








1 Environmental Biotechnology Laboratory, Institute of Biochemistry and Physiology of Plants and Microorganisms, Saratov Scientific Centre of the Russian Academy of Sciences (IBPPM RAS), 410049 Saratov, Russia
Abstract
Ligninolytic fungi are promising organisms in developing bioremediation technologies due to their ability to degrade various pollutants. Fungi and their extracellular enzymes in soil inevitably collide with metabolites produced by other organisms. Here, we investigated the effect of some natural metabolites on the degradation of a model mixture of polycyclic aromatic hydrocarbons (PAHs) by the fungus Pleurotus ostreatus var. Florida.
Fungus was grown in the liquid medium containing PAHs with or without the addition of natural metabolites. The degraded PAHs and the identification of metabolites were checked using high-performance liquid chromatography (HPLC). Enzymatic activities were measured spectrophotometrically using test substrates. The metabolite effects on the pure laccase and versatile peroxidase were also checked. All experimental treatments were analyzed using Excel 2019 (Microsoft Office 2019, USA).
Indole-3-acetic acid (IAA) and salicylic acid increased PAH degradation by 25–70%. However, tryptophan, a precursor to IAA biosynthesis, slightly increased the degradation of only three-ring PAHs. The tested flavonoids reduced the PAH degradations, which may have resulted from the inhibition of mycelial growth by these compounds. Ferulic and cinnamic acids, precursors to lignin biosynthesis, also inhibited PAH degradation. Of the tested fungal metabolites, only veratryl alcohol promoted PAH degradation: the four-ring PAHs became more accessible to fungal degradation (43.5 and 38.1% for fluoranthene and pyrene, respectively). Oxalic and malonic acids, the most actively produced fungal organic acids, reduced the degradation of all PAHs but fluoranthene. HPLC led us to identify 9,10-phenanthrenequinone, 9,10-anthraquinone, and 9-fluorenone as the main metabolites of PAH degradation. P. ostreatus is a strong producer of extracellular laccases and peroxidases, whose involvement in PAH degradation is also well known. In our study, the most vigorous laccase inducers were tryptophan and ferulic acid (40 and 60%, respectively), whereas IAA and salicylic acid were weaker inducers (about 20%). Ferulic and salicylic acids increased versatile peroxidase activity by 1.5–2 times, whereas other effectors reduced it to varying degrees.
These results are important for developing environmental biotechnologies that combine phyto- and mycoremediation.
Keywords
- polycyclic aromatic hydrocarbons
- degradation
- fungi
- laccase
- versatile peroxidase
- flavonoids
- phenolic compounds
- organic acids
Bioremediation is a promising green biotechnology for the cleanup of contaminated soils and water, which can successfully compete with traditional but more expensive remediation methods. Bioremediation is based on the ability of bacteria, plants, algae, and fungi to degrade, transform, accumulate, or immobilize various organic and inorganic pollutants, thereby reducing pollution [1]. Mycoremediation, a type of bioremediation, is based on the ability of fungi to utilize a wide range of pollutants through an extracellular pool of nonspecific enzymes. Fungi have unique properties that make them ideal for mycoremediation: (1) The fungal mycelial network can penetrate soils and access soil pores. (2) The multicellular fungal mycelium can act as a unit, grow around barriers, recycle dead hyphae, and grow toward nutrient-rich soil areas. (3) Fungal hyphae show indeterminate growth, and there is no limit on cell division in the hyphae of a mycelium as long as resources are available in the substrate. (4) Many fungal species can form rhizomorphs, which grow through vast distances under inhospitable environmental conditions, permitting colonization of areas far from the original mycelial source. (5) Fungi are more resistant than bacteria to high pollutant concentrations. (6) Fungi produce many bioactive compounds, including exoenzymes with low substrate specificity. (7) Fungi generally depend on the extracellular degradation of macromolecules, with many enzymes being produced and secreted into the substrate [1, 2].
Although many fungi can metabolize pollutants, strong natural lignin degraders are the most promising for mycoremediation. These are equipped with systems of phenol oxidases, including laccases, lignin peroxidases, and manganese peroxidases (Mn-peroxidases). These sets of enzymes have evolved specifically to degrade lignin. However, owing to the complexity of the lignin molecule, the enzymatic breakdown mechanisms are unspecific yet are likely to involve the formation of free radical ions at various levels. This explains why white-rot fungi can degrade a wide variety of organic pollutants in addition to lignin [1, 2, 3, 4].
Species of the genus Pleurotus are often considered the most promising for mycoremediation. These are widely grown, and their spent mycelia (after edible fruit bodies are obtained) can be used as an economical source of inoculum for application to contaminated soil. In addition to nutritional and therapeutic properties, they can convert agricultural residues into useful products and degrade a wide range of pollutants alongside lignin. This latter property is associated with their production of an extracellular nonspecific oxidative enzyme system, including laccases, Mn-peroxidases, and versatile peroxidases [2, 5, 6]. These enzymes degrade and transform polymeric substances into oxidized products that cells can easily absorb [3, 4].
The use of associations of different organisms (fungi, bacteria, and plants) in bioremediation has recently been discussed [7, 8, 9]. In this case, the mutual influence of these organisms is inevitable, including through their produced metabolites. In particular, compounds produced by other organisms, such as plants and soil bacteria, affect extracellular fungal enzymes in soil. In addition, these substances can induce or inhibit the production of these enzymes; thus, this influence must be considered when fungi are used for bioremediation [10].
Polycyclic aromatic hydrocarbons (PAHs) are among the most dangerous pollutants that Pleurotus spp. can degrade and mineralize [5, 11, 12]. PAHs consist of three or more benzene rings in a linear, angular, or cluster arrangement and have mutagenic and carcinogenic properties. PAHs enter the environment due to natural processes and human activities [2, 12]. The degradation and detoxification of PAHs involve oxidative nonspecific enzymes such as laccases and peroxidases, which are produced by Pleurotus spp. [5, 12].
This study aimed to identify the effects of natural metabolites from plants, bacteria, and fungi on the degradation of a mixture of PAHs and the production of ligninolytic enzymes by P. ostreatus var. Florida in a model system.
Pleurotus ostreatus var. Florida (IBPPM 540) was obtained from the IBPPM RAS Collection of Rhizosphere Microorganisms (http://collection.ibppm.ru; accessed on 23.07.2022).
A modified basidiomycetes-rich medium was used to store the fungus and obtain an
inoculum (g/L): NH4NO3, 0.724; KH2PO4, 1.0;
MgSO4
After 14 days, the contents of the flasks were extracted three times using 5 mL
of chloroform (HC quality, Chimmed, Moscow, Russia), and the extracts were evaporated.
The sorption of PAHs by the fungal mycelium did not exceed 5–7%. The PAH
content was analyzed using high-performance liquid chromatography (HPLC) on an
Agilent Technologies 1220 Infinity II LC chromatograph (Agilent Technology,
Waldbronn, Germany) equipped with a 4.6
The IAA content was analyzed by HPLC on the Agilent Technologies chromatograph
equipped with a 4.6
Laccase activity was measured using the oxidation rate for
2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS;
Fluka, Buchs, Switzerland) at 436 nm (
Electrophoretically homogeneous laccase and versatile peroxidase from P. ostreatus var. Florida were prepared in accordance with the previously published purification protocols [18, 19].
To examine the effect of different compounds on enzyme activities, we also added IAA, salicylic acid, ferulic acid, cinnamic acid, tryptophan, rutin, morin, quercetin, oxalic acid, malonic acid, or veratryl alcohol (final concentration, 100 µM) to the reaction mixture. In these experiments, enzyme activities were tested as described above.
All experiments on the enzymatic oxidation of PAHs were carried out in reaction test tubes containing 1 mL of 50 mM acetate buffer (pH 4.5) (HC quality, Chimmed, Moscow, Russia) and 1% (v/v) acetonitrile (HPLC-quality, Fluka, Buchs, Switzerland). In the case of versatile peroxidase, 200 µM H2O2 (HC quality, Chimmed, Moscow, Russia) was also added. A 100 µL portion of laccase or versatile peroxidase was added to each test tube to a final concentration corresponding to 1 U/mL. The PAHs were dissolved in acetonitrile and added to a final concentration of 10 µM. Effectors (ABTS, ferulic acid, or quercetin) were added to a final concentration of 100 µM. The closed reaction test tubes were incubated at 30 °C for the desired period. The PAHs and their oxidation products were extracted using 2 mL of chloroform, evaporated, and redissolved in 100 µL of acetonitrile. Control samples were prepared similarly, but the enzyme was deactivated by boiling for 10 min before being added.
All experimental treatments and all analyses were repeated at least three times. The obtained experimental data were statistically processed, average values were calculated, and the standard deviation (SD) was used for comparisons. Calculations were performed using Microsoft Excel 2019 (Microsoft, Redmond, WA, USA).
P. ostreatus var. Florida was previously described as a strong degrader of PAHs [20]. We investigated the effect of selected natural plant, bacterial, and fungal metabolites (IAA, salicylic acid, ferulic acid, cinnamic acid, tryptophan, rutin, morin, quercetin, oxalic acid, malonic acid, and veratryl alcohol) on the degradation of a model mixture containing three- (anthracene, fluorene, phenanthrene) and four-ring (pyrene, fluoranthene) PAHs by this fungus.
Phenolic compounds, including simple phenols, coumarin, lignin, condensed and hydrolyzable tannins, and flavonoids, may play a part in plant responses to environmental stress. These compounds are also believed to be important in detoxifying environmental pollutants by promoting the production of enzymes such as peroxidases and oxidases, which are involved in the metabolism of phenolic compounds [21]. High contents of phenolic compounds (1–6 mg/g dried mycelium) and flavonoids (0.9–3 mg/g dried mycelium) were found in some basidiomycetes, including P. ostreatus [22]. Salicylic acid with phenolic hydroxyl is a metabolite produced following PAH degradation by several bacteria and fungi [23]. Fungi, bacteria, and plants produce oxalic acid, which performs various functions in these organisms. In particular, oxalate may be implicated in bacterial–fungal communications [24]. Oxalic acid secreted by fungi and malonic acid promotes Mn-peroxidases reactions during lignin degradation [5]. Ferulic acid is the most abundant hydroxycinnamic acid in plants and is not only a precursor for lignin biosynthesis [25] but is also formed during fungal lignin degradation.
The addition of metabolites had little effect on the dry mass of mycelium in the PAH-free variants (Fig. 1). The exceptions were IAA and rutin, which reduced this indicator by 16.3 and 36.7%, respectively, and ferulic acid, which slightly increased it (by 17.8%).
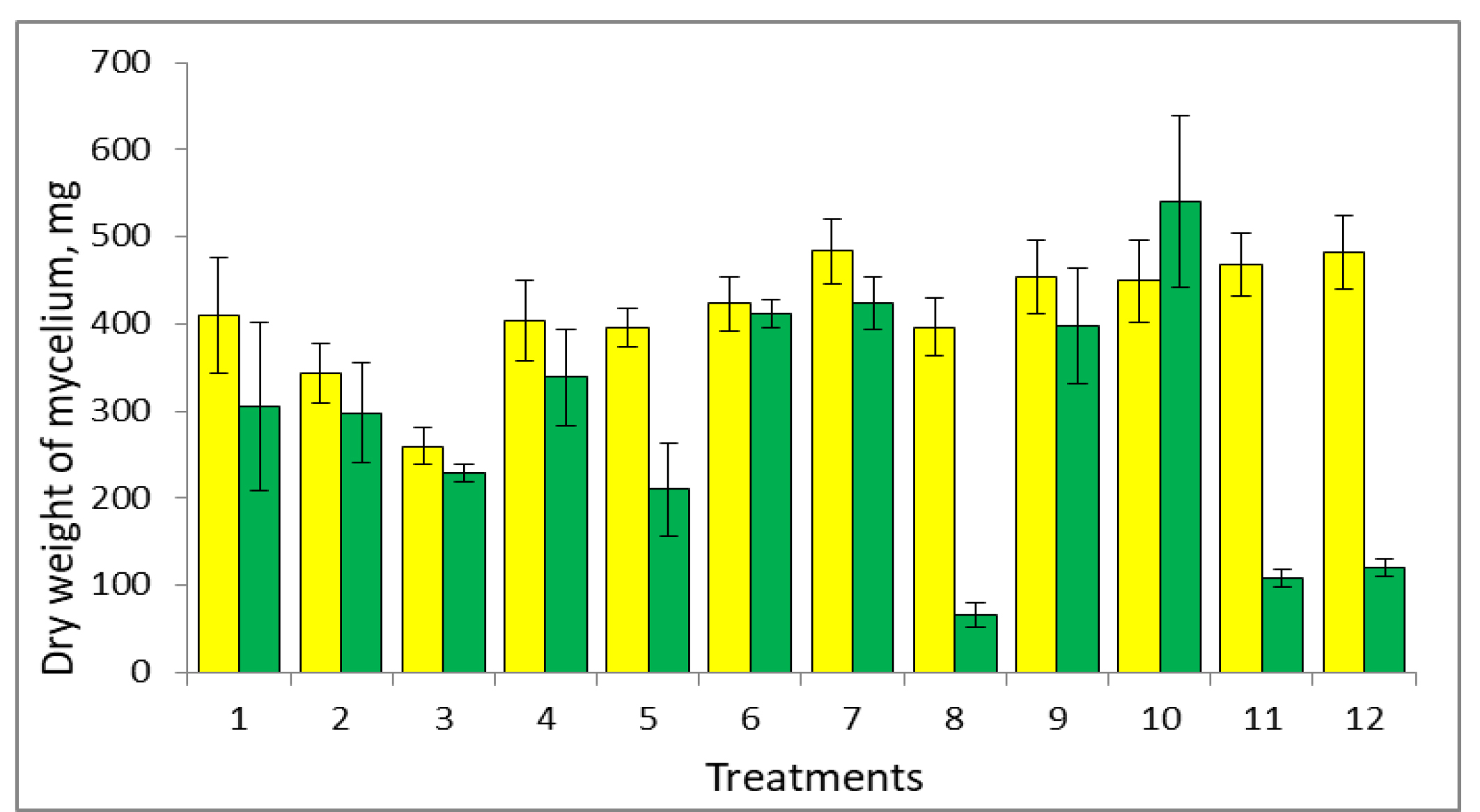 Fig. 1.
Fig. 1.
Effect of plant and fungal metabolites on the growth of
P. ostreatus var. Florida.
As evidenced by decreased mycelial dry weight, the PAHs inhibited fungal growth, contrasting with similar treatments without PAHs. The exception was salicylate, whose presence increased the dry weight by approximately 30% compared to corresponding treatments without PAHs. Analysis of mycelial dry weight in the presence of the PAHs and metabolites showed that quercetin and cinnamic acid reduced this indicator by 31.1% and 78.2%, respectively. Mycelial production increased in the presence of the PAHs and salicylic and ferulic acids, morin, tryptophan, and veratryl alcohol compared to corresponding treatments without metabolites (Fig. 1). Apparently, in this case, these plant and fungal metabolites neutralize, to some extent, the negative effect exerted on fungal growth by the PAH metabolites. Interestingly, veratryl alcohol is a known stabilizer of fungal ligninolytic enzymes [25, 26].
In the control treatment (without effectors), P. ostreatus var. Florida could only metabolize the three-ring PAHs (fluorene, phenanthrene, anthracene), while the four-ring PAHs (pyrene and fluoranthene) were poorly available (Table 1). Table 1 shows that only IAA and salicylic acid increased the degradation of all PAHs included in the study by 25–70%. However, even the four-ring pyrene and fluoranthene became more accessible to fungal degradation. The presence of tryptophan, a precursor to bacterial and fungal biosynthesis of IAA, only slightly increased the degradation of the three-ring PAHs. In contrast, the loss of the four-ring PAHs remained low. Both tested flavonols (morin and quercetin) and their glycoside (rutin) reduced the degradation of all PAHs, which may have resulted from the inhibition of mycelial growth by these compounds. Ferulic and cinnamic acids, precursors to lignin biosynthesis, also inhibited PAH degradation by P. ostreatus var. Florida, although these are known inducers of ligninolytic enzymes in this fungus [25, 26].
| Treatment | PAH disappearance (%) | ||||
| Fluorene | Phenanthrene | Anthracene | Fluoranthene | Pyrene | |
| Without effector | 70.3 |
58.3 |
40.9 |
17.7 |
1.9 |
| IAA | 85.9 |
68.9 |
85.2 |
73.9 |
69.0 |
| Rutin | 63.5 |
50.0 |
35.2 |
0 |
0 |
| Morin | 62.2 |
29.2 |
0 |
0 |
0 |
| Quercetin | 41.9 |
13.9 |
0 |
0 |
0 |
| Tryptophan | 82.4 |
72.2 |
48.8 |
11.3 |
18.1 |
| Ferulic acid | 37.8 |
47.2 |
47.7 |
0 |
0 |
| Cinnamic acid | 24.3 |
43.1 |
14.8 |
0 |
0 |
| Veratryl alcohol | 81.1 |
81.9 |
84.1 |
43.5 |
38.1 |
| Salicylic acid | 87.8 |
87.5 |
86.4 |
80.6 |
80.0 |
| Malonic acid | 25.7 |
16.6 |
28.1 |
14.5 |
0 |
| Oxalic acid | 31.1 |
16.7 |
28.4 |
14.5 |
0 |
Among the natural fungal metabolites used, only veratryl alcohol, a known inducer of the ligninolytic enzymes, promoted the degradation of the PAH mixture. However, the four-ring PAHs also became more accessible to fungal degradation (43.5% and 38.1% for fluoranthene and pyrene, respectively). Oxalic and malonic acids, the most actively produced organic acids by fungi, reduced the degradation of all PAHs except fluoranthene, whose content decreased by more than 4 times (Table 1); this identified effect requires further research.
HPLC identified 9,10-phenanthrenequinone, 9,10-anthraquinone, and 9-fluorenol as the main metabolites of PAH degradation, irrespective of the effector used or its administration. The formation of the corresponding quinones during PAH degradation is well-known and involves oxidative enzymes such as laccases and peroxidases [2, 5, 12].
IAA is the most important naturally occurring auxin, which is involved in many aspects of plant growth and development. Most plant growth-promoting rhizobacteria (or fungi) (PGPR or PGPF) synthesize IAA, and their effects on plants mimic those of exogenously applied IAA. Indeed, the l-tryptophan amino acid is a physiological precursor for auxin biosynthesis in plants and microbes [27]. Fungi, including Pleurotus members, are also known to produce IAA [28].
Our experiments evaluated the effect of natural metabolites on IAA production by P. ostreatus var. Florida during PAH degradation. IAA production was most vigorous in the presence of l-tryptophan and salicylic acid (0.4 and 0.68 µg/mL, respectively; Fig. 2). Apparently, tryptophan can also be a precursor for IAA biosynthesis in fungi and bacteria. Without PAHs, the production of IAA by P. ostreatus was non-significant and within the defined error level of the method.
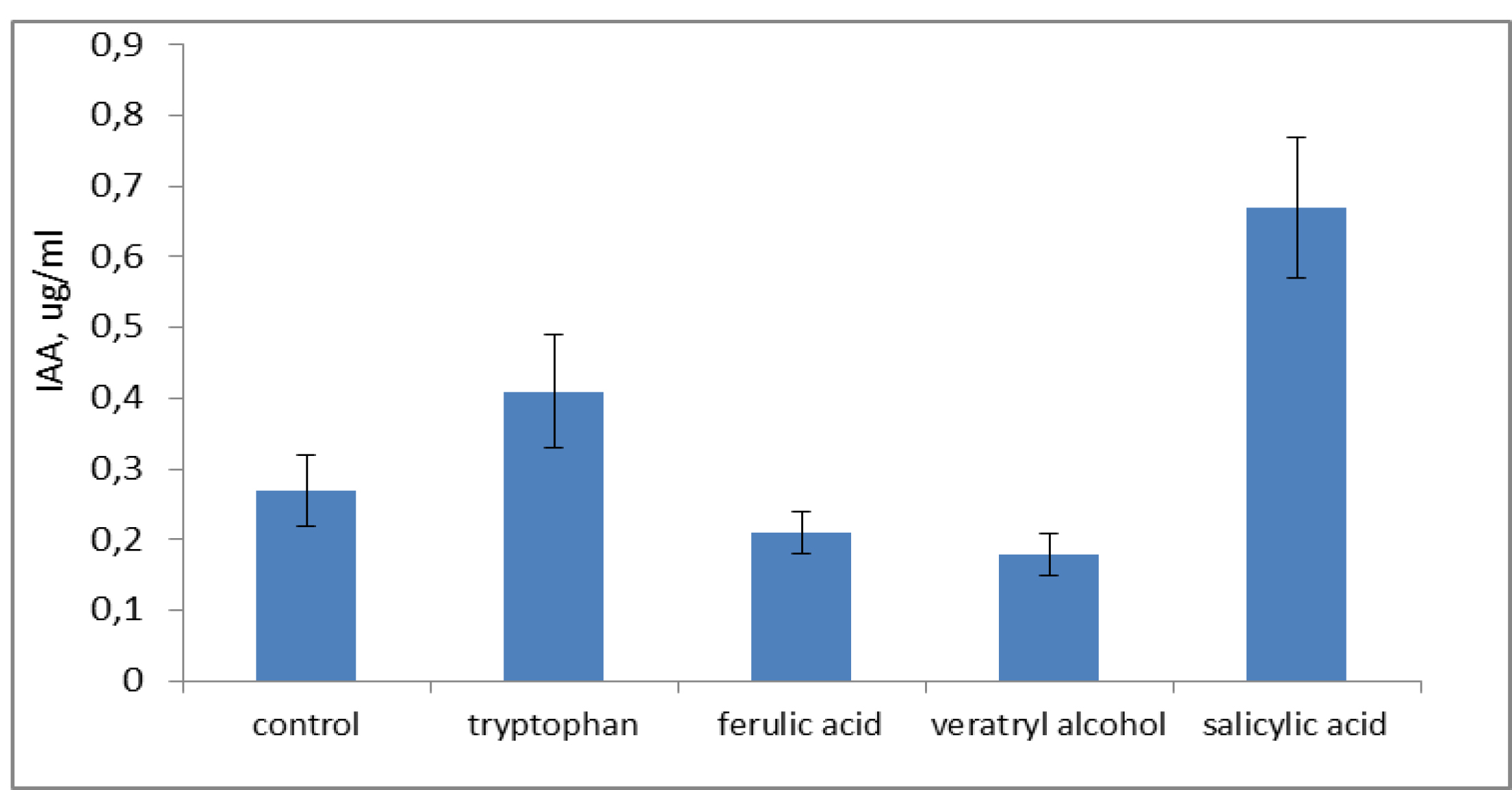 Fig. 2.
Fig. 2.
Effect of plant and fungal metabolites on IAA production by P. ostreatus var. Florida during PAH degradation. Control, without effector.
It is difficult to overestimate the role of extracellular enzymes, particularly oxidoreductases, in degrading those difficult-to-degrade pollutants for bacterial intracellular enzyme systems. However, partial oxidation of persistent pollutants, such as PAHs, by extracellular oxidative enzymes yields more polar and hydrophilic products, which can easily be utilized by bacterial intracellular enzyme systems [3, 4]. Among the most important extracellular enzymes involved in the degradation of pollutants in different environments are fungal laccases and peroxidases [4]. P. ostreatus is a strong producer of extracellular laccases and peroxidases, whose participation in PAH degradation is well known [12, 29]. We investigated the effect of a selection of natural metabolites on the production of laccase and versatile peroxidase by P. ostreatus var. Florida during PAH degradation. Tryptophan and ferulic acid (40–60%) were the strongest laccase inducers, which was expected since aromatic compounds are known to induce fungal laccases. IAA and salicylate induced this enzyme to a lesser extent (about 20%; Fig. 3). A similar picture emerged during PAH degradation: the most vigorous laccase inducers were tryptophan and ferulic acid (Fig. 3).
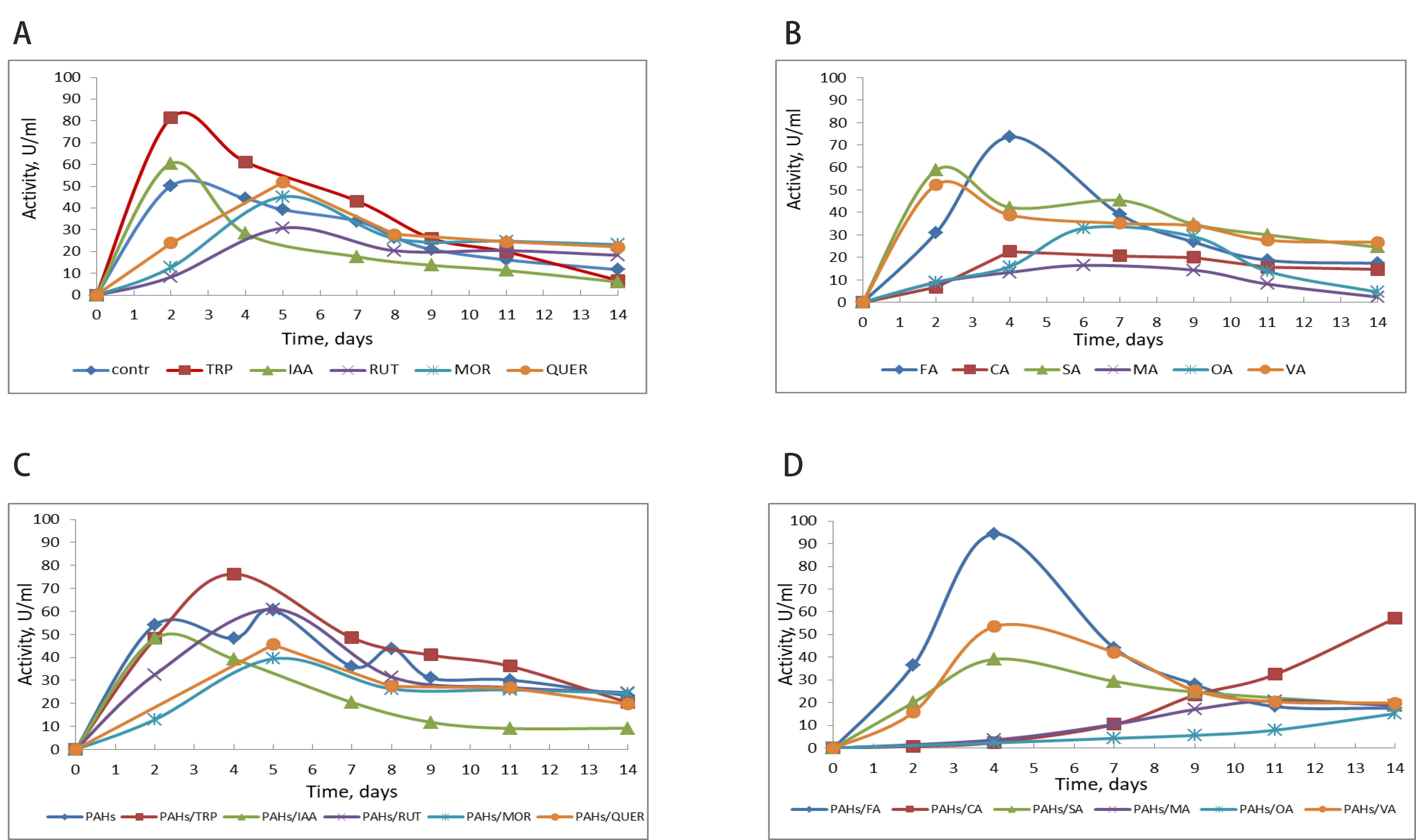 Fig. 3.
Fig. 3.
Effect of metabolites on laccase production by P. ostreatus Florida without PAHs (A,B) and during PAH degradation (C,D). Contr, control without effector; TRP, tryptophan; IAA, indole-3-acetic acid; RUT, rutin; MOR, morin; QUER, quercetin; FA, ferulic acid; CA, cinnamic acid; SA, salicylic acid; MA, malonic acid; OA, oxalic acid; VA, veratryl alcohol.
Versatile peroxidase is another enzyme identified in P. ostreatus var. Florida, which combines the properties of lignin and Mn-peroxidases. Generally, the activity of this enzyme was 2.5–3 times lower than that of laccase, regardless of the effectors used (Fig. 4). Ferulic acid and salicylic acid both induced versatile peroxidase activity by 1.5–2 times, whereas the other effectors reduced it to varying degrees (Fig. 4). During PAH degradation, versatile peroxidase activity increased in the presence of ferulic, salicylic, and cinnamic acids, whereas the other effectors inhibited it (Fig. 4). Organic acids, such as oxalate and malonate, which are secreted by fungi, are known to promote Mn-peroxidases reactions by stabilizing Mn3+, which diffuses from the surface of the enzyme and oxidizes the insoluble terminal substrate, lignin [5, 30]. Our studies showed no promotion of versatile peroxidase by these acids. In contrast to Mn-peroxidases, these organic acids are unnecessary for effective catalysis by versatile peroxidase.
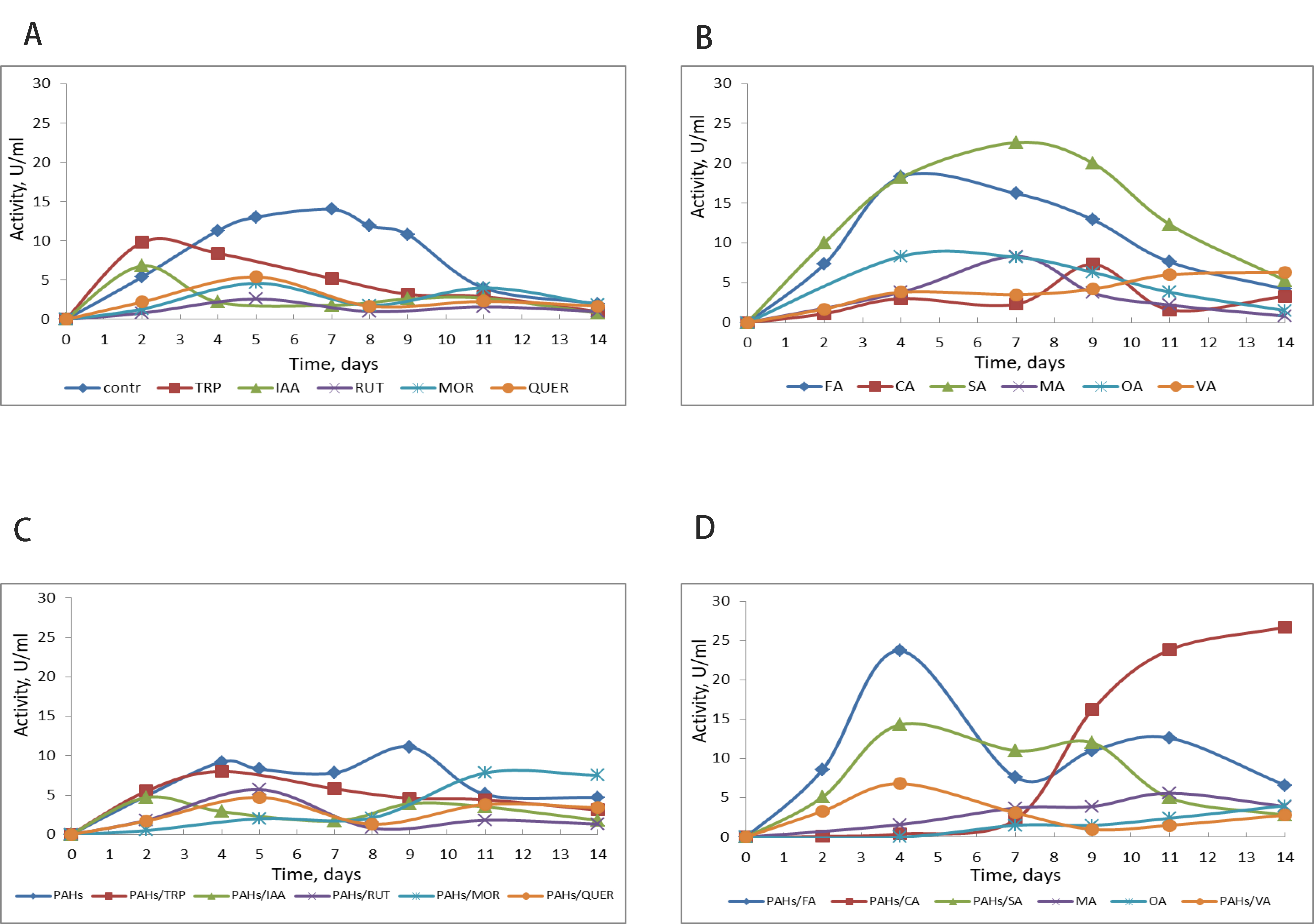 Fig. 4.
Fig. 4.
Effect of metabolites on versatile peroxidase production by P. ostreatus Florida without PAHs (A,B) and during PAH degradation (C,D). Contr, control without effector; TRP, tryptophan; IAA, indole-3-acetic acid; RUT, rutin; MOR, morin; QUER, quercetin; FA, ferulic acid; CA, cinnamic acid; SA, salicylic acid; MA, malonic acid; OA, oxalic acid; VA, veratryl alcohol.
The identified effects may have resulted from changes in the production of extracellular enzymes and from the influence of the tested metabolites on enzyme activity. For this purpose, we purified laccase and versatile peroxidase according to previously developed protocols [18, 19]. Morin and quercetin inhibited the laccase almost completely (Fig. 5). Furthermore, the nearly 45% increase in laccase activity in the presence of veratryl alcohol may have resulted from the stabilizing effect of this naturally occurring fungal metabolite, which has been described extensively previously [26]. Versatile peroxidase was unaffected by the flavonoids used and even increased in the presence of quercetin. A stimulatory effect mediated by ferulic acid was also noted (Fig. 5).
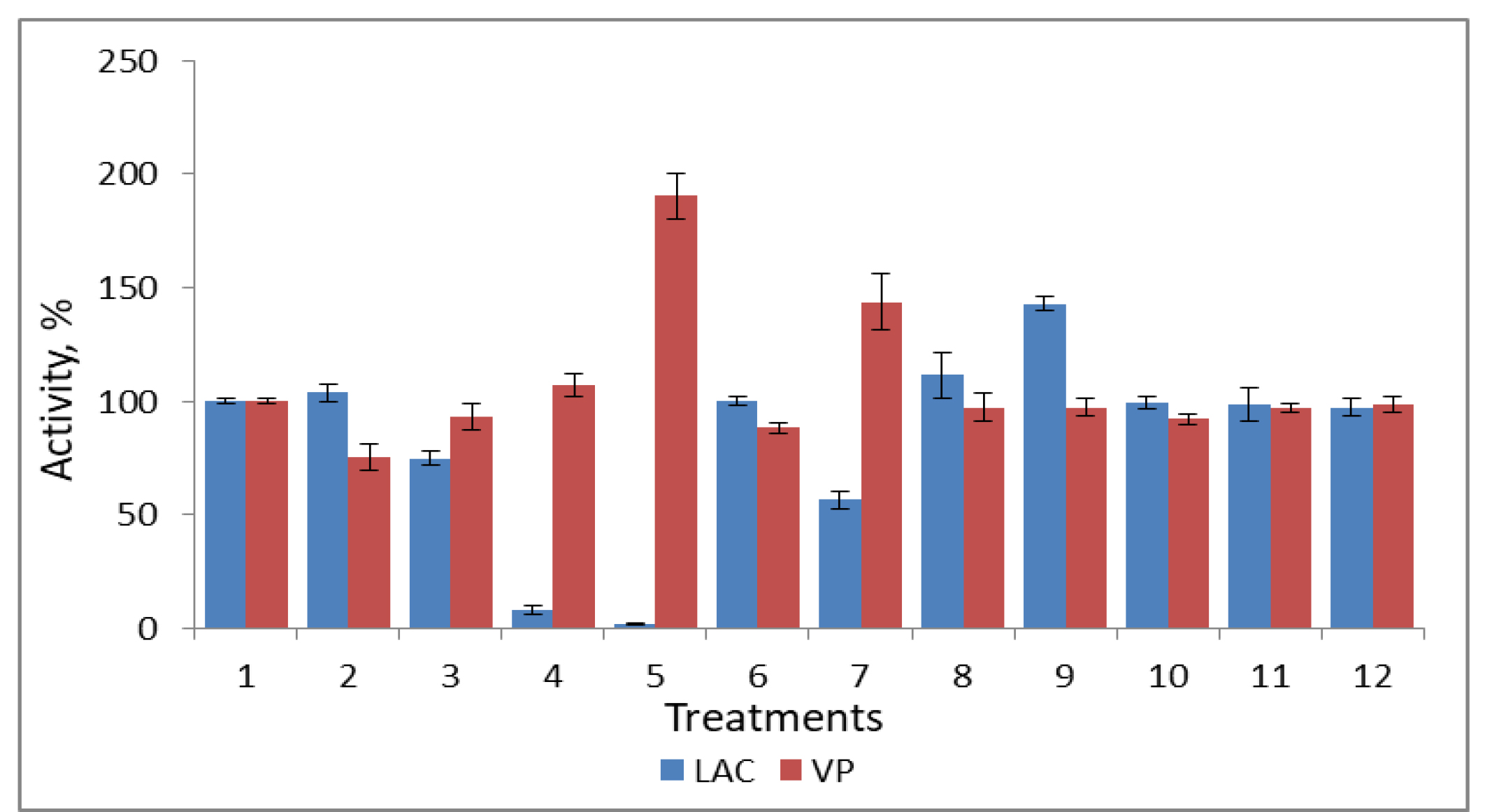 Fig. 5.
Fig. 5.
Effect of plant and fungal metabolites on the activities of
purified laccase (
Laccase directly oxidizes phenolic compounds, including lignin components, whereas versatile peroxidase can act on more recalcitrant nonphenolic aromatic compounds. Mediators, or co-oxidants, not only increase the catalytic ability of these enzymes but also largely expand their substrate scope to include those with higher redox potential or more complicated structures [30].
As a synthetic laccase substrate, ABTS can act as a redox mediator, allowing the oxidation of nonphenolic aromatic compounds, including PAHs, which are inaccessible to direct laccase oxidation. In addition to synthetic compounds, such as ABTS, 1-hydroxybenzotriazole, and violuric acid, some natural metabolites can act as redox mediators. For example, to overcome the redox potential barrier, Pycnoporus cinnabarinus produces the metabolite 3-hydroxyanthranilate, which mediates the oxidation of nonphenolic substrates by the laccase in this fungus [5].
Natural compounds that contain sulfhydryl groups, such as methionine, cysteine, and reduced glutathione, also act as mediators [31]. Indeed, fungal laccases can degrade some PAHs in the presence of phenolic compounds similar to those produced by ligninolytic fungi during lignin and humus degradation. The potency of these natural mediators, including vanillin, acetovanillone, acetosyringone, syringaldehyde, 2,4,6-trimethylphenol, p-coumaric acid, ferulic acid, and sinapic acid, was comparable to that of the synthetic mediators ABTS and 1-hydroxytriazole. The best results were obtained with 4-hydroxybenzoic acid and 4-hydroxybenzyl alcohol, which, along with other aromatic compounds, are produced and secreted by fungi. Meanwhile, a system consisting of laccase and hydroxybenzoic acid was shown to oxidize highly condensed PAHs, such as benzo[a]pyrene [31].
Our studies showed that the natural metabolite ferulic acid was an equally effective redox mediator of the laccase oxidation of PAHs (Fig. 6). Apparently, natural metabolites, which act as mediators, can be important in natural PAH degradation by fungi. Moreover, an increase was noted in PAH oxidation in the versatile peroxidase/redox mediator system. The oxidation of phenanthrene in the presence of quercetin or ferulic acid increased by 2 times compared with the control treatment (no mediator), while that of pyrene and fluoranthene increased by 4 times (Fig. 6). In this case, the identified effects can be attributed to both the stabilization of the enzyme in the presence of these substances and to the reactions catalyzed by the enzyme. However, the mechanism through which PAH oxidation occurs via versatile peroxidase in the presence of these compounds will be the subject of future research.
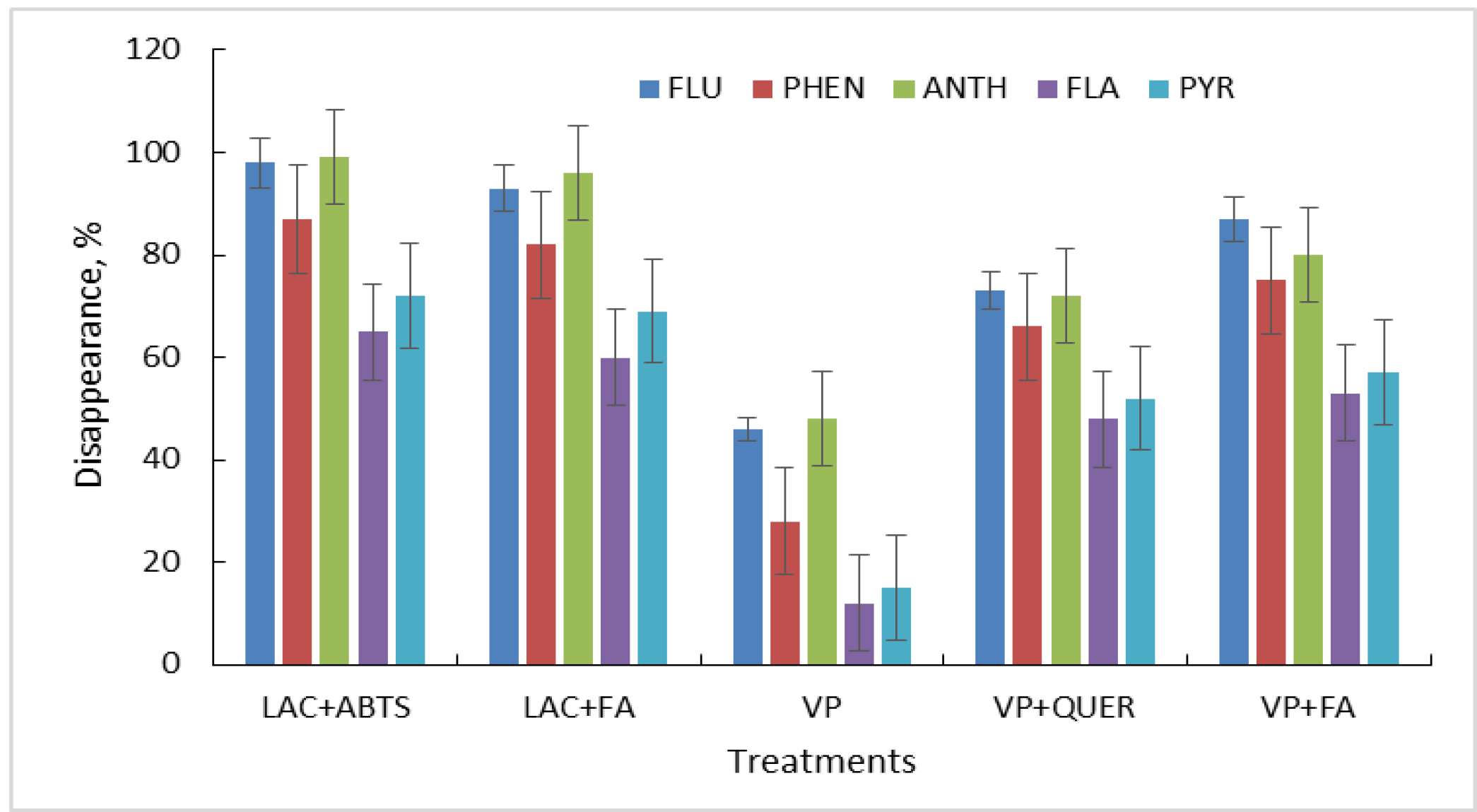 Fig. 6.
Fig. 6.
Effect of plant and fungal metabolites on the oxidation of PAHs by purified laccase (LAC) and versatile peroxidase (VP). These effects were assessed in the presence of 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS), ferulic acid (FA), quercetin (QUER), fluorene (FLU), phenanthrene (PHEN), anthracene (ANTH), fluoranthene (FLA), and pyrene (PYR).
The efficacy of pollutant biodegradation depends on the type of pollutant and fungus [3, 4]. The factors affecting degradation efficacy include but are not limited to the bioavailability of the compound, pH, temperature, aeration, and the presence of inhibitory compounds [12]. When different organisms are cocultured, inter-organismal relationships are added to these factors [8]. Importantly, degradation efficacy also depends on effectors, such as the natural metabolites of fungi, plants, and bacteria, which affect this process in a biocenosis. This study has tested the effect of some well-known natural metabolites on the degradation of a PAH mixture by P. ostreatus var. Florida, as well as the production and activity of extracellular laccase and versatile peroxidase, which catalyze the initial attack on the PAH molecule. In addition to the direct effect of these compounds on fungal growth and PAH degradation, we found inhibitory (for laccase, morin, and quercetin) and stimulatory (for versatile peroxidase, quercetin, and ferulic acid; for laccase veratryl alcohol) effects on the activity and stability of the extracellular enzymes. Moreover, some compounds (e.g., ferulic acid) can act as redox mediators, allowing and/or accelerating PAH oxidation.
PAHs, polycyclic aromatic hydrocarbons; IAA, indole-3-acetic acid; TRP, tryptophan; RUT, rutin; MOR, morin; QUER, quercetin; FA, ferulic acid; CA, cinnamic acid; SA, salicylic acid; MA, malonic acid; OA, oxalic acid; VA, veratryl alcohol; FLU, fluorene; PHEN, phenanthrene; ANTH, anthracene; FLA, fluoranthene; PYR, pyrene; ABTS, 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt; LAC, laccase; VP, versatile peroxidase; HPLC, high-performance liquid chromatography; SD, standard deviation; PGPR, plant growth-promoting rhizobacteria; PGPF, plant growth-promoting fungi.
The data and materials generated during the current study are available from the corresponding author.
NP designed the research study, performed the research, analyzed the data and wrote the manuscript.
Not applicable.
We are grateful to Dr. Dmitry N. Tychinin for his assistance in preparation of the English text of this paper.
This work was carried out under research theme no. 124020100146-9 of Ministry of Education and Science of the Russian Federation.
The author declares no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBE25804.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.