


 , Fareed Ahmad 1,2, Majid Alam 1,2, Abdul Wahid Ansari 1,2, Shahab Uddin 1,2, Martin Steinhoff 1,2,3, Joerg Buddenkotte 1,2,3,*
, Fareed Ahmad 1,2, Majid Alam 1,2, Abdul Wahid Ansari 1,2, Shahab Uddin 1,2, Martin Steinhoff 1,2,3, Joerg Buddenkotte 1,2,3,* , Aamir Ahmad 1,2,*, Angeliki Datsi 4,*
, Aamir Ahmad 1,2,*, Angeliki Datsi 4,*1 Translational Research Institute, Academic Health System, Hamad Medical Corporation, 3050 Doha, Qatar
2 Dermatology Institute, Academic Health System, Hamad Medical Corporation, 3050 Doha, Qatar
3 Department of Dermatology and Venereology, Hamad Medical Corporation, 3050 Doha, Qatar
4 Institute of Transplantation Diagnostics and Cell Therapeutics, University Hospital Dusseldorf, 40225 Dusseldorf, Germany
Abstract
Interleukin 31 (IL-31) is a proinflammatory cytokine, mainly secreted by Type II helper T cells. It signals through a heterodimeric receptor complex composed of IL-31 receptor α and oncostatin-M receptor β chain. The hallmark feature of IL-31, in its pathological role, is its ability to induce pruritus in mammals. Pruritus is a common symptom and major reason of morbidity in cancer patients, compromising their quality of life. Although, IL-31 is differentially expressed in different tumor types and could promote or inhibit cancer progression, high expression of IL-31 is a contributing factor to advanced stage tumor and severity of pruritus. The simultaneous existence of pruritus and cancer could either result from the aberrations in common proteins that co-exist in both cancer and pruritus or the therapeutic treatment of cancer could indirectly induce pruritus. Although the biology of IL-31 has predominantly been described in skin diseases such as atopic dermatitis and other inflammatory diseases, the precise role of IL-31 in the tumor biology of different cancer types remains elusive. Herein, we summarize the current understanding on the role of this cytokine in the pathogenesis of different cancers.
Keywords
- interleukin-31
- pruritus
- cancer
- interleukin-31 receptor α
- oncostatin-M receptor
Interleukin 31 (IL-31) belongs to the proinflammatory IL-6 superfamily of
cytokines in which most cytokines signal through receptor complexes containing
glycoprotein 130 (gp130) [1]. This family contains nine members that include
IL-6, IL-11, IL-27, IL-31, leukemia inhibitory factor, ciliary neurotrophic
factor, oncostatin M (OSM), cardiotrophin-like cytokine and cardiotrophin 1 [1].
Unlike other members of the family, IL-31 does not use the gp130 chain instead it
signals through the IL-31 receptor
IL-31 signals through a heterodimeric receptor complex composed of the IL-31RA
and oncostatin-M receptor (OSMR)
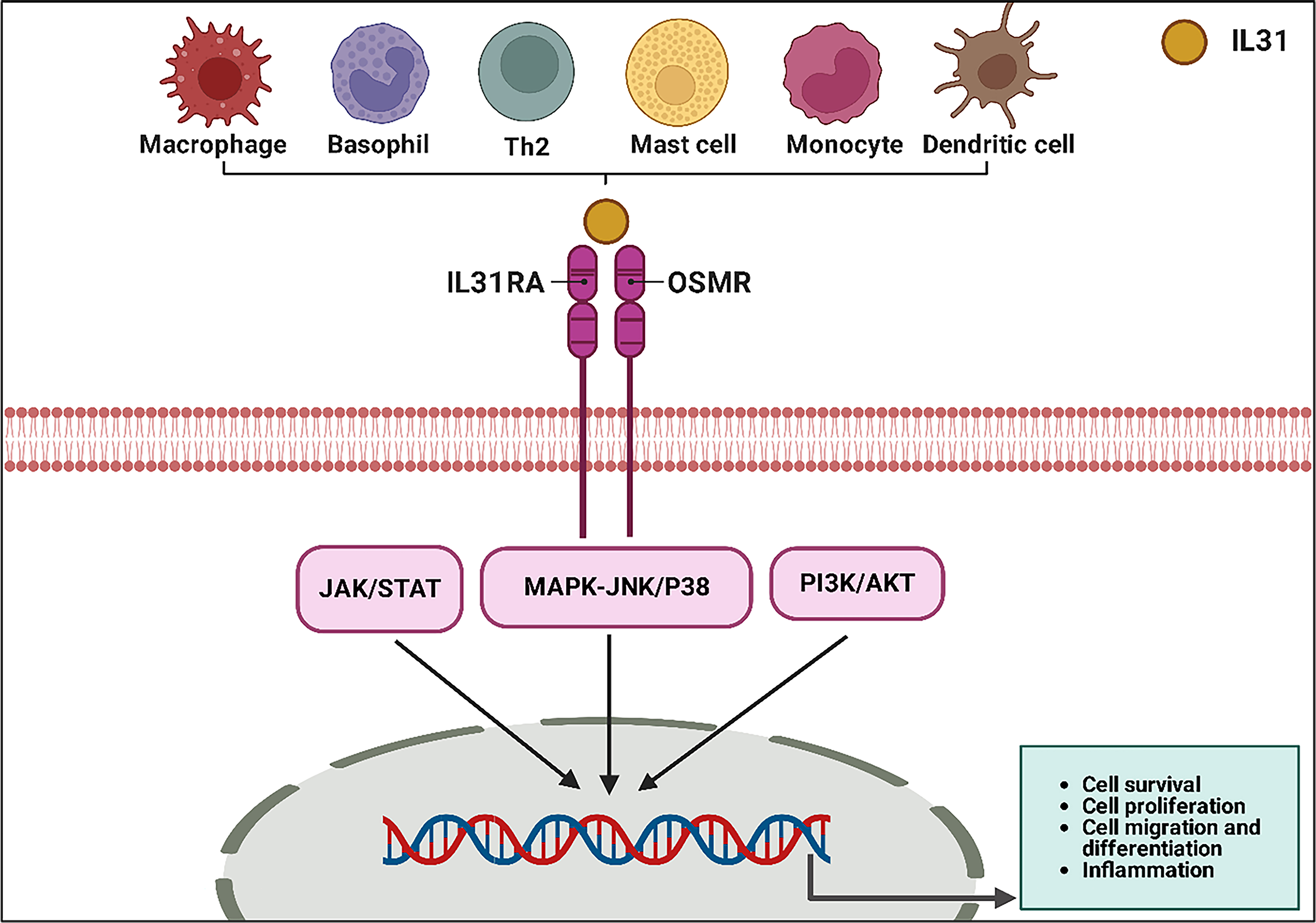 Fig. 1.
Fig. 1.
IL-31 dependent signal transduction. Different types
of immune cells produce IL-31 which signals through the heterodimeric receptor
complex comprising IL-31RA and OSMR. This results in stimulation of the JAK/STAT,
MAPK-JNK/P38 and PI3K/AKT pathways in the target cells. These pathways contribute
to cell survival, proliferation, migration, differentiation and inflammation.
IL-31, Interleukin 31; IL-31RA, IL-31 receptor
IL-31 is an immunoregulatory cytokine, primarily secreted by Th2 cells, and plays a paramount role in immune responses and chronic inflammation. T cell-derived cytokines are important to create effective immune responses but can cause/promote disease upon aberrant expression [14]. IL-31 is differentially expressed in different tumor types and could promote or inhibit cancer progression [15]. Since the microenvironment in tumors shifts towards a more inflammatory state, a compromised immune response of the host can play a critical role in tumor progression [16, 17, 18, 19]. Thus, cell-mediated immune responses are essential to prevent tumor development and progression, and an immunocompromised host immunity leads to cancer proliferation and metastasis [20]. The first involvement of IL-31/IL-31RA axis in cancer was realized in studies in cutaneous T cell lymphoma (CTCL) patients in which the serum levels of IL-31 were correlated with the disease stage of CTCL [21]. In addition to CTCL, IL-31RA mRNA is also expressed in melanoma, glioblastoma, osteosarcoma and myelomonocytic leukemia cell lines [14, 22, 23, 24]. However, the precise role of IL-31 in the tumor biology of different cancer types remains elusive. Herein, we summarize the current understanding on the role of this cytokine in the pathogenesis of different cancers [25].
IL-31 is 18 kDa protein that was first identified and cloned by Dillon
et al. in 2004 [14]. The gene encoding human IL-31 is located on the long region
of chromosome 12 while mouse IL-31 is in the syntenic region of chromosome 5 [13, 14, 24]. The IL-31 precursor protein contains 164 amino acids whereas the mature
IL-31 protein is composed of 141 amino acids that form 4 alpha helices [10, 26].
Mature mouse IL-31 protein shares 31% amino acid sequence identity with its
human counterpart, however, it does not interact with the human IL-31 receptor
and vice versa [14, 24]. Dillon and colleagues [14] identified IL-31 by employing
a functional cloning approach that was based on the proliferation of cells
expressing IL-31RA. They found that the IL-31 identified in the conditioned media
derived from the activated T cell line and activated human peripheral T cells,
induced proliferation only in the IL-31RA and OSMR
Pruritus is a common symptom of several inflammatory skin diseases which affects
patient’s quality of life. Numerous immune cells and mediators play a role in the
physiology of itching such as Th2 and mast cells. Among many cytokines, Th2
cytokine IL-4, IL-13, IL-25, IL-31, IL-33 and thymic stromal lymphopoietin are
not only pro-inflammatory cytokines but are also important regulators of itch
[27, 32, 33]. The significance of Th2 cytokines in mediating itch in cancer is
eminent from the findings that the administration of dupilumab (an antibody
against IL-4
The hallmark feature of IL-31, in its pathological role, is its ability to induce pruritus in mammals [36]. Pruritus is a common symptom and major reason of morbidity in cancer patients, compromising their quality of life [37]. Pruritus could potentially be associated with cancer in three situations. First, it could display the first signal of the presence of tumor cells due to the systemic release of oncologic mediators thus, constituting a paraneoplastic syndrome. Second, itch could be mediated by the physical presence of tumor mass. Third, pruritus could be the consequence of anti-neoplastic drugs [7]. The role of IL-31 in itch is based on the finding that IL-31 transgenic mice present with continuous scratching behavior resulting in atopic dermatitis-like skin lesions [14]. Moreover, continuous injection of IL-31 in BALB/c mice induces sustained scratching and development of dermatitis [38, 39]. Pruritus has been observed in several cancers such as chronic lymphocytic leukemia, Hodgkin’s lymphoma, chronic myelomonocytic leukemia, multiple myeloma and in solid tumors including gastric, lung and laryngeal tumors [40, 41, 42, 43, 44]. It is typical of some cancers such as CTCL, leukaemia, Hodgkin’s lymphoma and biliary cancer [7]. Itch is a common symptom of both Hodgkin’s and non-Hodgkin’s lymphoma affecting about 25% of patients with Hodgkin’s disease [45]. Considering the role of IL-31 in pruritus and cancer, targeting the IL-31 signaling pathway could be an effective way to develop therapies against them and for the clinical management of these diseases.
Leukemia is a hematologic disorder, characterised by an increased number of leucocytes in blood and/or bone marrow [46]. Lymphomas represent a heterogenous group of lymphoid malignancies, that display different clinical behavior and responses to treatment [47]. Lymphomas mainly originate in B cells but can also form in T cells or natural killer cells and are grouped into Hodgkin and non-Hodgkin lymphomas [48]. Cytokines play an important role in lymphoma as they mediate interactions between immune and non-immune cells, eventually inhibiting or promoting tumor growth [49].
Chronic myeloid leukemia (CML) originates in the hematopoietic stem cells during the chronic phase of the disease. It is characterised by myeloid cells of varying maturation stages in peripheral blood and bone marrow [50]. The first targeted therapy of CML, imatinib mesylate, was introduced in 2001 [51]. It is a tyrosine kinase inhibitor (TKI) that induces skin manifestation including pruritus, in CML patient, through the IL-31/IL-33 axis [52]. The potential injury to keratinocytes upon administration of imitanib mesylate causes the release of IL-33 which in turn interacts with its cognate receptor on mast cells, inducing the release of IL-31 (Fig. 2). Additionally, the serum levels of IL-31 in a 73-year-old CML patient were found to be higher when compared to 18 gender- and age-matched healthy controls [52]. However, to confirm the correlation of IL-31 and TKIs as cause of itch in CML, further studies are needed on different TKIs and a large cohort of CML patients.
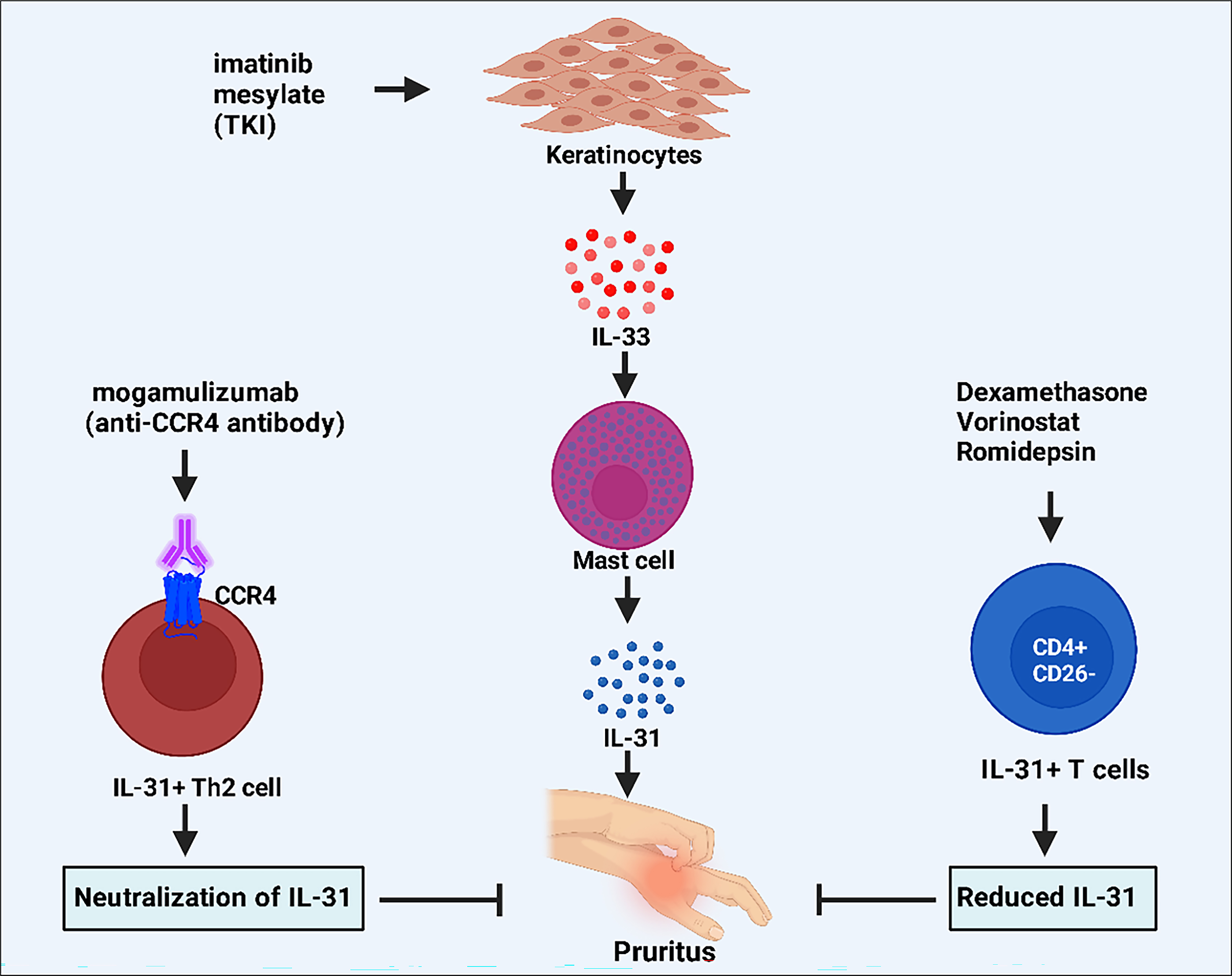 Fig. 2.
Fig. 2.
Role of IL-31 in pruritus. Indirect neutralization of the IL-31 in Th2 cells, through anti-C-C chemokine receptor type 4 (CCR4) antibody reduces Cutaneous T cell lymphoma (CTCL)-accompanied pruritus in the patients. The potential injury to keratinocytes upon administration of imitanib mesylate induces the release of IL-33 which further causes the secretion of IL-31 from mast cells, triggering pruritus in Chronic myeloid leukemia. Moreover, therapeutic administration of dexamethasone, romidepsin and vorinostat reduce IL-31 in CD4+ CD26– T cells of CTCL patients. TKI, tyrosine kinase inhibitor. Created with BioRender.com.
CTCL belongs to the diverse group of non-Hodgkin’s lymphomas presenting with a cutaneous infiltrate dominated by skin homing lymphocytes and is accompanied by episodes of intense, therapy-refractory pruritus [37, 53]. Pruritus is a hallmark symptom in both subtypes of CTCL; mycosis fungoides and Sézary Syndrome [54, 55]. However, patients with Sézary syndrome, and the erythrodermic form of mycosis fungoides report a more severe itch than patients with patch stages of mycosis fungoides [56]. CTCL patients endure a significant burden of physical symptoms that affect their emotional, social and physical well-being. Pruritus is one of the common symptoms in CTCL that affect about 88% of CTCL patients [57, 58]. High IL-31 expression in CTCL seems to be associated with disease severity since serum of advanced stage CTCL patients exhibits increased levels of IL-31 compared to early-stage CTCL patients [37]. Although two studies have reported no correlation of serum IL-31 expression in CTCL patients with pruritus [21, 59], an association of the IL-31 axis with pruritus at the advanced stages of the disease was reported. This could be explained by the hypothesis that IL-31 is released by multiple cells and the source of IL-31 at the early stages of the disease could be different than the later stages hence it results in different outcomes [37, 59].
Nattkemper and colleagues investigated the expression and distribution of the
IL-31 peptide and its cognate receptor in CTCL-affected skin. The group found an
increase in IL-31 expression in the epidermis of the skin along with a pronounced
lymphocytic infiltrate [58]. In addition, IL-31RA appears to form a greater and
denser expression pattern in the epidermis of patients with CTCL, implying that
permanent IL-31 exposure could account for the epidermal upregulation of the
heterodimeric IL-31RA/OSMR
Moreover, a pathophysiological role of IL-31 in CTCL is supported by the finding that therapeutic administration of dexamethasone and histone deacetylase inhibitors (romidepsin and vorinostat) markedly reduced IL-31 in CD4+ CD26– T cells of CTCL patients [60]. To further test if IL-31-producing T cells are involved in the pathogenesis in CTCL patients, the group indirectly eliminated the IL-31+ Th2 cell subpopulation by targeting the skin homing chemokine receptor type 4 (CCR4)+ Th2 cells with mogamulizumab, a humanized monoclonal anti-CCR4 antibody. Indirect neutralization of the IL-31 pathway in fact markedly reduced CTCL-accompanied pruritus in the patients, which indicates a pathological role of IL-31 in CTCL and highlights that the IL-31/IL-31RA axis as a potential therapeutic target in CTCL patients suffering from therapy refractory itch [60] (Fig. 2). However, if IL-31 signaling contributes to the development and/or progression of CTCLs in humans is thus far not known.
While several studies have been conducted on the involvement of IL-31 in T cell lymphomas, little is known about its role in B-cell lymphomas. Ferretti et al. [61], investigated the cytokine’s impact in follicular lymphoma (FL), a B-cell malignancy derived from the germinal center, and identified a role for the IL-31/IL-31RA axis in FL tumour progression. FL is a slow-growing, non-Hodgkin lymphoma that represents 35% of all non-Hodgkin lymphomas [62]. Despite being a B-cell malignancy, the reactive lymph nodes of FL are also inhabited by CD4+ T cells and CD68+ macrophages that express IL-31. The functional role of the IL-31/IL-31R axis in FL is demonstrated by an increase in Erk1/Erk2, Akt and STAT3 phosphorylation as well as subsequent cell proliferation upon incubation of purified FL cells (derived from lymphoma B cells) with human recombinant IL-31 [61]. In a translational approach, IL-31RA and OSMR expression was found in lymph nodes and in malignant B cells of FL patients. Lymph nodes obtained from later stage FL patients exhibited an increase of IL-31+ and IL-31RA+ cells than lymph nodes obtained from early stage FL patients indicating an association of the IL-31/IL-31RA signaling complex with FL progression [61].
Hodgkin Lymphoma (HL) is a B-cell lymphoma, typically found in cervical lymph nodes [63]. Ferretti et al. [64] described a correlation of IL-31 plasma levels in patients with HL. The neoplastic tissue in HL releases cytokines that shape the tumor microenvironment and leads to pruritus [64]. HL patients suffer from intense pruritus which is potentially correlated to IL-31 expression. IL-31 and its receptors are expressed on the surface and in the cytoplasm of Hodgkin cells. Moreover, IL-31 is expressed in B cells, CD4+ T cells and macrophages that were derived from lymph nodes of HL patients [64]. HL presents with an inflammatory profile characterized by Hodgkin cells that are surrounded by diverse immune cells of the innate and the adaptive immune system, including B and CD4+ T cells. The study suggests that IL-31 derived from tumor-infiltrating Th2 cells could provide a beneficial microenvironment for the growth of Hodgkin cells that express the IL-31R [64]. Moreover, the patients at higher risk of relapse also exhibited high levels of soluble IL-31. The group’s findings imply an autocrine/paracrine feed-forward loop that skews the tumor microenvironment towards a tumour progressive state [64].
Present data on the role of IL-31 in lymphomas indicate the pathophysiology of this cytokine and its potential involvement in tumor progression. Moreover, studies also suggest its role in inducing pruritus in both Hodgkin and non-Hodgkin lymphomas. However, the mechanisms through which IL-31 induces tumor progression in lymphomas are still not known and hence, warrant further investigation.
Solid cancers are highly prevalent and a major cause of morbidity and mortality worldwide [65]. These are the tumors from solid organs such as lung, breast, colon, prostate, ovaries etc., and are treated with multiple strategies among which chemotherapy, radiotherapy and surgical removal are the most common. However, their success is limited as different tumors develop distinct resistance mechanisms, which can then result in tumor relapse [65, 66, 67, 68]. The treatment of these cancers through the modulation of tumor-growth promoting cytokines, such as IL-31, could serve as an effective therapy. Therefore, in this section we will discuss the emerging role of IL-31 in solid tumors.
Breast cancer (BC) is the most common cancer and a major cause of mortality among women worldwide [69, 70]. The anti-tumor effects of IL-31 have been reported in BC through immunomodulation. To assess the immunomodulatory role of IL-31 under the normal setting, meaning tumor free BALB/c mice, mice were infused with recombinant IL-31 and compared to IL-31-infused tumor-bearing BALB/c mice. Kan and co-workers observed an IL-31-induced proinflammatory state in the myeloid and lymphoid lineage of tumor-bearing BALB/c mice compared to the control mice. They reported an immunomodulatory property of IL-31 due to an increased expression of CD8+, CD4+ T cells and central memory T cells (CD62L+CD44+) in contrast to a reduction of polymorphonuclear myeloid derived suppressor cells (Ly6CdimLy6G+) and macrophage (CD11b+F4/80+) frequencies in the spleen of BALB/c non-tumor bearing mice [12].
IL-31 induces an anti-tumor immunomodulation in tumors by increasing CD8+ T-cell numbers and granzyme B and lowering the frequency of CD4+ T cells, indicating cytotoxicity. Similarly, the intraperitoneal injection of recombinant IL-31 into tumor bearing C57Bl/6 mice resulted in an elevated infiltration of CD8+ T cells into the BC tumor which eventually reduced its growth. These findings shed light on the antitumorigenic mechanism of IL-31 by diminishing immunosuppressive cells and promoting cytotoxic T lymphocytes (CTLs) in the tumor and circulation [12]. The anticancer effect of IL-31 is also demonstrated by the finding that the IL-31/IL-31RA axis contributes to BC patient survival by modulating Th2-related cytokines such as IL-10 and IL-2. IL-31RA mRNA expression positively correlates with IL-2/IL-4 mRNA expression, granzyme B protein expression and an improved overall survival of BC patients [12]. Thus, the immunomodulatory property of IL-31 imparting antitumor effects in BC patients warrants to investigate the therapeutic potential of this cytokine in cancer patients in greater detail in the future [12].
IL-31 induces immunomodulation in myeloid derived suppressor cells (MDSCs) and macrophages that affects the motility and migratory properties of immunosuppressive MDSCs in mice, eventually reducing their ability to infiltrate tumors. Since MDSCs regulate CTL expansion and CTL activity [71], their absence in the tumor tissue due to IL-31 leads to an increase in the activation of CD8+ CTLs [12]. In IL-31 expressing BC, tumor infiltrating immune cell populations were made up of elevated numbers of central memory CD8+ T cells, increased proinflammatory M1 macrophages and decreased immunosuppressive M2 macrophages [12]. Besides the immunomodulation, IL-31 also affects the tumor size in BC as tumor growth was reduced in tumor-bearing mice when they were orthotopically implanted with stably IL-31-expressing BC cells [12].
In BALB/c mice injected with 4T1 BC cells, IL-31 inhibited tumor growth and microvessel density and led to the decrease of pulmonary metastasis [15]. In addition, IL-31 enhanced the therapeutic effect of paclitaxel in Non obese diabetic - severe combined immunodeficiency (NOD-SCID) mice bearing BC also by reducing microvessel density, and pulmonary micrometastases when compared to mice treated with paclitaxel alone. This finding indicates that IL-31 could serve as an adjuvant to classic chemotherapy potentiating the primary therapies’ antitumor effects [15].
However, IL-31 has also been reported to act as pro-oncogenic, for instance by supporting the progression of basal like BC. In BC patients, an elevated IL-31RA expression is found in basal like BC patients when compared to the patients with other BC subtypes. The pathological role of IL-31RA in basal like BC might depend on its activation of STAT3 since gene silencing of IL-31RA effectively represses STAT3 phosphorylation in MDA-MB-231 and MDA-MB-157 cells [72]. Moreover, IL-31RA knockdown in MDA-MB-231 cells prevented tumorsphere formation, as well as migratory and invasive properties of BC cells, upon the introduction of IL-31, supporting a putative role of IL-31RA in conferring cancer stem-like properties in basal like BC [72]. In addition, the injection of IL-31RA knockdown MDA-MB-231 cells into BALB/c nude mice decreased the primary tumor size and reduced pulmonary nodules when compared to mice receiving MDA-MB-231 cells with functional IL-31RA, suggesting that IL-31RA is required for tumor growth and metastasis [72]. In the same manner, the introduction of IL-31 in MDA-MB-231 cells and the overexpression of IL-31RA in MCF-7 luminal BC cells increased their tumorsphere formation, invasive and migratory properties [72]. Interestingly, the mRNA expression levels of IL-31RA and Twist, (an epithelial-to-mesenchymal transition marker) are elevated in basal like BC and correlate positively [72, 73]. Twist/BRD4 complex has also been reported to directly regulate gene transcription of IL-31RA [72].
Lung cancer is one of the malignant cancers and major cause of mortality in the world [74, 75]. The overall 5-year survival rate of lung cancer patients is less than 20% [75].
A study on a case presentation of a 61-year-old non-smoking woman with bronchioalveolar carcinoma demonstrated a possible association of adverse skin conditions with bronchioalveolar carcinoma and high IL-31 serum levels. Serum IL-31 levels in the patient were significantly higher than in 10 healthy controls (5725.9 pg/mL vs. 1722.9 pg/mL). IL-31 serum levels correlate with disease progression and cutaneous symptoms including skin rash, pruritus and xerosis during treatment with epidermal growth factor receptor tyrosine kinase inhibitors (EGFR-TKIs) (erlotinib and gefitinib) [76]. However, the presence of pruritus and skin rashes could possibly be associated mainly with the EGFR-TKIs as they appeared in the patient only during the treatment and disappeared after therapy discontinuation. EGFR-TKIs commonly induce pruritus in patients and thus compromise quality of life and treatment compliance [77]. However, further translational research studies in lung cancer are required to validate this initial observation, to understand the role of IL-31/IL-31RA across the various lung cancer subtypes and to identify cancer subtype-dependent mechanism(s) of IL-31 on a molecular level.
Gastric cancer (GC) is the sixth most common cancer and third leading cause of cancer-related deaths worldwide [78, 79]. Despite significant improvements in the treatment strategies, GC remains an aggressive malignancy with an approx. 5-year survival rate of 31% [80]. Studies proposed an antitumoral potential of IL-31 in GC based on the observation that IL-31 was markedly reduced in GC tissues compared to the adjacent non-cancerous gastric tissues [25, 81]. IL-31 expression levels appear to negatively correlate with the aggressiveness of GC since a reduced expression is observed in younger patients that have poorly differentiated and highly malignant tumors [25]. Younger patients present with a rapid disease progression, diffused infiltration, poor prognosis, advanced stage malignancy, and histologically undifferentiated GC than the older patients [82, 83, 84]. However, this could possibly be due to delayed diagnosis in younger patients [85].
The two opposite roles of IL-31 in different cancers can be explained by the signaling of IL-31 through different isoforms of the IL-31RA. IL-31 signals through the short and long isoform of IL-31RA. These two isoforms induce contrary functions as the long isoform of IL-31RA is reported to be involved in the aggressiveness of cancer whereas the signaling of IL-31 through the short isoform induces anticancer effects. Considering this hypothesis, it could be stated that IL-31 potentially mediate anticancer effects in GC through the short isoform of IL-31RA [25, 86].
Colon cancer has a high incidence and mortality rate in developed countries. It is often combined with rectal cancer due to common features of both cancers [87]. IL-31 appears to exert anti-tumor effects in colon cancer as IL-31-silenced MC38 cells showed a faster tumor growth in C57Bl/6 mice than IL-31-expressing control cells. The intraperitoneal injection of IL-31-IgG inhibited colon cancer growth in NOD-SCID mice that had formed HCT116 cells-induced colon carcinoma [15]. Similarly, mice infused with recombinant murine IL-31 showed complete inhibition of tumor growth compared to control mice infused with vehicle. The recombinant murine IL-31 treated tumor showed low percentage of endothelial cells and a low number of microvessels indicating that IL-31 could inhibit angiogenesis in colon cancer [15]. The silencing of IL-31 using siRNA in murine colon carcinoma cells, MC38 increased invasion and migration compared to control cells. In the same manner, culturing wild type MC38 with IL-31, reduced invasion and migration compared to untreated cells [15]. However, IL-31 does not affect the cell proliferation, apoptosis, and cell cycle of MC38 and CT-26 colon cancer cells [15].
Endometrial cancer (EC) is the second most common cancer in women. It has a high incidence rate and the 5-year survival rate among distant metastasis patients is 17% [88, 89, 90, 91]. IL-31 levels have been reported to be elevated in the serum of EC patients when compared to the serum of healthy controls. Moreover, the serum levels of IL-31 are directly correlated to the EC stage. However, an association of IL-31 with adverse clinical outcomes such as tumor invasion and metastasis, is less clear [89]. The ambiguous association of IL-31 with tumor invasion, metastasis and tumor stage could be explained by the hypothesis that in EC IL-31 originates from the mast cells and that its secretion is under control of IL-33. Indeed, IL-33 serum levels correlate better with clinical characteristics of EC such as tumor invasiveness and metastasis than IL-31 [89].
Studies have revealed a close link between IL-31 and IL-33 and their potential association with disease severity in autoimmune diseases, allergy and cancer [92, 93, 94]. The presence of one cytokine could stimulate the other, which amplifies inflammation and consequently induce detrimental effects. In human keratinocytes, IL-31 increases the expression of nuclear IL-33 and activates STAT3. Nuclear IL-33 binds to phosphorylated STAT3 and acts as STAT3 transcription co-factor, thereby inhibiting differentiation markers such as filaggrin [95]. This indicates that IL-31 exerts its pathological effects by activating STAT3 signaling through IL-33. Moreover, IL-31 and IL-33 play a synergistic role in the release of Atopic Dermatitis (AD)-related chemokines from basophils since the receptors for both cytokines are expressed on dermal fibroblasts. The interaction of fibroblasts and basophils results in the release of chemokines in response to EGFR-TK inhibitor therapy [94]. Besides, the co-dependency of these two cytokines and IL-31’s association to pruritus could also be explained by the mechanisms initiated by the administration of TKIs, as discussed earlier. Hence, the regulation of the IL-31/IL-33 axis could be an effective therapeutic strategy to manage inflammatory disorders.
Ovarian cancer (OC) is the third most common gynecologic cancer after cervical and uterine cancers [96]. It has high mortality rates owing to delayed onset of symptoms and advanced stage diagnosis [97, 98]. IL-31 displays a clinicopathologic correlation with immune infiltration and expression of p-STAT3 in OC. Immunohistochemistry of the tissues from six OC patients (3 poor and 3 well prognosis) revealed an elevated expression of IL-31 and p-STAT3 in tissues from patients with poor prognosis compared to patients with good prognosis [99]. Moreover, the expression of IL-31 and p-STAT3 is higher at immune cells populated areas of the tissues. The association of IL-31 with STAT3 in OC indicates that STAT3 is an important transcription factor in inducing adverse effects in gynecologic cancers. Apart from this finding, there is a lack of literature regarding the role of IL-31 in the development and progression of human epithelial OC. Therefore, further studies are needed to assess the role of the IL-31/IL-31RA axis in the pathomechanism of OC and to evaluate its status as a therapeutic target for the treatment of OC [99].
Based on the discussion presented in this section, it is evident that a role of IL-31 in different human cancers is increasingly being appreciated (Table 1, Ref. [12, 15, 37, 52, 58, 61, 64, 72, 76, 81, 89, 99]).
| Cancer type | Specimen/cell line | Expression/mechanism | Reference |
| Chronic Myeloid Leukemia | Serum | High IL-31 expression | [52] |
| Cutaneous T cell lymphoma | Serum | Elevated IL-31 expression in advanced stage CTCL patients | [37] |
| Skin tissue | High IL-31 and IL-31RA in the epidermis | [58] | |
| Follicular lymphoma | Follicular lymphoma cells | Increased phosphorylation of Erk1/Erk2, Akt and STAT3 and subsequent cell proliferation | [61] |
| Lymph nodes | High number of IL-31+ and IL-31RA+ cells | [61] | |
| Hodgkin Lymphoma | Hodgkin cells | IL-31 provides beneficial environment for the growth of IL-31R expressing HL cells | [64] |
| Breast cancer | C57Bl/6 mice | Confer anti-cancer property by diminishing immunosuppressive cells and promoting CD8+ T cells | [12] |
| MDA-MB-231 and MDA-MB-157 | IL-31 works as pro-cancerous cytokine as the gene silencing of IL-31RA effectively represses STAT3 phosphorylation | [72] | |
| Lung cancer | Serum | High IL-31 levels correlate to disease progression and cutaneous symptoms | [76] |
| Gastric cancer | Cancer tissue | Reduced IL-31 expression in GC tissues | [81] |
| Colon cancer | C57Bl/6 mice | IL-31 reduced growth, angiogenesis, invasion and migration | [15] |
| Endometrial cancer | Serum | High IL-31 levels in EC patients that correlates with EC stage | [89] |
| Ovarian cancer | Tumor tissue | Poor prognosis in patients exhibiting increased IL-31 and p-STAT3 | [99] |
CTCL, cutaneous T cell lymphoma; IL-31, Interleukin 31; IL-31R, Interleukin (IL)-31 receptor; IL-31RA, Interleukin
(IL)-31 receptor
Patients presented at the hospitals with pruritus have an increased likelihood of underlying malignancy beyond dermatological diseases, when compared to non-pruritic patients. Pruritus is strongly associated with cancers of the skin, liver, hematopoietic system, gall bladder and biliary tract [44]. The simultaneous existence of cancer and pruritus could arise from common protein aberrations, one of which is gastrin releasing peptide receptor, a member of the bombesin G protein coupled receptors, which mediates itch in mice and is abnormally overexpressed in numerous cancers [100]. Despite the evidence of pruritus being a major source of morbidity in various solid and hematologic cancers, it has not been studied in detail in cancer patients directly, which leads to unmet concerns in formulating effective cancer therapies, thus, raises opportunities and challenges for future investigations.
Although the biology of IL-31 has predominantly been described in skin diseases such as atopic dermatitis and other inflammatory diseases, this review summarizes the role of this cytokine in cancer onset and progression. Further, not only the ameliorating effect of IL-31 on tumour cells and potential growth inhibition of these malignant cells requires further insights, but also studies need to be conducted to investigate if cancer associated itch can be abrogated by one of the newly developed therapeutic agents targeting the IL-31/IL-31RA axis. Blockage of IL-31 could potentially prolong patient survival or even cure cancer in combination with already existent chemotherapies. Studies reveal unprecedented findings on the implication of IL-31 in cancer progression. They have reported the role and association of IL-31 and p-STAT3 in different solid cancers in which the pathological role of IL-31 is displayed through the aberrant activation of STAT3. Although IL-31 has largely been investigated for its oncogenic potential, other studies have also stated its anti-cancer role which could depend on the interactions of IL-31 within the immune microenvironment that needs to be further characterized.
The treatment of IL-31 overexpressing cancers by using TKIs induces pruritus which could compromise treatment compliance in patients, therefore, targeting IL-31 in those cancer types could help improve quality of life and treatment conformity in these patients. However, IL-31 also plays a role as an anti-tumor cytokine and enhances the anti-cancer potential of chemotherapy (paclitaxel) [15] which shows that the role of IL-31 in chemotherapy is increasingly being emphasized. The studies presented in this review give initial insights into the role of IL-31 in different cancer types, but there are still numerous aspects of this cytokine that remain elusive. Further investigations on the immune regulatory function of IL-31, rather than its pro-inflammatory effects, could provide better insight on the role of this cytokine in cancer.
Moreover, it is of utmost importance to understand potential differences, let it be downstream signaling pathways, expression patterns and variants of IL-31 or its cognate receptors between different cancer types, to understand the double-edged role of this pruritogenic cytokine in either tumor progression or tumor protection.
SA, FA, MA, AWA, SU, MS, JB, AA and AD conceptualized, wrote and edited the manuscript. SA prepared the figures. JB, AA and AD supervised the manuscript. All authors approved the final manuscript. All authors have participated sufficiently in the work to take public responsibility for appropriate portions of the content and agreed to be accountable for all aspects of the work in ensuring that questions related to its accuracy or integrity.
Not applicable.
Not applicable.
This research received no external funding.
Martin Steinhoff is a consultant or advisory board member for Pfizer, Janssen, Eli-Lilly, Novartis, Abbvie, Almirall, Avon, UCB, Celgene, Galderma, Leo, MenloTx, Mitsubishi, Maruho, Toray, Sanofi, Regeneron. Grants by Pfizer, Abbvie, Almirall, Novartis, Leo, Travi, Galderma, Toray, Mitsubishi, Maruho, Avon; Speaker for Pfizer, Janssen, Eli-Lilly, Novartis, Abbvie, UCB, Celgene, Galderma, Leo, MenloTx, Sanofi, Regeneron. Aamir Ahmad is an adviser for BioWiz Laboratories, Inc., Detroit, MI, USA. Given his role as a guest editor, Dr. Shahab Uddin had no involvement in the peer-review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process for this article was delegated to Prof. Alister C. Ward. The other authors declare no conflicts of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.