










 , Oksana Fuks 1, Natalya Timkina 1,2, Alina Islamova 1, Dina Sufieva 3, Оlga Kirik 3, Dmitrii Korzhevskii 3, Timur Vlasov 4, Tatiana Karonova 1,2
, Oksana Fuks 1, Natalya Timkina 1,2, Alina Islamova 1, Dina Sufieva 3, Оlga Kirik 3, Dmitrii Korzhevskii 3, Timur Vlasov 4, Tatiana Karonova 1,21 Institution of Endocrinology, Almazov National Medical Research Centre, 197341 Saint-Petersburg, Russia
2 Faculty Therapy Department, Pavlov First St. Petersburg State Medical University, 197022 Saint-Petersburg, Russia
3 Laboratory of Functional Morphology of Central and Peripheral Nervous System, Institute of Experimental Medicine, 197022 Saint-Petersburg, Russia
4 Faculty of Pathophysiology, Pavlov First St. Petersburg State Medical University, 197022 Saint-Petersburg, Russia
Abstract
Background: Acute and chronic brain damage in type 2 diabetes mellitus (DM) determines the need to investigate the neuroprotective potential of glucose-lowering drugs. The purpose was to directly compare the neuroprotective effects of glucagon-like peptide-1 receptor agonists (GLP-1RAs) with different duration of action and sodium-glucose cotransporter-2 inhibitors (SGLT-2i) in type 2 diabetic rats with and without stroke. Methods: DM was modelled using high-fat diet and nicotinamide+streptozotocin protocol. The following groups (n = 15 each) were formed: DM without treatment, treatment with liraglutide, dulaglutide, canagliflozin as well as control group without DM and treatment. After 8 weeks, 10 rats from each group underwent middle cerebral artery occlusion. In the reperfusion period neurological deficit, neuroglial damage markers and brain necrosis were evaluated. Brain slices from the remaining 5 animals in each group were histologically examined for microglial activation and neuronal damage. Results: Brain damage was similar in “DM” and “Control” (17.53 [14.23; 26.58] and 15.87 [13.40; 22.68] % of total brain volume, respectively). All study drugs diminished damage volume comparing with “DM” and “Control” whereas the necrosis volume in “DM+Liraglutide” was smaller than in “DM+Canagliflozin” and did not significantly differ from “DM+Dulaglutide” (2.9 [1.83; 4.71], 6.17 [3.88; 8.88] and 4.57 [3.27; 7.90] %). The neurological deficit was more prominent in “DM” than in “Control”, while all the drugs demonstrated similar positive effect. Neurofilament light chains (NLC) did not differ between “DM” and “Control”. Dulaglutide and canagliflozin caused a marked decrease in NLC. Protein S100BB level was similar in “DM” and “Control”. Liraglutide caused the largest S100BB decrease, while canagliflozin did not influence it. In chronic brain ischaemia, all drugs increased the number of normal neurons, but GLP-1RAs had a more pronounced effect. DM was accompanied by increased number of activated microglial cells in Cornu Ammonis (CA)1 hippocampal region. Both GLP-1RAs reduced the number of Iba-1-positive cells, with dulaglutide being more effective than liraglutide, whereas canagliflozin did not affect this parameter. Conclusions: GLP-1RAs and SGLT-2i have neuroprotective properties against acute and chronic brain damage in diabetic rats, although the infarct-limiting effect of GLP-1RAs may be more pronounced. GLP-1RAs and SGLT-2i exert their protective effects by directly influencing neuronal survival, whereas GLP-1RAs also affect microglia.
Keywords
- diabetes mellitus
- stroke
- chronic brain damage
- neuroprotection
- microglia
- GLP-1RAs
- SGLT-2i
Ischemic stroke comprises one of the most serious macrovascular complications and one of the major causes of death in type 2 diabetic patients. The risk of stroke in type 2 diabetes mellitus (DM) is twice as large as in the general population and increases with DM duration [1].
Moreover, almost one in two people with type 2 diabetes mellitus (DM) has chronic brain dyscirculation. This condition can be associated with more or less pronounced cognitive deficits in the absence of focal neurological symptoms and can lead to a dramatic reduction in quality of life and even life expectancy [2]. Chronic brain dyscirculation or chronic brain damage can be characterized by cerebral atrophy, decrease in grey matter volume, dilatation of the ventricles and subarachnoid space, decreased density of the space surrounding the lateral ventricles - leukoariosis, “periventricular space glow” [3]. Importantly, in chronic brain damage small focal lacunar infarctions in the white substantive and subcortical ganglia can be observed, so-called microstrokes or silent strokes. Due to their volume, they do not cause the classic symptoms of unilateral motor and sensory dysfunction, but their pathogenesis is very similar to that of clinically manifest stroke [4].
Nowadays, the concept of the so-called glucose-centric model of type 2 DM treatment has changed to a disease-modifying model, in which the organ-protective properties of glucose-lowering drugs are given priority. Thus, according to existing guidelines [5, 6], glucagon-like peptide-1 receptor agonists (GLP-1RAs) and sodium-glucose co-transporter type 2 inhibitors (SGLT-2i) are the two leading groups of glucose-lowering drugs with the widest spectrum of pleotropic organ-protective effects, including cardio- and nephroprotective properties. Nevertheless, data concerning neurotropic effects of the above-mentioned drug classes remain limited. Thus, according to the meta-analyses summarizing randomized controlled trials (RCTs) results, only two drugs can certainly decrease ischemic stroke risk in type 2 diabetic patients—the long-acting GLP-1RAs dulaglutide and injectable semaglutide [7]. However, accumulating evidence suggests that SGLT-2i inhibitors may favorably influence the course of atrial fibrillation and thus protect against the cardioembolic ischaemic stroke subtype [8, 9], and the SGLT-2i canagliflozin may reduce the risk of hemorrhagic stroke [10]. Data on the impact of GLP-1RAs and SGLT-2i on chronic brain dyscirculation are even more limited.
A number of experimental studies have demonstrated the possibility of GLP-1RA liraglutide to decrease the brain damage volume and neurological deficit in animals both with DM and without it [11, 12, 13, 14]. At the same time neuroprotective effect of long-acting GLP-1RA dulaglutide in diabetic animals undergoing acute ischemic-reperfusion injury is not fully elucidated, as well as experimental data concerning potential influence of SGLT-2i on stroke are very limited. To our knowledge, no experimental studies have directly compared the neurotropic potential of GLP-1RA and SGLT-2i in diabetic animals with stroke.
Furthermore, despite the growing interest in the potential of novel glucose-lowering drugs to protect the central nervous system (CNS), the potential mechanisms of this effect remain not fully elucidated, both in the conditions of acute and chronic brain damage. One of the pathogenetic pathways of neuroprotection, in addition to direct effects on neurons, may be the influence on microglial activation and microglial damage [15].
Therefore, the aim of our study was to compare the neuroprotective properties of GLP-1RAs with different duration of action, liraglutide and dulaglutide, and SGLT-2i canagliflozin in acute and chronic brain injury in diabetic Wistar rats and to evaluate potential mechanisms, including microglial involvement. We chose the low-selective SGLT-2i canagliflozin for our study because it affects not only the second type of sodium-glucose cotransporter, but also the first type expressed in the CNS [16], and therefore this drug may have a potential neuroprotective property.
DM was modelled in male Wistar rats using high-fat diet + nicotinamide/streptozotocin model as described in our previous works [17]. Wistar rats (body weight 220–250 g) underwent acclimatization period (14 days) during which animals’ activity, skin and mucus membranes condition, tail position and other behavioral and vital parameters were evaluated. Rats not having any abnormalities were included into the further experiment. From the end of acclimatization period and up to the end of the experiment animals were kept on a high-fat diet (22% of saturated fats).
Four weeks after the start of high-fat diet nicotinamide solution (Sigma-Aldrich, Milwaukee, WI, USA) 230 mg/kg was injected i.p., followed by streptozotocin (Sigma-Aldrich, Milwaukee, WI, USA) solution i.p. injection 60 mg/kg 15 min later. Glycemia was evaluated by tail vein puncture using glucometer AccuCheck Performa (Roche, Basel, Switzerland) on the 2nd and 3rd days after nicotinamide/streptozotocin injection. Animals whose glycemic level, measured twice two different days, reached 11.1 mmol/L or more, were considered diabetic [18, 19]. If lower glycemia was found at least once, an oral glucose tolerance test (OGTT) was made. Glycemia was determined fasting and 15, 30, 60 and 90 min after 40% glucose solution 3 g/kg intake using a gastric tube. When at any measurement point during OGTT glycemia was 11.1 mmol/L or more diabetes was confirmed, if hyperglycemia was less prominent the diagnosis was not confirmed, and the data from there rats were no longer taken and analyzed.
Four weeks after nicotinamide/streptozotocin injection animals were divided into the following groups (n = 15 each):
• “DM” — rats with DM that remained untreated
• “DM+CANA” — rats with DM that were administered canagliflozin 25 mg/kg per os once daily
• “DM+LIRA” — rats with DM that were administered liraglutide 0.06 mg/kg s.c. once daily
• “DM+DULA” — rats with DM that were administered dulaglutide 0.12 mg/kg s.c. every 72 hours
Since the time nicotinamide/streptozotocin were infected (4th week of the experiment) and up to the end of the study we measured blood glucose level (BGL) using AccuCheck Performa glucometer as described above every week (daytime measurement, not a fasting one).
The certain treatment continued for 8 weeks, while animals in “DM” group remined untreated for the same period of time.
Besides the “Control” group was formed (n = 15)—neither DM modelling nor any kind of treatment was performed in these animals, they received standard chow. Glycemia was also measured in this group every week.
If during the experiment any rat developed symptoms typical for absolute insulin insufficiency (extensive weight loss, polyuria) it was dropped out.
After 8 weeks 10 animals of each group underwent stroke modelling while 5 animals were subjected to euthanasia with the following brain extraction for immunohistochemical assay. The experiment design is demonstrated in Fig. 1.
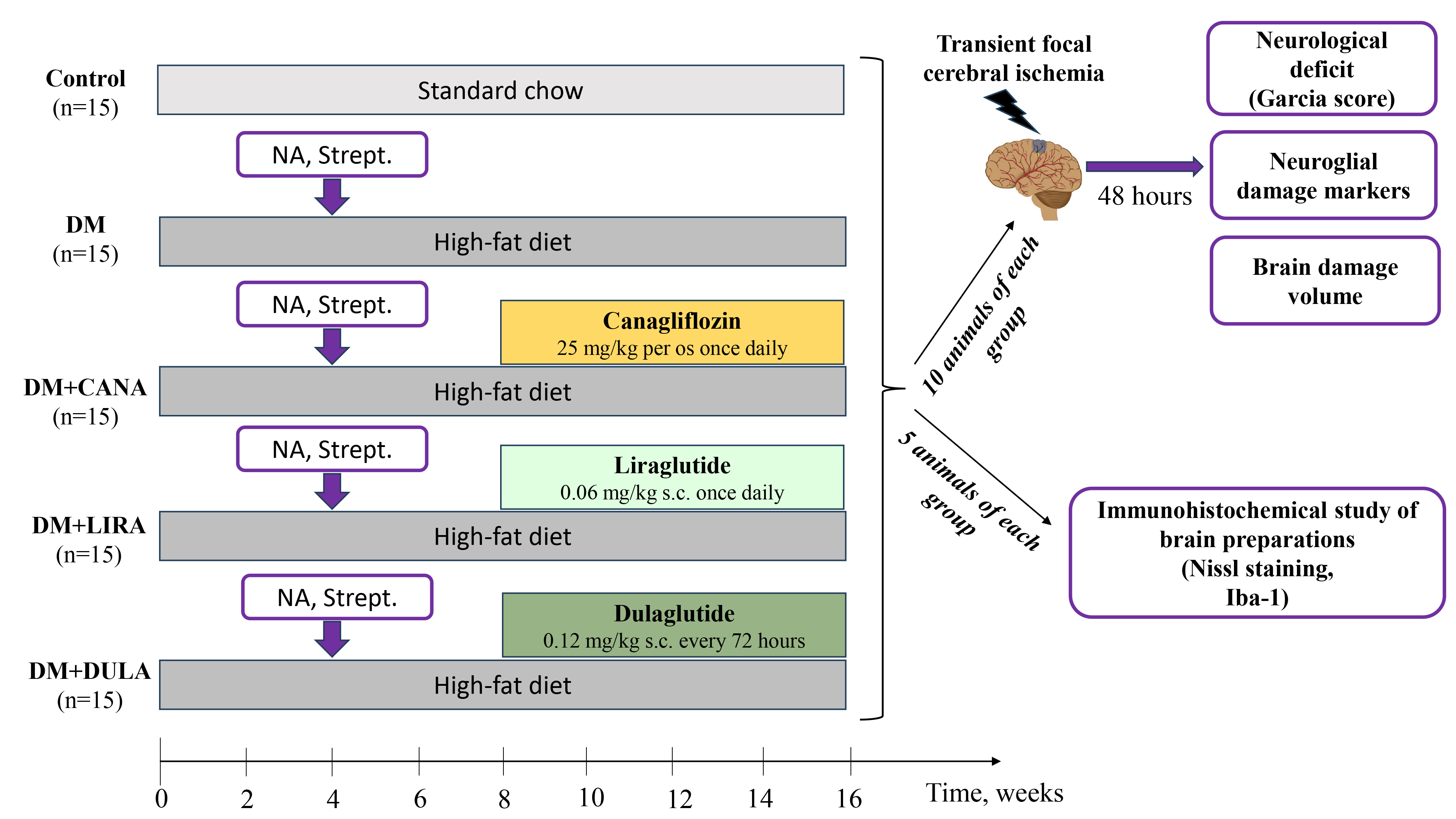 Fig. 1.
Fig. 1.Study design. After 4 weeks of high-fat diet male Wistar rats were subjected to nicotinamide 230 mg/kg and streptozotocin 60 mg/kg i.p. injection. Four weeks later rats were divided into groups “DM”, “DM+Canagloflozin (CANA)”, “DM+Liraglutide (LIRA)”, “DM+Dulaglutide (DULA)” (n = 15 each) to receive respective treatment for the following 8 weeks. Rats in “Control” group were fed with standard chow and did not receive medications. In 16 weeks of study 10 animals of each group were subjected to 30-min filament middle cerebral artery occlusion. After 48 hours of reperfusion neurological deficit using Garcia J.H. scores was measured, blood samples for neuroglial markers were obtained and brain damage volume was calculated by staining in 1% triphenyltetrazolium chloride solution. Other 5 animals of each group were subjected to immunohistochemical study of brain preparations for normal and pathologic neurons count (Nissl staining) and for microglial activation (immunohistochemical reaction to Iba-1). NA, nicotinamide; Strept., streptozotocin; DM, diabetes mellitus.
Transient focal brain ischemia was modelled in anesthetized rats (Zoletil + Xylazine i.m.) using the Koizumi and Longa method—30-minute intravascular left middle cerebral artery (MCA) occlusion [20]. Briefly, the model in based on the insertion of a 20–22 mm long thread (Doccol Corporation, Sharon, MA, USA) into the internal carotid artery and moving it to the MCA mouth occluding it. This leads to the development of ischemia in the arterial pool which is the most common localization of ischemic stroke in people. An occlusion-induced regional blood flow decrease was confirmed by means of Doppler ultrasound (Minimax Doppler-K model NB, Saint-Petersburg, Russia). We used the trepanation window to reach the left MCA cortical branch. If Doppler showed the linear blood flow velocity worsening for 70% or more, in comparison with the baseline value, we confirmed ischemia development in the curtain pool. After the 30-minute ischemia the occluder was taken away and the reperfusion period started.
Throughout ischemia modelling and thereafter, until anesthesia ended, the animals’ rectal temperature was maintained at 37.0 °C by means of a thermostatic table.
After 48 hours, neurological status was measured in all animals with the help of Garcia scores [21]. This scale consists of sensory and motor parameters, that allow to distinguish neurological impairments in animals that were subjected to stroke modelling. In Garcia scores a normal animal gets 18 points and an animal with maximal neurological deficit gets 3 points.
After neurological status assessment, the animals were anesthetized and blood samples from caudal vein were taken, centrifugated, aliquoted, serum samples were frozen for measurement of neurofilament light chains (NLC) level (Enzyme-linked Immunosorbent AssayKitFor Neurofilament, Light Polypeptide (NEFL), Cloud-Clone corp., Katy, TX, USA) as a neuronal damage marker and S100BB (Enzyme-linked Immunosorbent AssayKitFor S100 CalciumBindingProtein (S100), Cloud-Clone corp., Katy, TX, USA) concentration as a glial damage marker. After euthanasia the brains were extracted for determining the brain damage volume. Five brain slices, 2 mm thick each, were put into 1% triphenyltetrazolium chloride solution (MP Biomedicals, Santa Ana, CA, USA) for 15 min at 37 °C and pH 7.4. Thereafter both sides were photographed with an Olympus C4000 digital camera paired with an MBS10 microscope (BioLab, Moscow, Russian Federation). The received images were processed using ImageJ version 1.54j (https://imagej.net/ij/) and Adobe Photoshop 8.0 software (San Jose, CA, USA), which allow calculating the necrosis zone area. Then, by summing all the results, the brain damage volume was calculated in relation to the total brain volume (in %).
The rest 5 animals in each group after 8-week treatment were euthanized; the
brain was taken away with the preservation of the pia mater and arachnoid mater,
fixed in zinc-ethanol-formaldehyde [22], dehydrated, and embedded in paraffin.
Histological and immunohistochemical studies were performed on 5-µm frontal
sections of the telencephalon at –3.36 mm
Statistical data analysis was performed using the IBM SPSS Statistics-22
software package (IBM, Chicago, IL, USA) and Statistica-10 (Statsoft, Tulsa,
OK, USA). Results were obtained with nonparametric methods. Intergroup
differences significance was measured by means of the Kruskal–Wallace and
Mann–Whitney nonparametric tests for independent samples, using nonparametric
ANOVA (a post-hoc pairwise multiple comparison of groups using the Dunn’s test).
All values are presented as “median [25%; 75%]”. Significance was reached at
p
All experimental manipulations were performed in accordance with the Guide for the Care and Use of Laboratory Animals (NIH publication No. 85–23, revised 1996) and the European Convention for the Protection of Vertebrate Animals used for Experimental and other Scientific Purposes. The study protocol was approved by Institutional Animal Care and Use Committee of Almazov National Medical Research Centre (Protocol Number PZ_22_2, Feb 16 2022). All efforts were performed to protect the laboratory animals and minimize their suffering throughout the study. The experiments complied with the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines (https://arriveguidelines.org/).
We succeeded in modeling DM in 57 out of 60 rats, while DM criteria were not reached in the 3 animals: their BGL during OGTT reached the maximum values of 10.1, 10.7 and 10.9 mmol/L respectively, therefore they were excluded from the further experiment. For this reason, we had to include 3 more animals to maintain the stated sample size. None of the rats developed clinical manifestations typical for absolute insulin insufficiency throughout the experiment.
The BGL dynamics are shown in Fig. 2. For ease of analysis, we plotted blood glucose levels for every other week. All the rats in the diabetic groups had similarly elevated glycemic levels in the beginning. The start of therapy with canagliflozin, liraglutide and dulaglutide was characterized by BGL decrease and the glycemic profile remained normal in all the treatment groups throughout the rest of the experiment, with no differences between groups. In addition, BGL in “DM+CANA”, “DM+LIRA” and “DM+DULA” groups did not differ from that in “Control” group.
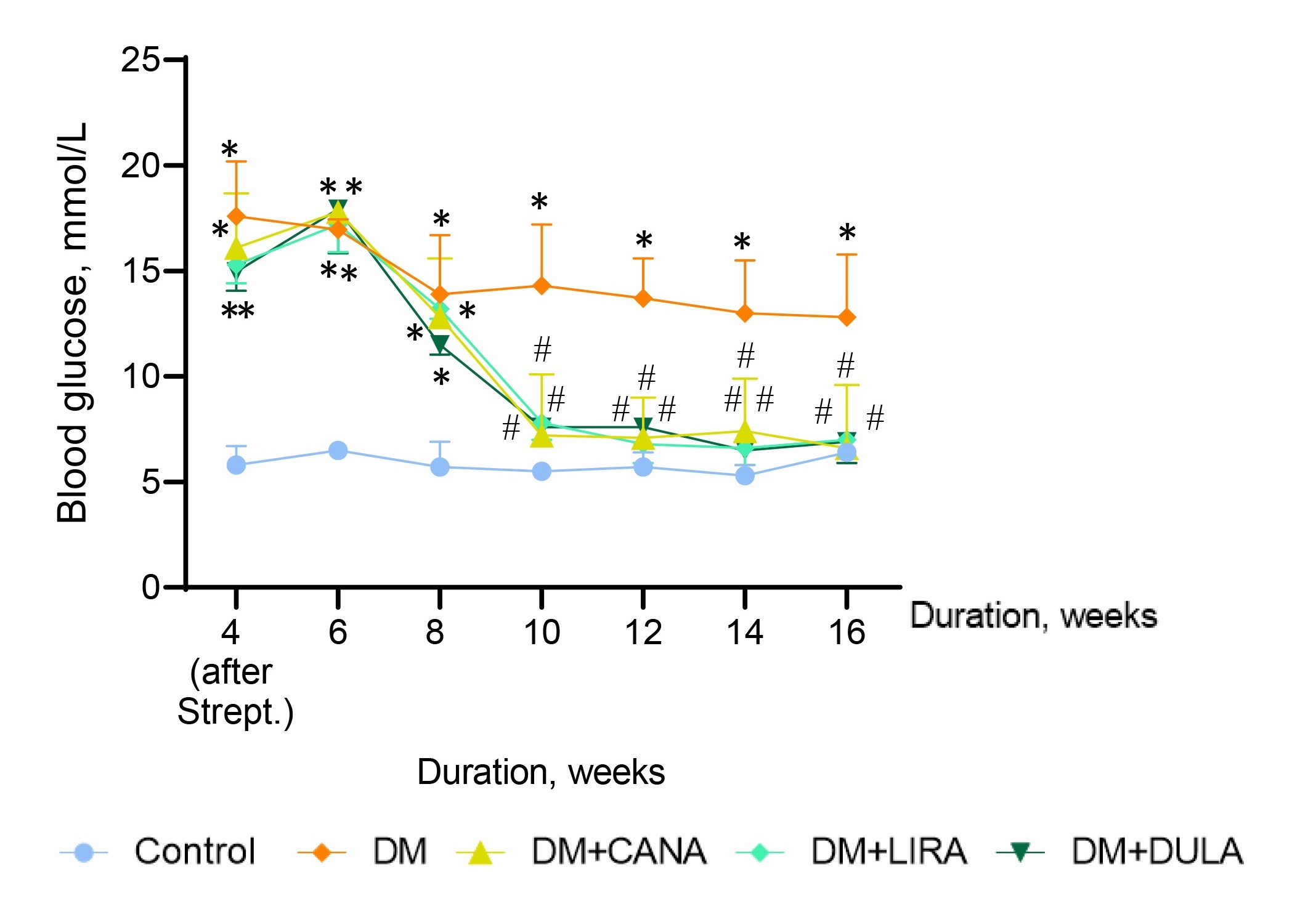 Fig. 2.
Fig. 2.Blood glucose level dynamics. *p
As mentioned above, neurological status 48 hours after acute cerebral ischemia was evaluated using the J.H. Garcia scale, that includes motor and sensory parameters and were the healthy animal receives 18 points and the most severe neurological deficit is described as 3 points.
Animals in “DM” group had more prominent neurological deficit than in “Control” (8.0 [4.0; 9.0] and 12.0 [10.0; 15.0] points, respectively, p = 0.02). All study treatments resulted in a significant improvement in neurological status. Thus, the neurological deficit in “DM+CANA”, “DM+LIRA” and “DM+DULA” was smaller than in “DM” (p = 0.000, p = 0.005 and p = 0.000, respectively) and even in “Control” group (except for “DM+DULA” where neurological deficit tended to be smaller than in “Control”, p = 0.077). Importantly, there were no between-group differences between treatment groups (Fig. 3).
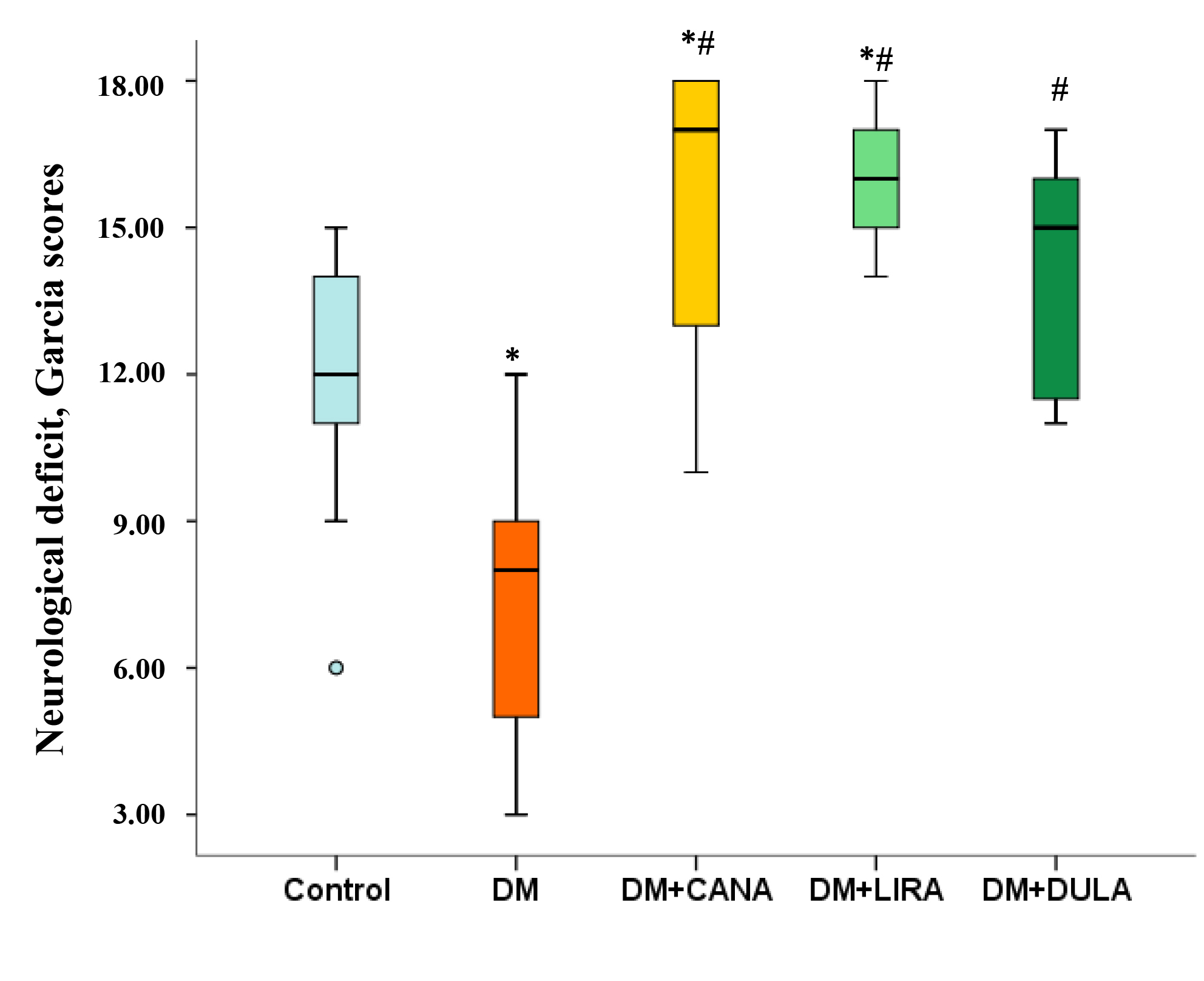 Fig. 3.
Fig. 3.Neurological deficit by Julio H. Garcia scale 48 hours after acute
brain ischemia. *p
Brain damage volume was similarly big in “Control” and “DM” groups (17.53 [14.23; 26.58] % and 15.87 [13.40; 22.68] % of total brain volume, respectively, p = 0.631). All the study drugs diminished brain damage volume comparing with “DM” and “Control”. Importantly, brain necrosis volume in “DM+LIRA” was smaller than in “DM+CANA” (2.9 [1.83; 4.71] % and 6.17 [3.88; 8.88] %, respectively, p = 0.015) and did not significantly differ from that in “DM+DULA” (2.9 [1.83; 4.71] % and 4.57 [3.27; 7.90] %, respectively, p = 0.165) (Fig. 4A,B).
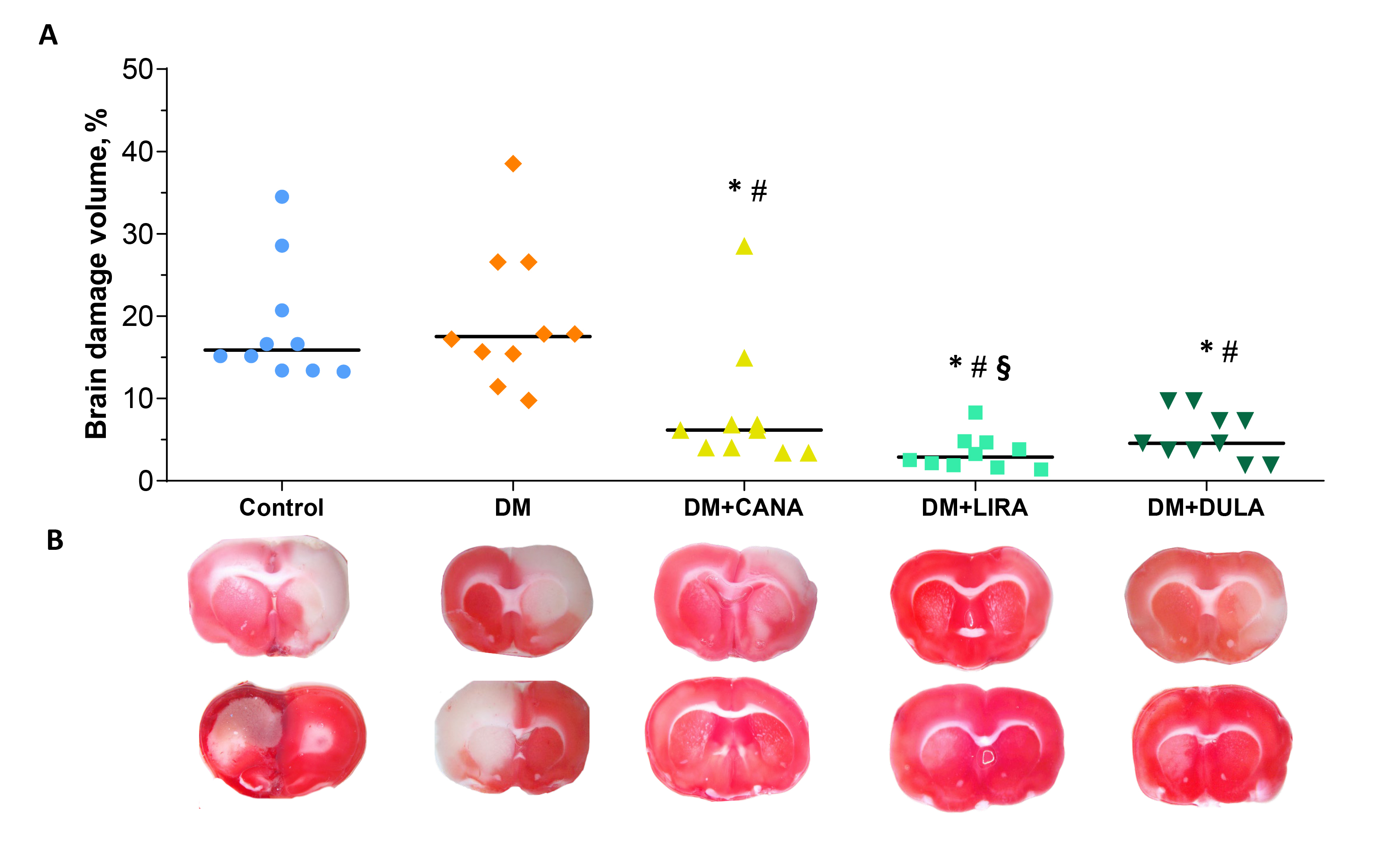 Fig. 4.
Fig. 4.Brain damage caused by ischemia-reperfusion injury. (A) Brain
damage volume measurement results presented as dot plots with median values. (B)
Representative images of brain slices stained with triphenyltetrazolium chloride.
*p
NLC level was similarly high in “Control” and “DM” groups (p = 0.963). Both the SGLT-2i canagliflozin and the long-acting GLP-1RA dulaglutide were associated with a decrease in this parameter (p = 0.029 comparing “DM+CANA” and “DM”, p = 0.026 comparing “DM+DULA” and “Control”). However, liraglutide had no effect on NLC concentration (Fig. 5A).
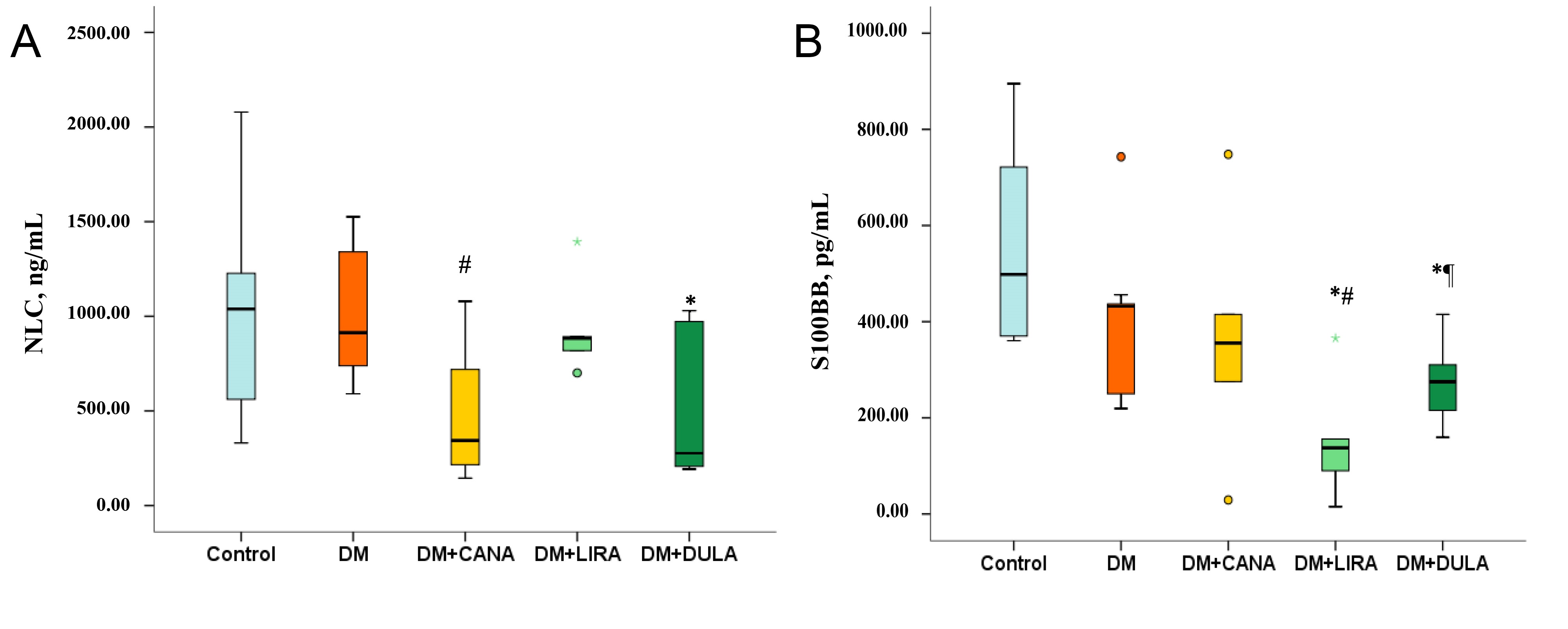 Fig. 5.
Fig. 5.Neurofilament light chains and S100BB protein concentrations 48
hours after ischemia-reperfusion injury. (A) Neurofilament light chains (ng/mL)
level, (B) S100BB protein (pg/mL) level. *p
Similarly, S100BB levels were high in both “Control” and “DM” with no differences between the groups (p = 0.113). Both GLP-1RAs caused a decrease in S100BB compared to “Control”, although the effect of liraglutide was significantly more pronounced than that of dulaglutide (p = 0.035). On the other hand, canagliflozin did not influence S100BB concentration (Fig. 5B).
A study of rat brain paraffin sections stained with toluidine blue using the Nissl method showed that in “Control” group neurons in the CA1 hippocampal zone were arranged densely in several rows, forming one layer of cells called the stratum pyramidale. These cells had a light, large nucleus and 1–2 metachromatically colored nucleoli. A uniformly distributed chromatophilic substance was clearly visible in the cytoplasm of the neuronal cell bodies. The initial segments of neuronal dendrites were often visible, extending into the stratum radiatum (Fig. 6A).
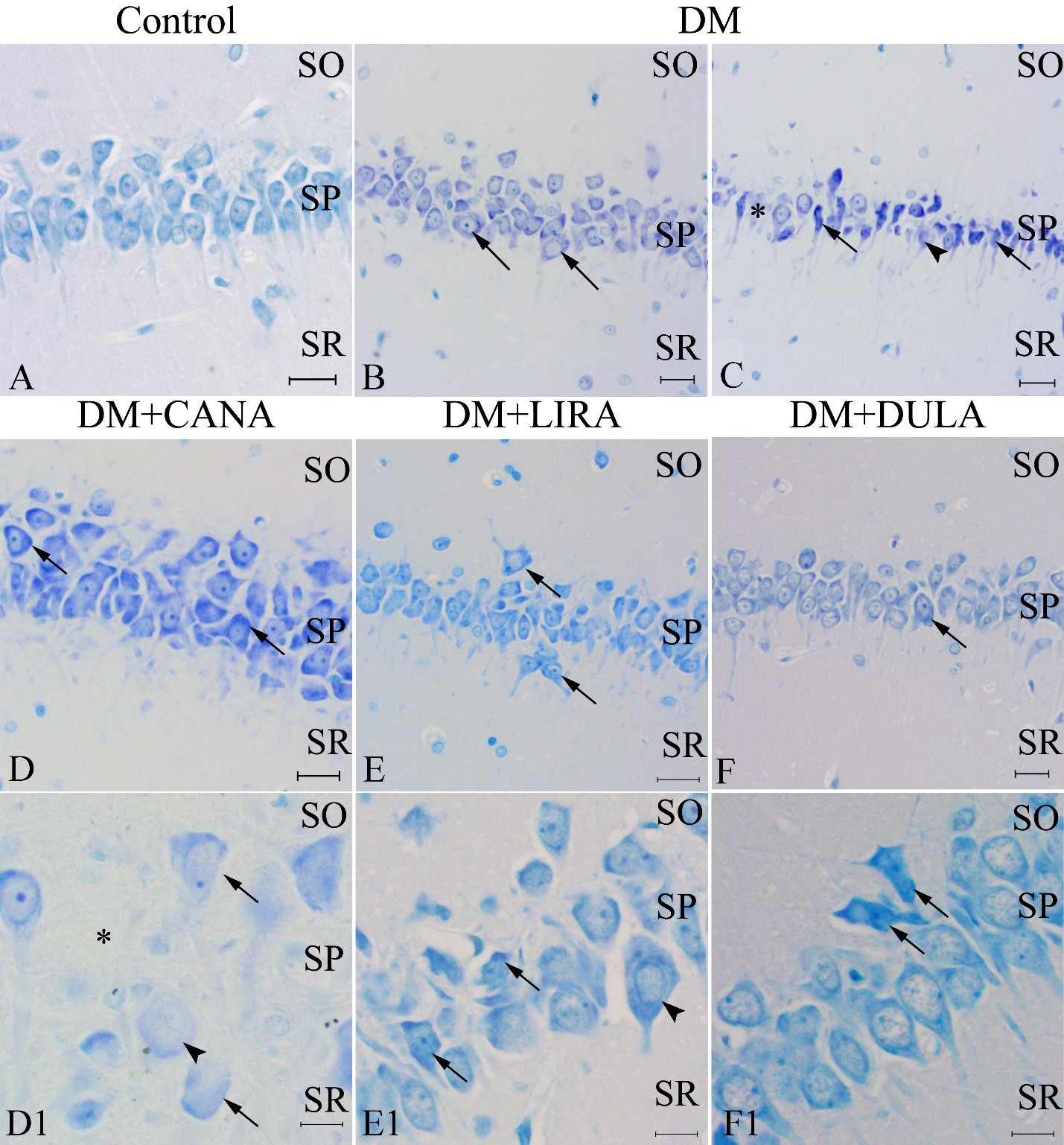 Fig. 6.
Fig. 6.Neurons of Cornu Ammonis (CA)1 zone of rat hippocampus. Nissle staining. “Control” group (A); diabetes mellitus, “DM” (B,C); diabetes mellitus + canagliflozin, “DM+CANA” (D,D1), diabetes mellitus + liraglutide, “DM+LIRA” (E,E1); diabetes mellitus + dulaglutide, “DM+DULA” (F) (D,E,F — lower magnification, D1,E1,F1 — higher magnification). Arrows point at expanded cells (B,D1), shriveled neurons (C,F1), hyperchromatosis (D,F,E1), neurons outside the pyramidal layer (E). Arrowheads point at hypochromatosis (C,D1) and expanded cell (E1). Asterisk indicates shadow cells (C) and empty area between neurons (D1). Scale bar 20 µm (A–F) and 5 µm (D1–F1). In the “Control” group neurons in the CA1 hippocampus zone were arranged densely in several rows, forming one layer of cells called the stratum pyramidale. These cells had a light, large nucleus and 1–2 metachromatically colored nucleoli. In the “DM” group there were hypo- and hyperchromatic, swollen, wrinkled neurons; in the pyramidal layer of CA1 zone neurons number was decreased, neurons located in several rows alternated with neurons located in 1–2 rows, areas were observed where neurons were absent and the pyramidal layer was interrupted. In the “DM+CANA” group, there were hypo- and hyperchromatic nuclei throughout the entire studied area, wrinkled neurons were less common than in “DM” group. In the “DM+LIRA” most neurons had normal morphology but there were cells with signs of hyperchromatosis, often with polygonal shape. In “DM+DULA” group most cells did not have pathological changes. There were single wrinkled neurons and cells with signs of hyperchromatosis or swelling. SO, stratum oriens; SP, stratum pyramidale; SR, stratum radiatum.
In the “DM” group hypo- and hyperchromatic neurons, as well as swollen (or edematous) neurons, were identified (Fig. 6B,C). Throughout the CA1 zone a large number of wrinkled neurons were observed – cells with pronounced signs of hyperchromatosis, uneven borders and a decrease in the cytoplasm and cell nuclear volume (Fig. 6C). There were also cells with cleared cytoplasm and the absence of chromatophilic substance (Fig. 6C, arrowhead). Rarely, shadow cells were found - cells in which toluidine blue staining was almost completely absent both in the cytoplasm and in the nucleus (Fig. 6C, asterisk). In the “DM” group, a decrease in the number of neurons was often observed in the pyramidal layer of the CA1 zone - neurons located in several rows could alternate with neurons located in 1–2 rows, while areas were observed where neurons were absent and the pyramidal layer was interrupted.
In the “DM+CANA” group, similar to the “DM” group, neurons were characterized by the presence of hypo- and hyperchromatic nuclei throughout the entire examined area (Fig. 6D). In the “DM+CANA” group wrinkled neurons were less common than in the “DM” group. Swollen cells and shadow cells were also found (Fig. 6D1).
In the “DM+LIRA” group most neurons had normal morphology. However, there were cells with signs of hyperchromatosis, often these cells had a polygonal shape. Individual cells showed signs of swelling. Rarely individual neurons or groups of neurons were located outside the pyramidal layer (Fig. 6E,E1).
In both the “DM+DULA” and the “DM+LIRA” groups, the vast majority of cells did not show pathological changes. There were individual wrinkled neurons and cells with signs of hyperchromatosis or swelling (Fig. 6F,F1).
Statistical analysis showed that the number of neurons with normal unaltered structure was lowest in the “DM” group. Administration of all the study drugs was associated with an increased number of normal cells while higher level was seen in the “DM+LIRA” and “DM+DULA” groups with no difference between them (Fig. 7A). The number of pathological cells was mostly similar in all the groups (Fig. 7B). We also calculated the ratio of normal to pathological cells and found that animals in the “DM” group had the lowest ratio, while all study drugs increased this ratio and the “DM+DULA” group was even characterized by a normal/pathological cell ratio similar to that in the “Control” group (Fig. 7C).
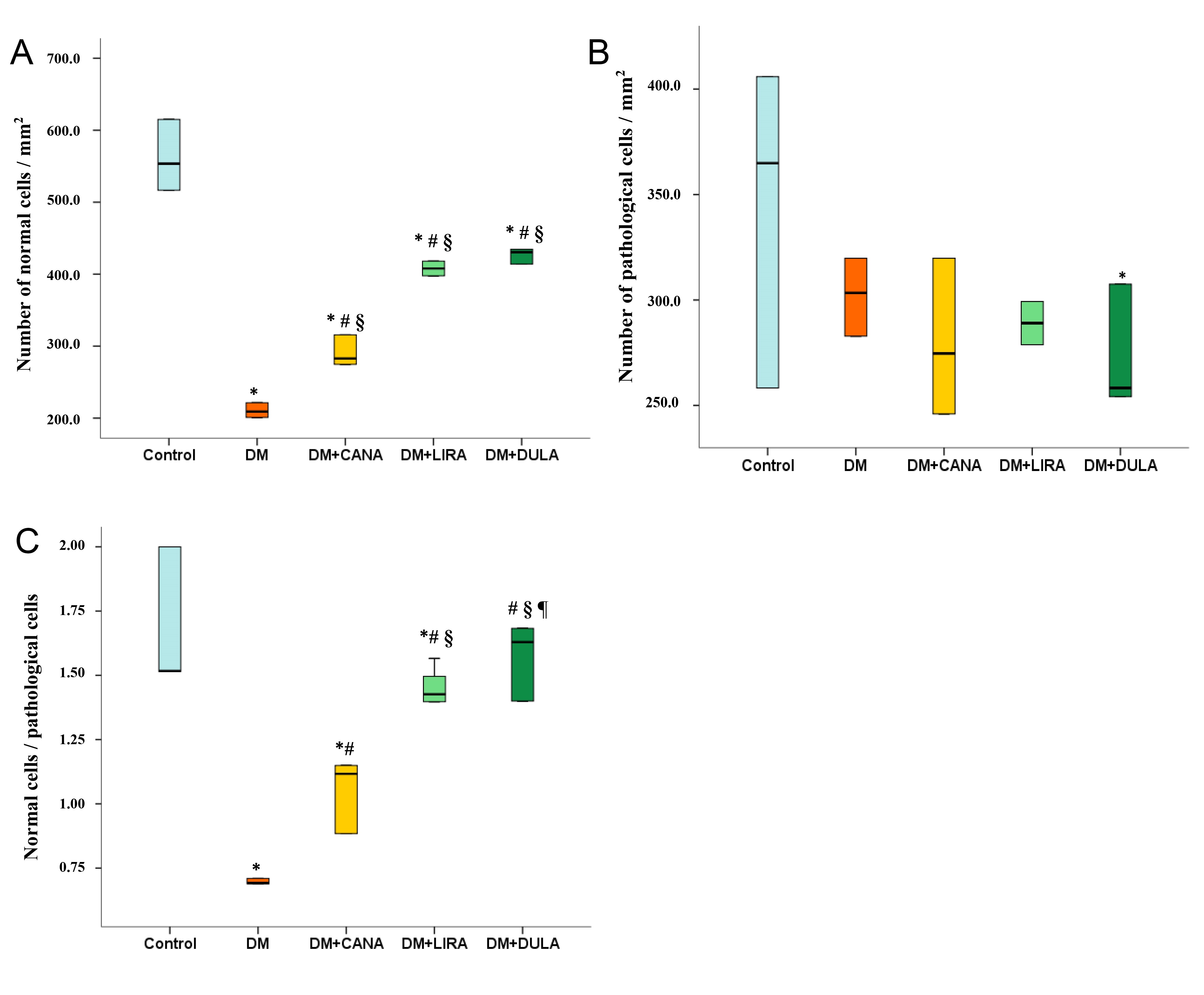 Fig. 7.
Fig. 7.Number of normal, pathological cells and normal/pathological
cells ratio of CA1 zone of rat hippocampus, Nissl staining. (A) number of normal
cells/mm
An immunohistochemical study of microglial cells in the CA1 hippocampus zone in the stratum radiatum layer revealed that in the “Control” group microglia were represented by the ramified type (Fig. 8, Control). These cells had a small cell body (and therefore cell bodies were rarely visible) and long, thin branching processes. In “DM” group microglia were characterized by a large cell body and often short and thick processes suggesting that microglia here were represented by activated forms (Fig. 8, DM). The number of microgliocytes also increased dramatically. In the “DM+CANA” group microglia were characterized by transitional forms. Cell bodies were small, but larger in size compared to “Control” group. The processes were long and strongly branched, slightly thicker compared to the “Control” (Fig. 8, DM+CANA). The number of microgliocytes was comparable to “DM” group. In the “DM+LIRA” and “DM+DULA” groups microglia were of the ramified type (Fig. 8, DM+LIRA, DM+DULA). The number of cells in the “DM+DULA” group was comparable to the “Control” group.
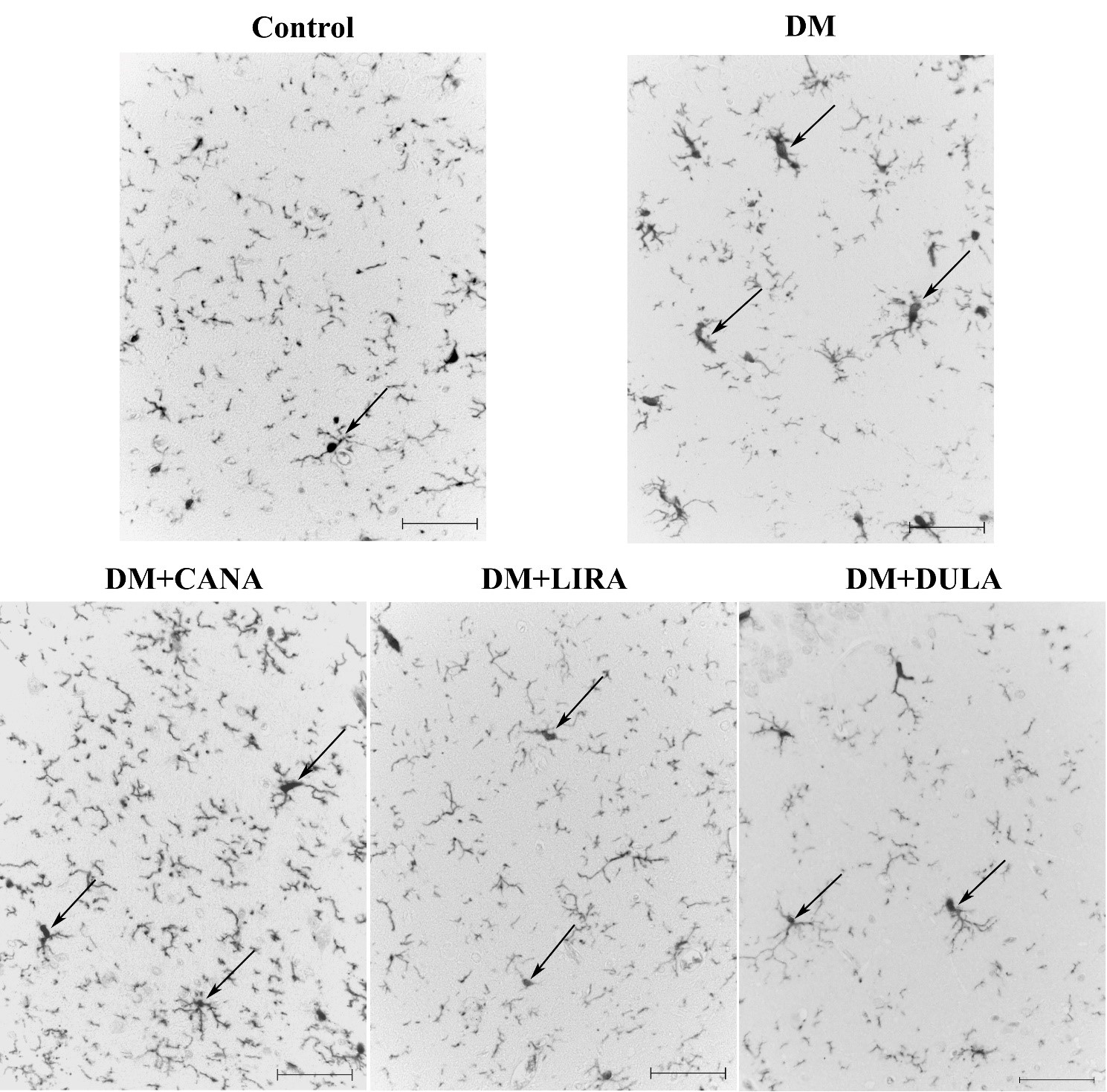 Fig. 8.
Fig. 8.Microglia of stratum radiatum of rat hippocampal CA1 zone. Immunohistochemical reaction to Iba-1. Effects of DM (diabetes mellitus), canagliflozin (DM+CANA), liraglutide (DM+LIRA) and dulaglutide (DM+DULA) on microglia activation. Arrows point at Iba-1+-cells. Scale bar 20 µm. In the “Control” group microglia were represented by the ramified type characterized by a small cell body and long, thin branching processes. In the “DM” group microglia were characterized by a bigger number of cells, larger cell body, short and thick processes typical for activated forms. In the “DM+CANA” group microglia were characterized by transitional forms. In the “DM+LIRA” and “DM+DULA” groups microglia were of the ramified type.
Statistical analysis confirmed the data described above: DM was characterized by an increased number of immunopositive cells. Canagliflozin treatment did not decrease the number of immunopositive cells compared to the “DM” group. On the other hand, both GLP-1RAs reduced the number of activated microgliocytes compared to the “DM” group, with dulaglutide being more effective than liraglutide (Fig. 9).
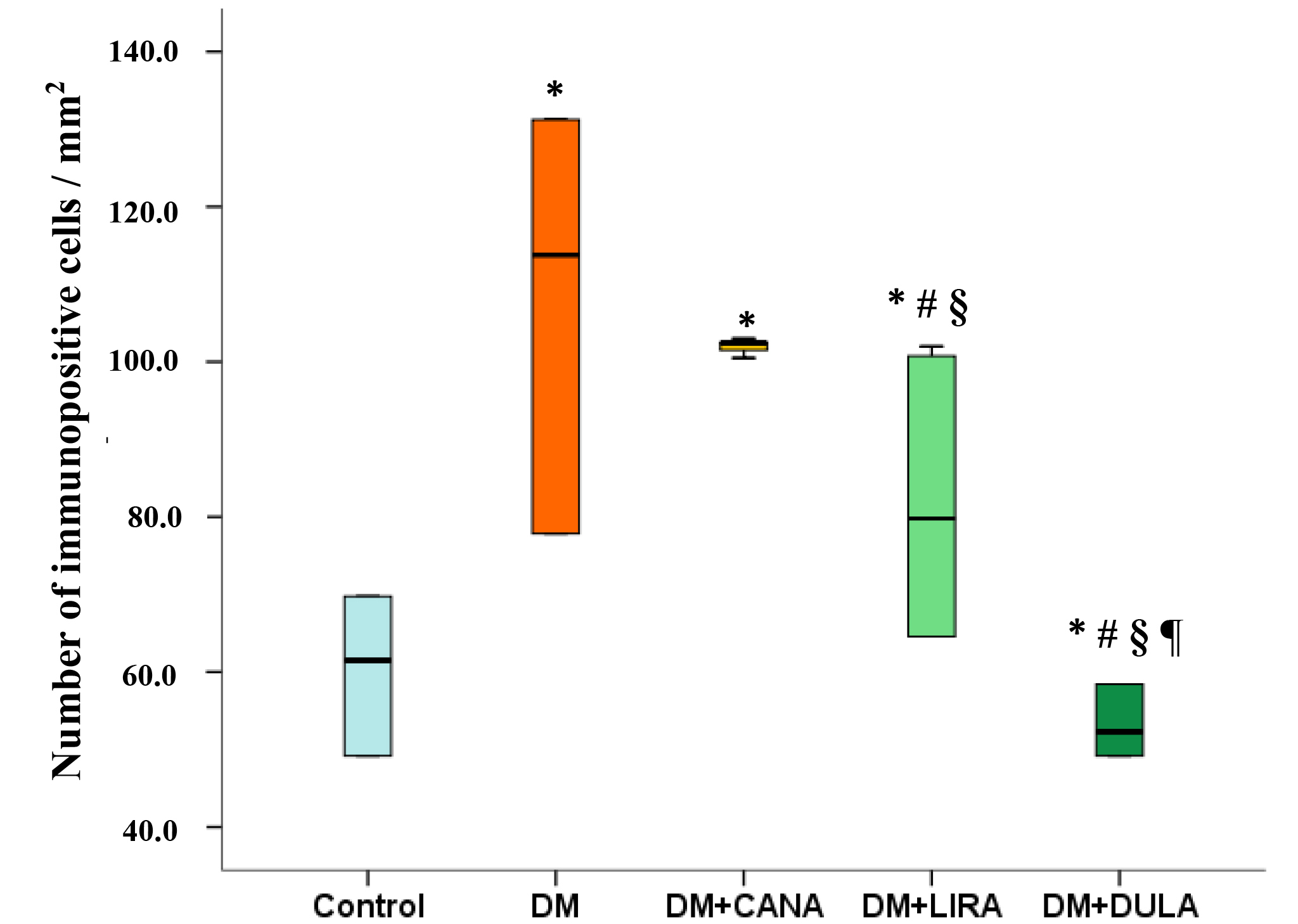 Fig. 9.
Fig. 9.Number of immunopositive cells (activated microgliocytes) in
stratum radiatum of rat hippocampal CA1 zone, immunohistochemical reaction to
Iba-1. *p
Our study was aimed to investigate the influence of diabetes mellitus on the severity of brain damage in conditions of acute and chronic brain ischemia as well as to hold the direct comparison of GLP-1RAs and SGLT-2i neuroprotective properties under these conditions.
In clinical practice, DM is known to be a major cause of acute cardiovascular accidents such as myocardial infarction and stroke, leading to significant disability or even death much more frequently than in the general population [24]. Surprisingly, the experimental data are not so unambiguous. Thus, it has been shown that the infarction area in diabetic animals was even smaller than in nondiabetic animals [25]. The explanation for such a phenomenon could be some kind of metabolic preconditioning that takes place in hyperglycemic conditions. Conversely, there are a number of experimental studies showing an outstanding negative impact of hyperglycemia in organ damage volume, including myocardial infarction and stroke, and poor outcome [26, 27, 28]. In our experiment brain damage volume was similar in untreated diabetic and non-diabetic animals, whereas the neurological deficit was more pronounced in diabetic animals, implying a worse recovery after stroke and therefore potentially a worse outcome.
To compare the neuroprotective effects of glucose-lowering drugs, we chose liraglutide and dulaglutide as two GLP-1RAs with different durations of action. According to RCTs, liraglutide has demonstrated the potential to reduce all-cause mortality, cardiovascular mortality and the incidence of myocardial infarction, without a certain impact on the risk of stroke. In contrast, dulaglutide is one of the long-acting GLP-1RAs that can reduce the risk of non-fatal stroke [29]. Interestingly, there is a large body of experimental work focusing on the neuroprotective effect of liraglutide in both diabetic and non-diabetic rats, used before or after stroke [11, 12, 14, 30]. Thus, liraglutide has demonstrated anti-ischemic properties in experimental conditions. On the contrary, we have not found any data on the neuroprotective potential of dulaglutide in stroke conditions in diabetic rats. Part of the rats in our experiment received canagliflozin as a non-selective SGLT-2i, which means that this drug suppresses not only the second type of sodium-glucose cotransporter, which is mainly found in the kidneys, but also the first type, which is widely expressed in the CNS and vascular endothelium [16]. In this context, as mentioned above, canagliflozin could be a perspective drug to realize neuroprotective properties. Surprisingly, although there are some publications on the neuroprotective properties of other SGLT-2i, for example the highly selective empagliflozin [31, 32], we could not find any work on the effect of canagliflozin in transient brain ischemia.
To our knowledge, our study is the first direct comparative investigation of the neuroprotective properties of GLP-1RAs and SGLT-2i in acute brain ischemia in diabetic rats.
Previously, our group performed a similar direct comparative study of SGLT-2i and GLP-1RA neuroprotective effects in stroke, but in non-diabetic conditions [33]. Such a model of non-diabetic rats undergoing stroke helps to phenomenologically confirm the presence of neuroprotective potential of some substances - in this situation, glucose-lowering drugs.
The current study held in diabetic rats helps to mimic the conditions close to the real-life ones. We succeeded in reaching similarly satisfactory glycemic profile in all treatment groups so that glycemic level did not differ significantly between the canagliflozin, liraglutide and dulaglutide treatment groups. We believe this allows us to analyze pleotropic neuroprotective effects of these drugs without strong association with glycemic control.
We found out that canagliflozin, liraglutide and dulaglutide all reduced the brain damage volume and neurological deficit in transient ischemia-reperfusion injury. Although all three drugs had a similar beneficial effect on neurological status, the infarct-limiting property of liraglutide was more pronounced than that of canagliflozin and did not differ from that of dulaglutide, suggesting that GLP-1RAs may be more protective than SGLT-2i in acute ischemic stroke. However, these data need to be investigated further to understand whether this is a drug-effect or a class-effect.
A possible explanation for the more pronounced infarct-limiting property of GLP-1RAs may be related to the broader expression of GLP-1 receptors in the CNS than that of SGLT-2. For example, GLP-1 receptors are found in the frontal cortex, thalamus, hypothalamus, hippocampus, cerebellum and substantia nigra–regions known to be involved in the regulation of energy homeostasis and autonomic functions [34]. Moreover, GLP-1 receptors are also present in the dorsal vagus complex, particularly in the nucleus of the solitary tract, and to a lesser extent in the subfornical organ and area postrema [35]. GLP-1RA are known to realize their protective effect in ischemic conditions through several ways including activation of anti-apoptotic genes and suppression of proapoptotic ones [36]. Anti-ischemic properties of SGLT-2i are much less studied and the specific mechanisms require further investigation. Nevertheless, low-selective SGLT-2i might by more promising regarding ischemic conditions as this is SGLT-1 activation during stroke that realizes proinflammatory action, increases ischemia-reperfusion injury and decreases neuronal survival [16].
Ischemia-reperfusion injury in our study was associated with increase in neurofilament light chains independently on glycemic profile which resulted in similar increase of this parameter in both control group and untreated diabetic group. NLC are the proteins comprising predominantly large-caliber myelinated axons and are the part of a complex called neurofilament [37]. Increased levels of NLC can be observed in biological fluids, including blood, in various conditions associated with nervous system damage, such as neurodegenerative diseases and traumatic brain injury [38]. It has been reported that serum NLC level correlates with the infarct volume in patients with acute ischemic stroke [39] and can be a biochemical predictor of unfavorable outcome and secondary neurodegeneration development after stroke [40]. In our study NLC level was lower in animals treated with canagliflozin and long-acting GLP-1RA dulaglutide. Surprisingly, liraglutide did not have similar action. The positive effect of both SGLT-2i and long-acting GLP-1RAs on NLC levels suggests that these drugs have a direct effect on neuronal tissue survival.
Another potential mechanism involved in ischemic stroke may be related to microglial damage. Microglia are resident CNS immune cells that act as critical sensors, effectors and regulators of CNS homeostasis under normal and pathological conditions [41]. In acute damage such as ischemic stroke microglia play a crucial role in a number of pathophysiological pathways leading to the activation of complement system and reactive oxygen species release. During the course of stroke, microglia undergo severe morphological changes and participate in neuroinflammation and at the same time in the regeneration process, thus realizing both alternating and beneficial effects [42].
To assess the microglial component of the damage, we measured serum levels of
S100BB. S100 are the brain glial proteins produced mainly by astrocytes. Cerebral
S100 is a combination of two closely related family proteins: S100A1
(S100
We also investigated the CNS damage in the conditions of chronic brain ischemia using DM model itself assuming that long-standing diabetes is characterized by the kind of CNS lesion by the mechanism of “silent” strokes as described earlier. Immunohistochemical assay approved that DM is characterized by pathological changes in the neurons. All the study drugs administration was associated with increased number of cells with unchanged structure, while GLP-1RAs had more pronounced effect, without differences between liraglutide and dulaglutide. This finding suggests direct positive influence of SGLT-2i and especially GLP-1RAs on neurons structure. Besides, we revealed pathological microglial activation in DM animals which was diminished by both GLP-1RAs with dulaglutide being more effective, whereas SGLT-2i canagliflozin did not realize this effect.
Thus, it can be assumed that microglial damage and pathological hyperactivation are involved in the pathogenesis of both acute and chronic brain damage. GLP-1RA can diminish microglial reaction while protective effect of SGLT-2i lies aside from the influence on microglia.
We can conclude that ischemic stroke course might be more severe in the presence of DM which represents in worse neurological outcome. Brain ischemia independently on the presence of DM is accompanied by both neuronal and glial damage. In acute ischemia-reperfusion injury both GLP-1RAs and SGLT-2i are neuroprotective while GLP-1RAs pleotropic effect might be more prominent as this group of drugs has more pronounced infarct-limiting effect. Microglia damage and pathological activation are parts of acute and chronic brain damage pathogenesis. We can suggest that both GLP-1RAs and SGLT-2i in acute stroke and in chronic brain damage have direct positive influence on neuronal structure and survival while only GLP-1RAs also decrease microglial damage and pathological hyperactivation.
All datasets on which the conclusions of a manuscript depend are included in the article in the form of graphic material including individual values or boxplots. Additional information is available from the corresponding author on reasonable request.
DK, TV and TK designed the research study. AS, OF, NT, AI, DS and OK performed the research. AS, OF, DS and OK analyzed the data. AS and DS wrote the manuscript. All authors contributed to editorial changes in the manuscript, read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All experimental procedures were performed in accordance with the Guide for the Care and Use of Laboratory Animals (NIH publication No. 85–23, revised 1996) and the European Convention for the Protection of Vertebrate Animals used for Experimental and other Scientific Purposes. The study protocol was approved by Institutional Animal Care and Use Committee of Almazov National Medical Research Centre (Protocol Number PZ_22_2, Feb 16 2022). All efforts were performed to protect the laboratory animals and minimize their suffering throughout the study. The experiments complied with the ARRIVE guidelines (https://arriveguidelines.org/).
Not applicable.
This research was funded by the Ministry of Health of Russian Federation, agreement no. 124021600064-8.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.