


 , Yuhan Hu 1,2,3,*
, Yuhan Hu 1,2,3,*1 Department of Pathology, The First Affiliated Hospital of Xinxiang Medical University, 453000 Xinxiang, Henan, China
2 Department of Pathology, School of Basic Medical Sciences, Xinxiang Medical University, 453000 Xinxiang, Henan, China
3 Department of Oncology, The Third Affiliated Hospital of Xinxiang Medical University, 453000 Xinxiang, Henan, China
Abstract
Colorectal cancer (CRC) is the most common malignancy of the gastrointestinal, however, the underlying mechanisms of CRC remain largely unknown. New evidence suggests that the PI3K/AKT/mTOR pathway is closely related to CRC. PI3K/AKT/mTOR is a classical signaling pathway that is involved in a variety of biological processes, such as regulating cellular metabolism, autophagy, cell cycle progression, cell proliferation, apoptosis, and metastasis. Therefore, it plays a crucial role in the occurrence and development of CRC. In this review, we focus on the role of the PI3K/AKT/mTOR pathway in CRC, and its application of to the treatment of CRC. We review the importance of the PI3K/AKT/mTOR signaling pathway in tumorigenesis, proliferation and progression, and pre-clinical and clinical experience with several PI3K/AKT/mTOR pathway inhibitors in CRC.
Keywords
- colorectal cancer
- PI3K/AKT/mTOR signaling pathway
- inhibitors
Colorectal cancer (CRC) has the third highest incidence and is the fourth highest cause of cancer-related deaths worldwide [1]. Studies suggest that the mortality rate from colorectal cancer will increase substantially by 2035. CRC metastasis, particularly colorectal liver metastasis, remains the most common cause of cancer death [2, 3]. In recent years, the proposal of targeted therapy has provided a new direction for the treatment of colorectal cancer. Finding key molecules involved in the regulation of colorectal cancer and developing targeted therapy drugs have become the key to clinical treatment of colorectal cancer and prolong the prognosis.
With the deepening of research, mTOR molecule and its participation in AKT signaling pathway play a key role in cancer, including colorectal cancer, making it a hot spot in targeted therapy research.
mTOR should belong to the phosphoinositide 3 kinase-related kinases (PIKK) protein family as it is very similar to phosphatidylinositol 3-kinase (PI3K) and phosphatidylinositol 4-kinase (PI4K) in the C-terminal chemical structural domain [4, 5, 6, 7]. The PI3K/AKT pathway serves as the upper axis of the mTOR pathway, and its upstream signaling affects the function of mTOR in regulating tumor and endothelial cell proliferation as well as cell survival [8]. PI3K/AKT/mTOR pathways regulating physiological functions can be activated by a variety of stimuli such as growth factors, nutrients, energy and stress signals. Studies have shown that mTOR is involved in multiple signaling pathways together with PI3K/AKT/mTOR, tuberous sclerosis complex subunit 1 (TSC1)/tuberous sclerosis complex subunit 2 (TSC2)/Rheb, Adenosine-5’-monophosphate-activated protein kinase (AMPK), VAM6/Rag GTPases, and others [9, 10]. The PI3K/AKT/mTOR pathway may be important in keeping cells alive when they are stressed. Tumor cells are in an environment with low pH, limited nutrients, and limited oxygen, so we cannot ignore the critical role of the PI3K/AKT pathway in cancer [11]. PI3Ks act as a family of activator enzymes capable of phosphorylating the 3-OH cluster of the inositol ring in phosphatidylinositol membrane lipids and helps to regulate signaling pathways. It includes three types, I, II, and III, with class I being strongly associated with cancer and subdivided into IA and IB, which are heterodimeric protein kinases with regulatory and catalytic subunits [12]. PI3K stimulates the signal transduction cascade and promote the activation of AKT. AKT is a vital signal regulator of cell growth by responding to various cellular stimuli [13].
Based on the key role of mTOR and its signal pathway in tumorigenesis and development, a variety of inhibitors, including rapamycin, have been developed and are expected to be applied to clinical treatment of a variety of malignant tumors, including breast cancer, lung cancer, gastric carcinoma, colorectal cancer, prostate cancer, head and neck cancer, gynecologic cancer, glioblastoma, lymphoma, urinary bladder cancer, renal cancer and medulloblastoma [14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25]. However, the resuits of mTOR inhibitors used as monotherapy in cancer are sometimes suppressed by several resistance mechanisms [26].
PI3K/AKT/mTOR is a common mutational pathway in tumors and is critical in CRC [27]. Neighboring targets of the PI3K/AKT/mTOR pathway are involved not only in the regulation of cell metabolism, proliferation, cell cycle, and apoptosis, but also in cancer development and metastasis as well as chemoresistance [28, 29, 30, 31, 32].
The core components of the PI3K/AKT/mTOR signaling network include
phosphatidylinositol 3-kinase (PI3K), AKT or protein kinase B (PKB), and target
of rapamycin-like (mTOR) [33]. PI3Ks are heterodimers consisting of regulatory
(p85) and catalytic (p110) subunits with four isoforms
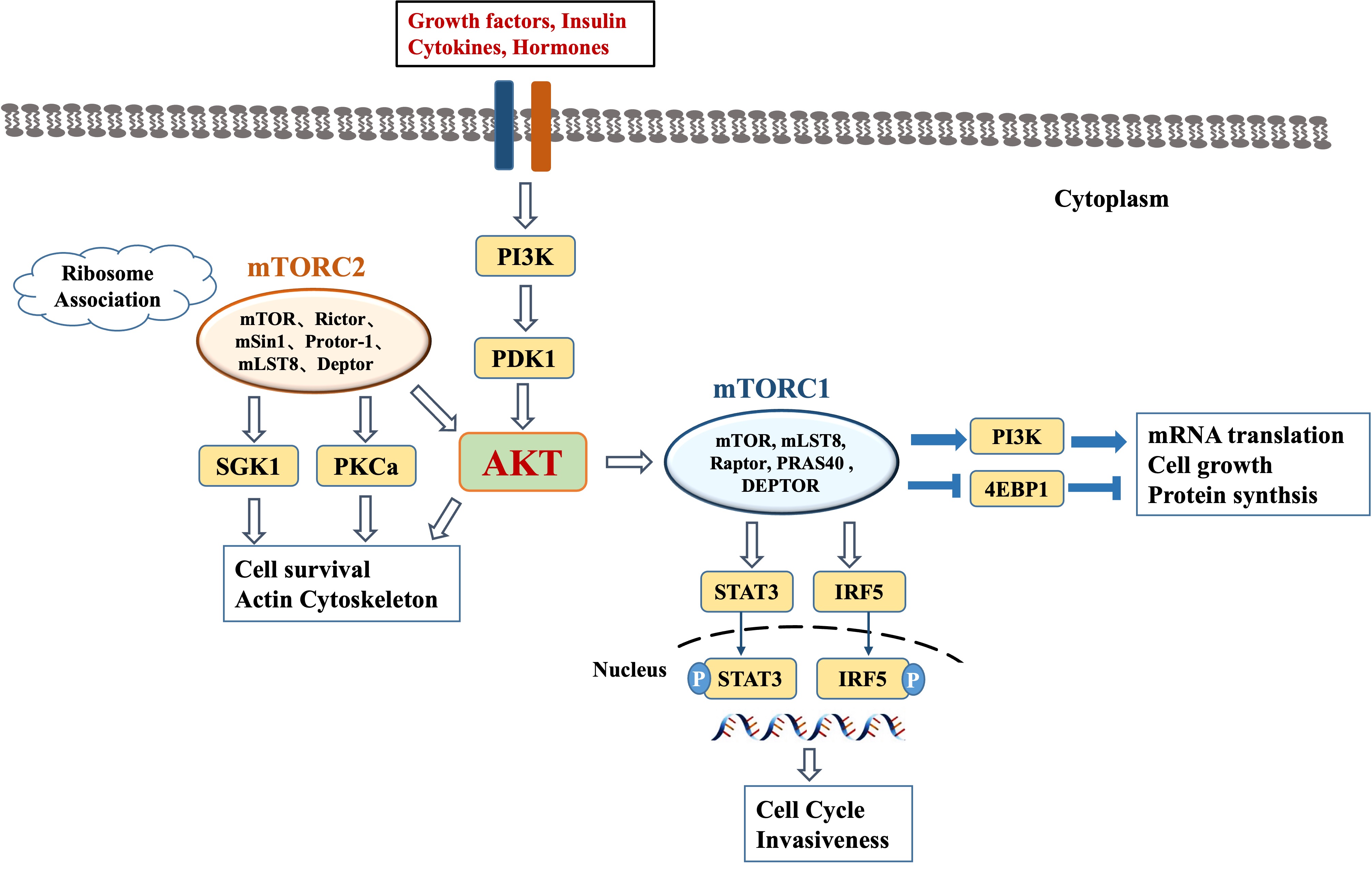 Fig. 1.
Fig. 1.The overview of PI3K/AKT/mTOR signaling pathway.
| mTOR complex | mTORC1 | mTORC2 |
|---|---|---|
| Components | mTOR, Raptor, G |
mTOR, Rictor, mLST8/G |
| Function | Controls many high-energy intracellular processes, promoting cell growth and increasing cell biomass in nutrient-rich conditions and allowing the autophagic use of cellular elements in nutrient-poor conditions | Interacts with growth factors to accelerate entry into the cell cycle, maintain cellular state, and promote actin cytoskeleton polarization and anabolism |
Abnormal PI3K/AKT/mTOR signaling pathways have been found in a variety of cancers [56]. Dysregulation of this pathway commonly leads to pathological changes. It is activated in several types of cancer, especially colorectal cancer [57, 58]. Studies have found that the PI3K/AKT/mTOR pathway is involved in controlling physiological processes such as cell growth, adhesion, migration, and survival [59, 60]. The PI3KAKT/mTOR pathway links many substances in the body to physiological processes in cells, nutrient synthesis and metabolism, angiogenesis, and tissue development [61, 62]. Studies have shown that the PI3K/AKT signaling pathway is actively involved in a variety of tumors [63]. mTOR can receive upstream signals from the PI3K/AKT pathway and holds the potential as a therapeutic marker for a variety of cancers including non-small cell lung cancer, colorectal cancer, kidney cancer, non-Hodgkin’s lymphoma and leukemia [64, 65, 66, 67, 68]. In addition, activation of the PI3K/AKT pathway can affect the prognosis of stage II colon cancer and may be a favorable prognostic factor in colon cancer [69].
PI3Ks are intracellular lipid kinases involved in the regulation of cell
proliferation, differentiation and survival. Overexpression of the PI3K/AKT/mTOR
signaling pathway has been reported in various cancer types, including CRC. PI3Ks
are known kinases that promote cell proliferation. Mutations in the
PIK3CA gene encoding the PI3K p110
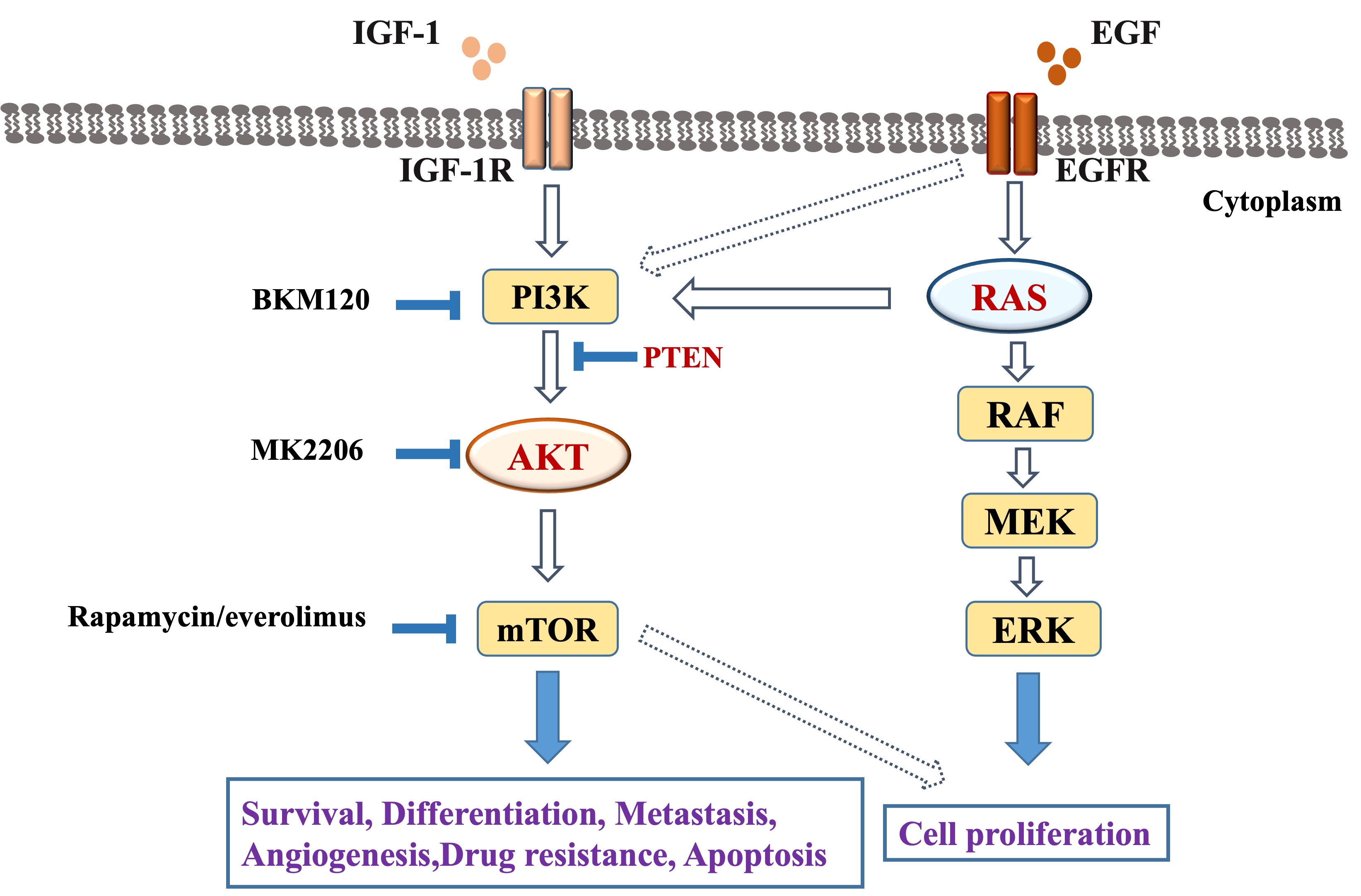 Fig. 2.
Fig. 2.Role of PI3K/AKT/mTOR signaling pathway in colorectal cancer.
Change in colorectal cancer cell genes mainly involve overexpression of insulin-like growth factors and reduced PTEN function. The PI3K/AKT/mTOR pathway regulates the uptake and release of glucose and multiple amino acids by colonic epithelial cells, as well as the response to complex extracellular signals. This pathway influences the metabolism and uptake of substances by converting stimuli received by cells into intracellular signals and is involved in a variety of cellular processes, so that it strongly influences tumorigenesis, progression, metastasis, and prognosis [74]. It is known that mTOR promotes protein synthesis, cellular proliferation, and angiogenesis in colon tumors and inhibits apoptosis and autophagy. The AKT/mTOR signaling pathway regulates the cell cycle by coordinating DNA replication and activating cyclin-dependent kinases (CDKs), which are the drivers of cell cycle progression [75].
Over-activation of mTOR signaling is common in human colorectal cancer, and it is closely associated with cancer initiation, progression, and drug resistance [76]. In CRC, mTOR significantly regulates the growth, proliferation, differentiation, survival and autophagy [77, 78, 79]. The PI3K/AKT/mTOR pathway inhibits cell proliferation by decreasing the levels of cell cycle proteins [80, 81]. The mTOR signaling pathway is involved in various cellular processes as well as cell growth, proliferation, and polarization [82]. Three subfamilies, MAPK/ERK, c-jun amino-terminal or stress-activated protein kinase (JNK or SAPK), and MAPK14, play important roles in colorectal cancer. ERK/MAPK regulates the proliferation, differentiation, survival, and death of colorectal cancer cells. The ERK pathway affects the proliferation, migration, and invasion of colon cancer cells [83].
VEGF is that the most potent pro-angiogenic tissue that contributes to tumor cell migration and increased vascular permeability. During the development of colorectal cancer, the mTOR signaling pathway is involved with the multi-stage regulation of VEGF-mediated angiogenesis [84]. During neoangiogenesis in colorectal cancer, AKT interacts with nitric oxide (NO) through various phosphorylation mechanisms and activates endothelial nitric oxide synthase, that produces gas and regulates endothelial cell migration and angiogenesis through AKT signaling [85]. The mTOR pathway strongly regulates the cloning, differentiation, invasion, and metastasis of tumor cells [86]. AKT in an activated state is often found in hypofractionated tumors, which connects oncogenic factors to surviving cells, increases tumor cell invasiveness and generates drug resistance [87].
A highly active state of the PI3K/AKT/mTOR signaling pathway increase drug resistance in colorectal cancer cells, inhibits apoptosis, and promotes tumor cell survival [88]. The gain or loss of function of proteins on this signaling axis leads to the transformation of multiple cancers, therefore investigating inhibitors of this pathway will have a role in treating these tumors [89]. Scientists expect the PI3K/AKT/mTOR pathway to become a drug target for tumors and are currently working to develop drugs that inhibit the PI3K/AKT/mTOR pathway [90]. The PI3K/AKT/mTOR pathway significantly affects human cancer. Pharmaceutical regulatory agencies have approved mTOR inhibitors for use in a variety of cancers [91, 92, 93].
Drugs that interfere with PI3K component signaling are undergoing clinical
evaluation. Drugs developed by investigators include PI3K inhibitors, dual
inhibitors of PI3K and mTOR, AKT inhibitors, and mTOR inhibitors [94, 95]. mTOR
inhibition prevents the loss and promotes regeneration of proliferating
colorectal cancer epithelial cells following radiation [96]. The effective,
irreversible PI3K-specific inhibitor BKM120 (IC50: 2–5 nM) inactivates PI3K by
covalently modifying the Lys-802 residue involved in the phosphotransfer
reaction, which inhibits cancer cell growth and has anticancer effects [95]. Dual
PI3K/mTOR inhibitors are theoretically able to completely shut down the PI3K
pathway, thereby eliminating the effect of negative feedback from mTORC1
metamorphosis inhibitors, which is not possible with the other three inhibitors
[97]. Rapamycin has been shown to have an inhibitory effect on mTOR [98]. MK-2206
may effectively inhibit AKT isoforms and thus have anti-tumor activity [99].
Selective inhibitors of AKT/PKB, act on inactivated or activated AKT and prevent
its translocation [100]. Together with Bad, GSK-3
mTOR inhibitors induce autophagy or apoptosis in colonic CSCs. The mTOR inhibitor Torin-1 impedes the growth, invasion and survival of CD326+/CD24+/CD49f+/CD29+ and CD326+/CD44+/CD166+ CRC subpopulations in vitro and inhibits tumor growth and angiogenesis in vivo [103]. The use of rapamycin or the stable inhibition of mTORC1 (Raptor) and mTORC2 (Rictor) inhibited CRC migration and invasion [104]. The selective tyrosine kinase inhibitor BKM120 blocks the PI3K signaling pathway in colon cancer. There are now many effective inhibitors of different types of AKT [105]. In study the antiproliferative effects of mTORC1 and mTORC2 inhibitors on colorectal cancer cells, researchers found that the two generations of mTOR inhibitors, mTORC1, and pp242 (mTORC1/2), do not have the same effect on colorectal cancer cells. Recent findings suggest that mTORC2 can directly regulate AKT activity thereby having a stronger effect on colorectal carcinogenesis and proliferation [106]. The apoptosis-inducing mTOR inhibitor Torin-1 inhibited tumor growth both in vivo and in vitro, raising expectations for their role in tumor therapy [107]. The highly active state of mTOR signaling in colorectal cancer promotes growth, proliferation, survival, and anti-apoptosis in a variety of tumor cells [108]. The PI3K/AKT/mTOR axis governs a range of tumor-associated cellular activities, such as promoting growth, proliferation, metabolism, angiogenesis, and inhibition of apoptosis. The PI3K/AKT/mTOR pathway is now used as a drug-binding site for the treatment of colorectal cancer [109].
Drug resistance in tumor cells is responsible for the poor survival of cancer patients. Although the number of targeted agents available for clinical use is gradually increasing, many of these drugs have performed poorly in clinical trials [110]. Due to the heterogeneous nature of most tumors, fixed drug regimens often fail to achieve the desired therapeutic effect in patients with the same cancer. Moreover, mTOR inhibitors can exert their antitumor effects through multiple mechanisms, which adds to the difficulty of accurately predicting the biological factors underlying inhibitor efficacy or resistance [111]. mTOR inhibitors are deficient in that they are deficient in that they only inhibit mTOR without blocking the activity of the mTOR-binding ligand, which may still transmit growth signals upon stimulation by cell death despite the loss of mTOR kinase activity by the binding ligand. Furthermore, crosstalk of the mTOR pathway with other pathways allows tumor cells to escape anti-tumor stimuli via other pathways. This suggests that signaling is complex and that therapies that merely inhibit feedback regulatory pathways often fail to achieve the desired effect in clinical practice. For example, the four inhibitors mentioned above hold great promise in theory as well as in clinical trials but are still not approved for use in practice. The first identified mTOR inhibitor, rapamycin, incompletely inhibited mTORC1 and was inactive against mTORC2 for a short period. PI3K inhibitors, dual PI3K-mTOR inhibitors, AKT inhibitors, or mTOR inhibitors block different sites of the PI3K/AKT/mTOR axis [112]. Although many drugs targeting the PI3K/AKT/mTOR axis are in development and a small number of them have been approved for clinical practice, there is skepticism regarding their therapeutic benefits [113]. Research and clinical trials related to PI3K/AKT/mTOR inhibitors have opened new oncological treatment options and revealed the limitations of established therapeutic approaches. The PI3K/AKT/mTOR axis is often cross-linked to multiple signaling pathways and the mechanisms of resistance to its inhibitors are mainly classified as intrinsic resistance and purchased resistance [114].
Central to the successful development of mTOR pathway inhibitors in colorectal cancer is the understanding of the site of action of each mTOR inhibitor to better select drugs for specific tumors [115]. The PI3K/AKT/mTOR signaling pathway is associated with autophagy, and glucose metabolism in many cancers, leading to their radioresistance [116]. These drugs have can help to treat cancer they can produce negative side-effects such as dermatitis, anemia, diarrhea, and neurotoxicity [117]. Further study of PI3K/AKT/mTOR pathway inhibitors is necessary to understand their role in cancer therapy [118]. Although the overall survival of colorectal cancer patients has improved and its mortality rate has decreased more intensive research in colorectal cancer is necessary to develop effective treatments that will continue to improve outcomes [119]. PI3K stimulates the signaling cascade and promotes the activation of AKT, an important regulator of cell growth signaling. mTOR is mainly regulated by the PI3K/AKT/mTOR signaling pathway. Research targeting the PI3K/AKT/mTOR signaling pathways is currently underway to develop new therapeutic regimens to cure and prolong survival in patients with CRC.
A meta-analysis is a statistical technique for statistical and comparative analysis of different results on the same subject to address variations in clinical studies. It combines published and unpublished studies to pool the results of interventions with clinical and intermediate outcomes by conduction a systemic review of a particular subject [120]. It is mostly beneficial when applied to randomized controlled trials (RCTs) [121, 122]. There are still many gaps in research involving the clinical and pathological implications of PI3K-AKT-mTOR mutations. A meta-analysis showed that mutations in the PI3K-AKT-mTOR pathway were significantly associated with the advanced Tumor Node Metastasis (TNM) stage but not with the tumor grade [123, 124]. Bias may arise either by not combining relevant studies or by including unconfirmed studies [125].
A meta-analyses showed that mTOR pathway protein expression could predict survival in patients with a complete mTOR pathway protein expression of 74.42 [126]. A systematic review and meta-analysis demonstrated that activation of the PI3K/mTOR/AKT pathway is associated with numerous solid tumors (e.g., colorectal cancer, breast cancer, gastrointestinal tumors, gynaecological tumors, prostate cancer and non-small cell lung cancer) and is related to survival [127, 128, 129].
Meta-analysis concluded that the PI3K/AKT/mTOR pathway was associated with
colorectal cancer. Moreover, simultaneous assessment of Phosphatase and Tensin Homolog (PTEN) levels and
activation of pS6 and AKT indicated a poorer prognosis. The overall expression of
74.42% of mTOR pathway proteins was overexpressed in most head and neck squamous cell carcinoma (HNSCC) and was
associated with reduced Overall Survival (OS) and DFS in HNSCC patients. mTOR pathway has the
potential to be a therapeutic target for HNSCC. The best estimate overall in a
meta-analysis is usually defined as SD. The premise that PI3K/AKT/mTOR inhibitors
are effective in Clinical Benefit Rate (CBR) is that the disease is stable [130]. The time to stable
disease progression is beneficial for disease assessment. They used a cut-off of
With advances in treatment, the survival of colorectal cancer patients has improved and mortality has decreased [131]. The development of targeted drugs targeting the PI3K/AKT/mTOR pathway is changing the treatment of colorectal cancer. A meta-analysis of the effectiveness of these drugs acting on patients with colorectal cancer, using SD as an outcome indicator requires consideration of time to disease stabilization. Screening for SD, therefore, is required for an accurate understanding of drug efficacy. Furthermore, the use of these inhibitors in cancer therapy has been associated with drug-related adverse effects in some patients. It remains to be seen whether the benefits outweigh the risks with these targeted therapies. We also observed better efficacy of PI3K and mTOR inhibitors compared to the other two inhibitors, but because of limited data, this result was not statistically significant [132]. Three separate experiments were conducted in the mTOR subgroup to investigate these inhibitors and found that tesilomox improved their effectiveness. Intravenous tesilomox improved drug utilization, but the mixed group of oral PI3K inhibitors had the highest CBR. Frequent mutations in the PI3K pathway may account for the slightly better CBR than the other inhibitors. Theoretically, dual PI3K/mTOR inhibitors may be more effective than single inhibitors and have a more promising future [133].
In conclusion, there were significant differences in the efficacy of the different types of inhibitors. We preferred PI3K and mTOR inhibitors among the four inhibitors and found that they have a good therapeutic effect, but this was only observed with limited data and was not statistically significant [134]. The strength of the current meta-analysis is that we studied a large number of advanced solid tumors and performed a systematic review in conjunction with the available information on studies related to PI3K/AKT/mTOR inhibitors [135].
The development of CRC is influenced by inflammatory cells and their inflammatory mediators. This review focuses on the PI3K/AKT/mTOR axis and colorectal cancer. The multiple roles of the PI3K/AKT/mTOR axis in colorectal cancer are closely linked to the development of drug resistance in tumor therapy. As colorectal cancer continues to be explored and related therapies evolve, a decrease in mortality in colorectal cancer patients can be observed.
Activation of the mTOR signaling pathway is closely associated with tumors and plays an important role in tumor cell proliferation and cloning, angiogenesis, invasion and metastasis, and inhibition of apoptosis. During the development of colorectal cancer, overexpression of the mTOR signaling pathway can lead to metastasis and growth of colorectal cancer cells.
Although research on the mechanism of CRC and its drug resistance has been advancing, there are still many unknowns in this field. The PI3K/AKT/mTOR axis will play an important role in developing therapeutic regimens to treat colorectal cancer and will undergo more in-depth study to improve clinical efficacy while limiting adverse drug reactions to improve survival in patients with CRC.
Conceptualization—JZ, YH; writing—original draft preparation—SD, YH, XZ, WD, XW, HZ, YC, YZ; writing—review and editing—JZ, YH; visualization—SD, YH; supervision—JZ. All authors have read and agreed to the published version of the manuscript.
Not applicable.
Not applicable.
This work was supported by the National Natural Science Foundation of China (No. U1804173), Henan province young and middle-aged health science and technology innovation talent project (No. YXKC2021044), Natural Science Foundation of Henan Province (No.212102310661), Joint construction project of Henan Medical Science and technology research plan (No. LHGJ20200504).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.