

Frontiers in Bioscience-Elite (FBE) is published by IMR Press from Volume 13 Issue 2 (2021). Previous articles were published by another publisher on a subscription basis, and they are hosted by IMR Press on imrpress.com as a courtesy and upon agreement with Frontiers in Bioscience.
 , Hermawan Wibisono 1, Yin Mon Khine 1, Tasuku Harada 1
, Hermawan Wibisono 1, Yin Mon Khine 1, Tasuku Harada 11 Department of Obstetrics and Gynecology, Tottori University Faculty of Medicine, Yonago, 683-8504, Japan
Abstract
Endometriosis results from the aberrant growth of endometrium outside the inner lining of the uterine cavity. Similar to humans, the primates also menstruate and hence, the primate models constitute the gold standard for studying the pathogenesis and potential treatment for this disabling disease in women. Due to the expense in carrying endometriosis research in primates, other models have been developed for understanding the pathobiology and potential treatment of endometriosis. This includes explanting human endometrial tissues in athymic nude mice or using homologous mouse models. Here, we examine the murine models of endometriosis, the impact of forced induced inflammation on its development, similarities in the gene expression profile in the endometriotic tissues in such models with that seen in human endometriosis, and the drugs that are being used in such models as potential new treatment for endometriosis
Keywords
- Endometriosis
- Murine Endometriosis-Like Lesion
- Experimental Model
- Cystic Lesion
- Review
The exact cause of endometriosis is not certain. However, the most commonly accepted explanation is the Sampson hypothesis which suggested viable endometrial tissue flows retrogradely through the fallopian tube and into the peritoneal cavity where it can attach to and invade tissues and organs within the cavity, all of which occured in a process called retrograde menstruation (1). At least 90% of women experience retrograde menstruation, but endometriosis occurs in only 10% to 14% of women under reproductive age, suggesting that additional factors impact its etiology (2). Based on the implantation theory, we generated the homologous murine endometriosis models by transplanting uterine tissue to mimic the pelvic environment in human endometriosis and analyzed the efficacy of several drugs (Figure 1, Table 1).
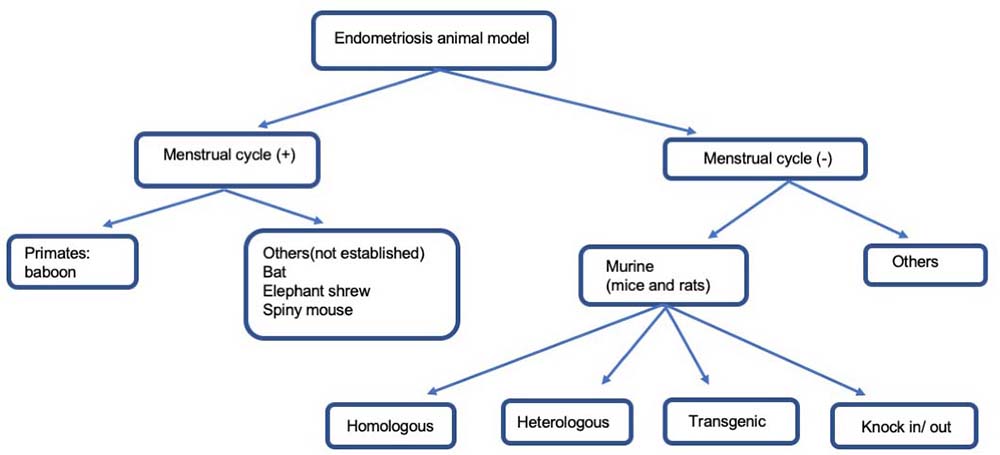 Figure 1
Figure 1A cascade of animal endometriosis mice model. Endometriosis animal model is divided into 2 groups without and those with cyclical menstruation.
| Drug | Characteristics of models | Reference |
|---|---|---|
| Mouse is both the donor and the host | ||
| Not tested for drug development | Syngeneic GFP transgenic mice, retrograde menstruation model | 78 |
| Not tested for drug development | Deep endometriosis mouse model | 13 |
| Not tested for drug development | Laparoscopic mouse model | 33 |
| Not tested for drug development | P-selectin KO, GFP transgenic mice | 18 |
| Not tested for drug development | GFP mice | 19 |
| Not tested for drug development | Adhesion scoring system | 79 |
| Not tested for drug development | Estrogen receptor KO | 40 |
| Not tested for drug development | SRC-1 KO | 58 |
| Omega-3 polyunsaturated Fatty acids (3-PUFA) | Transgenic Fat-1 mice (high levels of endogenous 3-PUFA) | 42 |
| Parthenolide | Large cyst formation | 30 |
| Retinoic acid | GFP mice | 59 |
| Temsirolimus | Deep infiltrating implants | 74 |
| Tricostatin A | Auto-transplant model | 65 |
| Bazedoxifene | No description available | 31 |
| Celecoxib, Rosiglitazone | No description available | 75 |
| Leptin receptor antagonist | No description available | 39 |
| Resveratol | No description available | 70 |
| Statins | No description available | 73 |
| Symvastatin | No description available | 9 |
| Vitamin D receptor Agonist | No description available | 72 |
| Human is the donor and mouse is the host | ||
| ENMD-1068, a protease-activated receptor 2 antagonist | Noninvasive fluorescent mouse model | 77 |
| Pentoxifylline | No description available | 76 |
| Cabergoline | Assessment nerve fibers | 71 |
Current medical therapeutic agents such as GnRH agonist, progestins, and androgen supplements are used to decrease the ovarian estrogen production and counteract estrogen effects as this hormone initiates endometriosis (3). However, these therapeutic agents have undesirable side effects that limit their long-term use (2). Likewise, the scope of surgical treatment is also limited by a high recurrence rate, which may eventually lead to extreme measures, such as removal of the uterus and ovaries. Today, as science advances, new potential source of drugs from natural resources—for example plants, which may provide less toxicity and side effects—are explored. Thus, animal models to elucidate the mechanisms of endometriosis and develop therapeutic agents to prevent the progression of the disease are very important.
Animal models, which are used in the early stages of drug development, usually rely on non-menstruating models with the induced endometriosis-like lesions. One of the major limitations in endometriosis research is the paucity of robust animal disease models. Ideally, a disease models should mimic human disease and allow scientific investigation of the effects of both intrinsic factors, such as genes, and extrinsic factors, such as environmental factors on disease progression (4).
Several animal models of endometriosis have been established, most of which consisted of transplantation of endometrium into the peritoneal cavity, which is by far the most common site of the disease. Besides rodents, primates such as monkeys that spontaneously develop endometriosis or that have been intraperitoneally transplanted with endometrium can be used as a good animal model for the development and optimization of drug candidates. As monkeys developed endometriosis spontaneously due to cyclic menstrual periods—this model is undoubtedly the most reliable one (4-16). However, laborious requirements for the maintenance of these primate animal models have inhibited the progression of potential drug developments. This being said, most researchers concentrated their studies on smaller mammalian models, such as rats and rabbit (17-22). Moreover, the murine model was considered as a possible candidate for this purpose, and it may provide advantages in terms of new therapeutic approaches (17,18,20,23-29). In recent decades, many knockout or transgenic mice have been generated. The availability of an endometriosis models in mice is crucial because it can be used to investigate some aspects of endometriosis.
Endometriosis is an estrogen-dependent disease. Thus, estrogen receptors play a significant role in the development of the disease. Moreover, among estrogen receptors, Estrogen receptor β (ERβ) showed an association with endometriosis progression. Han et al. used mouse models to observe the role of ER β in the development of endometriosis (54). This is based on the study of Burns et al. where they concluded that the expression of ERβ was required for endometriosis-like lesion in mice (42). This being said, the study used an ERBOE (ERβ over-expressed) mouse to determine the histologic and immunologic change effects of ERβ. Remarkably, they were able to elucidate that over-expression of ERβ can lead to downregulation of immune response such as Interferons, NF-κB, and other inflammatory response. Note that Interferon also plays a significant role in pregnancy, such as in embryo implantation (55) and NF-κB expression regulates the gene expression profiles for implantation and successful pregnancy (56-59). Notably, NF-κB in mouse plays an important role as its activation promotes implantation of the mouse uterus (60,61). Therefore, downregulation of NF-κB signaling by ERβ can contribute to endometriosis in women by impairing embryo implantation.
The successful utilization of mouse models to observe the effects of ERβ in the development of endometriosis open ERβ over-expressed biomarker for endometriosis. This mouse models also provides ERβ explored ER insights on the better understanding of complex molecular etiology of endometriosis induced by estrogen/ ERβ axis (62,63). Moreover, the availability of female mice (C57BL/6J) and its ease in animal laboratory handling provide a great advantage compared to other animal models.
Mice are the most commonly used animal models capable of investigating the pathophysiology of endometriosis. However, they do not spontaneously develop endometriosis. In order to induce endometriosis in mice, endometrial tissue must be transplanted into the peritoneal cavity using methods (10-13), which can be classified into two basic types, homologous and heterologous. Both models produce comparable phenotypes, which are then morphometrically evaluated.
In homologous or autologous models, endometrial tissue is transplanted into the peritoneal cavity of recipient mice and starts to grow in an estrogen-dependent manner. In almost all models, uterine endometrial fragments from a donor mouse are directly introduced via injection some by suturing (19,30-32) or inserted laparoscopically (33,34) into the peritoneal cavity of recipient mice. Our studies used a homologous mouse models of endometriosis.(47,48,53).
Care and treatment. According to the implantation theory, we made the readily available murine models which evaluate the development of endometriosis-like lesions (30). Female mice (6 weeks of age, BALB/c) were purchased. Before initiating the experiments, animals were allowed to acclimate to the following conditions for seven days. The mice were put in a controlled temperature range (72–74) on a 12-hour light, 12-hour dark cycle. They were given food and water ad libitum. Recipient mice were ovariectomized through two 0.5-cm dorsolateral skin incisions and were then divided into two treatment groups; estradiol valerate (0.5 μg/mouse·wk) in corn oil or only corn oil vehicle. The mice were dosed subcutaneously once per week for two weeks before they were induced experimental endometriosis. The donor uterus was removed en bloc after euthanasia, cleaned of excess tissue, and washed thrice in sterile phosphate-buffered saline (PBS). The uterus was slit with a linear incision longitudinally and minced (≤1.5 mm). Recipient mice were anesthetized using isoflurane/oxygen and given buprenorphine (0.1 mg/kg) for pain management. A 0.5-cm right dorsolateral incision was made, minced donor tissue (1:2 donor uterus to host ratio) in 500 μl PBS was injected into the peritoneal cavity of the recipient, and was gently massaged to disperse the tissue. An equivalent amount (~100 mg) of minced tissue was transferred into all recipients. The mice were treated for four additional weeks with estradiol valerate (E2) or the vehicle. After four weeks, the mice were euthanized with CO2, their peritoneal cavity was opened, and the endometriosis-like lesions were removed. To assess the effects of drug candidates on ectopic uterine tissue, ectopic lesions were photographed to document in situ endometriosis-like lesions. Endometriosis-like lesions were visualized, dissected, measured, and weighed. Endometriosis-like lesions were removed and were either fixed in 10% formalin or snap-frozen on dry ice and stored at −80 C until use.
In chapter six, we will describe the efficacy of SR-16234 (SR), a selective estrogen receptor modulator (SERM), parthenolide and BV6, an inhibitor of apoptosis protein inhibitor (IAP) on the murine endometriosis-like lesions by using homologous mouse models.
In heterologous models, human endometriotic lesions are transplanted into the peritoneal cavity of immune-deficient mice (35,36), and into the peritoneum of immunocompromised mice (37,38). The xenotransplantation models in the nude mouse is also used but is limited by the lack of a normal immune system. Despite the advantage of being based on human endometrial tissues, the number of endometriotic lesions developed in the heterologous models varies from one animal to another. In both models, with or without suturing the endometrial implant, the drug’s influence on the growth of endometrial or endometriotic transplants is evaluated.
However, these established endometriosis models in mice are under discussion. Indeed, mice lack a menstrual cycle and do not develop spontaneous endometriosis. It should be reviewed whether transplanting normal endometrium into the peritoneal cavity of a non-menstruating species reflects all pathophysiological aspects of human endometriosis.
The knockout and transgenic murine models provide a suitable environment to study biological factors such as the role of immune system and hormone balance that may affect endometriosis, which cause unbearable pain and possible infertility. These models also enable the evaluation of molecular mechanisms that are critical for disease initiation (39-42). With the existing limitation, additional and ideal models of endometriosis are needed. In the next section, we demonstrate the results using murine endometriosis model that expanded the capability of conducting both mechanistic and translational research.
The search for the most suitable candidate animal models to elucidate the disease mechanisms of endometriosis is still under rigorous evaluation. Although the use of small animals such as rabbit, rat, and mouse were a few of the animal models that are intensively studied, several disadvantages such as lesions sometimes being too unclear to be distinguished from surrounding normal tissues is still an issue to be addressed. This indistinctness makes it difficult to determine the weight of the lesions, which is essential in endometriosis studies. To address this issue, a transgenic mouse was developed. The use of green fluorescent protein (GFP) imaging of tumor growth, metastasis, and angiogenesis in mouse models have provided the new insights into the real-time growth and metastatic behavior of cancer. This study inspired the use of GFP imaging in the evaluation of endometriosis progression in mice. Moreover, the sensitivity of the method eliminates the surrounding normal tissues to metastatic tissue, thus making this transgenic mouse models can be a good candidate for the evaluation of endometriosis. In addition, this effort utilized the technology to quantitatively assess the human endometriotic tissue maintenance and regression in a non-invasive mouse model of endometriosis (54). Notably, they were able to demonstrate the feasibility of visualizing fluorescent endometriosis-like lesions in nude mice in a non-invasive manner.
With these scientific efforts, Hirata et al. developed the transgenic mice that ubiquitously express GFP for the evaluation of endometriosis (19). In their study, they provided several advantages of using this animal models; first, its sensitivity. Small endometriotic lesions are often more difficult to be identified by macroscopic examination. However, with the use of GFP-emitted light in the transgenic mice which can be easily detected in dark field, the location of the lesions can easily be discovered. In addition, the level of fluorescence of endometriotic lesions can be used for semi-quantitation of their size. The fluorescence intensity is positively correlated with weight of the lesions. And lastly, this animal models can clearly identify endometriotic cells originating from donor mice in histologic sections. The precise discrimination of endometriotic cells from other cells may have significant implications for the analysis of endometriotic tissues in animal models. However, a great disadvantage of using this model in drug development evaluation is that the GFP interactions to drugs might interfere with the study. Thus, additional evaluation should be conducted to provide a reliable animal model for drug assessment in endometriosis. Nevertheless, this transgenic mouse models can be utilized for the observation of endometriosis development and progression of disease.
Local inflammatory reaction in the peritoneal environment is also considered one of the contributing factors in the pathogenesis of endometriosis (44). Studies have demonstrated the effects of inflammatory mediators, such as LPS, or combined effects of E2 and LPS on promoting proinflammatory response in pelvis and growth of endometriosis (45,46). As mice with natural estrous cycle were used to be the innate hormonal environment in juvenile mice, in one study, we also analyzed the influence of LPS-induced inflammation using both recipient and donor mice with natural estrous cycle to avoid excess estrogen environment (47). For four weeks, recipient mice were treated with a single intraperitoneal (i.p) injection of LPS (2mg/kg) or vehicle (saline) twice weekly. We demonstrated that administrating LPS for four weeks significantly increased the total number (LPS: 2.8±0.27 vs. vehicle: 1.6±0.22/mouse), the average weight (65.0±9.7 vs. 30.0±5.2 mg/mouse), and the surface area (11.7±2.3 vs. 4.6±0.6 mm2/mouse) of endometriosis-like lesions compared with the vehicle group. LPS enhanced mRNA expression of Ptgs-2, Vegf, Ccl-2, and Il-6 in endometriosis-like lesions. LPS also increased the percentage of Ki67-positive cells and enhanced the intensity and rate of positive cells of CD3, F4/80, and PECAM. Intense expression of phosphor-p65 NF- κB (nuclear factor -kappa B) after LPS administration was observed.
Primate models would be the best fit for experimentation, however, the cost and lack of housing facilities provide great disadvantage to the progress of study of endometriosis. As an alternative model system, mice are readily available and comparatively inexpensive, but the relatedness to human endometriosis is an issue to consider for using them as a good animal models for endometriosis. With this, Pelch et al. investigated the gene expression profile of mouse models compared to women (32). Remarkably, the result showed that mouse endometriotic lesions resemble lesions in women. Moreover, the work has shown that mouse and human endometriotic tissue progresses from a move to a less biologically active form. In addition, the genes thought to be particularly important to endometriosis were altered in the same directions in both the human study and the use of mouse model study. With the remarkable findings of the study that the gene expression profile in the mouse endometriotic lesion is analogous to women, this makes mouse models a good candidate tool for studying endometriosis pathogenesis, pathophysiology, and treatment.
In addition to typical estrogen suppressing agents, combined targeting of E2 and LPS could be useful in the treatment of endometriosis. SR is a SERM, which is reported to have estrogen receptor (ER) α antagonistic activity with a weak partial agonist activity to ERβ (47). SERMs have been considered for using in the treatment of endometriosis due to their antiproliferative effects on the endometrium. Compared to other SERMs, which have ERα partial agonistic activity, SR is predicted to yield a superior effect in humans due to its pure antagonistic action on ERα. We investigated the effects of SR on the development of murine endometriosis-like lesions (48) by establishing the endometriosis models as previously described (30). Considering the synergistic effect between E2 and LPS, we also used the LPS to promote the pelvic inflammatory process. After induction, recipient mice were treated with vehicle or LPS (0.05 mg/kg, i.p; twice a week) without SR or with SR (1mg/kg; subcutaneously, daily). We demonstrated that after 4 weeks of treatment, SR effectively suppressed the growth and inflammatory-associated genes expression in endometriosis-like lesions without inducing endometrial growth (48).
We undertook a study of fever as a potential therapy for endometriosis that illustrated the effectiveness of the murine models. The medical herb feverfew has long been used as a folk medicine for treating fevers, migraine, rheumatoid arthritis, and dysmenorrhea. Parthenolide is considered the primary bioactive compound in feverfew having anti-tumor and anti-inflammatory properties (49). Parthenolide produces anti-tumorigenesis effects against human acute myeloid leukemia and solid tumors, such as breast and pancreatic cancer (43,50,51). We, therefore, used an experimental mouse models to evaluate the effect of parthenolide as a therapy for endometriosis (30).
We established the models by injecting tissue suspension intraperitoneally. Endometriosis-like lesions had grown in the abdominal cavity of all mice. The deposits appeared as cystic lesions bulging under the serosal coat. Most of the lesions were observed around the abdominal incision, the intestinal membrane, and the renal capsule. The size of lesions ranged from approximately 2 to 8 mm in diameter. Histologic sections stained with hematoxylin and eosin were endometriotic in character. The monolayer epithelial cell lining of the cyst was revealed by hematoxylin and eosin staining.30 We confirmed that cytokeratin (a marker of epithelial cells) and vimentin (a marker of stromal cells) in the mouse endometriosis-like lesions were positive, whereas calretinin (that of mesothelium cells) was negative, indicating that these cystic lesions originated from the injected endometrial tissues, not the peritoneal cells.
In view of the above considerations, we proposed a hypothesis that parthenolide may have an inhibitory effect on the development of endometriosis. After parthenolide treatment for 4 weeks, the total number of lesions (5.8 vs. 3.9/mouse) was significantly reduced, and the average weight (65.6 vs. 29.6 mg/mouse) and the surface area (50.3 vs. 25.7 mm2/mouse) of lesions decreased by approximately 50% of controls. To evaluate the proliferative activity of lesions, the ratio of Ki67-stained cells was calculated. In endometrial glands epithelia, the percentage of Ki67-positive cells decreased after parthenolide treatment (17.8 vs. 8.5%).
IAPs are a family of proteins implicated in multiple ways in cell death regulation, ranging from the inhibition of apoptosis to the regulation of cell cycle and inflammation. In the signaling pathway of apoptosis, IAPs have emerged as modulators in a step that involves direct inhibition of the terminal effectors, caspase-3, and -7. We previously reported higher expression of four IAPs (cIAP-1, cIAP-2, XIAP and survivin) in endometriotic tissue and endometriotic stromal cells (ESCs) than in eutopic endometrial tissues and eutopic endometrial stromal cells and demonstrated that IAP antagonist (BV6) repressed ESCs proliferation through NF-κB pathway (52).
To investigate the role of IAPs in the development of murine endometriosis lesions, we established the homologous ovariectomized murine model by transplanting uterine tissue as previously described (30). Recipient mice were treated with a single intraperitoneal injection of BV6 (10 mg/kg) twice weekly. After the treatment with BV6 for 4 weeks, the total number (Veh: 5.0 ± 1.6 vs BV6: 2.5 ± 0.6: P < .01), the average weight (Veh: 73.3 ± 27.8 mg vs BV6: 25.5 ± 10.7 mg: P < .01), and the surface area (Veh: 43.3 ± 17.1 mm2 vs BV6: 20.4 ± 6.2 mm2: P < .01) of lesions per mouse were significantly less than in the vehicle group. BV6 also decreased the Vegf, Il-6, Ccl-2, and Lif mRNA expression in the murine endometriosis-like lesions compared with the vehicle group (P < .05). BV6 also repressed the inflammatory and angiogenic activity and phospho-p65 NF-κB expression in endometriosis-like lesions (53).
Convenient and reliable endometriosis animal models are needed to accelerate emerging therapeutic alternatives. According to the Sampson’s “implantation theory”, we established the syngeneic immunocompetent mice model. In this model, we and the other investigators provided crucial evidence of both the development of endometriosis-like lesion growth and donor tissue responsiveness. Recently, Pelch et al described a detailed method of a surgically-induced endometriosis mouse model by auto-transplantation of its uterine tissue (32).
The rodent endometriosis models using mice or rats are widely used in research, but they may have limitations and may not mimic all aspects of human pathophysiology. For example, in homologous rodent models, “healthy’” uterus is cut into fragments and transplanted into the peritoneum, whereas it has been suggested that the eutopic endometrium of women suffering from endometriosis may already be abnormal (2). Nude mice lacking an intact immune system are employed in the heterologous model, which cannot mimic the inflammatory response normally seen in human endometriotic lesion (36). Although heterologous rodent endometriosis models are responsive to drugs and manipulations that induce a hypoestrogenic state, such as ovariectomy, GnRH agonists, aromatase inhibitors, danazol, and selective estrogen receptor modulators, it may be difficult to analyze novel target families in which, for example, the murine ligand does not bind to the receptor of the human transplant.
Whereas cell-based in vitro experiments provide a framework for testing molecular mechanisms, eventually, confirming their role in disease causality in vivo can only be accomplished by a suitable animal model. For a disease as diverse as endometriosis, a single animal model would unlikely be sufficient to represent the entire diversity in etiology, pathogenesis, and pathology. Each model has design-related strengths and limitations. For example, a recently described disease model consists of introducing endometrial tissue via direct injection into the peritoneal cavity of immunocompetent mice without suturing (28,32,40). In mice, the injected tissue forms cyst-like endometriosis lesions; however, the injection method does not seem to work in rats because the tissue fails to attach to and invade the peritoneal cavity (64). Since all endometriosis lesions in each model attached to the peritoneum and/or mesothelium with or without suturing, evaluating attachment or invasion of lesions would be difficult. Furthermore, in regard to quantifying the injected endometrial fragments, this model may be insufficient to evaluate the endometriotic lesions precisely.
The rodent model is used extensively to study the etiology, pathology, and risk factors of endometriosis (9,19,35,40,65-70) as well as to explore novel therapeutics (30,31,71-77). In conclusion, this model provides an important tool to evaluate a therapeutic approach to the disease. It will help to better understand disease evolution in the living animal and permit faster and more accurate characterization of a drug’s effect on experimental endometriosis.
To explain the pathophysiology of endometriosis and observe the endometriosis dispersion as early as possible without sacrificing the mice, we generated endometriosis mice model using transgenic eluc mice (with specific endometriosis genetic expression such as VEGF). Using in vivo imaging system (IVIS), the endometriosis disease progression, cell trafficking, and gene expression patterns in living animals can be easily observed. The differences between GFP mice and eluc mice is the endometriotic like lesion can not be detected in a deep intra peritoneum in GFP mice. (19) First, it needs a confirmation whether this animal model would become a reliable endometriosis model and whether the efficacy of drugs would be possible to be observed using this model. Our future plan and hope are to develop instruments to detect endometriotic lesions without laparoscopic intervention.
The authors have no conflicts to disclose relative to this work.
LPS
lipopolysaccharide
SR-16234
inhibitor of apoptosis proteins
phosphate buffered saline
estradiol valerate
intraperitoneal
nuclear factor
selective estrogen receptor modulator
In vivo Imaging System
Prostaglandin-endoperoxide synthase-2
Platelet Endothelial cell adhesion molecule
Vascular Endothelial Growth Factor
Reverse Transcription Polymerase Chain Reaction
Chemokine (C-C motif) ligand 2
interleukin 6
messenger Ribo Nucleic Acid
Protein that in human in encoded by the Mk167 gene
an IAP antagonist
Green Fluorescent Protein
Enhanced green-emitting Luciferase
Gonadotrophin Releasing Hormone
Estrogen Receptor-Beta
Estrogen Receptor- alpha
ERβ over-expressed