




 , Qingyang Wu 2,†, Hao Qian 1, Liang Wang 1, Guangcheng Liu 1, Bin Zhang 1, Wei Wu 1,*
, Qingyang Wu 2,†, Hao Qian 1, Liang Wang 1, Guangcheng Liu 1, Bin Zhang 1, Wei Wu 1,* , Shuyang Zhang 3,*
, Shuyang Zhang 3,*1 Department of Cardiology Medicine, Peking Union Medical College Hospital, Chinese Academy of Medical Science and Peking Union Medical College, 100730 Beijing, China
2 Eight-Year Medical Doctor Program, Chinese Academy of Medical Sciences and Peking Union Medical College, 100730 Beijing, China
3 Department of Cardiology, State Key Laboratory of Complex Severe and Rare Diseases, Peking Union Medical College Hospital, Chinese Academy of Medical Sciences & Peking Union Medical College, 100730 Beijing, China
†These authors contributed equally.
Abstract
About 20% of patients with coronary artery disease (CAD) experience adverse events within five years of undergoing percutaneous coronary intervention (PCI) for acute myocardial infarction. In these patients, the impact of metformin on long-term prognosis remains uncertain.
This study enrolled 22 metformin (Met)-CAD patients with diabetes mellitus (DM) who had been administered metformin for at least six months before PCI, 14 non-Met CAD-DM patients with DM who had never taken metformin or had stopped taking metformin for a year before PCI, and 22 matched healthy controls. A 5-year follow-up was conducted to collect clinical prognosis data. Fecal 16S rRNA sequencing and serum untargeted metabolomics analyses were performed. BugBase was utilized to analyze the possible functional changes in the gut microbiome. Multi-omics analysis was conducted using Spearman’s correlation to explore the interactions between metformin, gut microbiome, serum metabolites, and clinical prognosis.
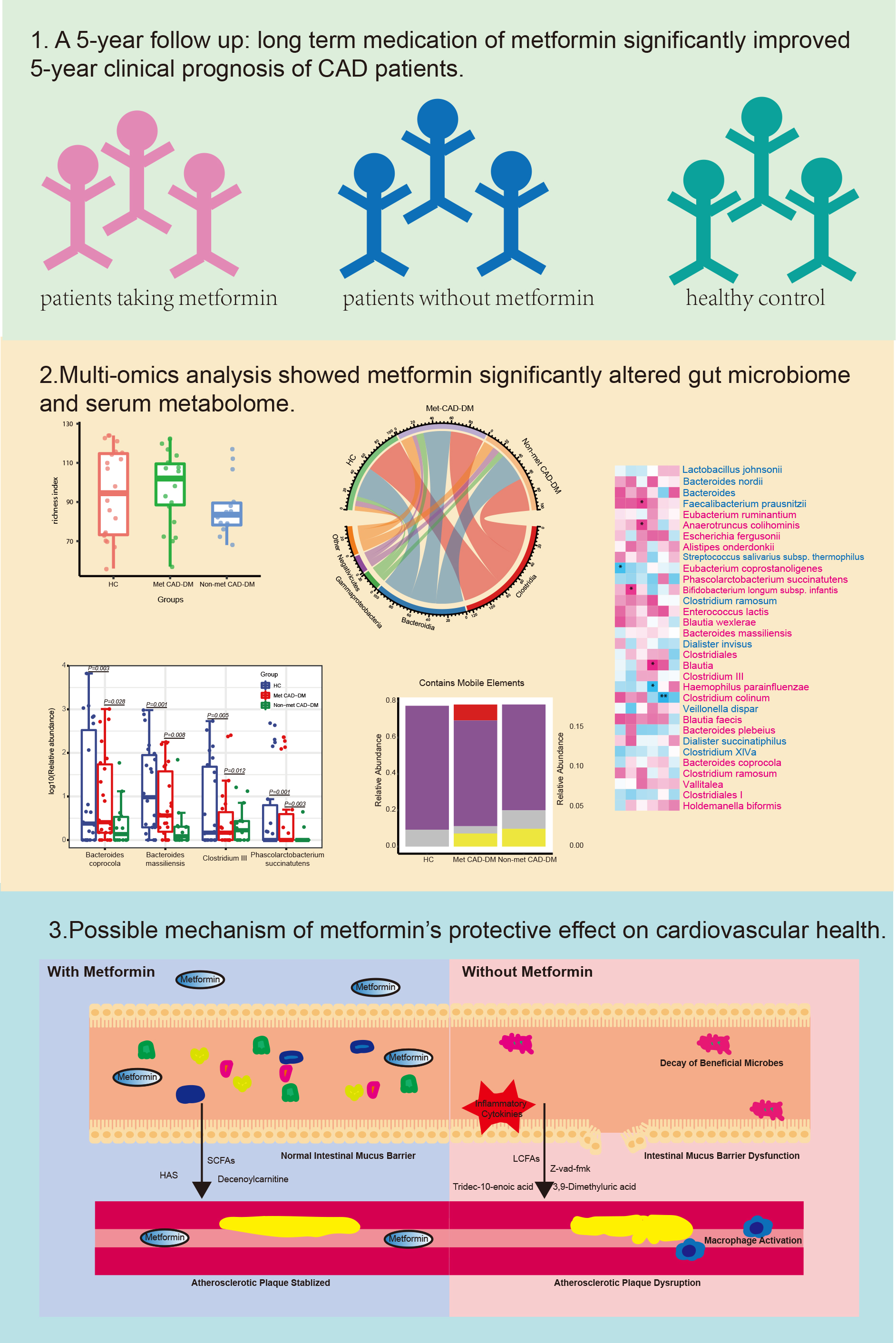
Metformin significantly lowered the 5-year major adverse cardiac events (MACEs) in Met CAD-DM patients. We found a higher abundance of Bacteroides coprocola, Bacteroides massiliensis, Phascolarctobacterium succinatutens, and Eubacterium coprostanoligenes in the Met CAD-DM patients, as well as an increase in hydroxy-alpha-sanshool (HAS) and decenoylcarnitine and a decrease in tridec-10-enoic acid, Z-vad-fmk (benzyloxycarbonyl–Val–Ala–Asp (OMe)–fluoromethylketone), 3,9-dimethyluric acid in blood serum. Multi-omics analysis revealed that alterations in the gut microbiome and serum metabolites are significantly associated with the 5-year prognosis of CAD-DM.
Metformin significantly improved the 5-year prognosis of CAD patients following PCI. Metformin tended to have more positive effects on the commensal flora and metabolic profiles, which may explain its beneficial effects on cardiovascular health. This study revealed the potential associations between metformin and the gut microbiome, an associated alteration in serum metabolome, and the impact on the host immune system and metabolic pathways.
Graphical Abstract
Keywords
- metformin
- gut microbiota
- coronary artery disease
- diabetes mellitus
- multiomic analyses
Coronary artery disease (CAD) is a major global health concern and a leading
cause of mortality worldwide [1]. Among CAD patients who underwent percutaneous
coronary intervention (PCI), approximately 20% experienced adverse events within
five years, with an all-cause mortality rate ranging from 2% to 11% following
stent implantation [2]. Chronic inflammation is a key driver of atherosclerosis
progression and plaque destabilization in diabetes mellitus (DM). This
inflammatory milieu promotes adverse remodeling of coronary arteries, thereby
increasing the risk of adverse events in CAD patients. Chronic low-grade
inflammation plays a pivotal role in the pathogenesis of coronary artery disease
[3]. The activation of the inflammatory response significantly contributes to
plaque destabilization and subsequent development of acute coronary syndromes
[4]. Several signaling pathways associated with the inflammatory response,
including the nucleotide-binding oligomerization domain (NOD)-, leucine-rich repeat
(LRR)- and pyrin domain-containing protein 3 (NLRP3)
inflammasome, toll-like receptors, and Notch and Wnt signaling pathways, have
been implicated in the development and subsequent regression of atherosclerosis
[5]. The polarization of M1 macrophages is considered to be involved in this
process. M1 macrophages are primarily activated by lipopolysaccharide (LPS) and
In recent years, the gut microbiota has received significant attention due to its potential involvement in various disease mechanisms [7, 8]. There is growing evidence to suggest that changes in the structure and function of the gut microbiota contribute to the onset and progression of CAD [9] and DM [10]. The gut microbiota can break down dietary components and generate metabolites that affect the host’s metabolism and immune responses [11]. These metabolites can influence various physiological functions and contribute to disease progression. An imbalance in the gut microbiota can increase inflammation, triggering immune activation and chronic inflammation [12]. This prolonged inflammatory state plays a crucial role in the development of numerous diseases [13]. In addition, disruptions in the gut microbiota can compromise the integrity of the gut barrier, leading to increased permeability and translocation of microbial byproducts. This process activates immune responses and may contribute to the emergence of systemic diseases. Therefore, exploring the connection between the gut microbiota and the progression of CAD and DM offers a novel perspective on understanding these diseases.
Metformin is a widely prescribed oral antidiabetic medication. In recent years, metformin was found to be beneficial for cardiovascular health. Metformin has been found to have beneficial effects on body mass index (BMI) and blood pressure, thereby helping to promote cardiovascular health [14]. Metformin also plays a crucial role in modulating the gut microbiota [15]. Metformin also affects microbial metabolism and virulence factors, which are essential for maintaining gut homeostasis. It has been suggested that metformin could influence the gut microbiota through various mechanisms. Metformin results in alterations to the structure of the gut microbiota by metabolic changes which may enhance the presence of beneficial bacteria, such as lactobacilli, and decreasing harmful bacteria [16, 17, 18]. Alternatively, metformin can affect microbial metabolism, stimulating the generation of metabolites such as short-chain fatty acids (SCFAs) which are crucial for maintenance of gut health [19]. Furthermore, a study showed that metformin may play a role in reducing the virulence of specific pathogenic bacteria, thereby decreasing inflammation [20]. However, research has primarily focused on the short-term effects of metformin on the gut microbiome. Metformin has been widely used as a first-line drug in diabetes and is often used long-term. Long-term use of metformin has a significant effect on the health of the host and their gut microbiome. However, the long-term effects of metformin have not been thoroughly investigated. Therefore, examining the mechanisms underlying CAD complicated by DM, along with alterations in gut microbiota linked to long-term metformin use, is of immense value.
The participants in the study were consecutively recruited at the Department of
Cardiology in the Peking Union Medical College Hospital from 2016 to 2018. The
inclusion criteria required patients to have
Data on metformin intake was collected. Typically, the gut microbiota undergoes significant changes within three months following the onset of the disease or alternations in medication, and these changes are maintained thereafter [22, 23]. Therefore, based on metformin usage for at least six months, the 36 CAD patients with DM were divided into two groups: (1) Metformin (Met) CAD-DM group (N = 22): CAD-DM patients who had taken metformin for at least six months before PCI. (2) Non-Met CAD-DM group (N = 14): CAD-DM patients who had never taken metformin or had stopped taking metformin for at least one year before PCI.
In addition, 22 healthy volunteers who met the following criteria were enrolled as the healthy control (HC): (1) did not take metformin, (2) did not suffer from CAD or DM, and (3) did not meet any of the above exclusion criteria.
All patients underwent 5-year follow-up. All the enrolled patients developed no additional comorbidities or changes in medication throughout the 5-year follow up period.
Written informed consent was obtained from all participants and the study adhered to the principles of the Declaration of Helsinki. The ethical approval was obtained from the Peking Union Medical College Hospital, and the protocol number is I-24PJ0927.
The samples were prepared and stored according to the protocol described in our previous study [21]. Microbial DNA was extracted from the fecal samples using the bead-beating method [24]. PCR was conducted to amplify the V3–V4 region of 16S rRNA genes [25]. The sequencing library was established as described previously [26], and purified products were sequenced using the Illumina Miseq system (Illumina Inc., San Diego, CA, USA). The downstream analysis of amplicons was performed using EasyAmplicon v1.0 (Fred Hutchinson Cancer Research Center, Seattle, WA, USA) [27]. For dereplication, the derep_fullength command in VSEARCH v2.15 (Technical University of Denmark (DTU), University of Copenhagen, Copenhagen University Hospital, Copenhagen, Denmark) was employed [28]. Operational taxonomic units (OTUs) were grouped using the -cluster_otus command in USEARCH (v10.0, Illumina, Inc, San Diego, CA, USA) with a 97% cutoff [29]. A feature table was generated using vsearch–usearch_global, and taxonomic classification was performed based on the Greengenes database using usearch–otutab [30].
All sample sequences were normalized to match the sample with the fewest
(10,560) sequences for diversity index calculations. An observed
species richness index was used to assess alpha diversity. Beta diversity was
examined through principal coordinate analysis (PCoA) and constrained PCoA
(CPCoA) using Bray-Curtis distances. Group compositions were visualized at the
phylum level as boxplots and at the genus level as a Chord diagram using the R
package ggplot2 (https://ggplot2.tidyverse.org/). To compare differences,
edgeR (https://bioconductor.org/packages/release/bioc/html/edgeR.html) was utilized to detect group
variances with the application of negative binomial distribution, and the
Benjamini-Hochberg method controlled the false discovery rate (FDR) [31]. A
significance level of p
A Wilcoxon rank-sum test was used for metabolomics analysis. Differential
metabolites were identified based on a Variable Importance in the Projection
(VIP) greater than 1 and a significance level of p
Serum metabolome analysis was performed using a Waters ACQUITY
ultra-high-performance liquid chromatography system (Milford, MA, USA) in
conjunction with a Waters Q-TOF Micromass system (Manchester, UK) operating in
both positive and negative ionization modes. Different modes, such as polar ionic
and lipid modes, were used based on the metabolite characteristics. The sample
preparation and experimental procedures for liquid chromatograph-mass
spectrometer (LC-MS) were previously outlined and explained. A peak-ion intensity
matrix was refined by eliminating peaks that displayed zero values in more than
80% of samples. A quality control sample coefficient of variation threshold of
30% was implemented. To identify metabolites showing significant differences
between groups, a Wilcoxon rank-sum test was used. Subsequently, partial least
squares discriminant analysis (PLS-DA) was conducted via SIMCA software (MKS Umetrics, Uppsala, Sweden). Peaks
were considered important based on a VIP-value greater than 1 and a significance
level of p
One-way ANOVA was used for analyzing continuous data that followed a normal
distribution across the three groups. Non-normally distributed continuous data
among three groups were analyzed using the Kruskal-Wallis H-test, while
comparisons between two groups were conducted using the Mann-Whitney U test.
Categorical variables were assessed through either the
The 36 CAD patients with diabetes were divided into two groups: those who took metformin (Met CAD-DM group, N = 22) and those who did not take metformin (non-Met CAD-DM group, N = 14). Both stool and blood samples were collected in 2019. At the time of sample collection, patients in the Met CAD-DM group had been taking metformin for at least one year and continued taking it for the following five years. In contrast, patients in the non-Met CAD-DM group did not take metformin for one year before fecal collection and did not take metformin for the subsequent five years. The clinical characteristics of the study cohort are presented in Table 1. Baseline clinical characteristics, including age, BMI, blood pressure, comorbidities (HTN (hypertension), HLP (hyperlipidemic pancreatitis), FLD (fatty liver disease)), CAD severity (Gensini score, New York Heart Association (NYHA) score), and cardiac biomarkers (CK-MB (creatine kinase MB isoenzyme), hsCRP (high sensitivity C-reactive protein)), were comparable between the two groups.
| HC | Met CAD-DM | Non-Met CAD-DM | p-value | ||
| (N = 22) | (N = 22) | (N = 14) | |||
| Demographics | |||||
| Age* | 55.86 |
64.82 |
64.29 |
0.003ab | |
| Gender (Male)§ | 11 (50.0) | 15 (68.2) | 9 (64.3) | 0.441 | |
| SBP, mmHg* | 117.86 |
131.36 |
127.43 |
0.003a | |
| DBP, mmHg† | 76.00 (20.50) | 73.00 (16.75) | 75.00 (13.25) | 0.912 | |
| Height, cm* | 165.27 |
164.73 |
166.29 |
0.870 | |
| Weight, kg* | 66.03 |
75.16 |
72.71 |
0.014a | |
| BMI, kg/m2* | 24.14 |
27.39 |
25.97 |
0.003a | |
| Waist, cm* | 82.14 |
95.64 |
93.29 |
||
| Gensini Score* | NA | 41.48 (24.59) | 43.85 (37.85) | 0.823 | |
| Smoke§ | 4 (18.2) | 12 (54.5) | 5 (35.7) | 0.043b | |
| Drink§ | 1 (4.5) | 13 (59.1) | 4 (28.6) | ||
| Family history§ | 7 (31.8) | 10 (45.5) | 6 (42.9) | 0.627 | |
| NYHA | 0.397 | ||||
| I§ | NA | 10 (45.5) | 9 (64.3) | ||
| II§ | NA | 11 (50.0) | 4 (28.6) | ||
| III§ | NA | 1 (4.5) | 1 (7.1) | ||
| IV§ | NA | 0 (0.0) | 0 (0.0) | ||
| Past/Personal history | |||||
| OMI§ | 0 (0.0) | 1 (4.5) | 2 (14.3) | 0.241 | |
| PAS§ | 2 (9.1) | 6 (27.3) | 4 (28.6) | 0.250 | |
| TGD§ | 2 (9.1) | 3 (13.6) | 3 (21.4) | 0.562 | |
| FLD§ | 6 (27.3) | 7 (31.8) | 4 (28.6) | 0.945 | |
| HLP§ | 7 (31.8) | 19 (86.4) | 10 (71.4) | ||
| HTN§ | 6 (27.3) | 15 (68.2) | 9 (64.3) | 0.014ab | |
| Laboratory data | |||||
| cTnI† | NA | 0.01 (0.05) | 0.01 (0.26) | 0.510 | |
| CK, U/L† | 111.00 (40.75) | 91.50 (48.00) | 108.50 (60.75) | 0.046a | |
| CK-MB, U/L† | 0.90 (0.63) | 0.70 (0.53) | 0.70 (0.70) | 0.810 | |
| TC, mmol/L* | 4.79 |
4.37 |
4.04 |
0.218 | |
| TG, mmol/L† | 1.37 (1.36) | 1.63 (1.49) | 1.16 (1.33) | 0.389 | |
| LDL-C, mmol/L† | 2.76 (0.81) | 2.19 (1.29) | 1.73 (1.23) | 0.034b | |
| HDL-C, mmol/L† | 1.20 (0.40) | 0.91 (0.23) | 0.97 (0.26) | 0.002ab | |
| hsCRP, mg/L† | 0.70 (0.77) | 2.78 (3.03) | 2.09 (2.41) | ||
| ALT, U/L† | 18.00 (13.25) | 25.00 (16.25) | 22.00 (21.25) | 0.140 | |
| AST, U/L† | NA | 23.00 (5.50) | 23.00 (23.00) | 0.693 | |
| GGT, U/L† | NA | 32.00 (19.25) | 22.00 (16.50) | 0.151 | |
| ALP, U/L* | NA | 75.36 |
72.62 |
0.565 | |
| LDH, U/L† | NA | 162.50 (29.50) | 193.00 (50.50) | 0.006 | |
| TBil, µmol/L* | 14.68 |
11.35 |
10.36 |
0.009ab | |
| DBil, µmol/L* | 4.79 |
3.22 |
2.94 |
||
| Cr, µmol/L† | 71.00 (20.50) | 69.50 (24.25) | 76.50 (23.25) | 0.735 | |
| Urea, mmol/L* | 4.82 |
6.49 |
5.44 |
0.001a | |
| HGB, g/L* | 145.23 |
136.41 |
136.36 |
0.084 | |
| WBC, |
5.37 |
7.20 |
5.83 |
||
| RBC, |
4.60 (0.47) | 4.46 (0.39) | 4.47 (0.49) | 0.248 | |
| HCT, %* | 41.97 |
39.71 |
40.00 |
0.124 | |
| Glucose, mmol/L† | 5.90 (1.95) | 9.20 (5.38) | 8.10 (3.43) | ||
| IL-18, U/L (pg/mL)* | 861.16 |
678.34 |
717.90 |
0.236 | |
| IL-1 |
3.26 (0.65) | 3.00 (1.15) | 3.04 (3.22) | 0.886 | |
| IL-6† | 2.81 (1.88) | 4.15 (9.64) | 3.48 (10.61) | 0.044b | |
| TNF- |
3.87 (9.77) | 19.88 (30.39) | 27.61 (41.55) | ||
*, mean
ap
HC, healthy control; Met, metformin; CAD, cardiovascular disease; DM, diabetes mellitus; SBP, systolic blood pressure; BMI, body mass index; OMI, old myocardial infarction; PAS, peripheral atherosclerosis; TGD, thyroid gland dysfunction; FLD, fatty liver disease; HLP, hyperlipidemia; HTN, hypertension; cTnI, cardiac troponin I; CK, creatine kinase; CK-MB, creatine kinase MB isoenzyme; TC, total cholesterol; TG, total triglyceride; LDL-C, low density lipoprotein-cholesterol; HDL-C, high density lipoprotein-cholesterol; hsCRP, high sensitivity C-reactive protein; ALT, alanine transaminase; AST, aspartate transaminase; GGT, gamma-glutamyl transferase; ALP, alkaline phosphatase; LDH, lactate dehydrogenase; TBil, total bilirubin; DBil, direct bilirubin; Cr, creatine; HGB, hemoglobulin; WBC, white blood cells; RBC, red blood cells; HCT, hematocrit value; IL, interleukin; TNF, tumor necrosis factor; DBP, diastolic blood pressure; IQR, interquartile range; NYHA, New York Heart Association.
After five years, in March 2024, we conducted a follow-up assessment with all the patients via telephone. The results showed that of the 22 individuals in the Met CAD-DM group, six patients had a major adverse cardiac event (MACE) event within five years, whereas out of the 14 individuals in non-Met CAD-DM group, nine had a MACE event. These results were statistically significant (p = 0.03). This indicates that patients in the Met CAD-DM group have a significantly better clinical prognosis than those in the non-Met CAD-DM group, indicating that metformin significantly enhanced the 5-year cardiovascular prognosis.
Despite similar baseline clinical characteristics, the Met CAD-DM group exhibited a significantly improved five-year clinical prognosis compared to the non-Met CAD-DM group. To investigate the potential role of the gut microbiome on this outcome, 16S rRNA sequencing was performed (Supplementary Table 1 and Supplementary Table 2). Alpha diversity analysis revealed a higher gut microbiome diversity in the healthy control group compared to both patient groups, with the lowest diversity observed in the non-Met CAD-DM group (Fig. 1A). Beta diversity analysis demonstrated microbial community structures across the three groups (Fig. 1B, Supplementary Fig. 1).
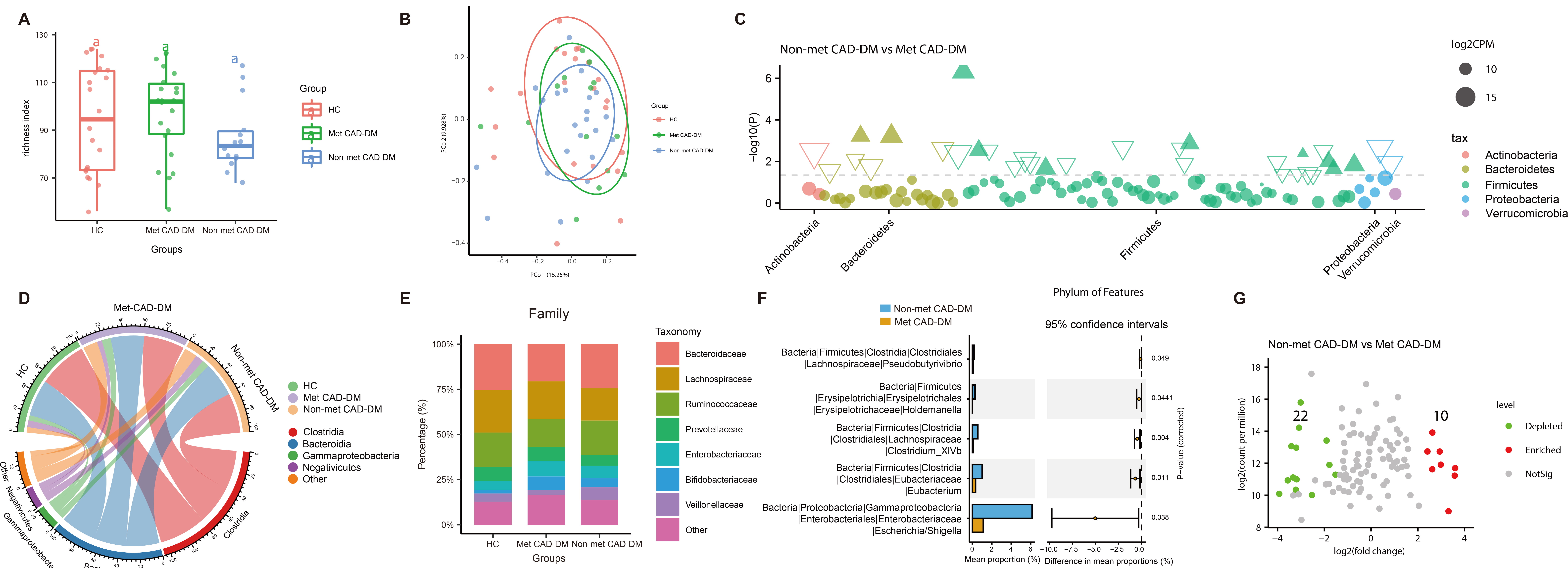 Fig. 1.
Fig. 1.
Medication of metformin affects the taxonomic features of gut
microbiota in patients with CAD-DM. (A) Observed Species Richness Index,
representing the number of operational taxonomic units (OTUs) identified in each
group. Box-plot features represent the mean
We further analyzed the specific structural composition of the three groups. The results showed that in the phylum level of gut microbiota (Fig. 1C), the non-Met CAD-DM group exhibited a greater incidence of Firmicutes compared to the Met CAD-DM group. At the class level of gut microbiota (Fig. 1D), Clostridia constituted the largest proportion across all three groups, followed by Bacteroidia. The proportion of Clostridia in the Met CAD-DM group and non-Met CAD-DM group were very similar but was much lower compared to the HC. The three least abundant classes were Gammaproteobacteria, Negativicutes, and Others. These three classes also had different proportions in the non-Met CAD-DM group, Met CAD-DM group, and HC group. At the family level of gut microbiota (Fig. 1E), Bacteroidaceae showed the highest abundance in all three groups, followed by Lachnospiraceae, Ruminococcaceae, Prevotellaceae, Enterobacteriaceae, Bifidobacteriaceae, and Veillonellaceae. We then compared the genus level of gut microbiota between the non-Met CAD-DM group and the Met CAD-DM group (Fig. 1F). The differential genera included Escherichia/Shigella, Eubacterium, Clostridium_XlVb, Holdemanella, and Pseudobutyrivibrio. To further investigate the role of metformin in shaping gut microbiome, we analyzed the differential species between the non-Met CAD-DM group and the Met CAD-DM group (Fig. 1G). The analysis identified a total of 32 distinct gut microbiome species between the two disease groups. The specific FDR-adjusted p-values are presented in Supplementary Table 3. 10 species were more enriched in the Met CAD-DM group than in the non-Met CAD-DM group, while 22 species were more enriched in the non-Met CAD-DM group than in the Met CAD-DM group. These results demonstrate that metformin significantly alters the abundance and composition of gut microbiota. The clinical data indicate no baseline differences between groups, suggesting that the observed microbial changes are likely due to metformin treatment rather than pre-existing variability.
The above results showed that the use of metformin modifies the gut microbiome. It is known that the composition and function of gut microbiome are inextricably linked. We carefully analyzed the differences in bacterial composition between the non-Met CAD-DM group and the Met CAD-DM group. We conducted a pairwise comparison of the bacterial composition between the groups. The results indicated that four species of bacteria were significantly different among all three groups. These four bacteria are Bacteroides coprocola (B. coprocola), Bacteroides massiliensis (B. massiliensis), Clostridium III, and Phascolarctobacterium succinatutens (P. succinatutens) (Fig. 2A). We found that all four bacterial species showed a decreasing trend in the following order: HC group, Met CAD-DM group, and non-Met CAD-DM group.
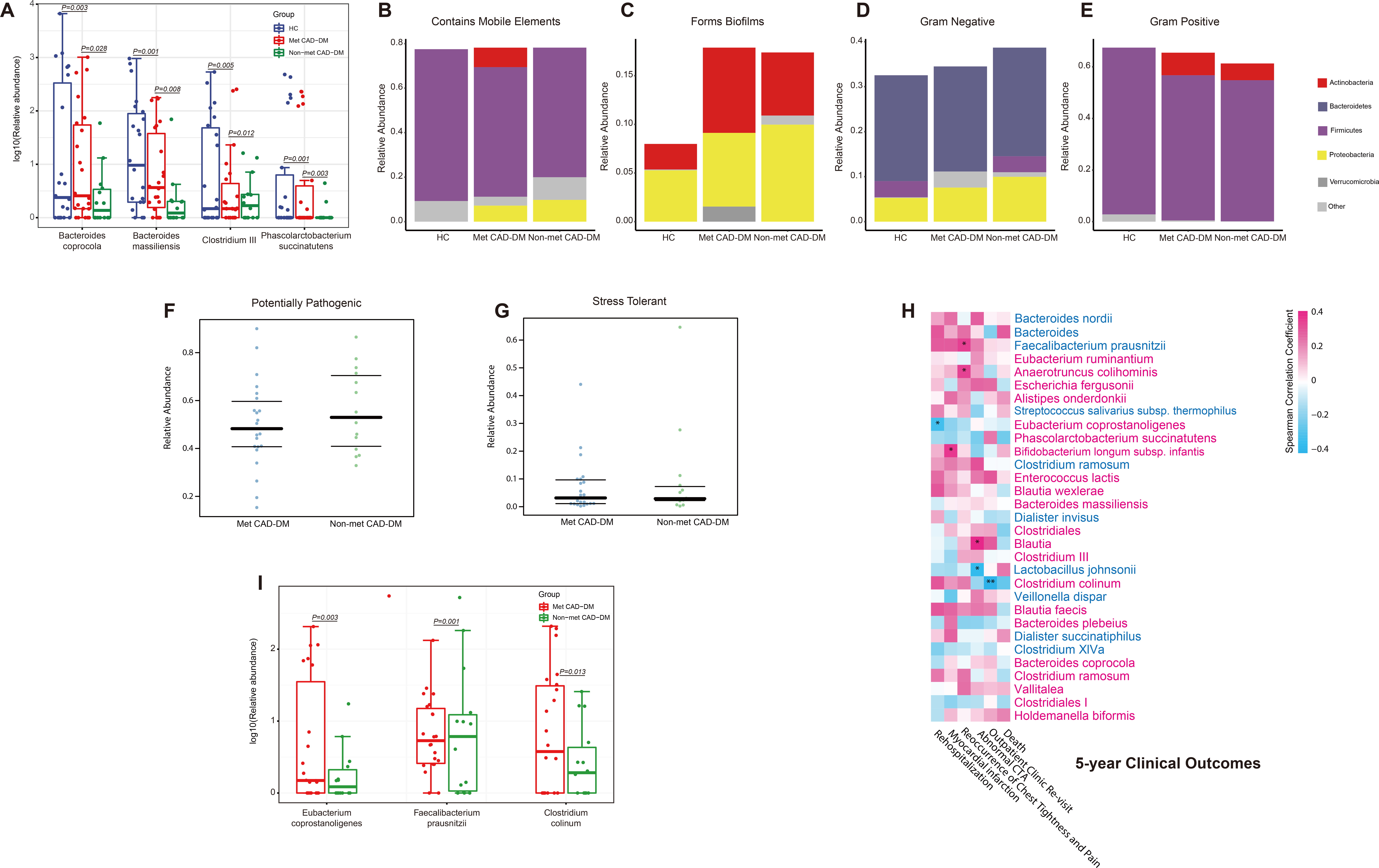 Fig. 2.
Fig. 2.
Metformin changed the abundance of specific microbes and altered
the potential function of the gut microbiome, associated with 5-year clinical
outcomes. (A) Abundance of four significantly different species among all three
groups. (B) Relative abundance of a form of the bacteria containing mobile
elements predicted based on BugBase database. (C) Relative abundance of a form of
the bacteria forming biofilms predicted based on BugBase database. (D) Relative
abundance of a form of the gram-negative bacteria predicted based on BugBase
database. (E) Relative abundance of a form of the gram-positive bacteria
predicted based on BugBase database. (F) Relative abundance of potentially
pathogenic bacteria predicted by BugBase (FDR-adjusted p = 0.43160). (G)
Relative abundance of stress-tolerant bacteria predicted by BugBase (FDR-adjusted
p = 0.81). (H) Spearman correlation between differential gut microbes
and 5-year clinical outcomes. The microbes or metabolites are highlighted in red
(enriched in Met CAD-DM) and blue (depleted in Met CAD-DM).
*FDR-adjusted p
We further predicted the function of flora using BugBase. The results showed that non-Met CAD-DM group had a higher proportion of potentially pathogenic gut microbiome than the Met CAD-DM group (Fig. 2B). This suggested that the use of metformin may reduce potentially pathogenic bacteria in the intestinal flora. We further analyzed the specific contributions of the gut microbiome to each phenotype. The majority of mobile genetic elements were found to be associated with Firmicutes, and their abundance was similar across all three groups (Fig. 2B). The main contributors to the biofilm formation are Actinobacteria and Proteobacteria (Fig. 2C). The data showed that the two diseased groups have significantly higher levels of biofilms than the HC group. The predominant gram-negative bacteria identified were Bacteroidetes and Proteobacteria (Fig. 2D) which were most abundant in the non-Met CAD-DM group. In contrast, the predominant gram-positive bacteria were Actinobacteria and Firmicutes (Fig. 2E) and were the most abundant in the HC group. The results indicated that the non-Met CAD-DM group had a higher proportion of potentially pathogenic gut microbiome than the Met CAD-DM group (Fig. 2F). This suggests that metformin use may reduce the abundance of potentially pathogenic bacteria in the intestinal flora. Furthermore, the findings revealed that the gut microbiota in the Met CAD-DM group exhibited a tendency toward increased, but not significant greater stress tolerance, than the non-Met CAD-DM group (Fig. 2G). This implies that the use of metformin has a potential role in enhancing the tendency of stress resistance of gut flora.
Correlation analysis between gut microbiome composition and 5-year clinical outcomes identified several bacterial species associated with adverse cardiovascular events (Fig. 2H, Supplementary Table 4). Bifidobacterium infantis was linked to myocardial infarction, while Faecalibacterium prausnitzii and Anaerotruncus colihominis were associated with recurrence of chest pain. Conversely, Eubacterium coprostanoligenes, and Clostridium colinum showed protective effects. In addition, we compared the relative abundance of both harmful and beneficial species. We found that metformin use was associated with a significant increase of abundance of beneficial bacteria and significant decrease of harmful species (Fig. 2I).
In this analysis, we found that significant changes were observed in the gut
flora of patients taking metformin. A significant correlation existed between gut
microflora alterations and patients’ clinical prognosis. Since alterations in gut
flora are known to significantly affect serum metabolomics [34], we further
performed an analysis of serum metabolomics. A total of 64 serum metabolites were
identified to be associated with moderate alcohol consumption with a threshold of
VIP
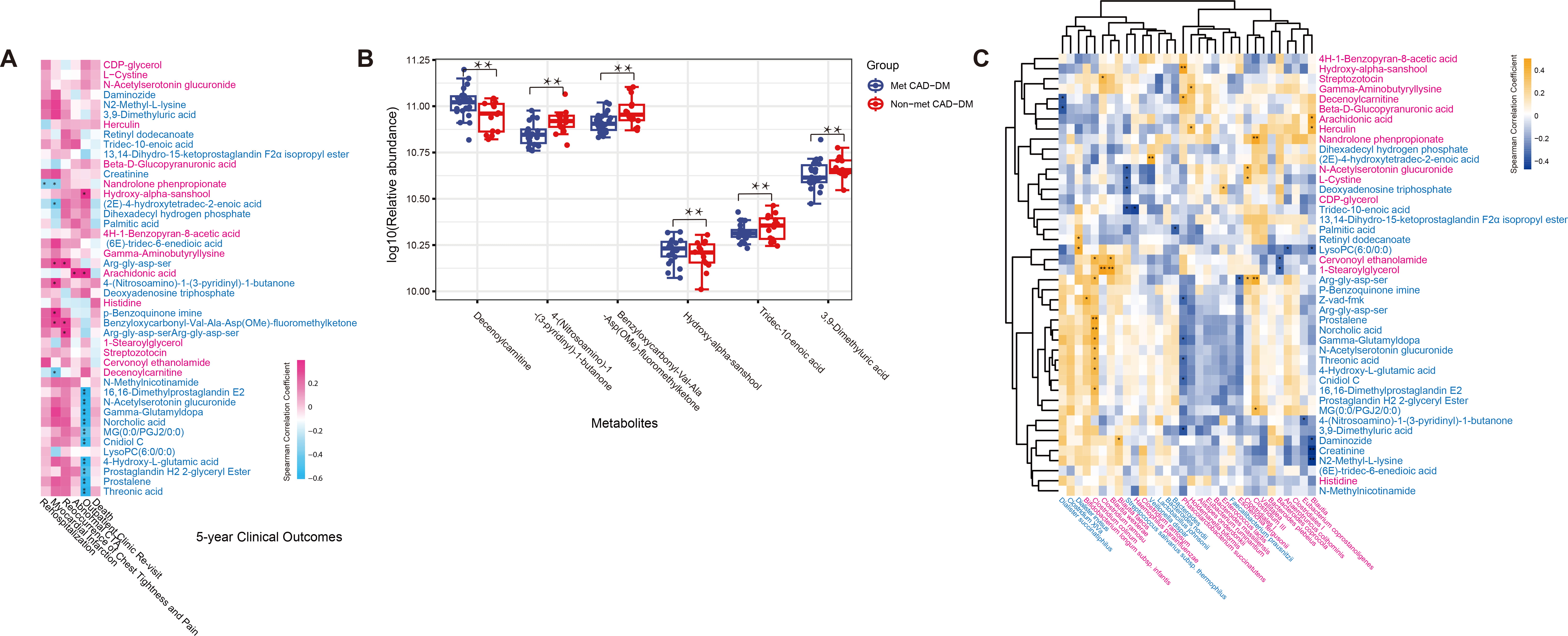 Fig. 3.
Fig. 3.
Spearman correlations between differential serum metabolites and
microbes associated with 5-year clinical outcomes. (A) Spearman correlation
between differential serum metabolites and 5-year clinical outcomes. (B) The
relative abundance of six key serum metabolites associated with patients’
clinical prognosis. (C) Spearman correlation between differential serum
metabolites and differential gut microbes. The microbes or metabolites are
highlighted in red (enriched in Met CAD-DM) and blue (depleted in Met CAD-DM).
*FDR-adjusted p
We performed a Spearman correlation analysis to study the
association between differential metabolites and differential gut microbiome
(Fig. 3C, Table 2, Supplementary Table 7). HAS is an unsaturated fatty
acid amide [35] that has been reported to attenuate neuronal oxidative stress.
HAS can also enhance the antioxidant enzyme activities and inhibit the
| Metabolite | Possible function | Abundance | Microbiota correlation |
| Hydroxy-alpha-sanshool | Attenuate neuronal oxidative stress, enhance the antioxidant enzyme activities and inhibit κB (p65 NF-κB) signaling pathway, regulate intestinal barrier dysfunction and gut microbiota dysbiosis | Enriched in Met CAD-DM | Positively associated with P. succinatutens |
| Decenoylcarnitine | Inhibit Mmp9 expression, associated with insulin resistance | Enriched in Met CAD-DM | Positively associated with P. succinatutens |
| 4-(nitrosoamino)-1-(3-pyridinyl)-1-butanone | - | Enriched in Met CAD-DM | Negatively correlated with Eubacterium Coprostanoligenes |
| Tridec-10-enoic acid | Long chain fatty acid | Depleted in Met CAD-DM | - |
| 3,9-dimethyluric acid | Have pro-inflammatory potential | Depleted in Met CAD-DM | Negatively correlated with P. succinatutens |
| Z-vad-fmk | Induce non-apoptotic cell death of macrophages and is not beneficial for atherosclerotic plaque stability | Depleted in Met CAD-DM | Negatively correlated with P. succinatutens |
P. succinatutens, Phascolarctobacterium succinatutens; Z-vad-fmk, benzyloxycarbonyl–Val–Ala–Asp(OMe)–fluoromethylketone.
In summary, within the Met CAD-DM group, the medication of metformin correlates with the growth of beneficial microbiomes and the decrease of harmful ones. Additionally, the increase in beneficial microbiomes correlates with the reduction of harmful metabolites and an increase in beneficial metabolites. These changes in the serum metabolome further correlates with better cardiovascular health. These correlations may imply a potential mechanism by which metformin improves cardiovascular health and is associated with 5-year clinical prognosis.
Coronary artery disease is closely related to chronic and systemic inflammation [42]. Metformin, a well-known drug used in treating diabetes, has been reported to benefit cardiovascular health [43, 44]. The underlying mechanisms may include enhancing insulin sensitivity and reducing cardiovascular risk factors [45], decreasing inflammation [46], and reducing oxidative stress [47]. Although much research has been performed on metformin and host health, the long-term benefits of metformin on cardiovascular health via gut microbiome have not been studied, especially the long-term benefits associating with the gut microbiome. There have been studies which showed that there was no significant difference in the prognosis between strategies of insulin sensitization and insulin provision. Frye et al. [48] compare insulin-sensitizing strategies (including metformin) with insulin-providing strategies in CAD patients and find no significant difference in outcomes, highlighting the need for individualized treatment strategies in this population. Jung et al. [49] specifically examined the effects of glucose-lowering agents on CAD outcomes and concluded that metformin significantly reduces the risk of repeat revascularization after PCI in type 2 diabetes mellitus (T2DM) patients. This supports the role of metformin in CAD management but does not address gut microbiota or metabolomics. A meta-analysis by Griffin et al. [50] evaluated the cardiovascular outcomes of metformin and concluded that while it is considered safe and potentially beneficial for CAD, evidence from long-term studies remains inconclusive due to limitations in study design and small sample sizes. Our study provides new evidence that long-term metformin use is associated with improved five-year prognosis in CAD patients who have undergone PCI. Despite similar baseline clinical characteristics, patients in the Met CAD-DM group experienced significantly fewer MACE within five years after PCI compared to those without metformin treatment. All the patients included in our study had follow-up for over 5 years. These findings underscore the potential of metformin as a therapeutic strategy for reducing cardiovascular risk in coronary artery disease.
Studies have shown that gut microbiome plays an important role in cardiovascular health and may affect clinical outcomes [51]. It has also been reported that metformin may play important role in alterations of the gut microbiome [52]. Thus, we looked further into the gut microbiome taxonomic features and serum metabolome profiles to reveal the possible associations between metformin intake and better 5-year prognosis. Our study showed that the long-term use of metformin plays an important role in altering gut microbiome taxonomic features. The alpha diversity and beta diversity both presented different taxonomic features between the non-Met CAD-DM group and the Met CAD-DM group.
The alterations of the gut microbiome associated with metformin tends to be
beneficial for the patient’s cardiovascular health. It’s worth mentioning that
the healthy control group served as a base line reference rather than a focus for
comparison. Specific beneficial gut microbiomes (such as B. coprocola,
B. massiliensis, Clostridium III, and P.
succinatutens) had the lowest abundance in the non-Met CAD-DM group and the
highest abundance in HC. The Met CAD-DM showed an intermediate abundance of the
above four beneficial microbes, which we attributed to the influence of
metformin. This suggests that metformin may slow the decrease of these specific
beneficial gut microbiota during disease progression. We specially focused on the
four gut bacteria: Bacteroides coprocola (B. coprocola),
Bacteroides massiliensis (B. massiliensis), Clostridium III,
and Phascolarctobacterium succinatutens (P. succinatutens). These
bacteria exhibited the highest abundance in the healthy controls, followed by the
Met CAD-DM group and the non-Met CAD-DM group. This showed a potential
association between metformin intake and the abundance of beneficial microbiota.
It can be inferred that metformin may be associated with the decrease of these
probiotics during disease progression. These four gut florae were reported to be
beneficial to host health in several studies. B. coprocola was reported
to be very promising as a potential preventive and therapeutic agent against
obesity [53], which is a risk factor in both CAD and DM [54, 55]. Furthermore,
B. coprocola was reported to be negatively associated with liver
fibrosis among male patients with metabolic dysfunction-associated fatty liver
disease (MAFLD) [56]. B. coprocola has been demonstrated to have
anti-oxidative properties and an improvement in intestinal barrier function [57].
Studies have found that oxidative stress is associated with the onset and
progression of coronary heart disease [58, 59] and diabetes mellitus [60, 61].
The imbalance in the generation and clearance of reactive oxygen species (ROS)
can lead to extensive and permanent damage, resulting in endothelial dysfunction,
and accelerating the occurrence and development of both CAD and DM [61]. These
studies indicate that the abundance of B. coprocola may correlate with
an anti-oxidative effect. Study on Clostridium III has shown that it is
linked to the attenuation of the NF-
The bacterial species Eubacterium Coprostanoligenes (E. coprostanoligenes) deserves special attention. Metformin is associated with the abundance of E. coprostanoligenes, which is linked to a better prognosis of CAD-DM, and metformin is significantly associated with elevated abundance of these beneficial microbes. E. coprostanoligenes was reported to be able to alleviate intestinal mucositis by enhancing the intestinal mucus barrier [72]. This enhancement of the intestinal mucus barrier is associated with activating the aryl hydrocarbon receptor/AU-rich element RNA-binding factor 1 (AhR/AUF1) pathway, consequently enhancing Muc2 mRNA stability [72]. The intestinal mucus barrier forms the first line of defense against bacterial invasion while providing nutrients to support microbial symbiosis [73]. On the one hand, the intestinal mucus barrier plays a key role in preventing preclinical diabetes from progressing into diabetes. It was reported that the integrity of the intestinal mucus barrier avoids dysregulated crosstalk between gut microbiota and immune cells, thus preventing the progression of preclinical diabetes [74]. Loss of gut barrier integrity triggers activation of islet-reactive T cells and autoimmune diabetes [75]. On the other hand, the dysfunction of the intestinal mucus barrier is observed in coronary artery disease, and restoration of the intestinal barrier is regarded as a potential therapeutic target in CAD [76]. The improvement of the intestinal barrier’s function also attenuates atherosclerosis by decreasing toxic lipid accumulation and reducing inflammatory cytokines [77]. The dysfunction of the intestinal mucus barrier is also associated with hyperlipidemia [78], which is a well-known risk factor in CAD. Thus, the enrichment of E. coprostanoligenes may significantly promote the restoration of the intestinal mucus barrier, thus alleviating autoimmune reactions and improving the prognosis of CAD-DM. This further demonstrates that metformin exerts a protective effect and improves the prognosis of patients suffering from coronary artery disease complicated with diabetes mellitus.
Not only does the gut microbiome affect the 5-year prognosis of CAD-DM patients,
but the gut microbiota-derived metabolites also play an important role in the
prognosis of the disease. HAS is positively related to P. succinatutens,
which is regarded as a beneficial microbe. Furthermore, HAS is negatively
correlated with rehospitalization and death. Thus, HAS is potentially beneficial
to CAD-DM patients and improves the 5-year prognosis of CAD-DM. It has been
reported that HAS regulates gut microbiota and metabolites by affecting lipid and
amino acid metabolism pathways [37, 79]. Lipid metabolism plays an important role
in CAD. Regulating lipid metabolism may influence the prognosis of CAD. Study has
shown that the intervention of HAS could also improve the intestinal and
metabolic functions [37]. HAS has also been reported to exert anti-diabetic
effects by increasing glycogen synthesis through regulation of
phosphoinositide-3-kinase/protein kinase B/glycogen synthase kinase-3
Our study showed that decenoylcarnitine, a subtype of acylcarnitine, is significantly negatively related to the incidence of 5-year myocardial infarction. Decenoylcarnitine is also negatively associated with rehospitalization, recurrence of chest tightness and pain, as well as an abnormal computed tomography angiography (CTA) within five years. This implies that the higher the abundance of decenoylcarnitine, the better the 5-year prognosis of CAD-DM. Acylcarnitine is essential in fatty-acid metabolism [82]. Although research has shown that acylcarnitine is associated with the development of heart failure [83] and accelerates the progress of atherosclerosis [84], decenoylcarnitine is differs from acylcarnitine and only a few studies have been done on decenoylcarnitine.
In our study, we also discovered several pathogenic metabolites, including
4-(nitrosoamino)-1-(3-pyridinyl)-1-butanone, Tridec-10-enoic acid,
3,9-dimethyluric acid, and Z-vad-fmk. These metabolites are positively associated with a
poorer 5-year prognosis. The enrichment of these four metabolites worsens the
5-year outlook of the disease. Tridec-10-enoic acid is a type of long-chain fatty
acid (LCFA). These fatty acids can be derived from food or produced by certain
types of gut microbiomes such as Fusimonas intestini, a commensal
species of the family Lachnospiraceae [85]. Thus, LCFAs are closely
associated with the gut microbiome. LCFAs are also recognized as significant
triggering factors for inflammatory disease because they regulate the
palmitoylation of signal transducer and activator of transcription 3 (STAT3)
through cluster of differentiation 36 (CD36)-mediated endocytosis [86]. Both CAD
and DM are closely associated with chronic inflammation. Consequently, LCFAs play
an important role in promoting inflammation in CAD and DM, thereby worsening the
5-year prognosis. Furthermore, LCFAs also contribute to M1 macrophage
polarization by activating NLRP3 inflammasome and the NF-
We focused on two specific metabolites: 3,9-dimethyluric acid and Z-vad-fmk.
Both these metabolites exhibited a significant negative correlation with
P. succinatutens, which is a well-known beneficial microbe.
3,9-dimethyluric acid is a type of dimethyluric acid that has been linked to
lipid metabolism and is known to have pro-inflammatory properties [40]. Increased
exposure to inflammatory cytokines can activate the
ROS-p38-p65 signaling pathway. This activation leads to endothelial cell
dysfunction, resulting in coronary atherosclerotic lesions and plaque rupture
[90]. Consequently, higher levels of dimethyluric acid contribute to systemic
inflammation, worsening the prognosis of patients with CAD-DM. In contrast,
Z-vad-fmk, has been shown to induce autophagy and necrotic cell death in smooth
muscle cells via macrophage activity. Research indicates that Z-vad-fmk-treated
macrophages overexpress and secrete various chemokines and cytokines, including
TNF-
Our results collectively demonstrate that metformin improves the 5-year prognosis of CAD-DM by influencing the gut microbiome. The potential mechanisms outlined in our analysis (Fig. 4) are as follows: (1) Metformin may play important role in influencing the abundance of beneficial microbes such as B. coprocola, B. massiliensis, Clostridium III, and P. succinatutens. (2) Metformin promotes the enrichment of SCFA-producing microbes while depleting pathogenic microbes. (3) Changes in the gut microbiome not only directly affect the 5-year clinical prognosis of CAD-DM, but also impact these outcomes by regulating the serum metabolome. (4) After metformin use, the serum metabolome shows higher levels of anti-oxidant metabolites and lower levels of pro-inflammatory metabolites. These, alterations in the gut microbiome and serum metabolome are significantly associated with lower levels of systemic inflammation, diminished macrophage activation, reduced apoptosis of smooth muscle cells, and increased atherosclerotic plaque stability. These cascading effects linked to metformin contribute to a better 5-year prognosis in patients with CAD-DM.
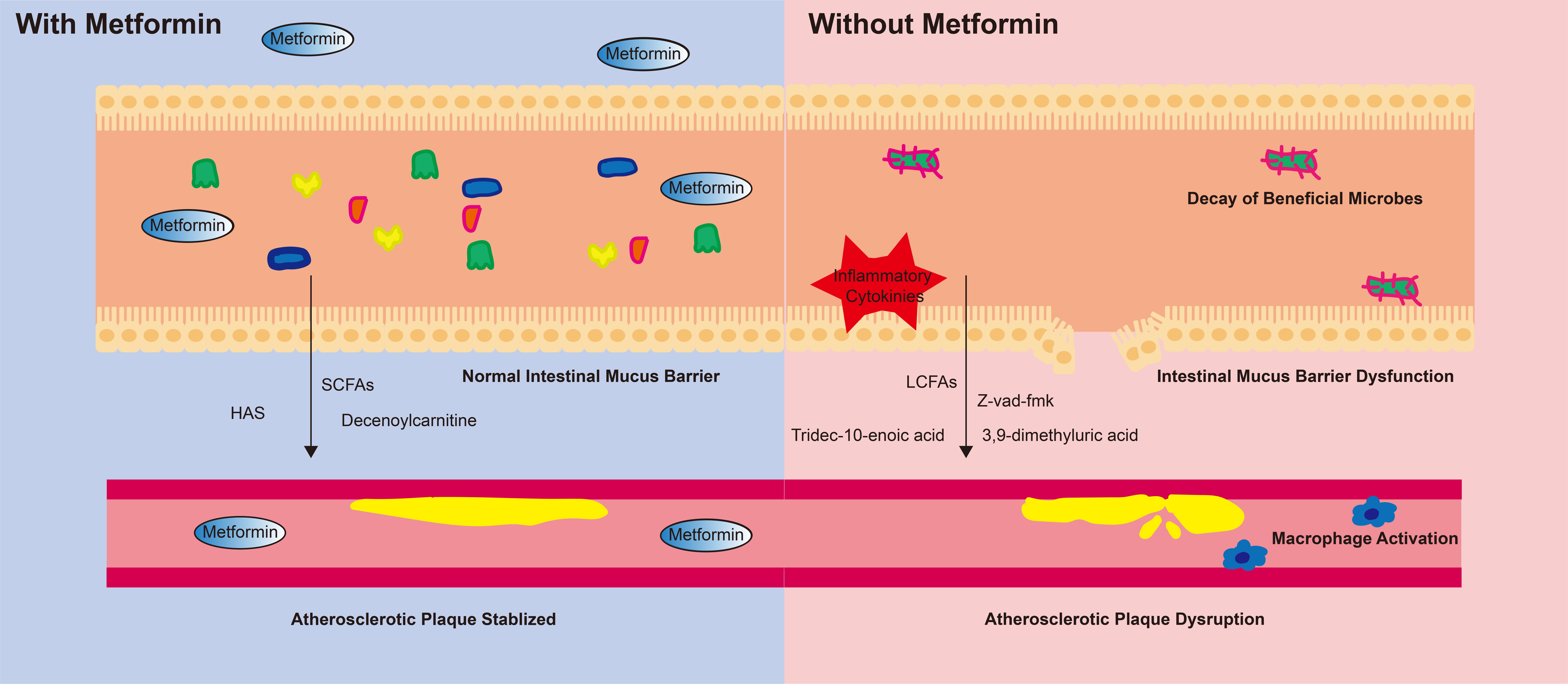 Fig. 4.
Fig. 4.
The potential mechanism of metformin’s beneficial effect on cardiovascular health. Metformin reshaped the gut microbiome by increasing the abundace of beneficial microbes and preserving the diversity of the gut microbiome. This change further maintained the stability of the intestinal mucus barrier and inhibited the production of inflammatory cytokines. The microbiome-derived metabolites helped stabilize atherosclerotic plaques. In contrast, individuals not taking metformin experienced disrupted gut microbiota diversity. The intestinal mucus barrier exhibited dysfunction, and the production of inflammatory factors was not inhibited. Moreover, the microbiome-derived metabolites promoted macrophage activation and further led to atherosclerotic plaque disruption. SCFAs, short-chain fatty acids; LCFAs, long-chain fatty acids; HAS, hydroxy-alpha-sanshool.
Our study emphasizes the long-term effects of metformin on the gut microbiome. While recent research has highlighted the interaction between metformin and the gut microbiome, it has largely overlooked its long-term impact. Although short-term use of metformin does alter the gut microbiome, these changes are minor and do not significantly affect disease prognosis. Only the long-term use of metformin and its lasting effects are convincing. The prolonged interaction between metformin and the gut microbiome leads to a state of homeostasis that ultimately influences clinical outcomes.
Several limitations of this study need to be acknowledged. First, the sample
size of patients was relatively small and the patients were all from a single
geographic area. This may affect the interpretation of gut microbiota. Second,
the study population included both CAD and DM. Hence, there are two distinct
disease variables. The age and BMI of the healthy control groups seemed to be
younger from the diseased group. Third, untargeted metabolomics had limited
accuracy in the annotation of serum metabolites. The use of Greengenes database
also has limited accuracy. We also used FDR
In conclusion, CAD patients who have been on long-term metformin therapy demonstrated a significantly improved prognosis within five years following PCI. This protective effect of metformin is further associated with gut microbiome and microbiome-associated metabolites. Multi-omics analysis revealed that metformin is strongly associated with the preservation of beneficial gut microbiota such as B. coprocola, B. massiliensis, P. succinatutens, and E. coprostanoligenes. Changes observed in serum metabolome exhibit anti-inflammatory properties, characterized by an increase in HAS and decenoylcarnitine and a decrease in Tridec-10-enoic acid, 3,9-dimethyluric acid, and Z-vad-fmk. These modifications in the gut microbiome and serum metabolome induced by metformin demonstrated beneficial effects on the five-year prognosis for CAD-DM patients on metformin therapy.
The dataset supporting the results of this article has been deposited in the Sequence Read Archive under BioProject accession code SRP167862 (https://www.ncbi.nlm.nih.gov/sra).
RLZ, QYW, SYZ, and WW conceived and designed the study. RLZ wrote the manuscript. RLZ and QYW contributed to the bioinformatics analysis and made the tables and figures. QYW conducted the literature search in chief. HQ, LW, GCL, and BZ contributed to literature search. WW critically revised the manuscript. All authors contributed to the conception and editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
The studies involving human participants were reviewed and approved by the Ethics Review Board at the Peking Union Medical College Hospital (protocol number is I-24PJ0927). Written informed consent was obtained from all participants and the study adhered to the principles of the Declaration of Helsinki.
Not applicable.
This work was supported by National High Level Hospital Clinical Research Funding (2022-PUMCH-D-002), Chinese Academy of Medical Sciences (CAMS) Innovation Fund for Medical Sciences (2021-I2M-1-003), and National High Level Hospital Clinical Research Funding (2022-PUMCH-B-098).
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/RCM26835.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.