
 , Hajra Zafar 3, Lei Xing 4, Qiaoyun Li 1, Ran Li 5, Shanshan Xu 6,*
, Hajra Zafar 3, Lei Xing 4, Qiaoyun Li 1, Ran Li 5, Shanshan Xu 6,* , Dan Chen 7,*
, Dan Chen 7,*1 Department of Neurology, Nanjing Gaochun Hospital of Traditional Chinese Medicine, 211300 Nanjing, Jiangsu, China
2 Department of Surgical, Nanjing Gaochun Hospital of Traditional Chinese Medicine, 211300 Nanjing, Jiangsu, China
3 Department of Pharmacy, School of Pharmacy, Shanghai Jiao Tong University, 200240 Shanghai, China
4 Department of Geriatrics, Nanjing Gaochun Hospital of Traditional Chinese Medicine, 211300 Nanjing, Jiangsu, China
5 Department of Pharmacy, School of Pharmacy, Jiangsu University, 212013 Zhenjiang, Jiangsu, China
6 Department of Neurology, People's Hospital of Gaochun, 211300 Nanjing, Jiangsu, China
7 Department of Rehabilitation Medicine, Xuancheng People's Hospital, 242000 Xuancheng, Anhui, China
Abstract
Background: Severe neurological condition like Alzheimer’s disease (AD)
has a significantly negative impact on families and society, wherein there is no
proven cure. As one of the principal active constituents of Achyranthes
bidentata Blume, ecdysterone (ECR) has demonstrated antioxidant and cognitive
dysfunction improvement effects. Nonetheless, the mechanism underlying the
improvement of cognitive dysfunction by ECR remains unclear. This study sought to
ascertain whether ECR may allebviate cognitive impairment by reducing oxidative
stress via activation of the nuclear factor erythroid-2-related factor-2 (Nrf2) antioxidant system through Akt/GSK3
Keywords
- Alzheimer's disease
- ecdysterone
- oxidative stress
- Nrf2
- Akt/GSK3
One of the most predominant disorders of the central nervous system that poses significant problems within the realm of global public health is Alzheimer’s disease (AD), which is a neuro-degenerative condition that is associated with cognitive and memory loss [1]. Amyloid plaques and intracellular neurofibrillary tangles (NFT), two significant neuropathological signs, are its defining characteristics [2, 3]. As the number of AD patients rises, their families experience considerable misery, and society as a whole is heavily burdened socioeconomically [4]. Currently, the fight against AD is neurology’s most pressing unmet medical need. For a long time, there has been no specific cure for AD, only a comprehensive approach to controlling the progression of the disease [5]. Therefore, it is critical in the realms of public health to develop effective treatment interventions for AD.
The pathophysiology of AD is heavily influenced by oxidative stress [6]. It
contributes to A
The cellular antioxidative response is thought to be mainly regulated by the
nuclear factor erythroid-2-related factor-2 (Nrf2) [10]. Normally, Nrf2 is
localized in cytoplasm, but it is translocated to the nucleus after exposure to
oxidatively stress conditions, where it activates genes that protect against
oxidation [11]. Due to Nrf2’s critical role in neuroprotection in AD, its
deletion or mutation worsens memory loss, cognitive decline, and A
In traditional Chinese medicine, the roots of the Amaranthaceae plant
Achyranthes bidentata Blume are frequently used to treat dementia [20].
Ecdysterone (ECR) is one of the main active ingredients of Achyranthes
bidentata Blume and its discovery has increased medicinal value while its
antioxidant activity has been reported [21]. Additionally, earlier research has
demonstrated that ECR can enhance rat C-fos expression, alleviate cognitive
impairment brought on by oral administration of the
Based on available literature, it was postulated that ECR may regulate Nrf2 in
an Akt/GSK3
Abcam (Cambridge, UK) provided antibodies for A
Changzhou Cavins Laboratory Animal Co., Ltd. (Changzhou, China) provided the male mice, namely senescence accelerated-resistant mouse (SAMR1) and Senescence-accelerated mouse prone 8 (SAMP8), which were without any specific pathogen, 4 months old and weight 30–35 g. All the animals were fed in cages in the same quiet environment with a light (12 L)/dark (12 D) cycle, respectively fed and drank sterilized feed and water. The experiment was carried out after 7 days of adaptation. The Jiangsu University (UJS IACUC) institutional committee for the care and use of laboratory animals reviewed and approved (approval number: UJS-IACUC-2022031401) for studies involving animals.
One week later, 48 qualified animals were selected, and 40 SAMP8 mice except for 8 SAMR1 mice in the control group were randomly divided into 5 groups. All the animals were divided into six groups: SAMR1 blank group (Control), SAMP8 Model group (Model), SAMP8 model + ecdysterone Low dose group (Low), SAMP8 model + ecdysterone Medium dose group (Medium), SAMP8 model + ecdysterone High dose group (High), SAMP8 model + Donepezil group (Positive). Mice in the administration group were given ECR intragastric administration (high, medium and low doses were 5, 10, and 20 mg/kg/day, respectively) [24]. The control group and model group were intragastrically given 0.9% sodium chloride solution of the same volume, and the positive group was intragastrically given Donepezil (1 mg/kg/day). The drug treatment group and the control group were fed the same way in different cages. And continued administration for 4 weeks. Animals were sacrificed after the treatment, and tissues were collected and kept in a –80 °C freezer until further analysis.
One of the most popular cell lines for neuroscience research is PC12, which is
employed in investigations on synaptogenesis, neurotoxicity, neuroprotection,
neurosecretion, and neuroinflammation [25]. STR profiling was used to confirm the
PC12 Cell Line, and mycoplasma testing came out negative. Every cell was
cultivated at 37 °C and 5% CO
In brief, we placed the platform in the middle of the fourth quadrant, while an automatic camera system with a computer connection was put in place above the pool to monitor and record the swimming activities of the mice. The mice’s swimming distance and travel time to the platform (to avoid the incubation period) were automatically estimated. Mice were thrown into the water with their backs to the pool wall from one of the four starting places. We looked at the movement patterns within 90 s before timing the duration within which the mice found the platform. The experimenter took the mice to the platform, where they rested for 15 s before the next training when they could not find the platform inside 90 s (90 s was recorded as the escape incubation period). We trained the mice 4 times a day at a fixed time period. Within 4 times of training, the mice were put into water from 4 different quadrants, while the interval of each training was 1 min. After three days of continuous training, the formal experiment began. Eventually, we recorded the time taken by the mice from the time they were placed into the pool to the time they found the platform. Once a day for 5 consecutive days, we recorded the time to be 90 s when the time for the mice to find the platform exceeded 90 s.
After the navigation experiment, the mice in each group rested for one day, and then tested their memory ability through the space exploration experiment. Mice were added to the pool from the third quadrant after the platform in the fourth quadrant was removed. Each mouse group was observed and counted as they passed the platform in the fourth quadrant over the course of 90 s.
Paraffin sections of mouse hippocampal tissue were dewaxed and immersed in water, dyed with 1% toluidine blue for 10 min, and rinsed with distilled water. The colour is then separated in 70% alcohol for seconds to minutes. Then in anhydrous ethanol dehydration, xylene is transparent. Finally, seal it with a neutral glue and observe the cornu ammonis 1 (CA1) and CA3 regions of the hippocampus under a microscope.
After administration, hippocampal tissue was taken from mice and placed in 4%
(w/v) paraformaldehyde (PFA) solution overnight. Before cutting the tissue into
slices (4 µm), we embedded them in paraffin. Dewaxing of paraffin sections
to water was carried out, before antigen repairing, blocking and sealing as well
as overnight incubation at 4 °C with primary antibodies of A
Measurement of ROS levels in tissues: At the end of the last administration, the hippocampus tissues of mice were extracted and cut into pieces, and then completely soaked in 2 mL digestive fluid (DMEM containing 1 mg/mL collagenase Ⅳ and 1 mg/mL DNA enzyme Ⅰ). The above mixture was then placed in a 37 °C water bath for digestion. Ice was added after 45 min to stop digestion. The digested brain tissue solution was run through a 40 µm cell filter in order to exclude cell masses and tissue masses that had not been adequately digested. To obtain a single-cell suspension, the filtrate was centrifuged at 1000 rpm for 5 min while the cell precipitation was resuspended in PBS. Wash with PBS twice, centrifuge at 1000 r/min for 5 min, remove the supernatant, add an appropriate volume of diluted DCFH-DA working liquid, and incubate at 37 °C for 30 min in the dark. Following incubation, the cells underwent two PBS washes in order to eliminate any remaining DCFH-DA from the cells. In 500 µL of PBS, the cells were suspended. The ROS positive rate was found using flow cytometry.
Measurement of ROS levels in cells: PC12 cells were placed in a 6-well plate,
3.5
The isolated hippocampal tissue was homogenized in cold phosphate buffer (pH 7.4) and centrifuged at 4 °C at 10,000 rpm for 15 min. The centrifuged supernatant (serum, cell culture supernatant) was collected and 100 µL supernatant was added to the plate. The levels of MDA, SOD, and GSH in supernatant, serum, and cell culture supernatant were measured using specific kits according to the manufacturer’s instructions.
Extraction of total protein from hippocampus and determination of protein
content were carried out respectively with RIPA lysis and BCA protein assay kit.
Later on, we performed electrophoresis of protein (40 µg) for 1 h on
SDS-PAGE (10%) to PVDF membrane with 120 V of transmembrane step. After being
blocked with 5% BSA, membranes were incubated with the primary antibodies
(BCL-2, Bax, HO-1, P-Akt, Akt, GSK3
Tissue or cell samples that have been fixed in PFA (4%) were subjected to drying, paraffin embedding, slicing, dewaxing, hydrating, antigen extraction, blocking, and all-night incubation at 4 °C with primary antibody. Afterwards, sections were subjected to three PBS washes before the FITC conjugate secondary antibody was used for their incubation for 1 h at 37 °C. Later on, an anti-fade mounting medium with 4, 6-diamidino-2-phenyl-indole (DAPI) was used to mount the slices after washing. Ultimately, we evaluated the sections with a fluorescent microscope.
Following the manufacturer’s instructions, TUNEL labeling was used to identify neuronal cell death in the hippocampal regions of mice. Under a fluorescent microscope, the sections were taken in pictures. To count the cells that were TUNEL positive, Image J was employed. Data was expressed as a ratio of the number of TUNEL positive cells to the square millimeter.
Assessment of the effects of various dosage forms on cell viability was accomplished with the MTT assay. Growing of PC12 cells was carried out in 96-well plates for 24 h with 8000 cells/well. Twenty-four (24) h after an in vitro cell model preparation, we exposed the cells to various amounts of drug-containing serum. Following a 24 h treatment period, we applied MTT (5 mg/mL, 20 µL) to each well, before the removal of supernatant after 4 h. Later on, we added DMSO (150 µL) to each well, while complete dissolution of dirty crystals was accomplished with 10 min of oscillations at low-speed. At a wavelength of 570 nm, we used an enzymatic-labeled meter to determine the absorbance OD values.
According to the manufacturer’s instructions, we detected cell apoptosis with
Annexin V-FITC/propyl iodide (PI) apoptotic kit. To put it simply, we inoculated
PC12 cells into 6-well plates with 3.5
To do the statistical analysis, GraphPad Prism 8.0 (GraphPad Software, Inc., San
Diego, CA, USA) was used. We expressed the data with mean
The aging process of SAMP8 mice is accompanied by complex physiological changes
related to cognitive dysfunction, such as brain A
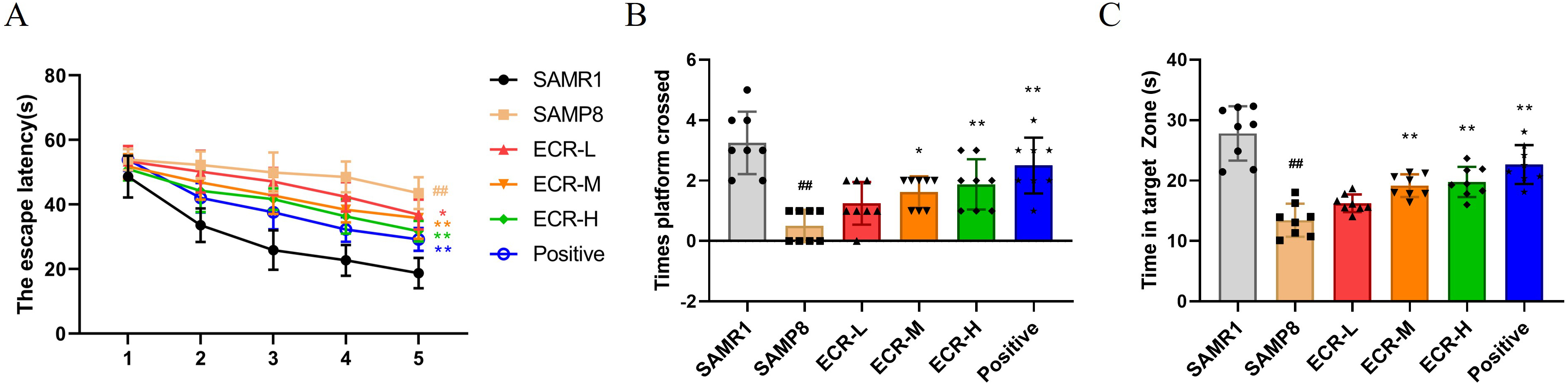
 Fig. 1.
Fig. 1.ECR improves cognitively deficient behavior in SAMP8 mice.
Effects of ECR treatment on (A) escape latency, (B) number of times crossed the
target platform position and (C) time spent in the target quadrant in Morris
water maze (MWM). (A) Data were analyzed using two-way ANOVA and a Bonferroni
test or (B,C) using a one-way ANOVA and Tukey’s post hoc test and presented as
the mean
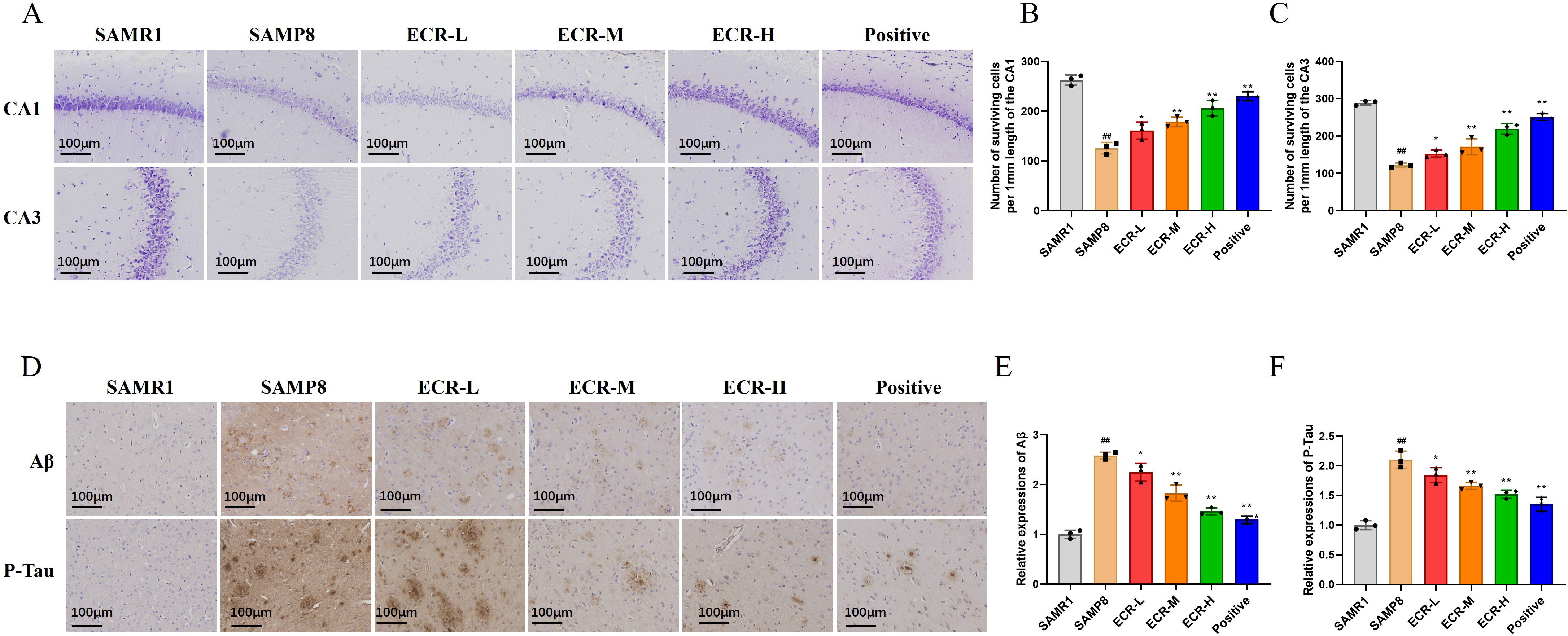
 Fig. 2.
Fig. 2.ECR improves cognitive deficits in SAMP8 mice. (A) Illustration
of images of Nissl staining of hippocampal regions (CA1 and CA3). Surviving cells
per 1 mm of (B) CA1 and (C) CA3 were analyzed quantitatively. In each group, n =
3, while the scale bar = 100 µm. (D) By using immunohistochemical staining,
A
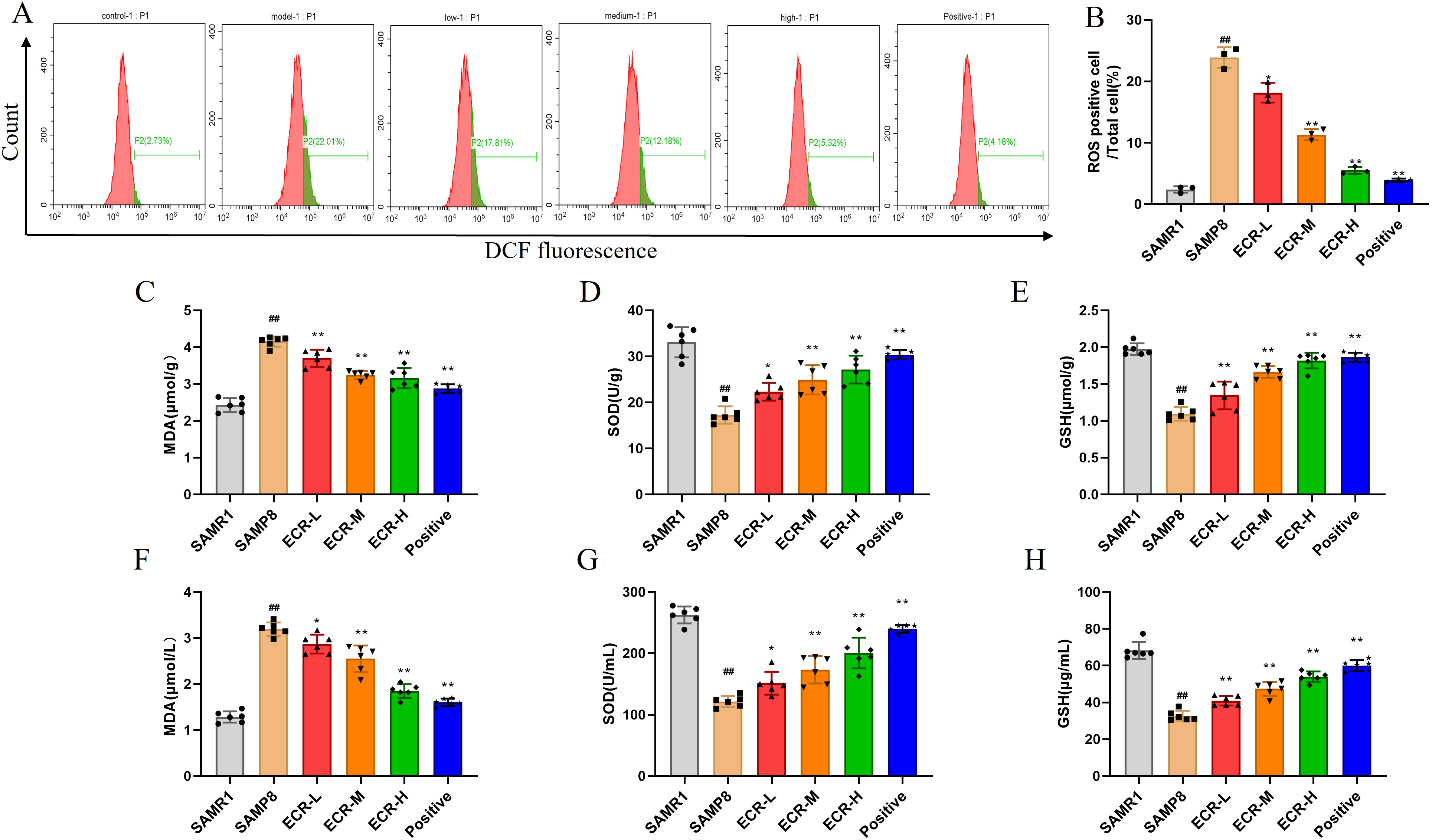
Scientists have shown that oxidative stress has a significant influence on the development of AD [7]. Based on this assertion, we evaluated oxidative stress-linked biomarkers to ascertain the involvement of this process in the ability of ECR to ameliorate cognitive deficits in SAMP8 mice. As shown in Fig. 3A,B, ROS levels considerably increased in the hippocampus of SAMP8 mice compared to the control group but decreased significantly after treatment with ECR or Donepezil, and ECR showed a dose-dependent manner. Important indexes that reflect oxidative system imbalance are GSH, MDA and SOD. As can be seen from Fig. 3C–E, SOD (Fig. 3D) and GSH (Fig. 3E) activities in the hippocampus of the model group were significantly decreased, while MDA (Fig. 3C) content was increased. These changes were significantly reversed by ECR or Donepezil, with a stronger reversal effect observed at higher doses of ECR. Similarly, the detection of MDA, SOD and GSH levels in the serum of mice was consistent with that in the hippocampus (Fig. 3F–H). Thus, inhibition of oxidative stress by ECR may facilitate its ameliorating effect on cognitive deficits in SAMP8.
 Fig. 3.
Fig. 3.Antioxidative effect of ECR against oxidative stress in SAMP8
mouse model. (A,B) Determination of levels of ROS in the hippocampus with flow
cytometric technique after administration of different dosage forms (n = 3). (C)
MDA, (D) SOD, and (E) GSH in the hippocampus were detected by biochemical kits (n
= 6). (F) MDA, (G) SOD, and (H) GSH in serum were detected by biochemical kits (n
= 6). The data were presented as the mean
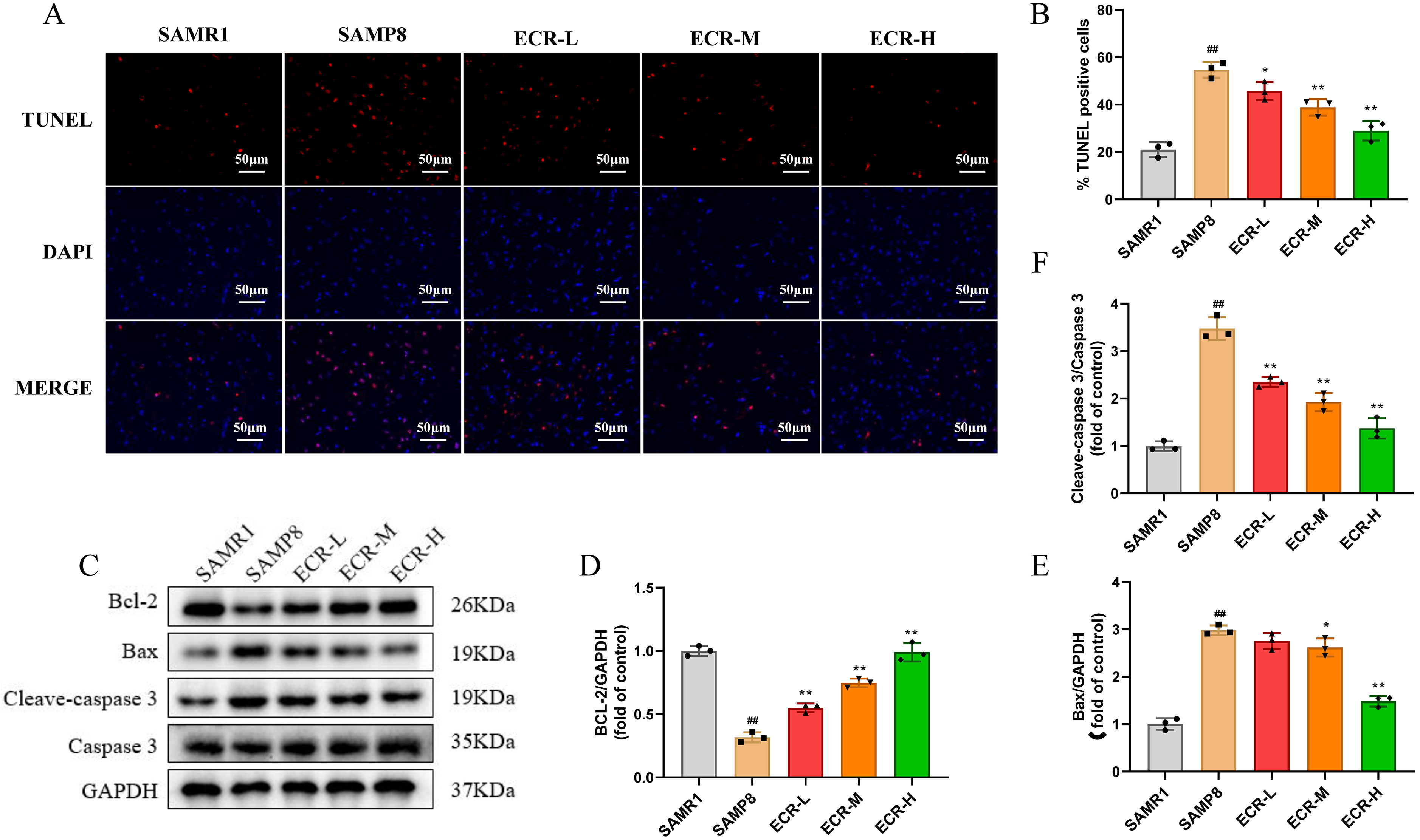
Much evidence suggests that peroxidation of lipids and proteins is the consequence of oxidative system imbalance, which ultimately culminates in apoptosis in cells [28]. To investigate whether inhibition of oxidative stress by ECR may further result in reduced cell apoptosis, we evaluated cell apoptosis and expression levels of apoptotic-linked protein in the hippocampus of the mice. To achieve this, we employed TUNEL staining to observe the ECR effect on the apoptosis of neurons in the hippocampus. In terms of results, we discovered a substantially increased number of apoptostic-positive cells in model mice compared to control, with the cells displaying distinctive morphological features of cellular apoptosis. Meanwhile, we observed a significantly decreased number of TUNEL positive cells in the hippocampus of mice that received ECR compared to model mice (Fig. 4A,B). Following that, we ascertained the alterations in proteins that have been linked to apoptosis with the western blotting technique. Regarding the findings, we saw that model mice expressed less Bcl-2 than control mice did, while the former group expressed more Bax and cleaved caspase-3 (Fig. 4D,E) . Nevertheless, ECR treatment could significantly reverse these changes, wherein the reversal was more obvious with an increase in ECR dose (Fig. 4C,F). These data suggest that, in the hippocampus of SAMP8 mice, ECR has a protective impact against cell apoptosis, and that this protective effect increased with increasing ECR concentration within a specific range.
 Fig. 4.
Fig. 4.ECR inhibits neuron apoptosis in SAMP8 mice. (A) Representative
images of TUNEL staining in each group. (B) Apoptotic cells were quantitatively
analyzed. In each group, n = 3, while the scale bar = 50 µm. (C) Detection
of apoptosis-linked proteins, namely Bax, Bcl-2, cleave caspase-3, and caspase-3
in the hippocampus with western blotting technique. (D,E) Normalization of
quantified levels of protein to GAPDH (n = 3). (F) Normalization of the
quantified level of protein to caspase-3 (n = 3). The data were presented as the
mean
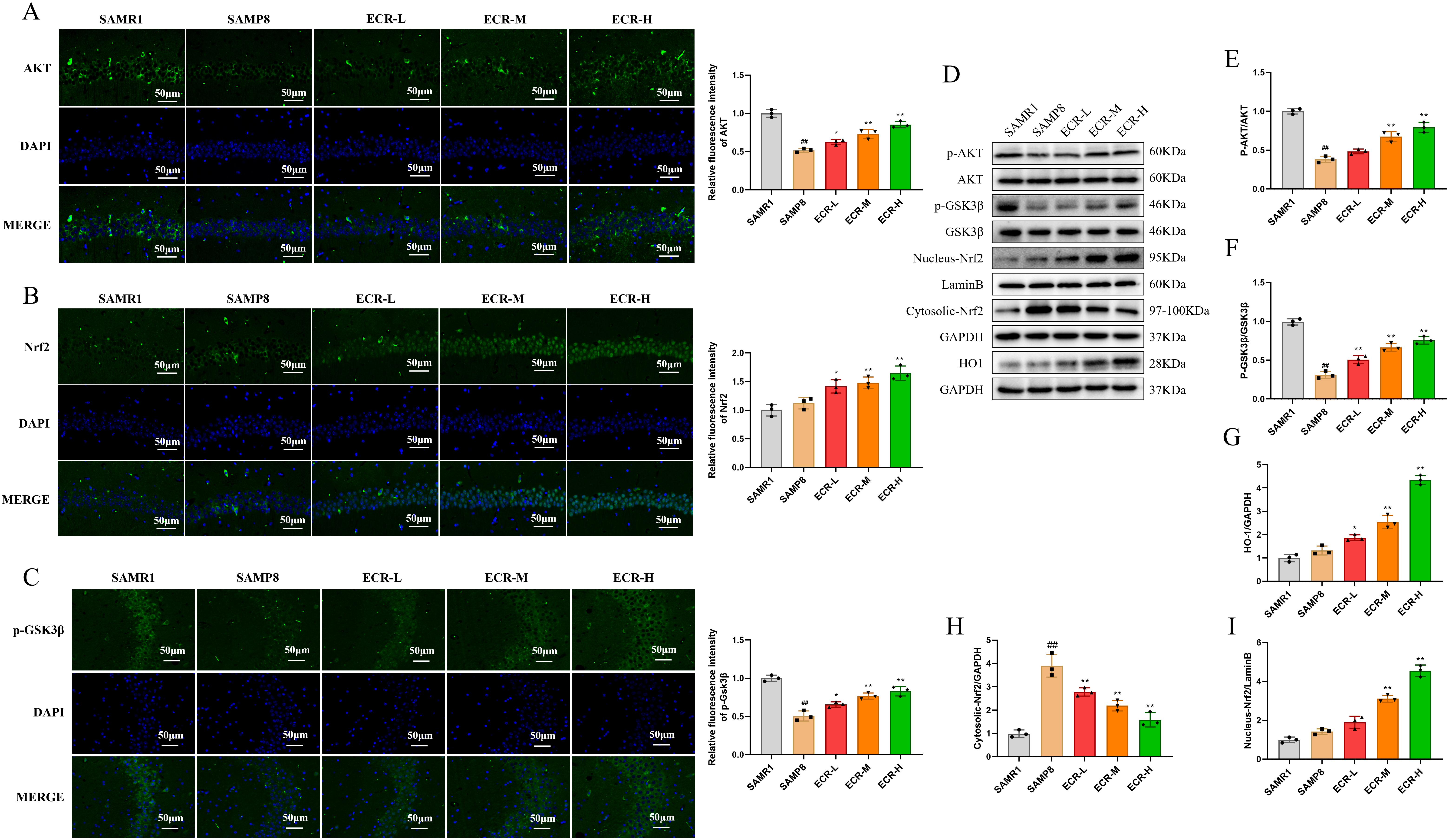
Immunofluorescent staining and western blotting techniques were utilized to
detect the expression of proteins linked to the Akt/GSK3
 Fig. 5.
Fig. 5.ECR improves cognitive deficits in SAMP8 mice by activating
Akt/GSK3
After demonstrating the potential of ECR to alleviate cognitive impairment via
prevention of oxidative stress and death of neurons in vivo, we
conducted several in vitro experiments to investigate the probable
mechanistic action of ECR in cognitive impairment improvement. First, we screened
the cell use concentrations of A
In addition, we observed that PC12 cells treated with A
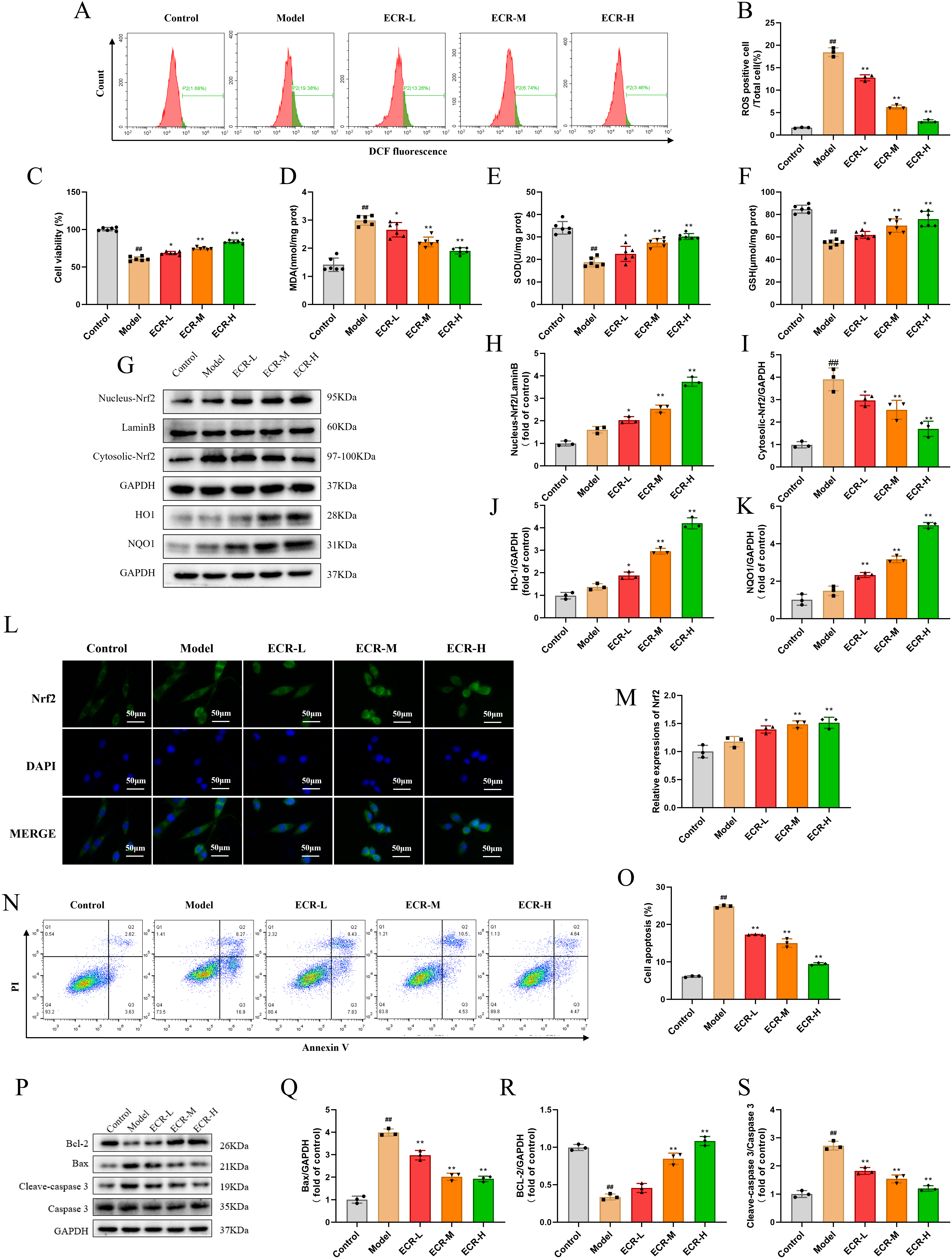
 Fig. 6.
Fig. 6.Protective effect of ECR on oxidative stress of neurons in
A
Collectively, ECR demonstrated a beneficial effect on oxidative stress and cell
apoptosis accordingly induced in PC12 cells by A
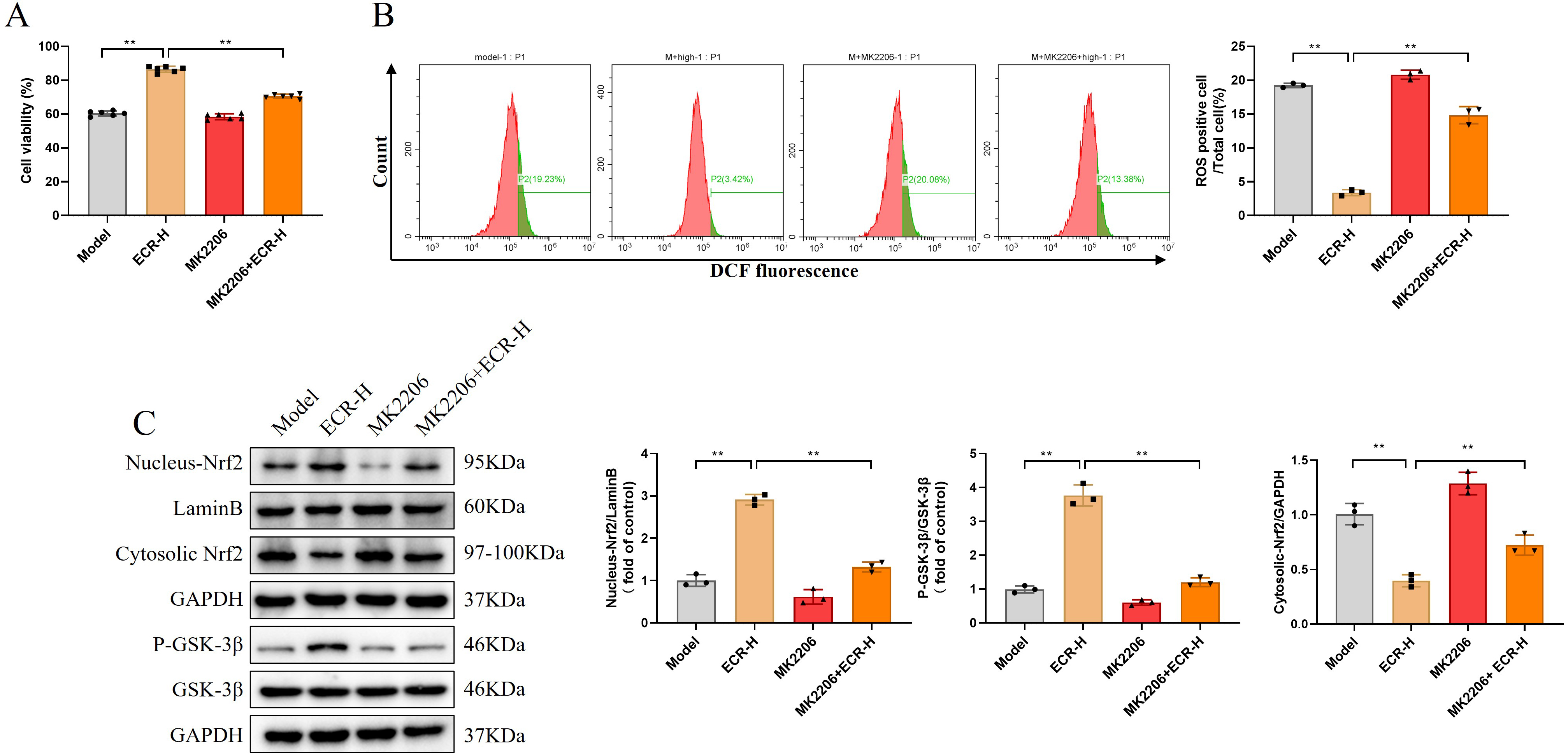
During this experiment, we employed Akt inhibitor MK2206 to further clarify the
possibility that ECR protected PC12 against oxidative stress and apoptosis
induced by A
 Fig. 7.
Fig. 7.ECR regulates Nrf2 by activating the Akt/GSK3
Herein, we attempted to understand the mechanism underlying the potential of ECR
to ameliorate cognitive impairment by conducting a series of experiments
(in vitro and in vivo). Results obtained from SAMP8 animal
models in vivo suggest that ECR may regulate Nrf2 to prevent oxidative
stress in an Akt/GSK3
The leading cause of dementia and a growing global health concern, AD has significant effects on both individuals and society [29]. Despite significant research efforts to find a therapeutic approach to stop the course of AD or to cure it, currently available medications are only effective in treating its symptoms and their efficacy is still insufficient [5]. Hence, efficacious treatment options for AD are urgently needed. ECR is a natural substance and the main steroid hormone. It has anti-oxidative and neuroprotective properties [22, 30]. However, the therapeutic impact of ECR on AD and its molecular mechanism have not been sufficiently investigated. The SAMP8 mouse model’s cognitive deficiencies were found to be improved by ECR and positive treatment in this study, and ECR was demonstrated in a dose-dependent manner.
Oxidative stress, which is caused by an excess of ROS, has been associated with
various disorders. Due to the high oxygen consumption in the brain, these free
radicals may cause more visible damage than what cell molecules can scavenge
[31]. Oxidative stress has emerged as a crucial approach in the prevention and
treatment of AD since it plays a significant role in the disease progress and may
be regarded as a major factor in its development [32, 33, 34]. Therefore, we evaluated
the ECR effect on oxidative stress in the hippocampal of SAMP8 mice or PC12
celles induced by A
Apart from crucially regulating the antioxidative system, Nrf2 has been found to
additionally control the response of inflammation, homeostasis of intracellular
redox and other processes of biological importance [36]. Besides, research has
demonstrated that one of the body’s most crucial antioxidant defense mechanisms
is the Nrf2/HO1 pathway [37]. It has been posited that the antioxidative system
of Nrf2 (for example) might reduce the severity of several diseases by preventing
oxidative stress when downstream genes like NQO1 and HO1 are controlled [38].
Additionally, one of the several upstream signaling pathways that target Nrf2 is
the Akt/GSK3
In conclusion, this study suggested that ECR reduced oxidative stress by
activating the antioxidative system of Nrf2 via the Akt/GSK3
The datasets used and analyzed during the current study are available from the corresponding author upon reasonable request.
HX wrote the original draft. HX, SX and DC designed the research study. HX and GC performed the research. FR and HZ provided help and advice on the experiments. LX, QL and RL analyzed the data. HX and DC acquired the funding. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
The Jiangsu University (UJS IACUC) institutional committee for the care and use of laboratory animals reviewed and approved (approval number: UJS-IACUC-2022031401) for studies involving animals.
Not applicable.
This project was supported by Nanjing Young Talents Training Plan of Traditional Chinese Medicine Project (ZYQ20065) and Anhui Provincial Key Research and Development Plan (202104j07020008).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.