













 , Cemil Gocmen 2, Figen Cicek 3
, Cemil Gocmen 2, Figen Cicek 31 Department of Pharmacology, Faculty of Pharmacy, Cukurova University, 01330 Adana, Turkiye
2 Department of Medical Pharmacology, Faculty of Medicine, Cukurova University, 01330 Adana, Turkiye
3 Department of Biophysics, Faculty of Medicine, Cukurova University, 01330 Adana, Turkiye
Abstract
The administration of neocuproine (NC), a copper(I)-binding agent, can induce phasic contractions in the neonatal rat bladder, while NC treatment can also promote tonic contractions in the adult rat bladder. These data may indicate that copper is important in regulating developmental contractile responses. Thus, investigating how responses to copper(I)-binding agents change during development could offer valuable insights into the mechanisms underlying overactive bladder. Therefore, this study aimed to induce developmentally different contraction responses in isolated rat whole-bladder tissues using NC.
Neonatal (1–2 weeks old) and adult (250–300 g) Wistar rats were used in the experiments. Isolated rat bladders were placed in organ baths containing 2 μM atropine and 2 μM guanethidine Krebs solution. The resulting tone changes in the preparation were recorded using a pressure transducer (cmH2O). Differences between groups were evaluated using one-way analysis of variance (ANOVA). The Student's t-test was used to assess the paired groups (GraphPad Prism); p-values smaller than 0.05 were considered statistically significant.
NC administration caused a significant increase in basal spontaneous contraction responses in neonatal rat whole-bladder tissue, whereas in adult whole bladder, NC treatment promoted a significant tonic contraction over basal tonus. In isolated whole-bladder tissue from neonatal rats, the increases in the amplitude and area under the curve (AUC) of spontaneous contraction responses induced by 50 μM NC were significantly reduced by the addition of 1 μM nifedipine, 50 μM adenosine triphosphate (ATP), 100–200 μM suramin, or 30 μM NS1619, a large-conductance Ca2+-activated K+ [BK] channel opener, to the medium. The number of spontaneous contractions decreased in the presence of NC; however, this finding was reversed in the presence of the aforementioned drugs. Moreover, the tonic contractions observed in the presence of NC in the adult bladder preparations were significantly reduced following the addition of 1 μM nifedipine, 50 μM ATP, 100 μM suramin, or 30 μM NS1619. However, the addition of 100 μM Nw-nitro-L-arginine (LA) to the medium had no significant effect on the amplitude, AUC, and frequency of NC-induced tonic contractions or spontaneous contraction responses.
These results suggest that copper may play an important role in the regulation of phasic/tonic contractions in the bladder during postnatal development.
Keywords
- copper
- urinary bladder
- potassium channels
- ATP
- muscle contraction
The bladder can produce spontaneous contractions, either small or large. It has been observed in experimental studies that increased spontaneous contractions are similar to contractions that occur without urination due to afferent pathway activation in vivo in the overactive bladder. The literature suggests that the phasic contractions occurring in isolated bladder tissue could be linked to activities during the filling phase of the micturition cycle, without leading to urination; these contractions are referred to as non-voiding contractions [1, 2, 3, 4]. While the functionally normal bladder contracts only during urination, phasic contractions occur in the detrusor even during the filling phase in the case of an overactive bladder.
The neural control and contractility of the bladder undergo significant changes during postnatal development. In bladder preparations from neonatal rats, it has been observed that the amplitude of spontaneous contractions is low during the first week of postnatal life, but the amplitude increases progressively during the second week [5, 6, 7]. Rat bladders in the early postnatal period exhibit symptoms similar to those seen in human overactive bladders. Therefore, the neonatal rat bladder can be used as an experimental model for investigating and solving overactive bladder problems [8]. Some articles suggest that neonatal and adult bladders have different characteristics [7, 9, 10, 11]. In adult bladders, contractions tend to be of low amplitude and high frequency, whereas neonatal bladders exhibit phasic contractions with higher amplitude and lower frequency. Contraction parameters in the newborn change to adult parameters approximately 4 to 5 weeks after birth. However, neonatal parameters may recur in adults with pathological conditions such as chronic urethral obstruction [9] or spinal cord injury [10]. In a healthy human bladder, atropine significantly inhibits parasympathetic nerve-mediated contractions. This inhibition indicates that cholinergic mechanisms are more dominant in parasympathetic-mediated contractions occurring in the healthy bladder. However, there are also publications suggesting that the response is not blocked by atropine (atropine-resistant) and is related to purinergic signaling [12, 13]. In an experimental study conducted on adult and neonatal rat bladders, it was observed that purinergic activity was higher in neonates compared to adults [14].
Copper is a trace element that plays an essential role in various cellular functions [15]. Studies have shown that the targets of copper include neuronal postsynaptic neurotransmitter receptors [16, 17, 18] and voltage-gated ion channels [19]. In a study conducted by Morera and colleagues [15] on skeletal muscle, it was shown that external Cu2+ causes inhibition of BKCa channel activity. Therefore, in our study, the role of Ca2+-activated K+ channels, specifically large-conductance Ca2+-activated K+ channels (BKCa), in the mechanism of spontaneous contractions observed in the presence of neocuproine has been demonstrated.
Research on copper chelators and rat bladder function has indicated that copper-dependent pathways might influence or contribute to adenosine triphosphate (ATP) or nitric oxide (NO)-driven responses [20]. Moreover, copper ions have been reported to alter the activity of P2X4 purinergic receptors expressed in Xenopus oocytes [21, 22]. This observation suggests that other P2X receptor subtypes present in the bladder could likewise be susceptible to modulation by copper ions.
A literature review revealed that the effects of copper on neonatal rat bladder tone and its mechanism of action have not been previously investigated. Therefore, this study compared the spontaneous tone changes induced by neocuproine, a copper-binding agent, in the bladders of adult and neonatal rats. While neocuproine (NC) induced spontaneous phasic contractions in the neonatal bladder, it caused an increase in tone in the adult bladder. The role of purinergic pathways, membrane Ca2+ channels, and K+ channels in this response difference was investigated.
Preliminary studies were conducted in the Department of Medical Pharmacology at the Faculty of Medicine in 2008. Experiments were initiated in the Department of Pharmacology at the Faculty of Pharmacy between 2022 and 2023, and previously conducted experiments were repeated. Data from these preliminary studies were not used.
A total of 56 neonatal (1–2 weeks old) and 42 adult (250–300 g) Wistar rats (Çukurova University Health Sciences Experimental Application and research center-SABIDAM-Adana/Turkiye), both female and male, were used in the experiments. Animals were maintained under standard laboratory conditions with a 12-hour light/dark cycle. The experimental protocols were approved by the ethics committee of Cukurova University Animal Experiments Local Ethics Committee (03-11-2004/136) and conducted in accordance with the Principles of Laboratory Animal Care as outlined by the National Institutes of Health (NIH publication 86–23, revised 1984).
Neocuproine, guanethidine, atropine, diamide, Nw-nitro-L-arginine, suramin, and ATP were prepared in distilled water. Nifedipine and NS1619 were dissolved in 75% ethyl alcohol, ensuring that the final concentration in the bath medium did not exceed 0.08%. All compounds were supplied by Sigma-Aldrich (St. Louis, MO, USA).
A total of 98 rats were anesthetized with 4–5% (v/v) halothane and decapitated. Following a midline laparotomy, the urinary bladder, together with the urethra, was excised as a single unit. Whole-bladder preparations were obtained using methods adapted from earlier studies [9, 11] A 26-gauge (26G) needle was then inserted into the bladder through the urethra and secured with 5-0 silk sutures. The other end of the needle was connected to a pressure transducer and an infusion pump (Harvard Apparatus) via a polyethylene cannula. Each preparation was placed in a temperature-controlled organ bath filled with Krebs solution (composition in mM: NaCl 113, NaHCO3 19.8, glucose 11.1, KH2PO4 1.2, KCl 4.7, MgCl2 2.5, CaCl2 1.7), maintained at 37 °C and continuously gassed with 95% O2 and 5% CO2. The bath fluid also contained atropine (2 µM) to block muscarinic cholinergic receptors and guanethidine (2 µM) to inhibit adrenergic neurotransmission, allowing assessment of non-adrenergic, non-cholinergic (NANC) activity. Preparations were allowed to equilibrate for 30 minutes prior to the initiation of the filling protocol. After this period, the tissues were slowly infused with Krebs solution using an infusion pump at a rate of 50 µL per minute. At the end of each infusion, electrical field stimulation (Grass S88; EFS, 50 volts, 1.5 ms duration, 32 Hz, 15–30 seconds) was applied. The bladder was infused until the contraction induced by EFS reached its maximum. Once the bladder reached a certain level of fullness, the infusion was stopped, and the bladder was washed with Krebs solution. All experiments were carried out with the bladder fully filled. Following a 30-minute equilibration period, drug treatments were applied while maintaining the bladder in this fully filled state. All drugs were administered 20 minutes before neocuproine (NC, 50 µM). Bladder pressure was monitored in cmH2O via a pressure transducer connected to a Data Acquisition System (BIOPAC MP 30). Spontaneous contraction amplitude and frequency were determined during a 5-minute analysis window within a 10-minute observation period. Contraction amplitude was expressed relative to the maximal response elicited by 80 mM KCl at the end of the experiment. Frequency was calculated as the number of contractions per 5 minutes. Nifedipine (1 µM; L-type Ca2+ channel blocker), N
In experiments conducted on neonatal and adult rat bladder tissues, the amplitude of the spontaneous or tonic contractions induced by the agents used was measured in cmH2O. The spontaneous and tonic contractions obtained were expressed as a percentage relative to the contraction induced by 80 mM KCl applied at the end of each experiment. Additionally, the AUC for the spontaneous or tonic contractions in neonatal and adult rat bladders was expressed as cmH2O
In experiments involving more than two groups, differences between groups were evaluated using one-way analysis of variance (one-way ANOVA). These comparisons were performed using the Tukey test (GraphPad Prism; version 5.0; GraphPad Software, San Diego, CA, USA). For the statistical evaluation of paired groups, the Student’s t-test was used. The statistical significance of the data was assessed at the level of p
Following basal tone, the administration of 50 µM NC into the organ bath caused non-reproducible spontaneous contractions in all isolated whole-bladder tissues (Fig. 1a). The amplitudes and AUCs of these spontaneous contractions showed a significant increase compared to the basal spontaneous contractions (Fig. 1b,c, p
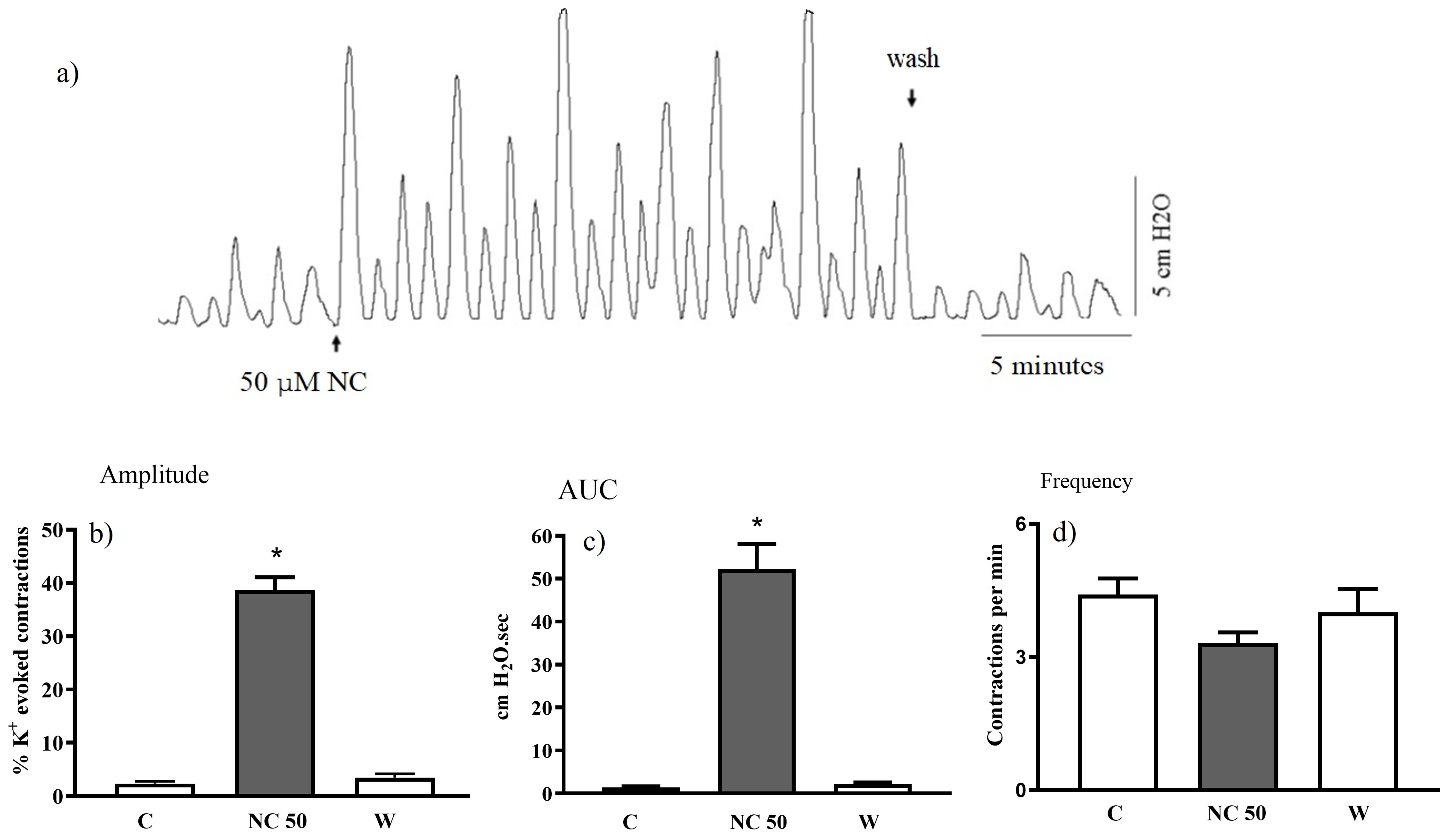 Fig. 1.
Fig. 1. Effects of 50 µM NC (NC 50) in neonatal rat bladders. (a) Representative tracing showing the effects of NC, (b) Amplitude, (c) AUC in cmH2O
In adult whole-bladder tissues, NC application did not cause phasic contractions, unlike in the neonatal bladder, but did increase basal tone (Fig. 2a). No significant changes were observed in the amplitude, AUC, and frequency of spontaneous contractions as a result of NC application (Fig. 2b–d).
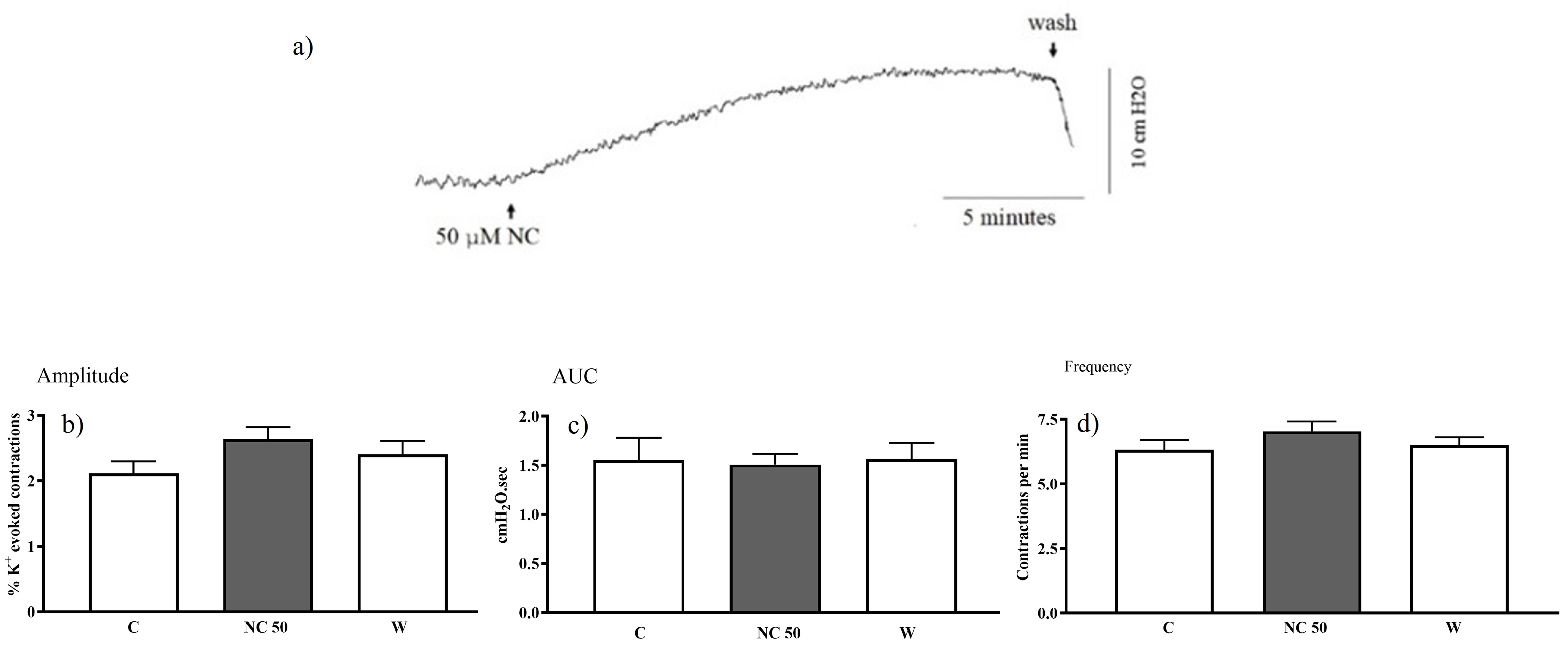 Fig. 2.
Fig. 2. Effects of 50 µM NC (NC 50) in adult rat bladders. (a) Representative tracing showing the differential effects of NC in adult bladder, (b) amplitude, (c) AUC in cmH2O
Suramin 100 µM (SUR100) or 200 µM (SUR200) significantly reduced the amplitude and AUC of spontaneous contractions induced by NC in neonatal rat bladders in a dose-dependent manner, while no significant change was observed in the frequency (Fig. 3a–c, p
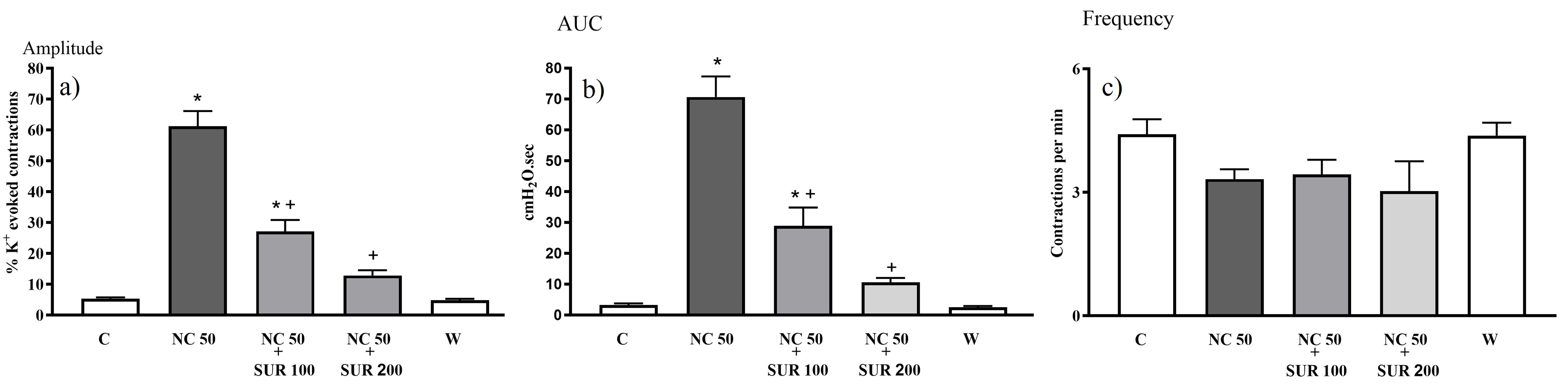 Fig. 3.
Fig. 3. The inhibitory effect of suramin 100 µM (SUR100), or 200 µM (SUR200) on the responses to 50 µM NC (NC 50) in neonatal rat bladder. (a) Amplitude, (b) AUC in cmH2O
The increase in tone caused by NC in adult bladder tissue was significantly reduced in the presence of 100 µM suramin (Fig. 4, p
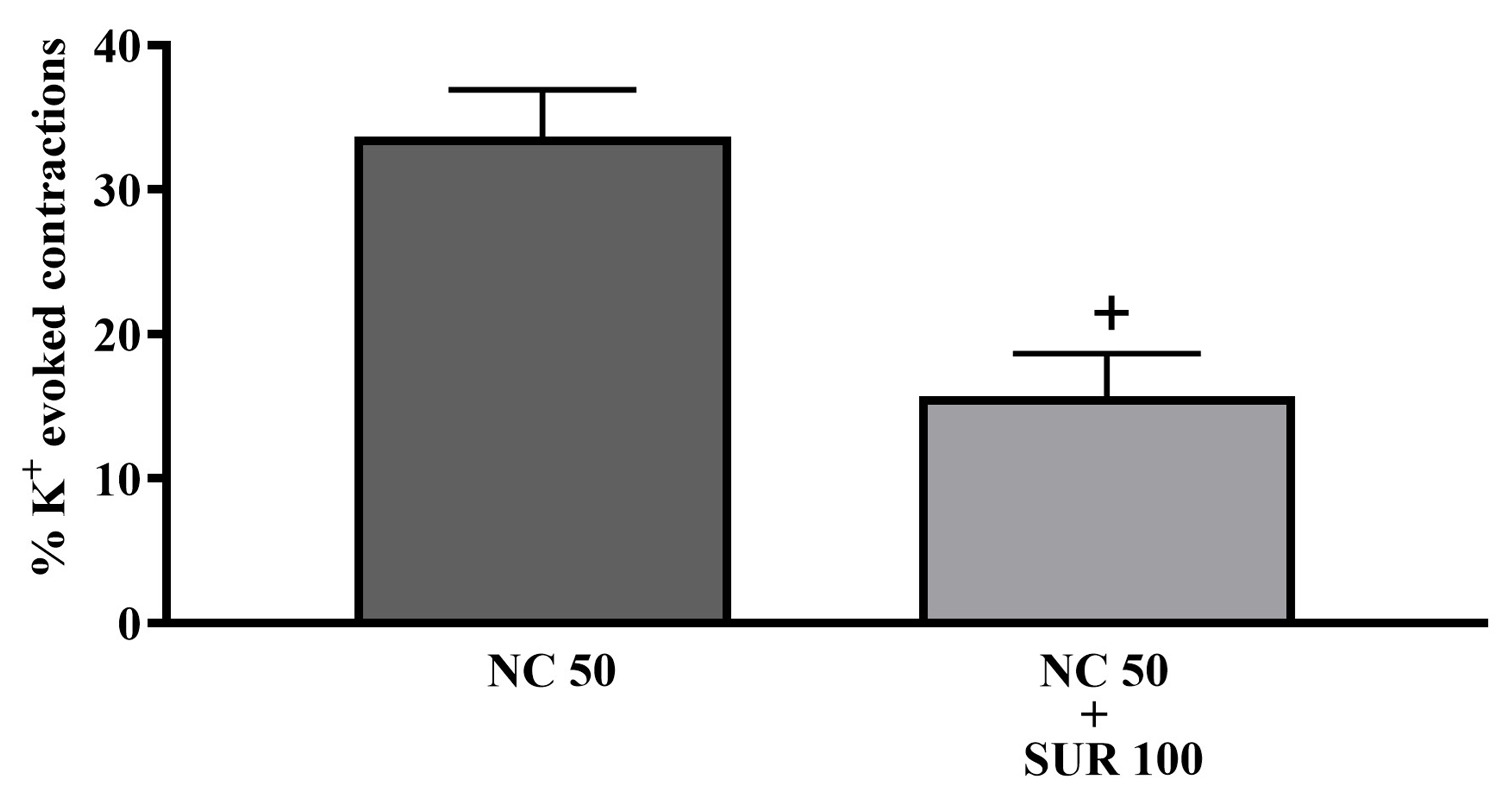 Fig. 4.
Fig. 4. The effects of suramin 100 µM (SUR100) on tonic contractions of isolated whole-bladder preparations induced by 50 µM NC (NC 50) in adult rats (n = 8). +: p
In neonatal bladder tissue, ATP (50 µM) significantly inhibited the amplitude and AUC of spontaneous contractions induced by NC (Fig. 5a,b, p
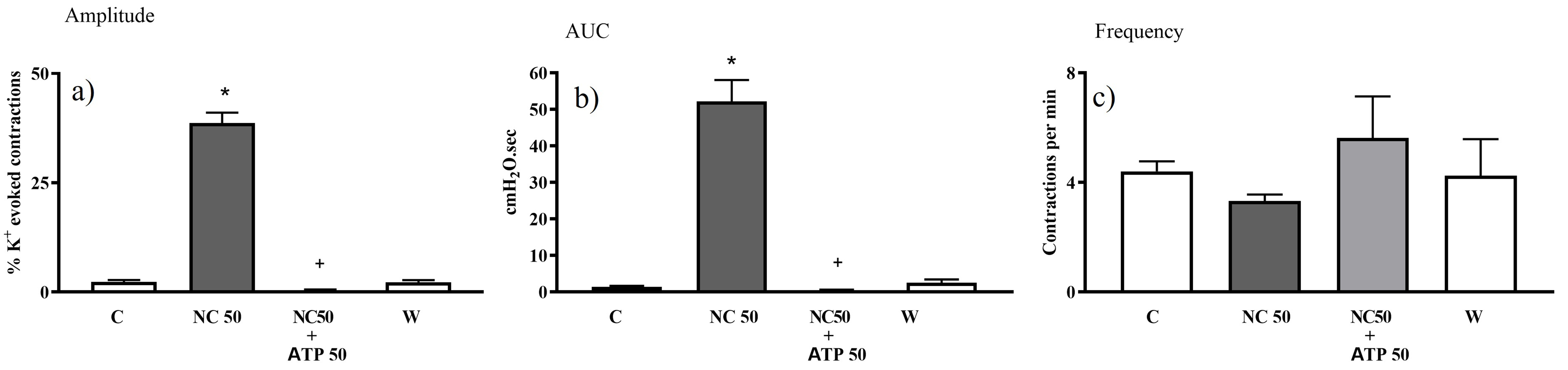 Fig. 5.
Fig. 5. The inhibitory effect of 50 µM ATP (ATP 50) on the responses to 50 µM NC (NC 50) in neonatal rat bladders. (a) Amplitude, (b) AUC in cmH2O
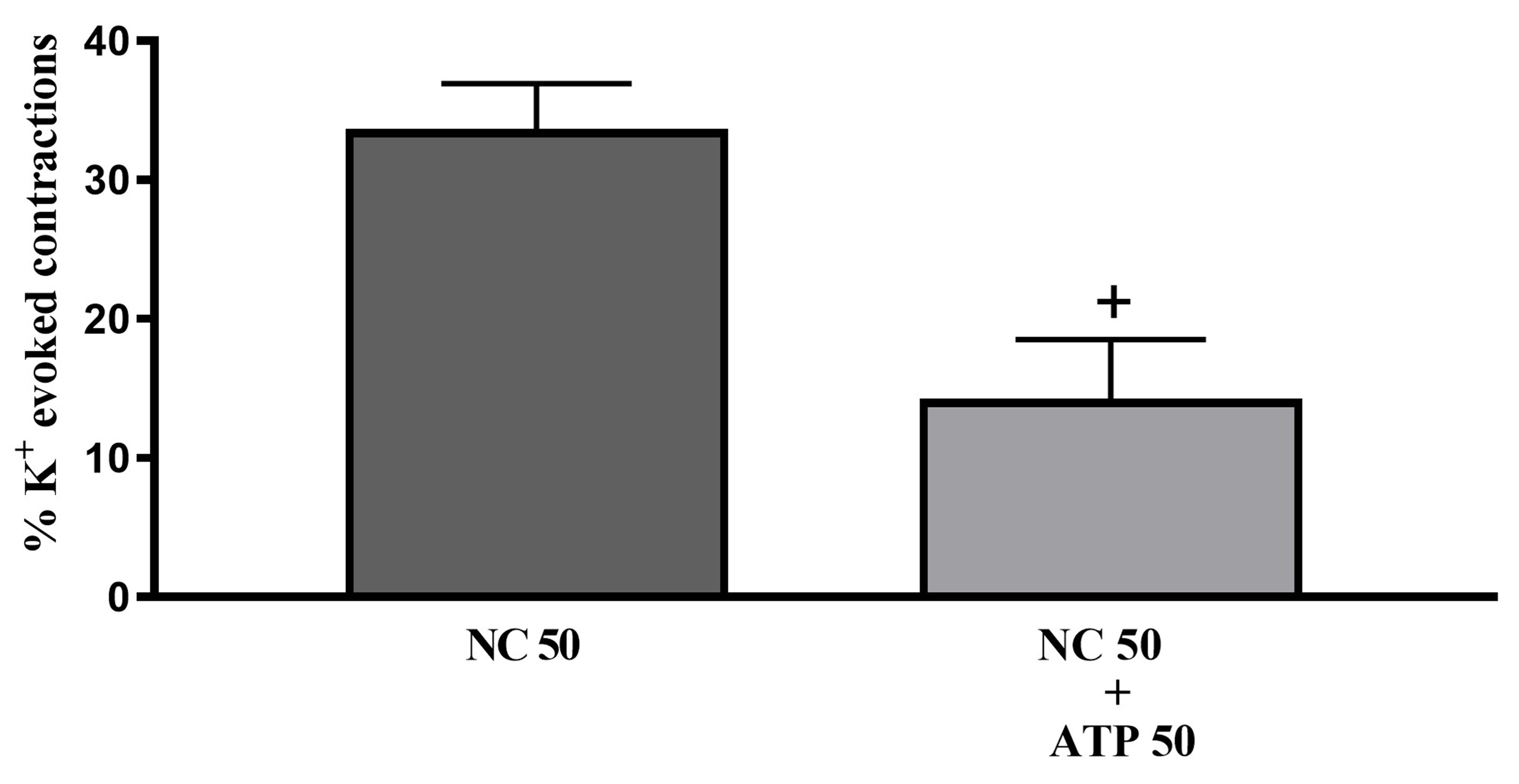 Fig. 6.
Fig. 6. The effects of 50 µM ATP (ATP 50) on tonic contractions of isolated whole-bladder preparations induced by 50 µM NC (NC 50) in adult rats (n = 8). +: p
In neonatal rat bladder tissues, NS1619 (30 µM) significantly inhibited the amplitude and AUC of spontaneous contractions induced by NC (Fig. 7a,b, p
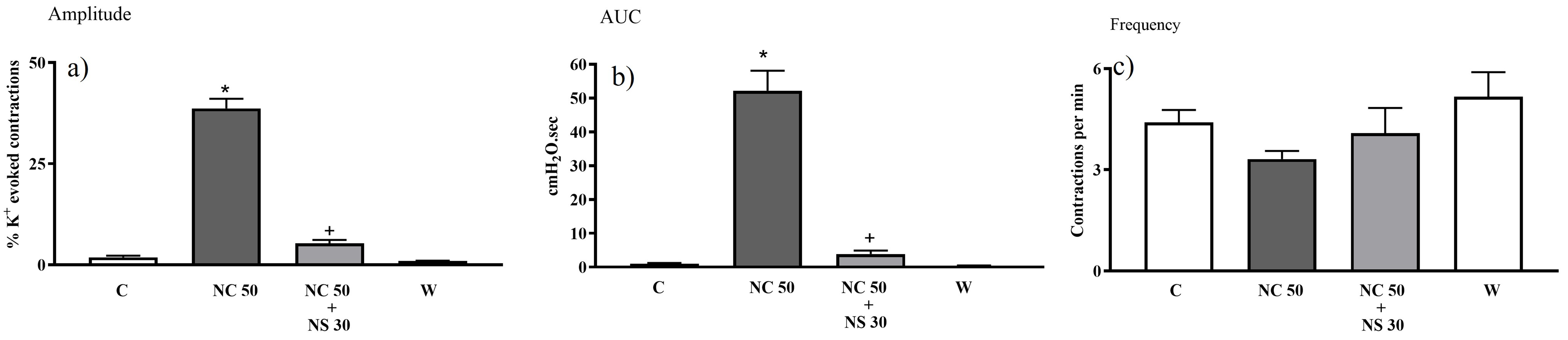 Fig. 7.
Fig. 7. The inhibitory effect of 30 µM NS1619 (NS 30) on the responses to 50 µM NC (NC 50) in neonatal rat bladders. (a) Amplitude, (b) AUC in cmH2O
In adult bladder tissue, the increase in tone induced by NC was significantly reduced in the presence of 30 µM NS1619 (Fig. 8, p
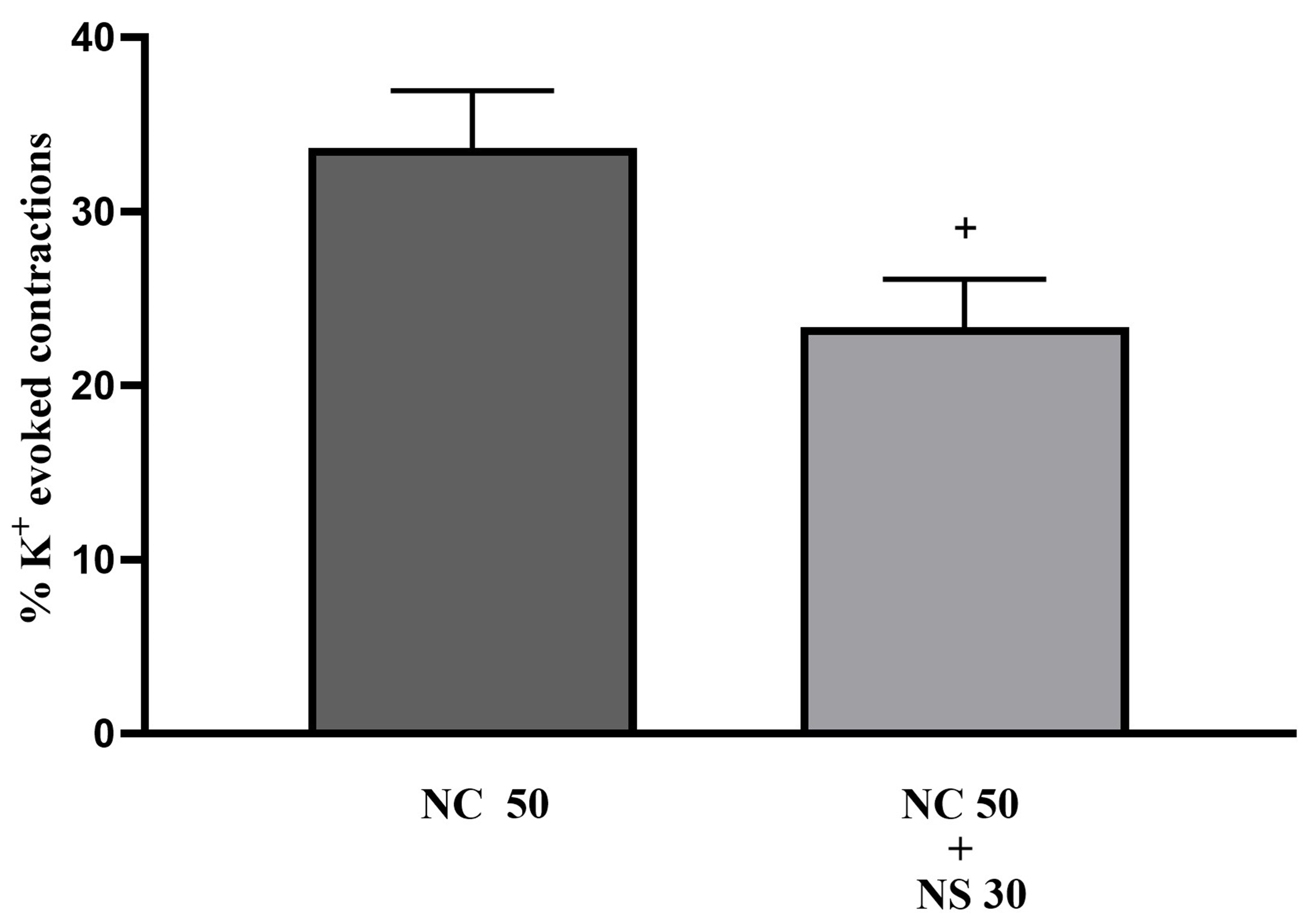 Fig. 8.
Fig. 8. The effects of 30 µM NS1619 (NS 30) on tonic contractions of isolated whole-bladder preparation induced by 50 µM NC in adult rats (n = 6). +: p
In neonatal rat bladder tissue, 1 µM nifedipine (NIF 1) significantly inhibited the amplitude and AUC of spontaneous contractions induced by NC (Fig. 9a,b, p
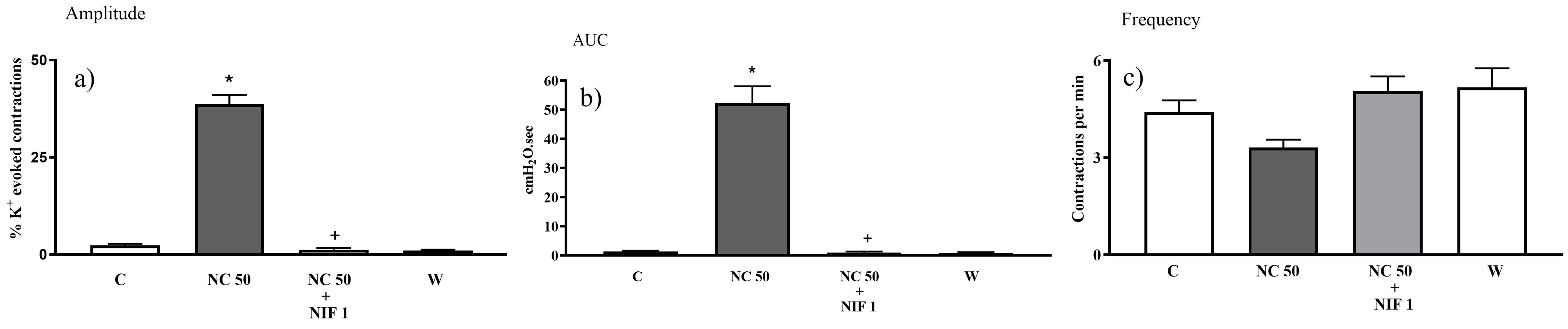 Fig. 9.
Fig. 9. The inhibitory effect of 1 µM Nifedipine (NIF 1) on the responses to 50 µM NC (NC 50) in neonatal rat bladder. (a) Amplitude, (b) AUC as cmH2O
In adult rat bladder tissue, the tonic contraction response induced by NC was significantly reduced in the presence of 1 µM nifedipine (NIF 1; Fig. 10, p
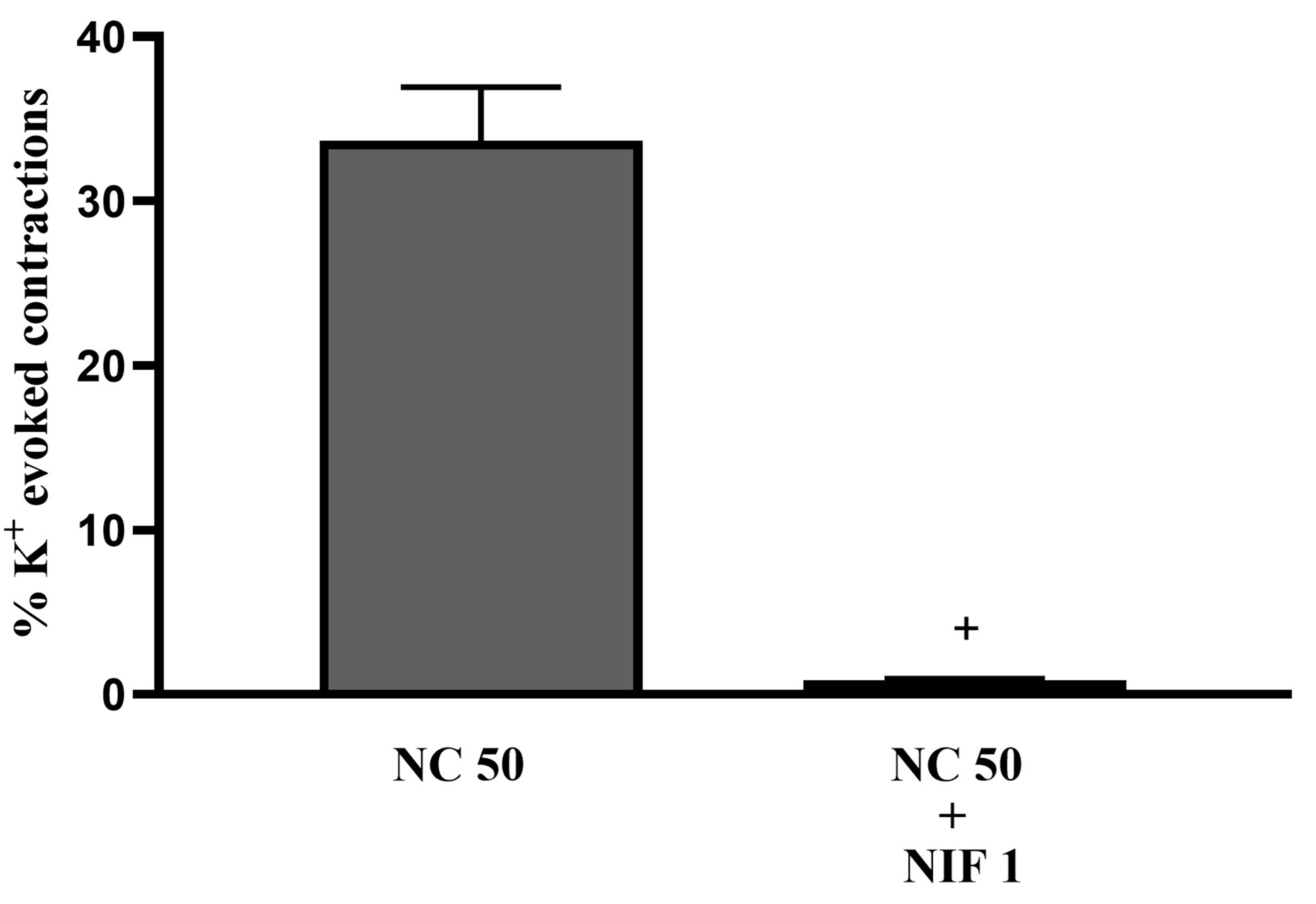 Fig. 10.
Fig. 10. The effects of 1 µM nifedipine (NIF 1) on tonic contractions of isolated whole-bladder preparations induced by 50 µM NC (NC 50) in adult rats (n = 6). +: p
In neonatal rat bladder tissue, 100 µM L-NA (L-NA 100) did not cause any significant changes in the amplitude, AUC, or frequency of spontaneous contractions induced by NC, as shown in Fig. 11a–c.
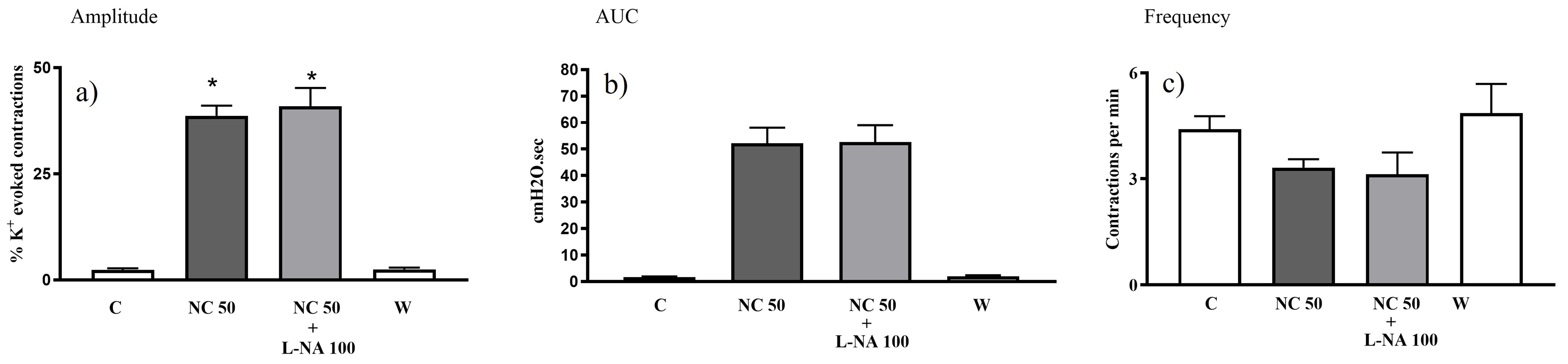 Fig. 11.
Fig. 11. The effect of 100 µM L-NA (L-NA 100) on the responses to 50 µM NC (NC 50) in neonatal rat bladder. (a) Amplitude, (b) AUC in cmH2O
In adult bladder tissue, 100 µM L-NA also did not have a significant effect on the increase in tone induced by NC (Fig. 12).
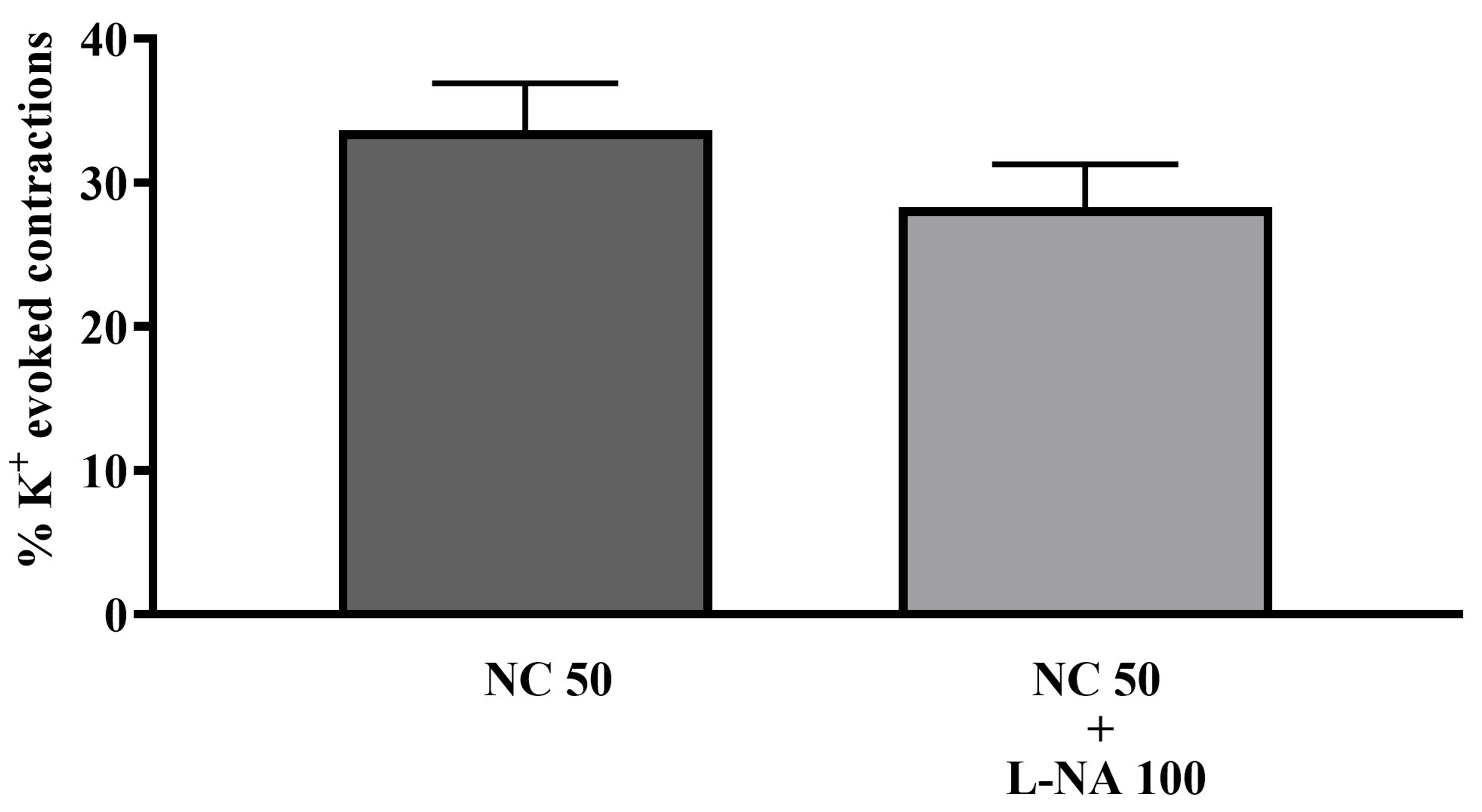 Fig. 12.
Fig. 12. The effects of 100 µM L-NA (L-NA 100) on tonic contractions of isolated whole-bladder preparations induced by 50 µM NC (NC 50) in adult rats (n = 6).
Significant changes occur in the neural control and contractile properties of the bladder during the postnatal period. Studies have shown that neonatal rat bladder preparations exhibit low-amplitude spontaneous contractions during the first week of life, but from the second week onward, these contractions gradually increase in amplitude and frequency, acquiring distinct phasic characteristics [5, 6, 7]. In contrast, low-amplitude but high-frequency contractions predominate in adult rat bladders. Therefore, in the early postnatal period, rat bladders exhibit symptoms similar to those seen in human overactive bladders, and the neonatal rat bladder can be considered a suitable experimental model for studying the pathophysiology of overactive bladder [8].
Based on these effects, this study demonstrated that neocuproine, a copper(I)-binding agent, has significantly different effects on the whole-bladder preparations isolated from neonatal and adult rats. While NC induced large amplitude phasic contractions in neonatal bladders, it only caused an increase in tone in adult bladders. Both phasic and tonic responses to NC were attenuated by an L-type Ca2+ channel blocker and a K+ channel activator, aligning with previous evidence that copper can modulate smooth muscle contractility via Ca2+ and K+ channel regulation [15, 24].
Calcium channels are essential for normal bladder function, and the detrusor muscles are the smooth muscles in the bladder responsible for bladder contraction and relaxation [25]. Preclinical and clinical studies have demonstrated that calcium influx via L-type calcium channels plays a crucial role in bladder contractions [26, 27]. In this study, the complete inhibition of copper-mediated NC effects in neonatal bladders by nifedipine, an L-type Ca2+ channel blocker, demonstrates that L-type channels are essential for these excitatory responses.
In the presence of atropine (cholinergic muscarinic receptor antagonist) and guanethidine (adrenergic neuron blocker), NC responses were unchanged in both neonatal and adult bladders, which suggests that this effect is not related to cholinergic or adrenergic mechanisms. However, ATP and suramin, a purinergic receptor antagonist, significantly inhibited NC-induced phasic/tonic contractions in the whole bladders. These findings suggest that NC enhances spontaneous/tonic activity by facilitating ATP release from nerves and/or activating purinergic pathways. Previous studies have shown that NC can modulate postjunctional purinergic signaling in bladder smooth muscle and urothelial cells [20, 28, 29] and can potentiate purinergic responses in other smooth muscle tissues, such as the rat vas deferens [30]. Research has shown that neuronal-mediated contractions of the detrusor smooth muscle persist in the presence of atropine and guanethidine, and the initial component of these contractions is suppressed by suramin, a P2X purinergic receptor antagonist [12, 31, 32, 33]. Husted and Nedergaard [34] also reported that ATP can desensitize contractile responses—a finding that may explain the attenuation of NC-induced contractions following ATP application in the present study. Moreover, ATP released from the urothelium is known to regulate afferent pathways via P2X3 receptors, influencing bladder reflex control [29, 35, 36]. Therefore, this study hypothesizes that NC may cause bladder overactivity by increasing the release of ATP from the urothelium or the effects of ATP on afferent nerves. The observed reduction in phasic contractions in neonatal rat bladders with suramin also supports this hypothesis. Considering the inhibitory effects of ATP and suramin on the phasic and tonic contractions induced by NC in neonatal and adult bladders, it can be suggested that the purinergic pathway plays a role in this mechanism. Additionally, our study indicates that a copper(I)-sensitive mechanism plays a role in the facilitatory effects of neocuproine on bladder activity. Previous studies have suggested that copper can modulate the function of P2X4, another subtype of purinergic receptors, in Xenopus oocytes [21, 22]. These results indicated that the purinergic efferent pathway in rat bladder tissue plays an important role in a copper (I)-sensitive mechanism.
This study also examined the role of the nitrergic system. Although nitric oxide (NO) released from the urothelium can influence afferent excitability [37, 38], and phasic contractions in neonatal bladders have been reported to be sensitive to NO [39], another study on the adult bladder showed that the bladder is relatively insensitive to nitric oxide [40]. The findings of the present study indicate that NC-induced facilitation persists in the presence of the NO synthase inhibitor L-NA in both age groups. This is consistent with studies on the vas deferens [30] and the mouse bladder [41] showing that NC effects are independent of NO.
Finally, studies have shown that nonselective K+ channel blockers and specific Ca2+-activated K+ channel blockers modulate spontaneous contractions in neonatal rat bladders [11, 42]. NS1619 is a potent BK channel activator with the potential to modulate cellular excitability in neurons and smooth muscle [43, 44]. BKCa channels have been shown to be important in the down-regulation of spontaneous bladder contractions postnatally during maturation [13]. Studies have demonstrated that spontaneous contractions in both normal adult and diabetic bladders can be increased by the BKCa channel blocker iberiotoxin [45, 46, 47]. A study using human detrusor smooth muscle showed that the BK channel is an important physiological regulator of smooth muscle excitability and contractility [48]. Research by Soder and Petkov [49] proposed BK channel activation with NS1619 as a therapeutic approach to control myogenic and neurogenic contractions of detrusor smooth muscle (DSM). Therefore, activation of BK channels may be an important step in controlling bladder function in the pathophysiology of detrusor overactivity.
Our results suggest that NC-induced phasic contractions in neonatal bladders depend on both purinergic signaling and BK channel activity, in a manner reminiscent of pathological excitability in adult bladders. Future work should explore the molecular interplay between copper-sensitive purinergic mechanisms and K+ channel regulation.
This study showed that neocuproine (NC), a copper-binding compound, produces different effects in isolated neonatal and adult bladder tissues. In neonatal bladders, NC-induced phasic contractions are mediated by Ca2+-activated K+ channels and/or purinergic pathways, with no significant involvement of the NO system. The bladder activity observed in neonates parallels changes seen in chronic bladder outlet obstruction and spinal cord injury models, and the early postnatal intrinsic bladder activity reappears in adult pathological states. These findings highlight the potential of copper(I)-binding agents as targets for developing new treatments for overactive and neurogenic bladder conditions.
This study investigated the possible effect of copper(I) in isolated neonatal and adult rat bladder tissue. NC, a copper(I)-binding agent, produced developmentally different responses in neonatal and adult bladders. While NC produced large amplitude low-frequency rhythmic contractions in the neonatal rat bladder, it increased basal tone in the adult rat bladder. Copper may play an important role in the regulation of phasic/tonic contractions in the bladder during postnatal development. Therefore, understanding the effect of copper-binding agents on the neonatal bladder should provide us with information about new treatment approaches in bladder pathologies.
The datasets used and analyzed during the current study are available from the corresponding author upon reasonable request.
HSB designed and performed the experiments, analyzed the results, and drafted the manuscript. CG designed and performed the experiments. FC analyzed the results. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
The study was approved by the Ethics Committee of Cukurova University Animal Experiments Local Ethics Committee (Decision No. 03-11-2004/136) and was conducted in accordance with the Principles of Laboratory Animal Care as outlined by the National Institutes of Health (NIH publication No. 86–23, revised 1984).
We thank Ahmet Kantur and Zeynep Akıllı from the Department of Pharmacology, Cukurova University for technical assistance and the Cukurova University Health Sciences Experimental Application and Research Center (SABIDAM) for the supply of rats.
This study was supported by the Cukurova University Research Foundation (TF2004D6).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.