


1 Department of Zoology, Acharya Prafulla Chandra College, New Barrakpur, Kolkata, 700131 West Bengal, India
2 Department of Botany, Acharya Prafulla Chandra College, New Barrakpur, Kolkata, 700131 West Bengal, India
3 Department of Zoology, University of Calcutta, 700019 Kolkata, India
4 Department of Botany, Dinabandhu Andrews College, Garia, Kolkata, 700084 West Bengal, India
† These authors contributed equally.
Abstract
Being polymorphic, deoxyribonucleic acid is worthy of raise a variety of
structure like right-handed B to left-handed Z conformation. In left-handed
contour of DNA consecutive nucleotides substitute between syn-arrangement and
anti-arrangement, through the chain. 2D gel electrophoresis comprising d(PCpG)n
of topo isomers of a plasmid inserts d(pCpG)n, in this ‘n’ ranges among 8 to 21,
indicate the change of B-Z DNA. The high denseness of salt is required for
conversion of B configuration d(CG)n toward Z configuration. The rate of B to Z
transition is measured by “Cytosine Analogues” and “Fluorescence
Spectroscopy”. h-Z
Keywords
- Alzheimer’s disease
- Wang model
- Z-DNA-binding protein
- Z-DNA-forming sequence
- Review
In 1979, a left-handed crystal deoxyribonucleic acid structure was published, which convey a unique zigzag, sugar-phosphate backbone, it’s named as Z conformation of deoxyribonucleic acid (Z-DNA) and it’s all biological relevance had yet to be established [1, 2]. It was already known that normal right-handed B conformation can assume a diverse number of configurations, under certain torsional stress [3]. Z configuration exists in high energy state than the common B-DNA configuration. This conformation has negative super helicity which soothes the structure. In contrast to B form with anti-conformation, in Z-DNA convey anti-conformation and syn-conformations alternately by rotating around glycosyl bonds, along with the chain [4]. Under bound condition non-B-DNA structure like cruciform, triplex, hairpin, etc. are formed by collapsible monotonous DNA sequence. This unusual structure has effects on several biological progressions [5]. Super helicity is the most significant inducer for Z contour in usual DNA. Non-super helical, natural DNA holds practically no Z-DNA, but other hand the same DNA under extreme negative super helicity, as in “form V” may have as much as 35–40% of its sequence in Z arrangement [6]. Except for Z-DNA, X-ray fiber diffraction outlines were framed and differentiates several conformations of DNA. Most DNA enters the A-DNA conformation which’s per turn contain 11 bp through right-handed helix [7]. The single-crystal method resolute the complementary structure, oligo deoxy nucleosides, d(GGTATACG) and d(IODO-CCGG) [7, 8] (Table 1).
| Parameter | A-DNA | B-DNA | Z-DNA |
| Helix sense | Right-handed | Right-handed | Left-handed |
| Axial raise [in Armstrong] | 2.55 | 3.4 | 3.7 |
| Helix pitch | 28 |
34 |
35 |
| Base pair tilt | 20 |
-6 |
7 |
| Rotation per residues | 33 |
36 |
-30 |
| Diameter of helix [in angstrom] | 23 | 20 | 18 |
| Glycosidic bond configuration | Anti | Anti | Anti |
| da, dT, dC, dG | Anti | Anti | Syn |
| Inserted phosphate phosphate distance [in Armstrong] | 5.9 | 7.0 | 7.0 |
| da, dT, dC, dG | 5.9 | 7.0 | 5.9 |
| Suger pucker | C3’-endo | C2’-endo | C2’-endo |
| da, dT, dC, dG | C3’-endo | C2’-endo | C3’-endo |
Existence of B-Z transition and Z-DNA is further deep-rooted by the
specific ZBP discovery [10]. In vitro, Z-DNA was postulated for
identification of proteins that bind with it in a structure-precise manner, act
as a cis-element and aid in biological development. RNA Double Strand adenosine
deaminase 1 is a type of the ZBP [11]. This ADAR1 has a Z
The optical investigation originally proposed the Z-DNA. The result of
the experiment exhibited that a 4 mL NaCl solution contains a polymer which
consists of discontinuous cytosine and guanine residue and formed a nearly
inverted circular dichroism gamut [25]. Until 1979, the invention of Z-DNA
remained unknown. Original atomic steadfastness exposed that it was not the same
right-handed B-DNA which was invented by James D. Watson & H.C. Crick in 1953.
Despite that, this new left-handed helical structure named as Z deoxyribonucleic
acid. This Z form consists of extremely immunogenic antibodies to recognize the
configuration, unlike B form of DNA [26]. There have some familiar features of B
form with the d(Cg)
Various conformational topographies differ the Z-DNA from the B-DNA (Fig. 1). The double-helical Hexa-nucleoside Penta Phosphate molecules allied with the crystal. Crystal of Z arrangement contains discontinuous cytosine and guanine residues’-DNA is dinucleotide while B-DNA is mononucleotide with anti-configuration. All deoxycytidine has anti-configuration whereas all the deoxyguanosine has anti syn-configuration.
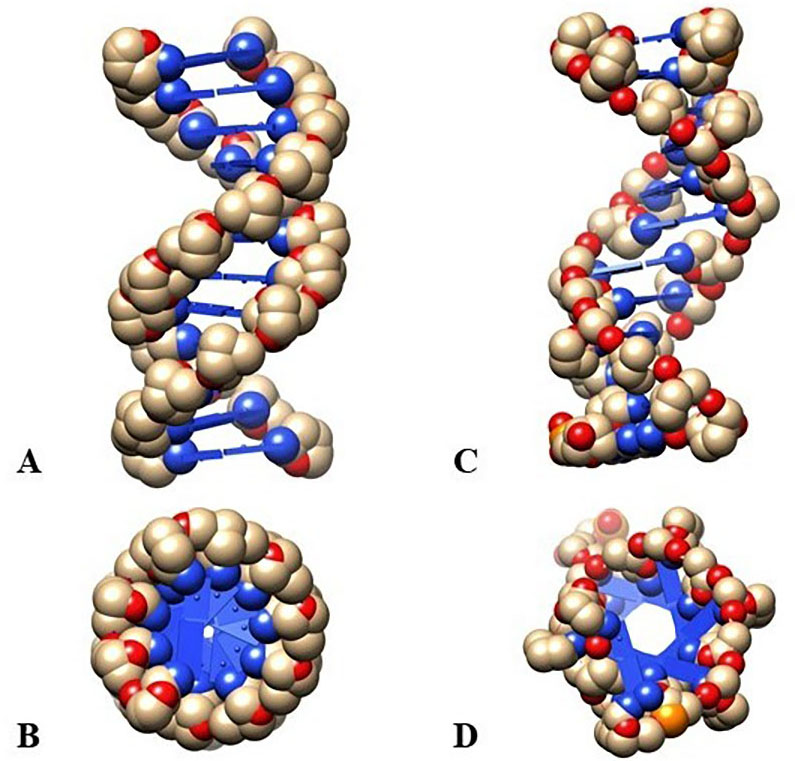 Fig. 1.
Fig. 1.Evaluation of the structure between Z and B-DNA configuration. (A, B) Z-DNA configuration showing left-handed elongated spiral with a lateral view and a polar view. (C, D) B-DNA configuration showing right-handed elongated spiral with a lateral view and a polar view.
In Z arrangement the base pair is lifted from the center, so the guanine
imidazole ring is originated at the edge, but in case of B-DNA those bases are at
the center. In B configuration 34Å pitch with 10.5 bp is present where Z
configuration convey 44.6Å pitch with 12 bp per turn [9]. Six levels of base
pairing have been seen in the d(Cg)
Earliest Harvey model is used for examining B-Z transition. This model defines the procedure which is engrossed by another longitudinal DNA conscious models. Base pairs opening was another early observed mechanism regarding this model before the Wang model. At the present portrait of Zipper Model demonstrated that Z-DNA contains high energy joint that grows through the DNA polymer until the full B-Z trans polymer gets transferred into Z-DNA. Though there are some problems in the model because it does not disclose many vibrant structural details, so it has limited applications in thermodynamics. There are several facts regarding the transition of B-Z such as the high concentration of salt in the solution which balance the Z-DNA due to massive reduction in electrostatic repulsion in the pillar of phosphate. Negative helicity of deoxyribonucleic acid needs energy that can also uncoil B form to configure the Z form. Z-DNA can also be stabilized by transcription.
Maruyama and colleague establish the B-DNA to Z-DNA transition communed by a method called “cationic graft copolymer” where the Poly (L-lysine)-graft dextran (PLL-g-Dex), begins with two-step method including the creation of a clear intermediate [27]. Amid DNA phosphate group electrostatic repulsion reduce by the cationic backbone of the copolymer and the transition is a result of these 2 factors. The most plausible Z form created negative supercoiling, utilizing B-DNA occurs during several metabolisms like Transcription and replication processes [28]. For reducing the transition stress, unusual such DNA as Z-DNA is formed [6, 29]. Lee et al. (1992) used “Magnetic-tweezers” and FRET combinedly to examine at molecule level of negative supercoiling [30]. Magnetic tweezers are a very useful technique for investigating wind/unwinding procedure of twisted DNA through precisely controlling infinite tension [28, 31]. Therefore, B-Z change can be active by tiny negative super helicity and approximately one Pico Newton Tension. This outcome suggests that in tension Z arrangement is formed more easily in vivo [32]. Methanol, Ethanol, Ethylene Glycol (Dehydrating agent) balance the Z-DNA configuration. Due to adjacent clustering counteractions all over the DNA, though more strong ionic properties, thus it provides additional mutually repelling phosphate groups [6]. Antibodies and ZBP can bind the Z form of DNA selectively. This conformation has triggering capability. The Qu group had been reported that Alzheimer amyloid protein brings about the Z-B transition. Forming the Z-form is correlated with Alzheimer’s disease [33, 34]. Bae et al. analyzed to transition from B-Z conformational change occurs by Z-DNA binding protein unravel the detailed binding machinery and whether the protein industriously initiates Z-DNA’s or passively traps transitionally performed Z form. Therefore, it proved that the conformational selection mechanism stabilized the Z-DNAs by alternating the “induced fit” mechanism. A chemical modification also stabilizes Z-DNA transformation [4]. Bulky group’s introduction precise in a certain base also steady the growth of Z arrangement by increasing static hindrance.
Double-stranded adenosine deaminase RNA is an enzyme of the deaminase
family which edited the appearance of the ds-mRNA by converting adenosine to
inosine and creating diversity between RNA and Protein [11]. It is noted as a
naturally stirring protein with obvious specificity for methylated and
hemi-brominated DNA contains discontinuous deoxy guanosine-deoxycytidine residues
[13]. ADAR1 carry two binding motifs for Z-DNA, Z
A few numbers of investigations were completed to show the interface
between the solution of DNA and Z
Kim et al. in 2005 developed a DNA duplex with 15 bp and with
two hanging nucleotides [17]. This DNA duplex is co-crystallized with the
Z
Another investigation also proved that B-Z DNA junction can be produced
by oligomeric sequences in the aqueous solution at 3 M or high salt
concentration. The 5.5 M NaCl with a 95 mM combination induces the A-T sequence
into the Z-DNA conformation [18]. This study re-establishes that when NiCl
In living body, Z-DNA can form and role as a dynamic component in
various genome’s metabolic courses under certain biological circumstances [21].
Z-DNA is used in many precise activators or repressors enrolment for directive
gene countenance, genome uncertainty control [22]. Another study proved that in
cells of mammal’s ZFS fetch genetic uncertainty. Repair mechanism can proceed
with the Z-DNA development in the mammal’s body, which creates a large genomic
alteration. These sorts of changes are relevant to the breakage and translocation
near ZFS in human lymphoma and leukaemia [9]. In humans, Z-DNA links with the
transcription of the c-myc genes, which means when the Z-DNA development is
turned off the cell gives a signal as a result, c-myc transcription also starts
to down-regulate [23]. In the same way, Z-DNA development is also associated with
the corticotropin-releasing hormone (CRH) gene transcription [37]. On the other
hand, the human body also shows the activation of the Nrf2 gene which is relevant
to the HO-1 gene’s promoter, which allied with Z-DNA development [24]. A few
numbers of immunoglobulin-related genes (example-ETV6) are enriched by the Z-DNA
sequence. But in blood cancer, these genes are related to translocation of the
chromosome [22]. Interferonopathies disease like Aicardi-Goutières Syndrome
is caused by Mutation, which reduces p150 Z-binding with impaired enzymatic
activity. This is induced by dsRNAs and most commonly these dsRNAs derive from
Alu retroelement. The Z-DNA and Z-RNA both are essential for limiting Alu
retroelement intrusion of primate genomes [38]. Z-DNA provides a base for
therapeutically reducing the chances of Arthritis, Cancer, and cardiac
hypertrophy. This role is believed to be arbitrated by the downregulation of
ADAM-12. It was observed that ADAM-12 protein expression is raised when there are
pieces evidence of arthritis, cancer, and cardiac hypertrophy. Whereas ADAM-12
expression level Is exceptionally low in certain adult tissue. The regulation of
ADAM-12 is related to the highly conserved region containing a stretch of
dinucleotide repeat sequence and known as negative regulatory element (NRE),
which serves as a repressor of ADAM-12 expression. There is a certain Z-DNA
binding protein-like MeCP2. It modulates the ADAM-12 repression by recruiting NF1
transcriptional factors. Loss of ZFS leads to a low level of MeCP2 which results
in metastatic breast cancer [22, 39]. Apart from this, HIF1
Z-DNA is a double-helical structure that preserves antiparallel backbone of sugar-phosphate chains with Watson Crick pairing. Despite that, it has a contour which is fundamentally dissimilar from B configuration of DNA. Two-dimensional Gel Electrophoresis offers us a powerful method to examine the super helicity-induced physical revolution in the DNA. Besides this, B-Z conversion is also designated here. One of a reasons for transition is a cause of free unfavourable energy. Affected advances are unrestricted from the uniting effect of genomics, human genetics, biophysics, and molecular studies on non-B-DNA configurations through mutation causing agents, intricate in Genetic diseases. Autoimmune processes may be suspected in all clinical conditions where specific anti-Z-DNA antibodies are found, but for further investigation, larger population is wanted to prove such an immunological hypothesis. Future prominence will challenge to tune the acceptance of the non-B-DNA configurations at a definite location of genes to correlate this behavior extra thoroughly with the generation reposition terminuses. Also, the analysis to recognize the kind of non-B-DNA structures that obtain certain sort of mutations and the fascinated enzyme on the evolution of therapeutics, to ameliorate the disturbing corollaries of these disorders.
PC and RR conceptualize this review article. RR analyzed and interpreted the information regarding Z-DNA structure and B-Z DNA transition. PC performed a study on B-Z DNA hybrid junction formation and effects of Z-DNA on human disease and was a major contributor in writing the manuscript. AC developed the figure based on available data. PC prepared the final draft of the manuscript under the supervision of JS. All authors read and approved the final manuscript.
The work reported here in the manuscript is original and free from any plagiarism. All the data in the article are real and authentic. All the co-authors have read and agree to publish all the items listed above.
Reetabrita Roy and Pallab Chakraborty contributed equally to this article.
We don’t have any funding support from any organizational or institutional level. On behalf of all listed authors, the corresponding author declares that there is not any sort of financial and non-financial conflict of interest in the subject materials mentioned in this manuscript.
The authors declare no conflict of interest.
A