
 , Zhili Yang 1,2,3,4,†, Lei Li 1,2,3,4, Guangzhi Zhang 1,2,3,4, Zhenyu Cao 1,2,3,4, Tianyue Liu 1,2,3,4, Xuewen Kang 1,2,3,4,*
, Zhili Yang 1,2,3,4,†, Lei Li 1,2,3,4, Guangzhi Zhang 1,2,3,4, Zhenyu Cao 1,2,3,4, Tianyue Liu 1,2,3,4, Xuewen Kang 1,2,3,4,*
1 Department of Orthopedics, Lanzhou University Second Hospital, 730000 Lanzhou, Gansu, China
2 The Second Clinical Medical College, Lanzhou University, 730000 Lanzhou, Gansu, China
3 Key Laboratory of Orthopedics Disease of Gansu Province, Lanzhou University Second Hospital, 730030 Lanzhou, Gansu, China
4 The International Cooperation Base of Gansu Province for The Pain Research in Spinal Disorders, 730030 Lanzhou, Gansu, China
†These authors contributed equally.
Abstract
Nucleus pulposus (NP) cell apoptosis and extracellular matrix (ECM) degradation constitute the two major pathological hallmarks of intervertebral disc degeneration (IVDD). Inhibiting these deleterious processes represents an effective strategy for attenuating IVDD progression. Sirtuin 7 (SIRT7), a member of the sirtuin family, plays a critical role in modulating gene expression, mediating cellular stress adaptation, and facilitating DNA repair. While SIRT7 has demonstrated therapeutic potential across diverse pathological contexts, its specific contribution to IVDD pathogenesis remains elusive. This study aimed to delineate the functional contribution of SIRT7 to IVDD progression and unravel its molecular mechanisms.
We quantified SIRT7 levels by immunohistochemistry (IHC) in degenerative human and rat NP tissues, and by quantitative reverse transcription polymerase chain reaction (qRT-PCR) in tert-butyl hydroperoxide (TBHP)-treated NP cells. To evaluate the protective capacity of SIRT7 overexpression, we conducted multifaceted analyses encompassing oxidative stress markers, apoptotic indices, ECM turnover, and nuclear factor kappa B (NF-κB) cascade activity in TBHP-challenged NP cells, utilizing reactive oxygen species detection probes, mitochondrial membrane potential indicators, Hoechst 33342 nuclear staining, qRT-PCR, western blotting, and immunofluorescence techniques. Its therapeutic potential was subsequently validated through magnetic resonance imaging and comprehensive histopathological evaluation (hematoxylin and eosin, Safranin O/Fast Green, and Masson trichrome staining) combined with IHC in a rat IVDD puncture model.
SIRT7 was consistently downregulated in degenerated human and rat NP tissues as well as TBHP-treated NP cells, concomitant with elevated NF-κB pathway activation. SIRT7 overexpression in TBHP-stimulated NP cells effectively attenuated oxidative stress, apoptosis, and ECM degradation. Mechanistically, SIRT7 overexpression may exert inhibitory effects on NF-κB signaling. Consistently, SIRT7 overexpression in the rat IVDD model decreased NF-κB activity, reduced NP cell apoptosis and ECM depletion, eventually ameliorating disc degeneration.
Our findings demonstrate that SIRT7 expression declines progressively during IVDD development. SIRT7 overexpression protects against NP cell apoptosis and ECM degradation, and this protective effect correlates with inhibition of the NF-κB pathway. These findings suggest tha SIRT7 is a guardian of NP homeostasis and highlight its substantial promise as a molecular target for IVDD therapeutics.
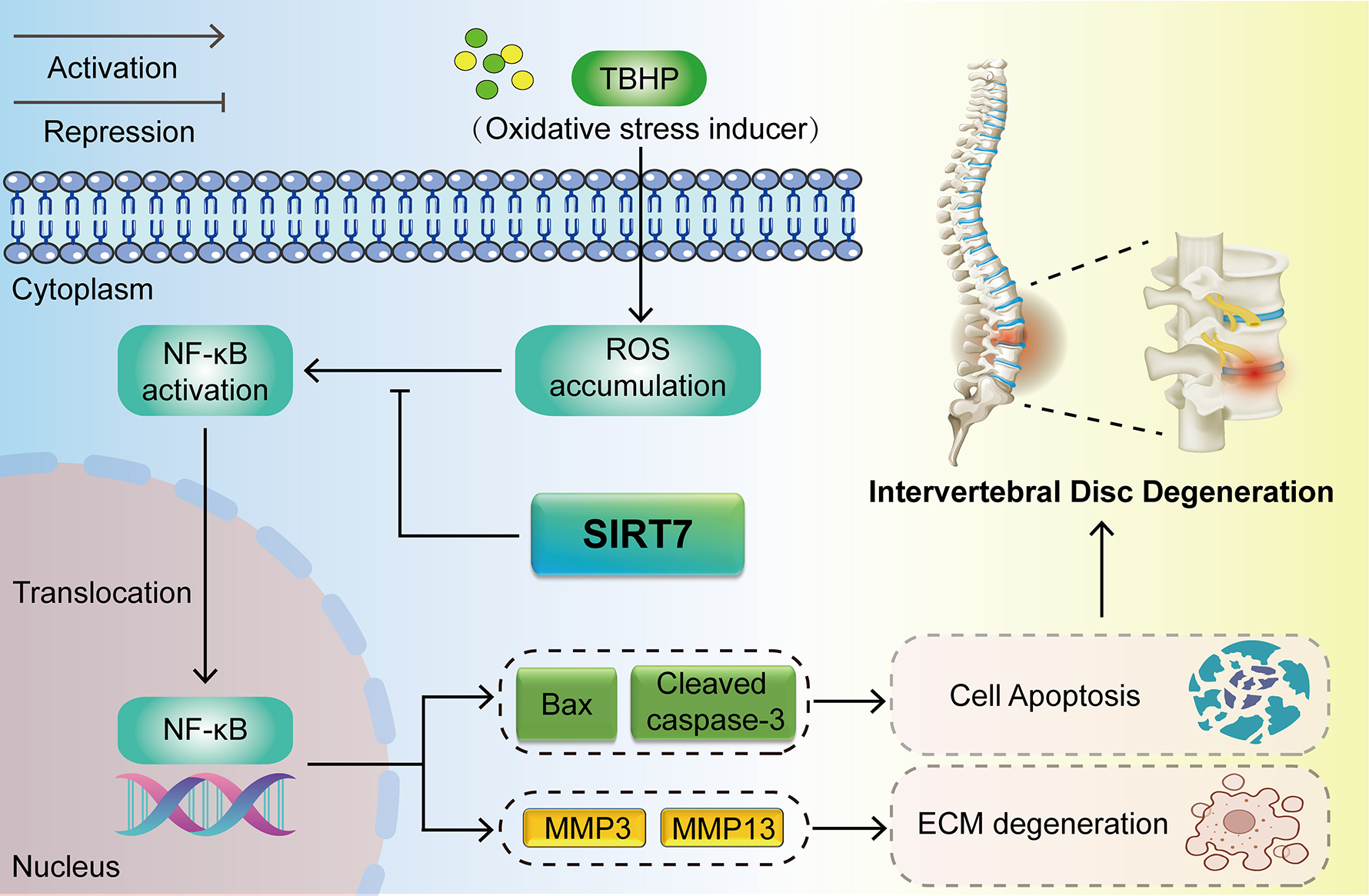
Graphical Abstract
Keywords
- intervertebral disc degeneration
- sirtuin7
- NF-κB pathway
- apoptosis
- extracellular matrix
Intervertebral disc degeneration (IVDD) is a slowly advancing pathological process involving progressive intervertebral disc (IVD) structural failure and functional impairment. This debilitating disorder culminates in chronic low back pain, imposing considerable socioeconomic burdens on healthcare systems worldwide [1]. Current management strategies, encompassing nonsteroidal anti-inflammatory agents and surgical procedures, provide merely symptomatic relief without halting disease advancement [2]. Therefore, the identification of safe and efficacious molecular targets holds substantial clinical and societal imperative.
Anatomically, the IVD comprises three distinct constituents: nucleus pulposus (NP), annulus fibrosus (AF), and cartilaginous endplates (CEP) [3]. The gelatinous NP serves as a critical shock-absorbing structure that facilitates uniform mechanical load distribution across the IVD, thereby safeguarding vertebral integrity. Through continuous synthesis of extracellular matrix (ECM) enriched in proteoglycans (predominantly aggrecan) and collagen II, NP cells preserve IVD functionality [4]. IVDD pathogenesis ensues when these resident cells succumb to apoptosis or alternative cell death modalities, accompanied by ECM degradation. Thus, diminished NP cell viability and ECM depletion represent the major pathological hallmarks of this disorder [3, 5].
The nuclear factor kappa B (NF-
Sirtuin 7 (SIRT7) belongs to the evolutionarily conserved sirtuin protein
family, functioning as a nicotinamide adenine dinucleotide (NAD+) -
dependent deacetylase [11, 12]. Despite its recent discovery, SIRT7 governs
essential cellular functions including transcription, stress adaptation, and DNA
repair, with promising implications for human disease treatment [13]. Yamamura
et al. [14] have demonstrated that SIRT7-mediated deacetylation of GATA4
attenuates pathological cardiomyocyte hypertrophy. Additionally, SIRT7 fortifies
cardiomyocyte resilience under pathological conditions, mitigating apoptosis and
inflammation-mediated cardiac injury in murine models [15]. Yu et al.
[16] established that SIRT7-mediated NRF2 activation via deacetylation enhances
cellular antioxidant capacity and confers protection against chemotherapy-induced
hepatic damage. Moreover, SIRT7 exerts protective effects against pulmonary
hypertension through Krüpple-like factor 4 (KLF4) deacetylation and dampens
inflammatory responses via NF-
Accordingly, the present investigation sought to evaluate SIRT7 expression and
explore its role in IVDD. We found that SIRT7 expression was markedly
downregulated during IVDD progression. Functional rescue experiments revealed
that SIRT7 overexpression could significantly inhibit NF-
We obtained 12 NP tissue samples from patients who underwent surgery at the Department of Orthopedics, Lanzhou University Second Hospital, in 2025. During their lumbar IVD surgery, NP tissue was removed and transported to the laboratory. The samples were divided into the following two groups: a mild degeneration group (Grade II), which included NP tissues from three males and three females aged 22–46 years (mean age: 32.9 years), and a severe degeneration group (Grade V), which included NP tissues from three males and three females aged 47–69 years (mean age: 60 years). Exclusion criteria included history of spinal tumors, tuberculosis, or infectious diseases, consistent with previous studies [20].
For rat NP cell isolation, 4–6 weeks old Sprague–Dawley rats were deeply anesthetized via inhalation of 3–4% isoflurane until loss of pedal reflex, followed immediately by cervical dislocation as a secondary physical method to ensure definitive euthanasia [21], in accordance with the American Veterinary Medical Association (AVMA) Guidelines for the Euthanasia of Animals (2020). NP tissues were harvested from the IVD and digested with 0.5% type II collagenase (BS164; Biosharp, Hefei, Anhui, China) at 37 °C for 2 hours. The resulting cell suspension was cultured into DMEM/F-12 medium (C11330500BT, Gibco, Thermo Fisher Scientific, Suzhou, Jiangsu, China) supplemented with 10% FBS (FSP500, ExCell Bio, Suzhou, Jiangsu, China) and maintained in a humidified incubator at 37 °C with 5% CO2 atmosphere. Upon reaching 80–90% confluency, cells were subcultured employing 0.25% trypsin–EDTA solution (T1300, Solarbio, Beijing, China). The cultured rat NP cells were validated for the NP cell–specific marker aggrecan by immunofluorescence staining (Supplementary Material) and tested negative for mycoplasma contamination. An oxidative stress environment was established in vitro by exposing the NP cells to the indicated concentrations of tert-butyl hydroperoxide (TBHP) (458139; Sigma-Aldrich, St. Louis, MO, USA) for 48 h. All in vitro experiments were independently performed three times.
Total RNA was extracted with RNAiso Plus Reagent (9109, Takara Bio Inc.,
Kusatsu, Japan) and assessed for purity and concentration using NanoDrop 2000.
After genomic DNA removal, the first-strand complementary DNA (cDNA) was
synthesized using the Hifair® III 1st Strand cDNA Synthesis
SuperMix for qPCR (gDNA digester plus) kit (H4509060, Yeasen Biotech, Shanghai,
China). Quantitative polymerase chain reaction (PCR) proceeded on a QuantStudio 3
instrument (Thermo Fisher Scientific, Waltham, MA, USA) with
Hieff® qPCR SYBR Green Master Mix (Low Rox Plus) kit (11202ES08,
Yeasen Biotech, Shanghai, China) following the manufacturer’s protocol. Relative
expression was calculated by 2–ΔΔCt
method with
| Gene | Forward primers 5′-3′ | Reverse primers 5′-3′ |
| SIRT7 | TCCGTCAGGTGTCACGCATC | CCCTGAAGCTCGGTCACCA |
| Aggrecan | TGTCACTTCCCAACTATCCAGC | CCATGCATCACTTCACACCGAT |
| Collagen II | CCCCGGTCTTCCTGGTGTCA | CGCTCTCACCCTTCACACCT |
| MMP3 | AACACTATGGAGCTGATGCAC | CTCAATGGCAGAATCCACAC |
| MMP13 | CTACCATCCTGTGACTCTTGCG | TTTGCCAGTCACATCTAAGCC |
| Bax | GCCCCAGGACGCATCCACC | ACATGTCAGCTGCCACACGGAA |
| Bcl2 | GGATGACTTCTCTCGTCGCTAC | ACACATGACCCCACCGAAC |
| CATTGTCACCAACTGGGACGA | CATCTTTTCACGGTTGGCCTT |
qRT-PCR, quantitative real-time polymerase chain reaction.
NP cells were homogenized in ice-cold radioimmunoprecipitation assay (RIPA)
buffer (P0013B, Beyotime, Shanghai, China) supplemented with protease inhibitor
phenylmethylsulfonyl fluoride (PMSF) (ST506, Beyotime, Shanghai, China) and
phosphatase inhibitors (P1091, Beyotime, Shanghai, China). Protein concentrations
were spectrophotometrically measured via bicinchoninic acid (BCA) method (PC0020,
Solarbio, Beijing, China). Equal amounts of protein were electrophoretically
separated on polyacrylamide gels, transferred to polyvinylidene difluoride (PVDF)
membranes (Millipore, Billerica, MA, USA), and blocked with 5 % skim milk or
BSA. Membranes were subsequently probed with primary antibodies targeting SIRT7
(1:1000, 12994-1-AP, Proteintech, Wuhan, Hubei, China), matrix
metalloproteinase-3 (MMP3) (1:4000, 66338-1-Ig, Proteintech, Wuhan, Hubei,
China), phospho-p65 (p-p65, Ser536, 1:1000, 3033, CST, Danvers, MA, USA), total
p65 (1:1000, 8242, CST, Danvers, MA, USA), or
Intracellular ROS accumulation was evaluated utilizing dihydroethidium (DHE) fluorescent probe (S0064S; Beyotime, Shanghai, China). Following loading with 10 µM DHE for 30 min, NP cells underwent triple PBS washes prior to immediate assessment of DHE-derived fluorescence via flow cytometric analysis (BD Biosciences, Franklin Lakes, NJ, USA) or inverted fluorescence microscopy (Zeiss, Jena, Thuringia, Germany).
SIRT7-overexpressing lentiviruses (Lv-SIRT7) and a control lentivirus (Lv-NC) were purchased from GeneChem (Shanghai, China). At ~30% confluency, NP cells were transfected with Lv-SIRT7 (MOI = 50). Medium was changed after 48 h. SIRT7 overexpression was verified by qRT-PCR and Western blotting.
After exposure to TBHP, NP cells were stained with 2 µM Hoechst 33342 (C1025, Beyotime, Shanghai, China) for 15 min at 37 °C in darkness. They were then rinsed thrice with PBS and observed under an inverted fluorescence microscope (Zeiss, Jena, Thuringia, Germany). Apoptosis was identified based on nuclear condensation, fragmentation, and intense blue fluorescence.
NP cells plated in 24-well or confocal dishes underwent fixation with 4%
paraformaldehyde for 20 min, followed by permeabilization using 0.5% Triton
X-100 and blocking with 10% goat serum for 1 h. Overnight probing at 4
°C was conducted employing primary antibodies targeting aggrecan (1:200, DF7561, Affinity Bioscience, Changzhou, Jiangsu, China), collagen II
(1:200, 28459-1-AP, Proteintech, Wuhan, Hubei, China), matrix
metalloproteinase-13 (MMP13) (1:200, YT2796, Immunoway, Suzhou, Jiangsu, China),
Bax (1:200, 60267-1-Ig, Proteintech, Wuhan, Hubei, China), Bcl-2 (1:200,
68103-1-Ig, Proteintech, Wuhan, Hubei, China), cleaved caspase-3 (1:200, 9664,
CST, Danvers, MA, USA), or p65 (1:400, 8242, CST, Danvers, MA, USA). Subsequent
to triple PBS rinses, specimens were exposed to cyanine 3- conjugated goat anti-rabbit IgG (1:200, GB21303, Servicebio, Wuhan, Hubei, China) or cyanine 3-conjugated goat anti-mouse IgG (1:200, GB21301, Servicebio, Wuhan, Hubei, China) for 1 h and nuclei were labeled with DAPI. Microscopic examination was
carried out utilizing inverted fluorescence or confocal microscopy (Zeiss, Jena,
Thuringia, Germany). For quantitative analysis, fluorescence intensity was measured using ImageJ software. Images were acquired under identical exposure settings across all groups. For NF-
The rat model of IVDD was established according to a previously published
protocol [22]. Twenty-eight 8-week-old male Sprague–Dawley rats (obtained from
the Laboratory Animal Center of Lanzhou University) were randomly allocated into
four groups: Sham, IVDD, IVDD + Lv-NC, and IVDD + Lv-SIRT7. The rats were sedated
with 2% isoflurane (R510-22-10, RWD Life Science, Shenzhen, Guangdong, China)
and subsequently anesthetized with 40 mg/kg intraperitoneal pentobarbital (SCRC
69020180, Sinopharm Chemical Reagent Shanghai Co., Ltd, China). Following sterile exposure of the Co7/8 disc, the NP was punctured with a 21G needle parallel to the CEP (360° rotation, 60 s), then injected with 3 µL of Lv-NC or Lv-SIRT7 (titer: 1
MRI was conducted to evaluate the impact of SIRT7 on disc degeneration. At the
eighth week post-operation, T2-weighted sagittal images were acquired using a uMR
9.4T small-animal MRI scanner (United Imaging Life Science Instrument, Wuhan,
Hubei, China) via fast-spin echo (TR/TE = 3000/40 ms; matrix, 224
Human NP and rat IVD samples were preserved in 4% paraformaldehyde, with rat samples additionally decalcified prior to processing. Following dehydration and paraffin embedding, 4 µm sections were stained with: hematoxylin and eosin (H&E) (G1120, Solarbio, Beijing, China) for general tissue architecture and cellularity; Safranin O/Fast Green (G1371, Solarbio, Beijing, China) and Alcian blue (G1560, Solarbio, Beijing, China) for matrix proteoglycans; Masson staining (G1346, Solarbio, Beijing, China) for collagen and fibrosis. Protocols followed manufacturer specifications, with Olympus microscopy used for imaging.
Paraffin sections of human NP and rat IVD samples underwent dewaxing,
rehydration, and citrate-based antigen retrieval. Peroxidase quenching utilized
3% hydrogen peroxide (10 min) prior to overnight primary antibody binding at 4
°C: SIRT7 (1:200, DF6161, Affinity Bioscience, Changzhou, Jiangsu,
China), aggrecan (1:100, DF7561, Affinity Bioscience, Changzhou, Jiangsu, China), collagen II (1:200, ab307674, Abcam, Cambridge, UK), MMP3 (1:200,
ab52915, Abcam, Cambridge, UK), Bax (1:400, 60267-1-Ig, Proteintech, Wuhan,
Hubei, China), or p65 (1:400, 8242, CST, Danvers, MA, USA). Immunoreactivity was
visualized using an IHC detection kit (KIT-9710, Maixin Biotech Co., Fuzhou,
Fujian, China) and DAB peroxidase substrate kit (KIT-0038, Maixin Biotech Co.,
Fuzhou, Fujian, China) per manufacturer instructions. For quantitative analysis, at least three fields within the NP region were randomly selected from each section, and positively stained cells (brown, DAB) and total cells (hematoxylin counterstain) were counted in each field. The percentage of positive cells was calculated as (positively stained cells / total cells)
Quantitative results represent mean
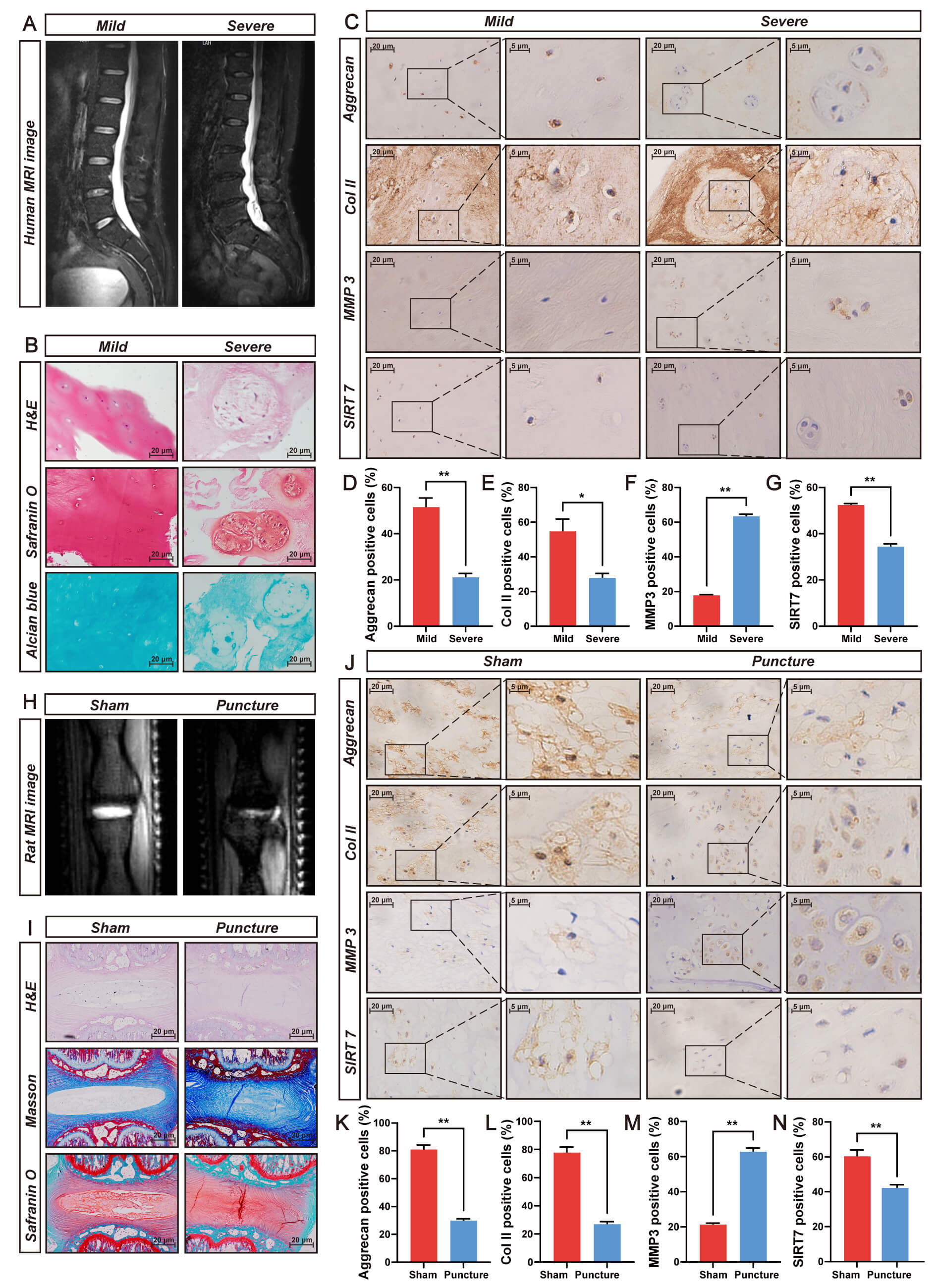
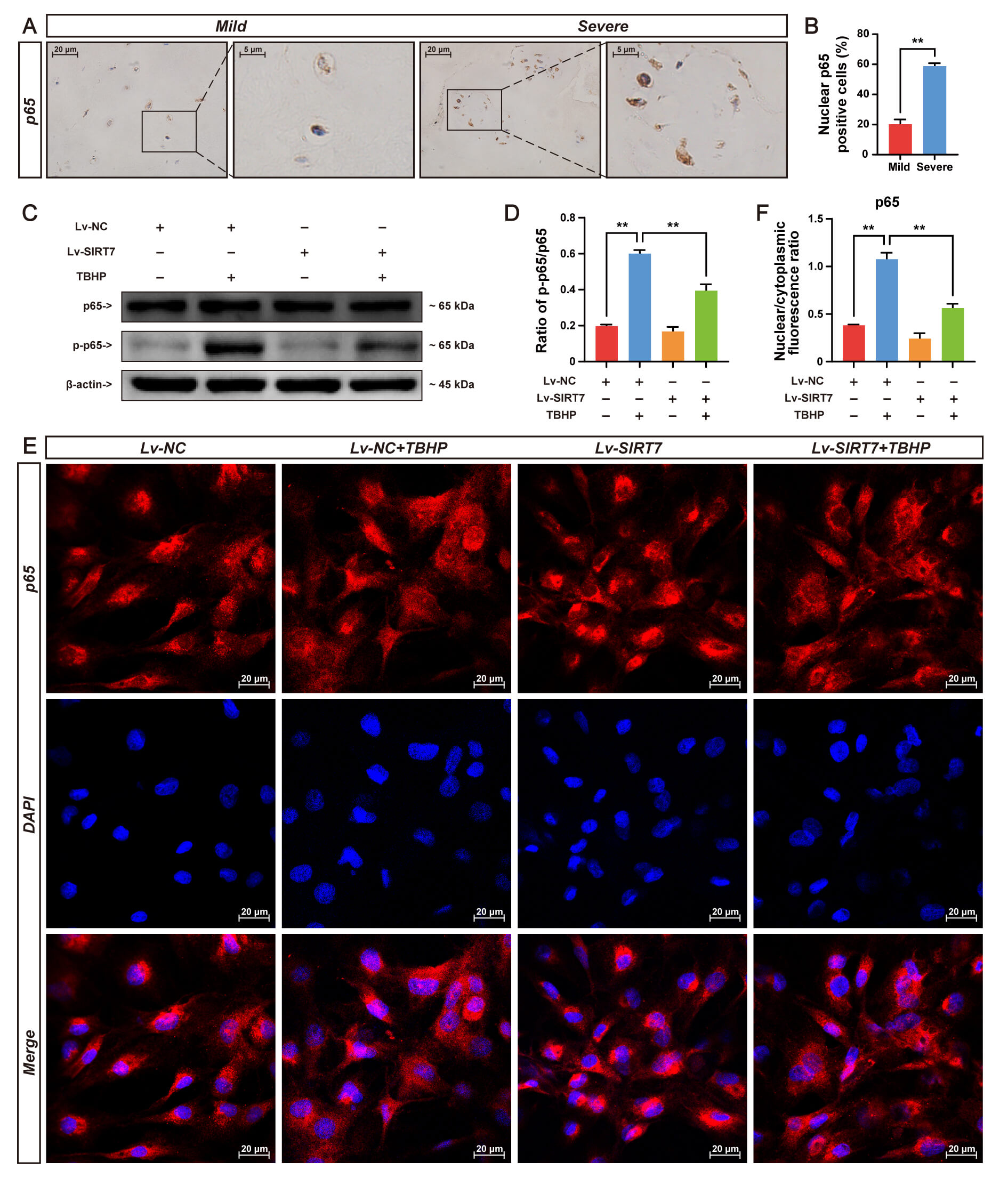
SIRT7 expression and its association with IVDD were evaluated using human NP tissue of varying degenerative severity. Pfirrmann classification by T2-weighted MRI (Fig. 1A) was histologically confirmed through H&E, safranin O/fast green, and Alcian blue staining (Fig. 1B) and validated via IHC staining for aggrecan, collagen II, and MMP3 (Fig. 1C–F). IHC analysis revealed significantly lower SIRT7 levels in severe versus mild NP degeneration (Fig. 1C,G), indicating SIRT7 expression decline during disease progression.
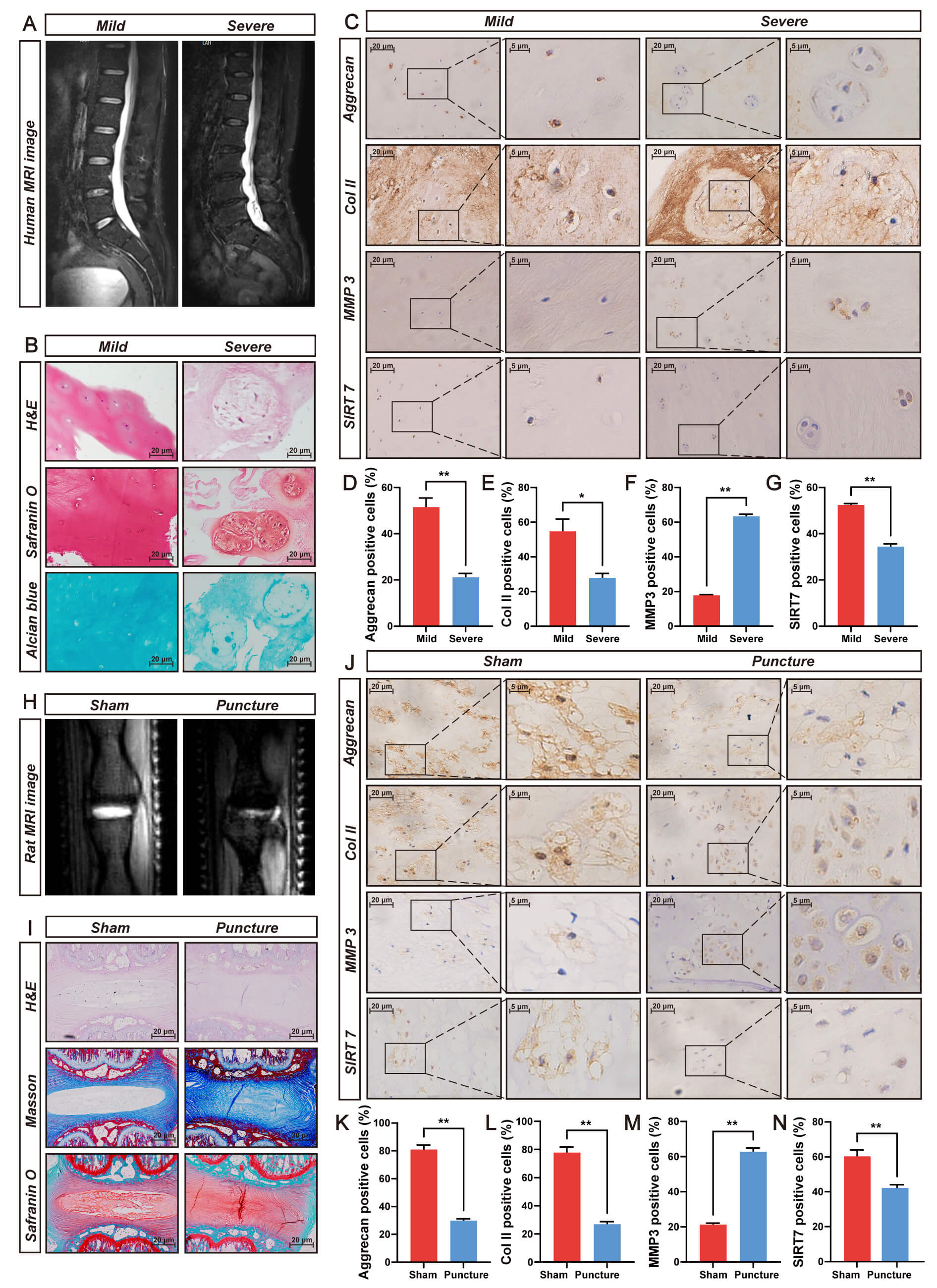 Fig. 1.
Fig. 1.
SIRT7 expression is markedly downregulated in degenerated NP
tissues from humans and rats. (A) Representative T2-weighted MRI scans
illustrating mild (Pfirrmann Grade II) and severe (Pfirrmann Grade V) disc
degeneration in patients. (B) Histological characterization of human NP specimens
employing H&E, Safranin O/fast green, and Alcian blue staining. Scale bar: 20
µm. (C) IHC staining of aggrecan, Collagen II (Col II), MMP3, and
SIRT7 in degenerative human NP tissue. Scale bar: 20 µm (main images) and 5 µm (insets). (D)
Quantitative analysis of aggrecan immunoreactivity in degenerative human NP
tissue. (E) Quantitative analysis of Col II immunoreactivity in degenerative
human NP tissue. (F) Quantitative analysis of MMP3 immunoreactivity in
degenerative human NP tissue. (G) Quantitative analysis of SIRT7 immunoreactivity
in degenerative human NP tissue. (H) Typical T2-weighted MRI images captured from
rats at 8 weeks post-needle puncture-induced IVDD surgery. (I) Histopathological
evaluation of rat NP tissue utilizing H&E, Safranin O/fast green, and Masson’s
trichrome staining. Scale bar: 20 µm. (J) IHC detection of aggrecan,
Col II, MMP3, and SIRT7 expression patterns in degenerated rat NP tissue
sections. Scale bar: 20 µm (main images) and 5 µm (insets). (K) Quantitative analysis of positive
staining rates for aggrecan in degenerative rat NP tissue. (L) Quantitative
analysis of positive staining rates for Col II in degenerative rat NP tissue. (M)
Quantitative analysis of positive staining rates for MMP3 in degenerative rat NP
tissue. (N) Quantitative analysis of positive staining rates for SIRT7 in
degenerative rat NP tissue. Quantitative results represent mean
These findings were verified in a puncture-induced rat IVDD model. Eight weeks post-surgery, MRI revealed reduced T2 signal intensity in punctured versus sham discs (Fig. 1H). Histological assessment via H&E, safranin O/fast green, and Masson staining revealed disrupted NP architecture, proteoglycan depletion, and fibrosis (Fig. 1I). Concurrent IHC analysis indicated decreased aggrecan, collagen II, and SIRT7 levels alongside elevated MMP3 (Fig. 1J–N). Together, these data suggest SIRT7 downregulation during IVDD.
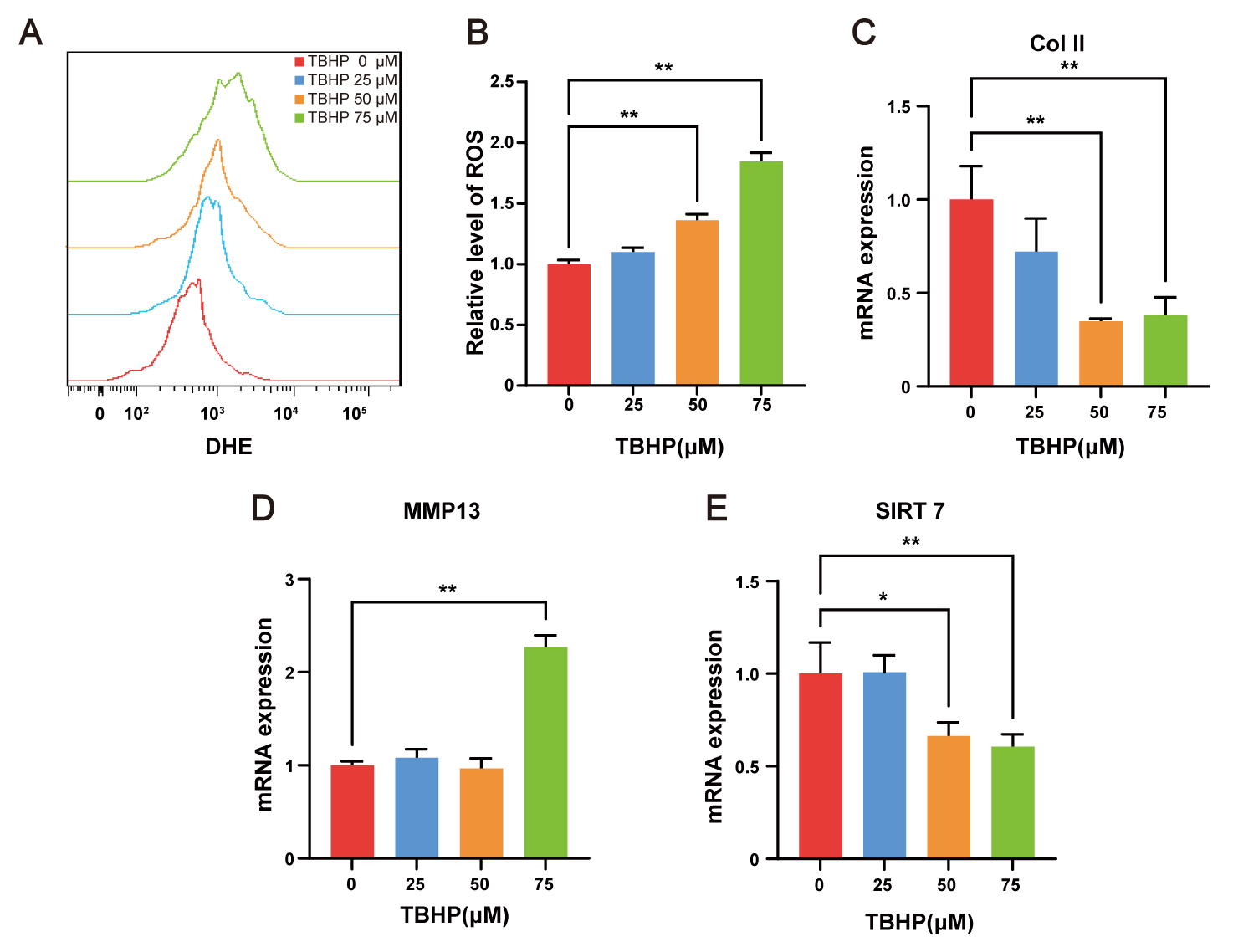
ROS are considered a major pathological driving factor of IVDD. TBHP, an exogenous ROS donor, is widely used to establish cell models of IVDD in vitro [23]. To induce oxidative stress, NP cells were exposed to indicated concentrations of TBHP for 48 h. Intracellular ROS levels were subsequently quantified by flow cytometry following DHE labeling, revealing dose-dependent ROS accumulation (Fig. 2A,B). The qRT-PCR analysis demonstrated that 75 µM TBHP markedly suppressed collagen II expression (Fig. 2C) while upregulated MMP13 expression (Fig. 2D) in NP cells, thereby confirming the degenerative phenotype. Following validation of the cellular model, transcript level analysis revealed that SIRT7 expression was significantly downregulated in NP cells treated with 50 and 75 µM TBHP (Fig. 2E). These in vitro findings align with prior in vivo observations, collectively indicating decreased SIRT7 expression in NP cell degeneration. Accordingly, 75 µM TBHP was selected for subsequent experiments.
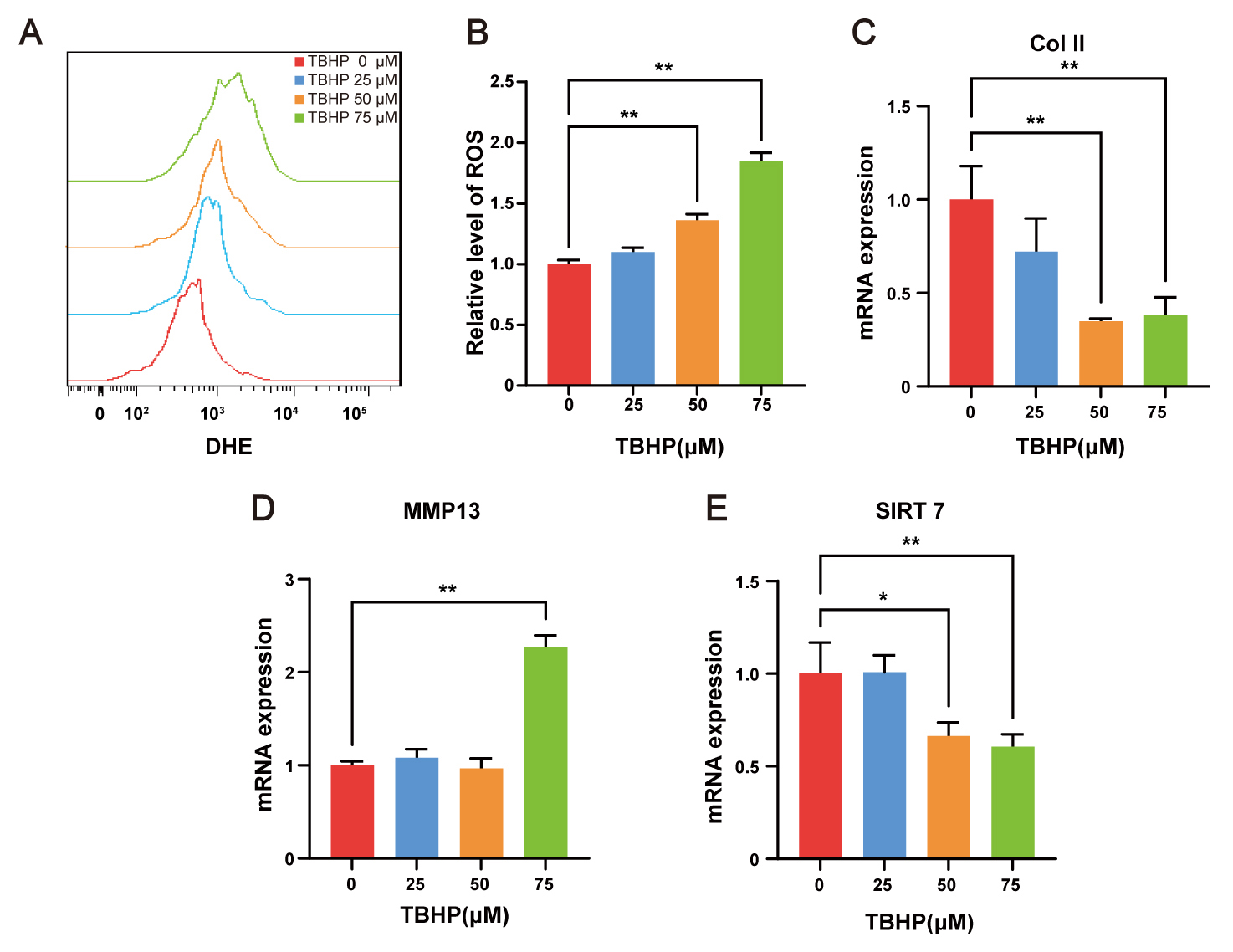 Fig. 2.
Fig. 2.
SIRT7 is downregulated in TBHP-treated NP cells. (A) Flow
cytometry histograms of DHE fluorescence after 48 h TBHP treatment. (B)
Quantitative analysis of relative intracellular ROS levels, expressed as fold
change in DHE mean fluorescence intensity relative to the control group. (C)
qRT-PCR results for Col II in NP cells. (D) qRT-PCR results for MMP13 in NP
cells. (E) qRT-PCR results for SIRT7 in NP cells. Values represent mean
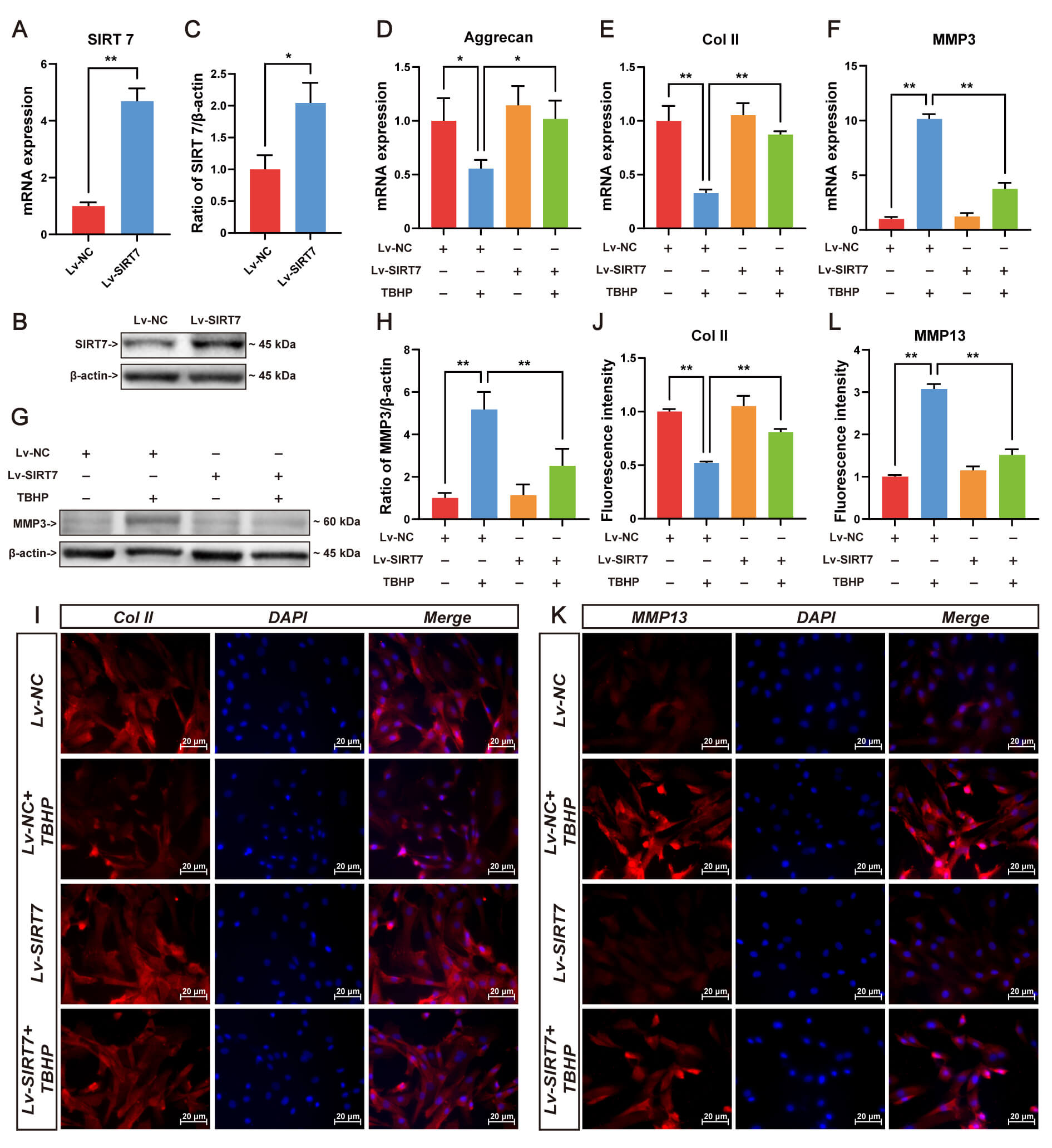
ECM homeostasis disruption underlies IVDD pathogenesis. To elucidate SIRT7’s regulatory role in matrix metabolism, lentiviral-mediated SIRT7 overexpression was established in NP cells. The qRT-PCR (Fig. 3A) and Western blotting were performed to verify efficient SIRT7 upregulation in NP cells (Fig. 3B,C). Following oxidative challenge with TBHP, transcriptional profiling revealed substantial repression of aggrecan and collagen II alongside MMP3 induction, while SIRT7 overexpression reversed these TBHP-induced alterations (Fig. 3D–F). At the protein level, SIRT7 attenuated MMP3 accumulation (Fig. 3G,H), enhanced collagen II deposition (Fig. 3I,J), and diminished MMP13 immunoreactivity (Fig. 3K,L). Collectively, these findings suggest that SIRT7 upregulation preserves matrix integrity under oxidative stress, thereby retarding degenerative progression.
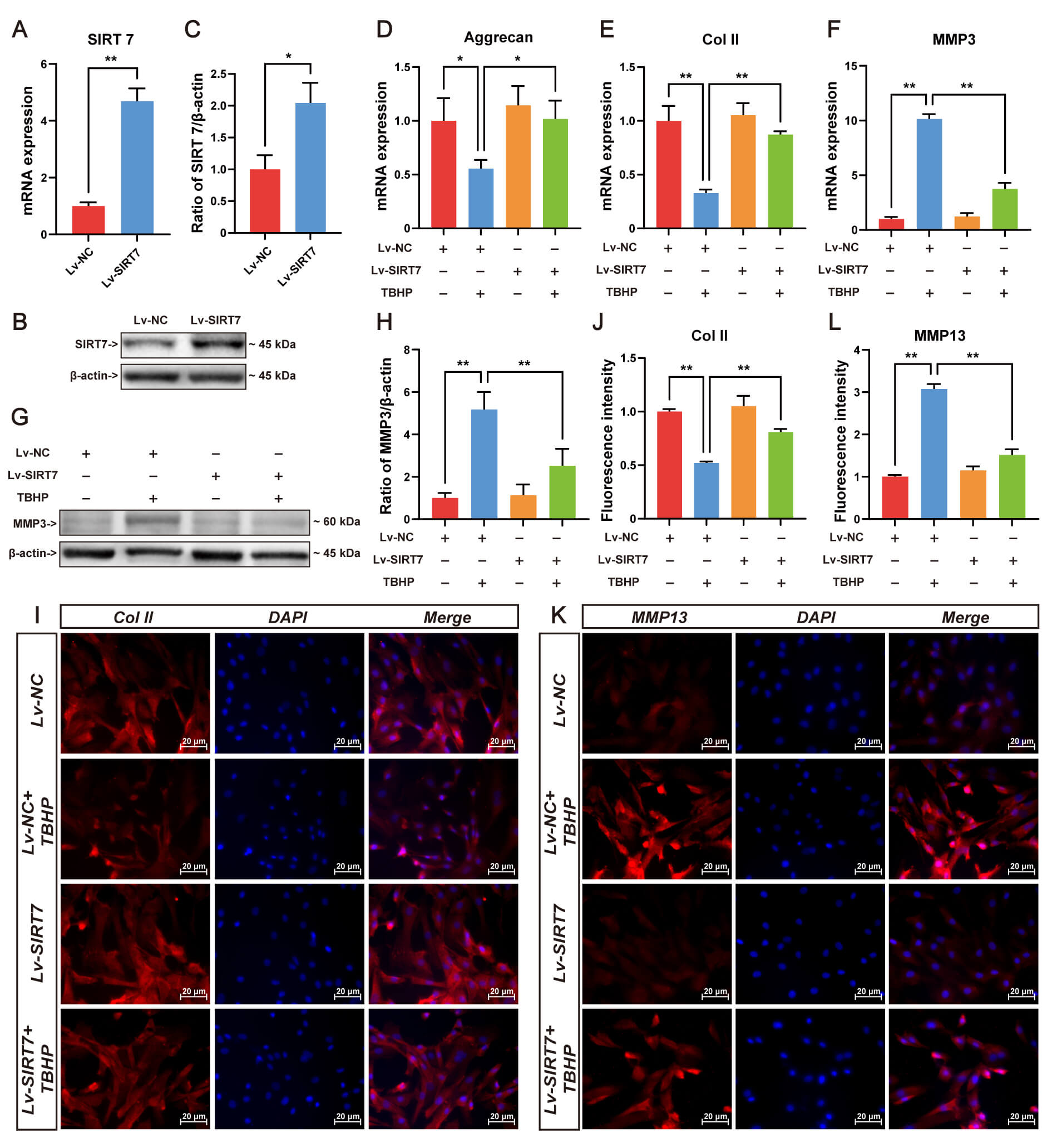 Fig. 3.
Fig. 3.
SIRT7 overexpression attenuates TBHP-induced ECM degradation in
NP cells. (A) SIRT7 transcript levels following lentiviral delivery evaluated by
qRT-PCR. (B) Representative Western blot showing SIRT7 protein expression in
transfected NP cells. (C) Quantification of SIRT7 protein levels normalized to
loading control. (D) Aggrecan mRNA quantification by qRT-PCR. (E) Col II gene
expression assessed through qRT-PCR. (F) MMP3 mRNA quantification by qRT-PCR. (G)
Representative Western blot of MMP3. (H) Relative MMP3 protein quantification by
Western blot. (I) Representative immunofluorescence staining images of Col II.
Scale bar: 20 µm. (J) Quantitative immunofluorescence analysis of
Col II in NP cells. (K) Representative immunofluorescence staining images of
MMP13. Scale bar: 20 µm. (L) Quantitative immunofluorescence
analysis of MMP13. Values represent mean
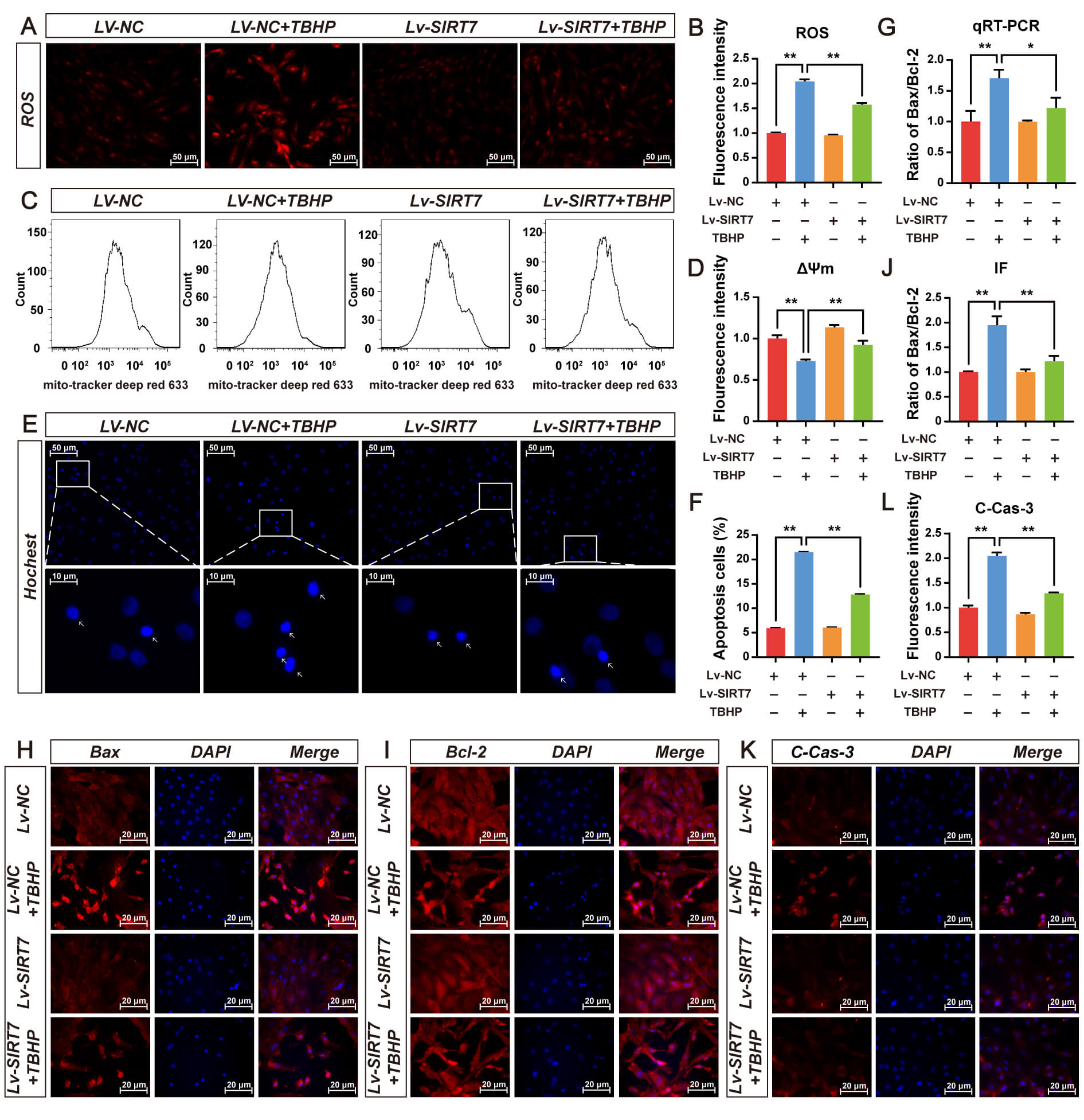
DHE staining showed that TBHP significantly increased intracellular ROS levels,
whereas SIRT7 overexpression effectively inhibited TBHP-induced increase in ROS
(Fig. 4A,B). Increased ROS triggers a decrease in
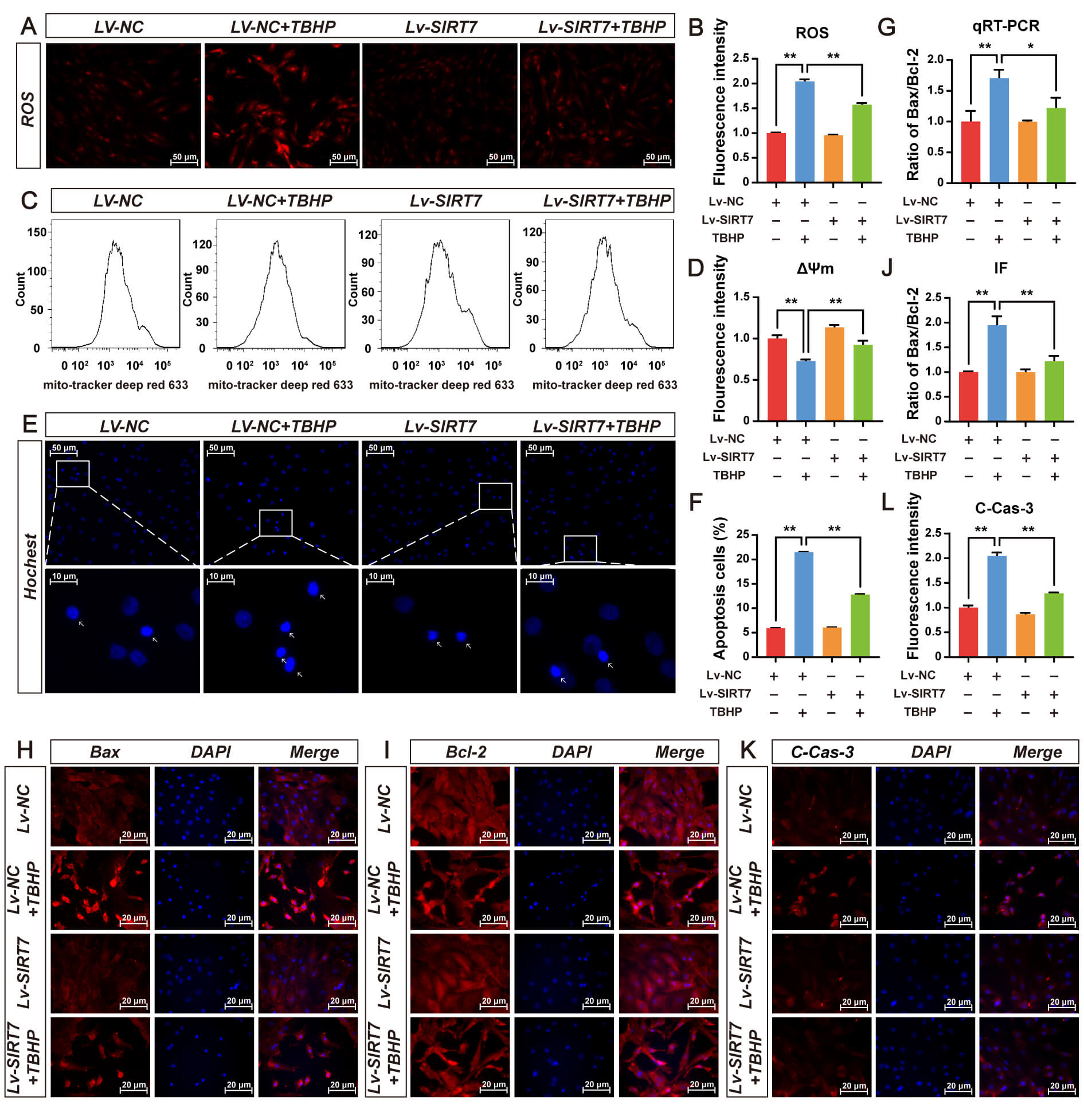 Fig. 4.
Fig. 4.
SIRT7 overexpression protects NP cells from oxidative stress and
apoptosis induced by TBHP. (A) Representative fluorescence micrographs of
intracellular ROS detected by DHE staining in Lv-NC and Lv-SIRT7 groups treated
with or without 75 µM TBHP for 48 h. Scale bar: 50 µm. (B)
Quantitative assessment of intracellular ROS level in (A). (C) Representative
flow cytometry histograms of
Accumulating evidence has established NF-
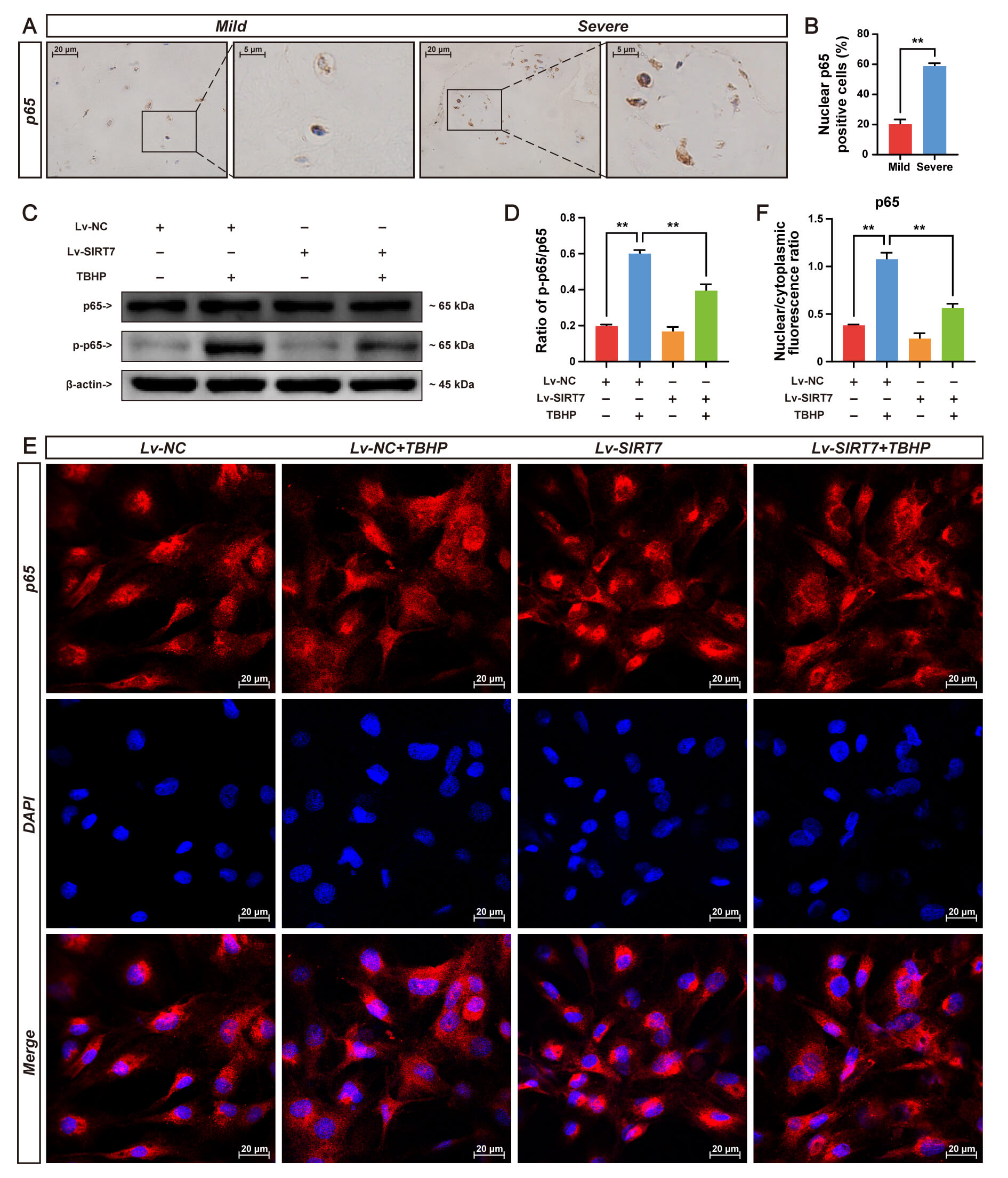 Fig. 5.
Fig. 5.
SIRT7 overexpression inhibits NF-
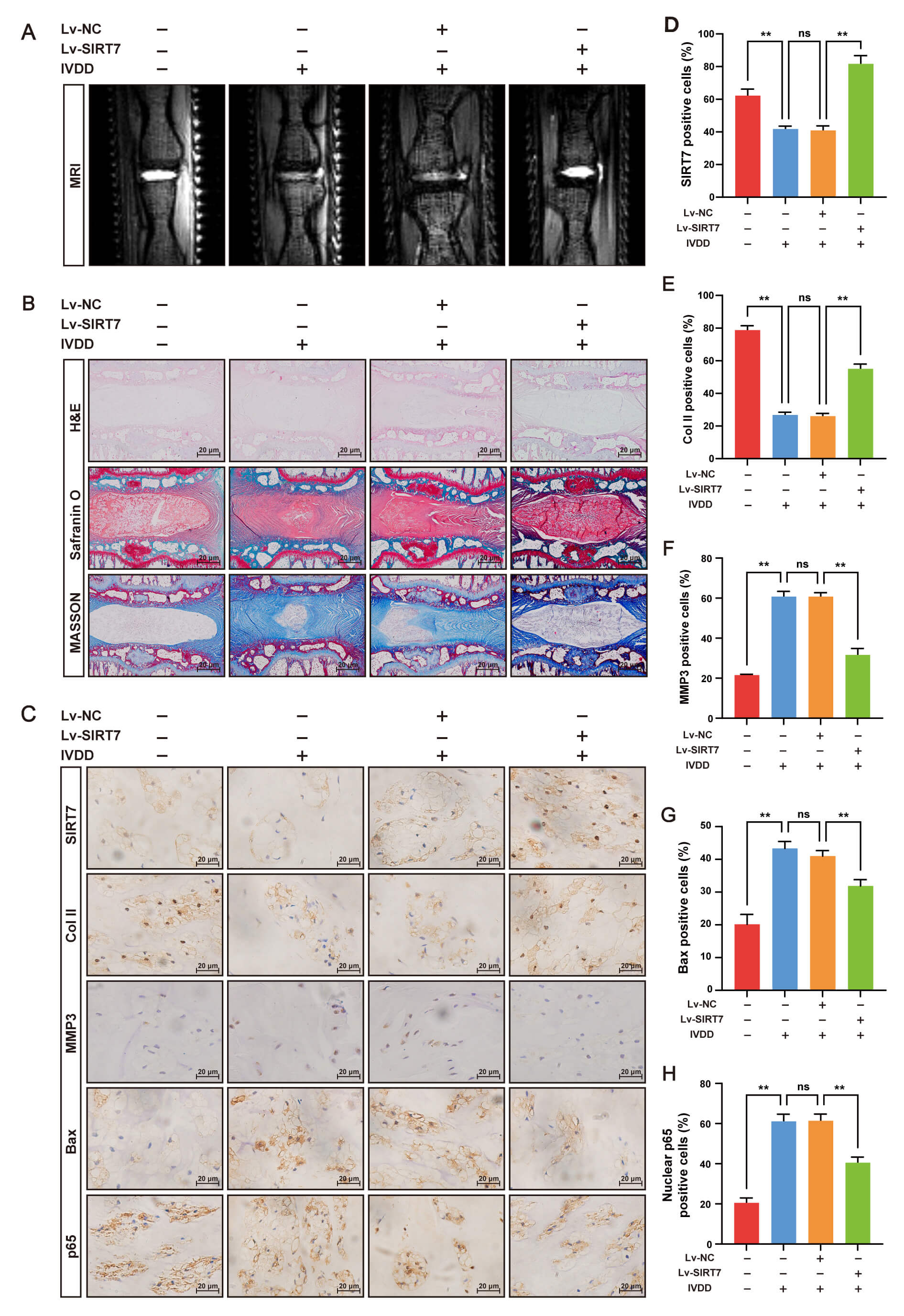
We established an IVDD rat model via needle puncture to evaluate the therapeutic
potential of SIRT7 in vivo. Twenty-eight Sprague–Dawley rats were
randomly assigned to sham, IVDD, IVDD + Lv-NC, or IVDD + Lv-SIRT7 groups. With
the exception of sham-operated animals, rats in the other groups were injected
with PBS, Lv-NC, or Lv-SIRT7, respectively. Eight weeks later, T2-weighted MRI
was performed, and the IVD was harvested for histological and IHC analyses. As
shown in Fig. 6, SIRT7 overexpression significantly attenuated IVDD progression.
IHC analysis revealed that SIRT7 expression was markedly upregulated in the NP
tissue of the IVDD + Lv-SIRT7 group compared with that in the other groups (Fig. 6C,D), confirming the successful overexpression of SIRT7. MRI showed that the
T2-weighted signal intensity was considerably preserved in the IVDD + Lv-SIRT7
group compared with that in the IVDD and IVDD + Lv-NC groups (Fig. 6A).
Consistent with this, histological analysis by H&E, Safranin O/fast green, and
Masson staining revealed severe NP collapse, proteoglycan loss, and fibrosis in
the IVDD and IVDD + Lv-NC groups; however, these degenerative changes were
significantly attenuated in the IVDD+Lv-SIRT7 group (Fig. 6B). Additionally, IHC
for collagen II, MMP3, and Bax showed that collagen II expression was
downregulated, whereas MMP3 and Bax expression were upregulated in the IVDD and
IVDD + Lv-NC groups, indicating increased ECM degradation and NP cell apoptosis.
SIRT7 overexpression reversed these pathological changes (Fig. 6C,E–G).
Importantly, nuclear p65 protein levels were significantly lower in the IVDD +
Lv-SIRT7 group than in the IVDD and IVDD + Lv-NC groups (Fig. 6C,H), indicating
that SIRT7 inhibits NF-
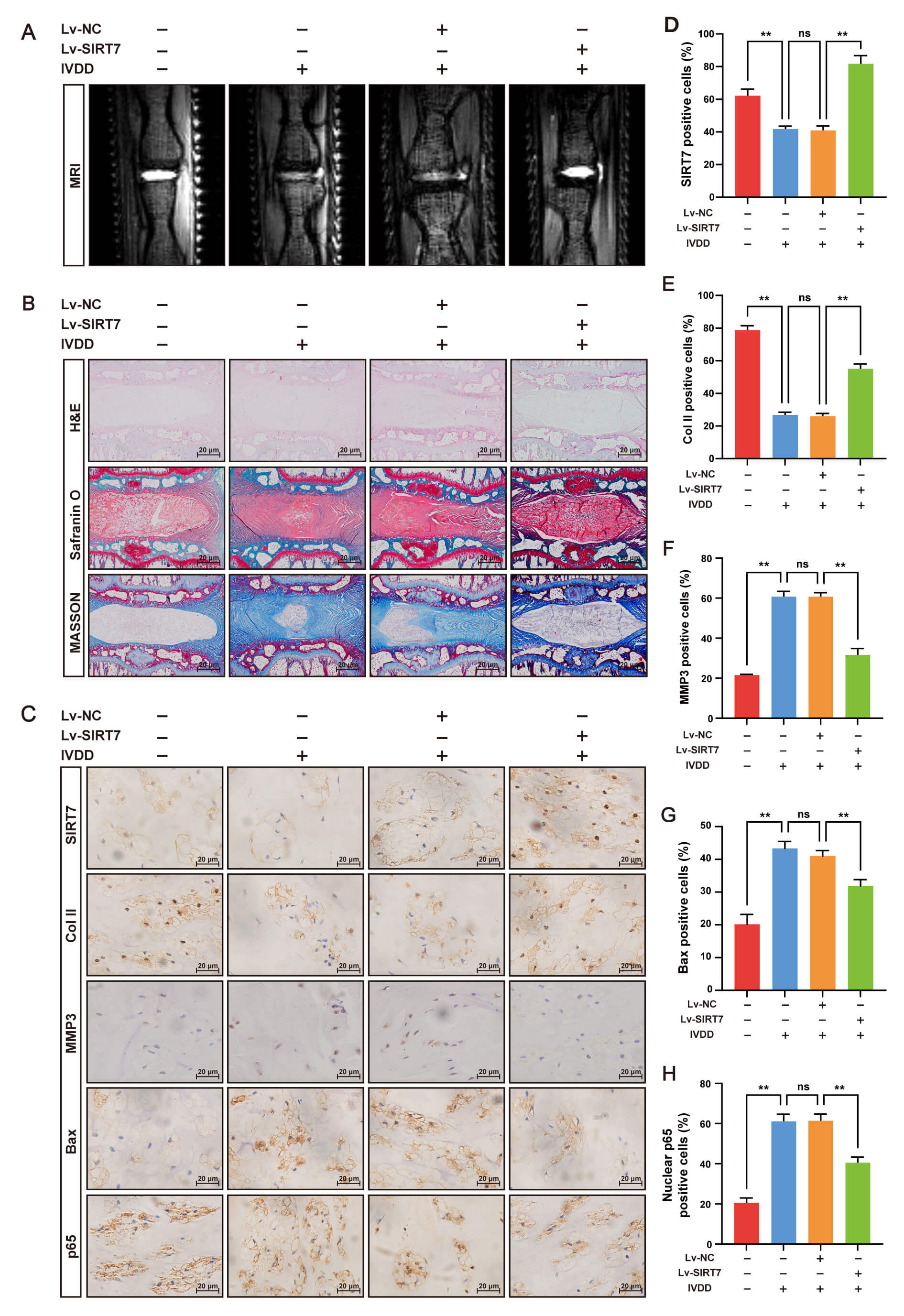 Fig. 6.
Fig. 6.
SIRT7 overexpression alleviates IVDD in rats by suppressing the
NF-
To the best of our knowledge, this study represents the first attempt to
characterize SIRT7 expression patterns in IVDD and to investigate its
cytoprotective potential in NP cells. Our data revealed a significant reduction
in SIRT7 expression within human degenerative NP specimens. Comparable outcomes
were observed in both the needle-puncture-induced rat IVDD model and TBHP-induced
NP cell degeneration model. Functional experiments showed that SIRT7 overexpression protected NP cells from apoptosis and ECM breakdown, and this protective effect correlated with NF-
IVDD constitutes the predominant pathological basis of chronic low back pain.
Despite incomplete mechanistic understanding, NP cell apoptosis and ECM
deterioration represent hallmark pathological processes in this disease. Thus,
suppression of cell apoptosis and restoration of matrix homeostasis constitute
fundamental therapeutic strategies for this disease [25, 26]. Oxidative stress,
driven primarily by ROS overproduction, critically impairs NP cell function [27].
Oxidative stress not only induces NP cell apoptosis but also compromises ECM
integrity, thereby accelerating IVDD. TBHP is an exogenous oxidant and
extensively employed for in vitro degeneration modeling [23]. It rapidly
elevates intracellular ROS levels and recapitulates apoptotic and catabolic
phenotypes observed in human pathological specimens. Based on our observation
that SIRT7 expression was decreased in degenerative NP tissues from both humans
and rats, as well as in TBHP-stimulated NP cells, we conducted functional rescue
experiments to evaluate the potential protective effects of SIRT7 overexpression
against TBHP-induced injury. We found that TBHP challenge induced robust ROS
generation,
NF-
The needle puncture-induced rat model of IVDD is extensively employed for
investigating disease pathogenesis and evaluating therapeutic interventions [28, 29],
owing to its technical simplicity, reproducibility, and cross-species
applicability [30]. We therefore assessed whether SIRT7 overexpression exerts
protective effects against IVDD in vivo. MRI revealed that
needle puncture markedly diminished IVD hydration, whereas SIRT7 overexpression
restored water content. Histological evaluation by H&E, Safranin O/Fast Green,
and Masson’s trichrome staining further confirmed that SIRT7 overexpression
attenuated IVDD. Notably, NF-
This study has several limitations. First, we did not use NF-
As far as we are aware, no prior investigation has implicated SIRT7 in the
preservation of NP homeostasis. The present study demonstrates that the protective effect of SIRT7 overexpression against IVDD progression correlates with inhibition of the NF-
SIRT7, sirtuin 7; IVD, intervertebral disc; IVDD, intervertebral disc degeneration; NP, nucleus pulposus; AF, annulus fibrosus; CEP, cartilaginous endplates; cDNA, complementary DNA; DHE, dihydroethidium; ECM, extracellular matrix; H&E, hematoxylin and eosin; IHC, immunohistochemistry; Lv-NC, negative control lentivirus; Lv-SIRT7, SIRT7-overexpressing lentivirus; MMP3, matrix metalloproteinase-3; MRI, magnetic resonance imaging; MMP13, matrix metalloproteinase-13; PCR, polymerase chain reaction; qRT-PCR, quantitative reverse transcription polymerase chain reaction; ROS, reactive oxygen species; SD, standard deviation; TBHP, tert-butyl hydroperoxide;
The data of this study are available upon reasonable request from corresponding author.
DW: Conceptualization, Data curation, Investigation, Methodology, Writing original draft. ZY: Data curation, Formal analysis, Methodology, Writing-review & editing. LL: Data curation, Investigation, Writing review & editing. ZC: Data curation, Formal analysis, Writing-original draft. GZ: Data curation, Formal analysis. TL: Data curation, Formal analysis. XK: Con ceptualization, Data curation, Funding acquisition, Methodology, Resources, Supervision, Writing-review & editing. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All the experimental protocols involved in this study were approved by the Ethics Committee of Lanzhou University Second Hospital (2025A-1331). Human-related procedures followed the principles of the Helsinki Declaration, and written informed consent was obtained from every patient before tissue collection. All animal experiments adhered to the ARRIVE guidelines and were reviewed and approved by the Animal Care and Use Committee at Lanzhou University Second Hospital (No. D2025-898).
Not applicable.
This research was funded by the National Natural Science Foundation of China, Grant No.: 82272536, 82560433; the Natural Science Foundation of Gansu province, Grant No.: 25JRRA626; the Cuiying Scientific and Technological Innovation Program of the Second Hospital & Clinical Medical School, Lanzhou University, Grant No.: CY2024-YB-A02; Major Scientific Research Project for Technological Innovation in the Health and Healthcare Sector of Gansu Province, Grant no.: GSWSQNPY2025-02.
The authors declare no conflicts of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBL50425.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.