notable upregulation of CXCR4 in eosinophils isolated from infants with moderate to severe AD [71]. The CXCL13-CXCR5 axis governs the localization of Tfh cells by attracting them to the T and B cell border, thereby promoting B cell activation and proliferation. In the context of AD, particularly in children with exogenous forms of AD, there is a significant reduction in IL-10+ regulatory B cells, and Breg cells exhibit a diminished capacity to inhibit Tfh cell differentiation in an IL-10-dependent manner [72]. The involvement of CXCR5 in the functional interplay between B cells and Tfh cells, along with their critical roles in secondary lymphoid tissues, suggests that targeting this axis could represent a promising therapeutic strategy for AD.
Psoriasis is a chronic inflammatory skin disorder characterized by erythema, scales, cracks, and itching. The interaction between CXCR2 and BLT1 (leukotriene B4 receptor 1) facilitates the early infiltration of neutrophils in psoriasis [73]. CXCR3 and its ligand CXCL10 are prominently expressed during the active phase of psoriatic plaques [74]. Increased CXCR3 expression in psoriatic lesions drives the recruitment of plasmacytoid dendritic cells and T cells to developing lesions [8]. CXCR3A levels in psoriatic skin correlate with epidermal T cell count [75]. Anti-IL-17 therapy results in a marked reduction of CXCR3+ Th17 cells in peripheral blood [76]. CXCR3 may play a more pivotal role in psoriatic arthritis (PsA), with its upregulation in synovial CD8+ T cells and elevated levels of CXCL9 and CXCL10 in PsA synovial fluid [77]. Recent finding suggests that patients with PsA exhibit higher levels of CXCR3+ memory Th cells in circulation compared to those without joint involvement [78]. Single-cell transcriptome analysis reveals CXCR4 expression in both CD8 Tc17 cells and keratinocytes, mediating psoriasis through inflammatory cytokine production [79]. Early study propose that CXCR4 mitigates psoriasis inflammation by inhibiting keratinocyte proliferation and suppressing Th17 cytokine expression, thereby alleviating IL-23-induced inflammation [80]. Recent research highlights that high CXCR4 expression in neutrophils, driven by CREB1, exacerbates skin infiltration and amplifies psoriasis inflammation [81]. CXCR6 is predominantly expressed in innate lymphoid cells (ILCs), CD8+ T cells, and neutrophils within psoriatic lesions. CXCR6+ ILC1 cells expand rapidly and upregulate IL-23 receptors during early psoriasis, contributing to disease progression [82, 83]. CXCR6 upregulation in CD8+ T cells mediates T cell recruitment and intensifies psoriasis inflammation [84]. Tissue-resident memory T cells (Trm) are implicated in psoriasis relapse [85], with CXCR6 facilitating the survival and residency of CD8+ Trm cells, marking them as key mediators of this process [86]. While research on CXCR7 in psoriasis remains limited, study suggest CXCR7 mRNA upregulation in microvascular endothelial cells in psoriatic lesions [87], underscoring the need for further exploration into its role in the disease.
Acne is an inflammatory disorder affecting hair follicles and sebaceous glands, with pathogenesis involving an innate immune response triggered by Toll-like receptor interactions and inflammasome activation, followed by the onset of adaptive immunity. Research indicates that a high-fat diet enhances neutrophil infiltration around hair follicles via CXCR2 signaling, contributing to acne and folliculitis [88]. Although the role of the CXCR family in acne pathogenesis remains inadequately explored, the existing evidence provides a foundation for further investigation into CXCR involvement in acne.
Infectious skin diseases, caused by various pathogens such as bacteria, viruses, fungi, and parasites, exhibit diverse clinical manifestations depending on the specific pathogen. CXCR1 and CXCR2 play key roles in these diseases by mediating neutrophil recruitment and activation. Inflammation in infected skin upregulates the expression of CXCL9, CXCL10, and CXCL11, facilitating the recruitment of CXCR3-expressing effector T cells to the site of infection. In the absence of inflammation, the CXCR3-CXCL10 axis may promote memory T cell accumulation in the skin, while tissue-resident memory CD8+ T cells in the epidermis form a barrier against infections, independent of CXCR3. CXCR3 signaling is critical for preventing excessive angiogenesis during the pre-remodeling phase of post-infection wound healing. Keratinocytes inhibit vascular endothelial growth factor (VEGF)-induced endothelial cell motility and tubule formation via CXCR3 activation and PKA-mediated calpain inhibition, suggesting that CXCR3 plays a role in refining vasculature and facilitating wound repair [89]. The CXCL12-CXCR4 axis governs immune cell recruitment and migration, which is vital for managing symptoms of skin infections. However, gain-of-function mutations in CXCR4 can result in severe human papillomavirus (HPV) infections, manifested as warts, hypogammaglobulinemia, infections, and myelokathexis (WHIM) immunodeficiency syndrome. Targeting CXCR4 with antagonists offers therapeutic potential for WHIM syndrome, alleviating symptoms of HPV-induced epithelial neoplasia by specifically inhibiting the CXCR4 receptor [90]. Additionally, CXCR4 mediates HIV infection of T cells. While no direct evidence links CXCR5 to infectious skin diseases, CXCR5’s regulation of Tfh cells and B cells is significant in such conditions. CXCR6, predominantly expressed on CD8+ T cells, plays a pivotal role in immune surveillance within non-lymphoid tissues, such as the skin. It is essential for the survival and long-term residency of CD8+ T cells, which are vital for monitoring skin pathogens and optimizing T cell responses in non-lymphoid peripheral tissues [49].
SLE is a chronic autoimmune disease that affects multiple organ systems. Skin manifestations include butterfly-shaped erythema, photosensitivity, rashes, ulcers, and occasionally pruritus or scaling. The underlying mechanism in SLE involves an aberrant immune response, where the body’s immune system attacks its own tissues, leading to inflammation and tissue damage. This is primarily driven by the abnormal activation of B and T cells. Study indicates an increased presence of CXCR3+ cells in patients with SLE, with the accumulation and dysfunction of CXCR3+ T cells serving as potential prognostic biomarkers for the disease [91]. Recent research using CXCR3-deficient mice suggests that CXCR3 plays a critical role in autoantibody production by enhancing the percentage of abnormally activated Tfh cells and B cells, promoting CD4+ T cell migration and helper function in lupus [92].
Vitiligo is a common dermatological condition characterized by the loss of skin
and, in some cases, hair pigmentation. In vitiligo, CXCR family receptors
regulate the migration and localization of immune cells such as T cells and
dendritic cells, which are pivotal in targeting and destroying melanocytes, the
pigment-producing cells. Activation or inhibition of CXCR receptors may therefore
influence the extent of this autoimmune response. Elevated levels of CXCR3
ligands, including CXCL9–11, have been observed in the serum of patients with
vitiligo [93]. Recent study also highlights the role of the
IFN-
Medications targeting the CXCR family have demonstrated considerable promise in treating skin diseases by modulating inflammatory pathways through specific chemokine receptor inhibition. This approach offers more targeted and potentially effective therapeutic strategies. However, the field remains in its early stages, necessitating further clinical trials and research to comprehensively evaluate their efficacy and safety.
In inflammatory skin disorders such as psoriasis and AD, CXCR pathways represent promising therapeutic targets. Both conditions are marked by dysregulated immune microenvironments, characterized by increased Th1 and Th17 cells and decreased Treg cells. Research in RA models has shown that the CXCR3 antagonist AMG487 effectively reduces inflammation, significantly decreases the proportions of Th1, Th17, and Th22 cells, and increases Treg cell levels, highlighting its potential for treating inflammatory skin conditions [96].
In autoimmune diseases, CXCR inhibitors have shown substantial therapeutic promise. A randomized, double-blind, placebo-controlled study evaluating the CXCR5 inhibitor PF-06835375 in SLE and RA found that it effectively depleted B cells, Tfh cells, and circulating Tfh-like cells [97]. These findings underscore the clinical promise of CXCR5 inhibition as a strategy for managing autoimmune diseases.
While pigmentary disorders have been relatively underexplored in this context, the therapeutic potential of CXCR pathways in other diseases suggests possible applications in dermatology. For instance, during melanogenesis, CXCR2 plays a key role in regulating transcriptional programs that promote tumor growth. The CXCR1/CXCR2 antagonist SX-682 has been shown to enhance the expression of the tumor suppressor transcription factor Tfcp2l1, potentially reducing tumor incidence [98]. These findings indicate the broader therapeutic relevance of CXCR modulation in skin-related conditions.
This review provides a comprehensive analysis of the multifaceted roles of the CXCR family in skin diseases, emphasizing the pivotal functions of these receptors in skin immune responses and pathological processes. CXCR receptors are integral in regulating immune cell migration and activation, playing critical roles in skin inflammation, autoimmune skin diseases, and skin tumors. The involvement of CXCR3 and CXCR4 in various inflammatory and autoimmune skin conditions is well-established, while CXCR5 and CXCR6 also demonstrate significance in specific pathological contexts.
Despite considerable advances in understanding the role of the CXCR family in skin diseases, several key questions remain unresolved. For instance, the differential effects of CXCR receptors through distinct signaling pathways in various skin diseases require further exploration. Future research should focus on the interactions between CXCR receptors and their ligands, particularly how these interactions vary across different skin layers and microenvironments.
Additionally, the therapeutic potential of the CXCR family as targets for treatment requires further investigation. While several CXCR-targeting drugs have shown promise in clinical trials, the development of new therapeutic strategies for a broader spectrum of skin diseases, especially those with limited treatment options, remains a crucial area of research. Furthermore, investigating the side effects and long-term outcomes of targeting the CXCR family will be an essential focus of future studies.
JX and CW designed the review. YX, BL and JY participated in the acquisition, analysis and interpretation of research data, and were responsible for systematic literature retrieval and screening, as well as the design and production of research charts. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This research received no external funding.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.