
 , Jiayao Zhou 1,2,†, Shiyao Zhang 1,2, Weitian Zhang 1,2, Zhipeng Li 1,2,*
, Jiayao Zhou 1,2,†, Shiyao Zhang 1,2, Weitian Zhang 1,2, Zhipeng Li 1,2,*
1 Department of Otolaryngology-Head and Neck Surgery, Shanghai Sixth People’s Hospital Affiliated to Shanghai Jiao Tong University School of Medicine, 200233 Shanghai, China
2 Otolaryngological Institute, Shanghai Jiao Tong University, 200233 Shanghai, China
†These authors contributed equally.
Abstract
Chronic rhinosinusitis with nasal polyps (CRSwNP) is characterized by persistent tissue remodeling, but the mechanisms underlying impaired fibrinolysis were not fully understood. SERPIN family B member 2 (SERPINB2/plasminogen activator inhibitor type 2 [PAI-2]) is a known inhibitor of tissue plasminogen activator (tPA). However, its role in CRSwNP pathogenesis remains unclear. This study investigated whether SERPINB2 contributes to fibrinolytic dysfunction in CRSwNP.
tPA and SERPINB2 expression levels were assessed in nasal polyp and control turbinate tissues using qRT-PCR, Western blot, and immunofluorescence. Primary human nasal epithelial cells were stimulated with IFN-γ, IL-13, or IL-17A to evaluate cytokine-mediated regulation. The direct effects of SERPINB2 on tPA expression were examined using recombinant protein treatment and siRNA-mediated knockdown. tPA enzymatic activity and fibrinolytic function were measured using a fluorometric substrate assay and D-dimer ELISA, respectively.
tPA expression was significantly reduced in nasal polyp tissues compared to control turbinate and inversely correlated with SERPINB2 levels. Immunofluorescence analysis revealed decreased tPA-positive and increased SERPINB2-positive cells in the nasal epithelium. Both the Th1 cytokine IFN-γ and the Th2 cytokine IL-13 downregulated tPA while upregulating SERPINB2 in primary nasal epithelial cells, whereas IL-17A showed no significant effect. Notably, recombinant SERPINB2 dose-dependently suppressed epithelial tPA expression, while SERPINB2 knockdown rescued cytokine-induced tPA downregulation. Functionally, SERPINB2 inhibited tPA enzymatic activity in a dose-dependent manner and significantly impaired fibrinolytic function.
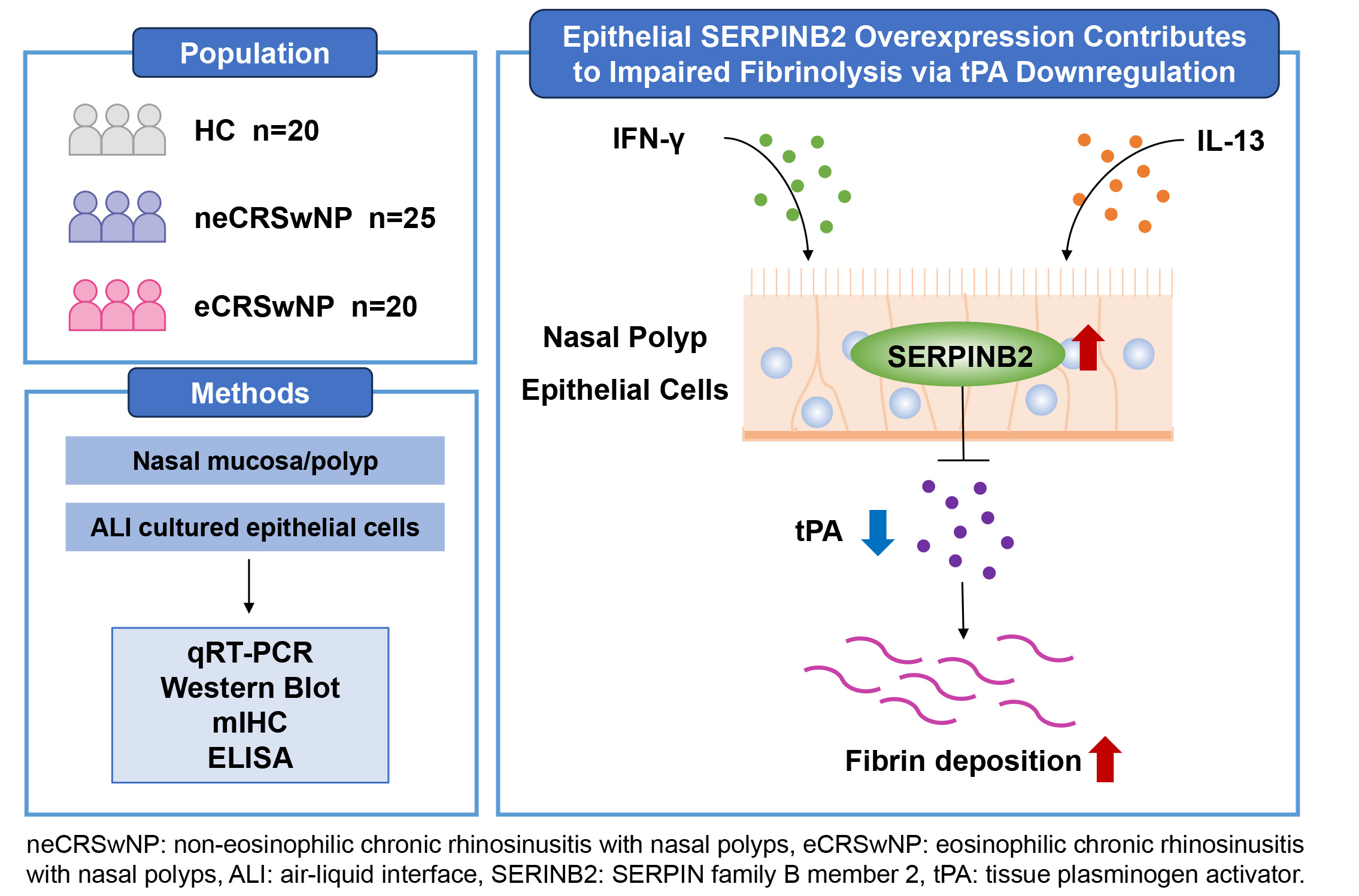
This study identifies a novel SERPINB2-tPA regulatory axis in nasal epithelial cells. The convergent regulation by both Th1 and Th2 cytokines suggests that fibrinolytic dysfunction occurs across different CRSwNP inflammatory endotypes. These findings provide mechanistic insights into fibrin accumulation in nasal polyps and identify SERPINB2 as a potential therapeutic target for the prevention of polyp formation and recurrence.
Graphical Abstract
Keywords
- rhinosinusitis
- nasal polyps
- plasminogen activator inhibitor 2
- tissue plasminogen activator
- fibrinolysis
Chronic rhinosinusitis with nasal polyps (CRSwNP) is an inflammatory condition characterized by outgrowth of sinonasal tissue [1]. According to the degree of eosinophilic infiltration, CRSwNP can be further divided into eosinophilic CRSwNP (eCRSwNP) and non-eosinophilic CRSwNP (neCRSwNP) [2]. The eCRSwNP phenotype is characterized by predominant Th2 immune responses, while neCRSwNP displays a heterogeneous inflammatory profile involving Th1, Th2, and Th17 pathways [3]. Regardless of these distinct immunological patterns, both subtypes demonstrate substantial subepithelial fibrin accumulation [4]. Fibrin degradation is facilitated by plasmin, which is generated from plasminogen upon cleavage by either urokinase plasminogen activator (uPA) or tissue plasminogen activator (tPA) [5]. Previous reports have shown significant downregulation of tPA, but not uPA, in both eCRSwNP and neCRSwNP patients [6]. Exposure to both Th1 and Th2 inflammatory mediators has been shown to downregulate tPA in cultured nasal epithelial cells [7]. However, the mechanisms underlying their regulation in CRSwNP remain poorly understood. Elucidating the mechanisms driving fibrin accumulation in CRSwNP could lead to improved therapeutic strategies.
SERPIN family B member 2 (SERPINB2), also known as plasminogen activator inhibitor type 2 (PAI-2), is a coagulation factor that inactivates uPA and tPA [8]. Previous studies have shown that SERPINB2 is dramatically elevated in bronchial epithelial cells of asthmatic patients and is positively associated with pivotal clinical markers, such as FEV1, FeNO, eosinophil counts, and asthma severity [9]. Single-cell RNA sequencing analysis has identified SERPINB2 as one of the core upregulated genes in human bronchial epithelial cells after segmental allergen challenge [10]. We have previously shown that SERPINB2 upregulated by IL-13 promotes the expression of 15-lipoxygenase-1 (15LO1), CCL26 and iNOS through activation of the STAT6 pathway in nasal polyp epithelial cells [11]. We also reported that elevated SERPINB2 in allergic rhinitis patients regulates MUC5AC expression via STAT6 signaling in nasal epithelial cells [12]. Based on these findings, we hypothesize that elevated SERPINB2 in nasal epithelial cells may suppress the expression and activity of tPA, thereby contributing to fibrin deposition in nasal polyps.
To test this hypothesis, SERPINB2 and tPA expression were analyzed in nasal polyp (NP) tissues from CRSwNP patients and turbinate tissues from healthy controls. Their colocalization was examined by immunofluorescence. Using air-liquid interface (ALI) cultures of primary nasal epithelial cells, the regulatory effects of SERPINB2 on tPA were examined through cytokine stimulation, recombinant protein treatment, and RNA interference. tPA enzymatic activity and fibrinolytic function were measured by fluorometric assays and D-dimer ELISA. Our results demonstrated that increased SERPINB2 in nasal polyp epithelial cells suppresses tPA expression and reduces tPA activity in CRSwNP.
The Ethical Committee of Shanghai Sixth People’s Hospital Affiliated to
Shanghai Jiao Tong University School of Medicine granted approval for this
investigation, and all subjects provided written informed consent prior to
participation. The study cohort comprised 20 eCRSwNP patients, 25 neCRSwNP
patients, and 20 healthy controls (HCs) (Supplementary Table 1). CRSwNP classification adhered to the EPOS2020 criteria. Eosinophilic
CRSwNP was defined as tissue eosinophil counts of
Multiplex immunohistochemistry (mIHC) was performed using the TSA Fluorescence
Staining Kit on formalin-fixed paraffin-embedded tissue sections. Briefly, tissue
sections underwent deparaffinization, antigen retrieval, and sequential antibody
staining. Primary antibodies against tPA and SERPINB2 were sequentially applied,
followed by HRP-conjugated secondary antibodies and TSA-fluorophore
amplification. Microwave treatment was performed between each cycle to strip
antibodies. Nuclei were counterstained with DAPI. Confocal images were acquired
using Zeiss LSM 780 confocal laser scanning microscope. SERPINB2+ and
tPA+ cells within the epithelial compartment were manually counted in 5
randomly selected fields (200
Turbinate epithelial scrapings (IT/MT) from control subjects served as the
source for primary NEC isolation. All primary NECs were isolated and cultured
following established protocols. Cell identity was confirmed by characteristic
epithelial morphology under microscopy and tested negative for mycoplasma.
Cultures were maintained under ALI conditions following established protocols.
Inflammatory stimuli (IFN-
Gene expression was quantified using SYBR Green-based qRT-PCR on a Roche
LightCycler 480 II system.
Fresh tissue samples and cell lysates underwent 10% SDS-PAGE separation followed by immunoblotting with antibodies against tPA, SERPINB2, and GAPDH. Band intensities were quantified using ImageJ software (version 1.54j; National Institutes of Health, Bethesda, MD, USA). Detailed procedures are provided in the Supplementary Material.
tPA protein levels in culture media were measured with quantitative sandwich ELISA kits (DTPA00, R&D Systems, Minneapolis, MN, USA) according to the manufacturer’s instructions.
tPA enzymatic activity was measured fluorometrically using the synthetic substrate Z-GGR-AMC (carbobenzoxy-Gly-Gly-Arg-7-amino-4-methylcoumarin). Recombinant human tPA (5 nM) was pre-incubated with increasing concentrations of recombinant SERPINB2 (0, 6.25, 25, or 100 nM) for 30 min at 37 °C to allow SERPINB2-tPA complex formation. Following pre-incubation, Z-GGR-AMC substrate was added to a final concentration of 100 µM. Fluorescence (Ex/Em 360/460 nm) was continuously monitored for 60 min at 37 °C using a microplate reader. The rate of AMC release, reflecting tPA proteolytic activity, was quantified as relative fluorescence units (RFU) over time.
To assess the functional impact of SERPINB2 on fibrinolysis, human serum was treated with recombinant tPA (50 nM), recombinant SERPINB2 (0, 6.25, 25, or 100 nM), or both and incubated at 37 °C for 300 s. The higher tPA concentration (50 nM vs. 5 nM in the enzymatic assay) was used to overcome endogenous serum protease inhibitors and ensure measurable fibrinolysis. D-dimer levels, a specific marker of fibrin degradation, were quantified by ELISA according to the manufacturer’s instructions.
Statistical analyses and figure generation were performed using GraphPad Prism
software (version 8.0; GraphPad Software, San Diego, CA, USA). Parametric data
are presented as mean
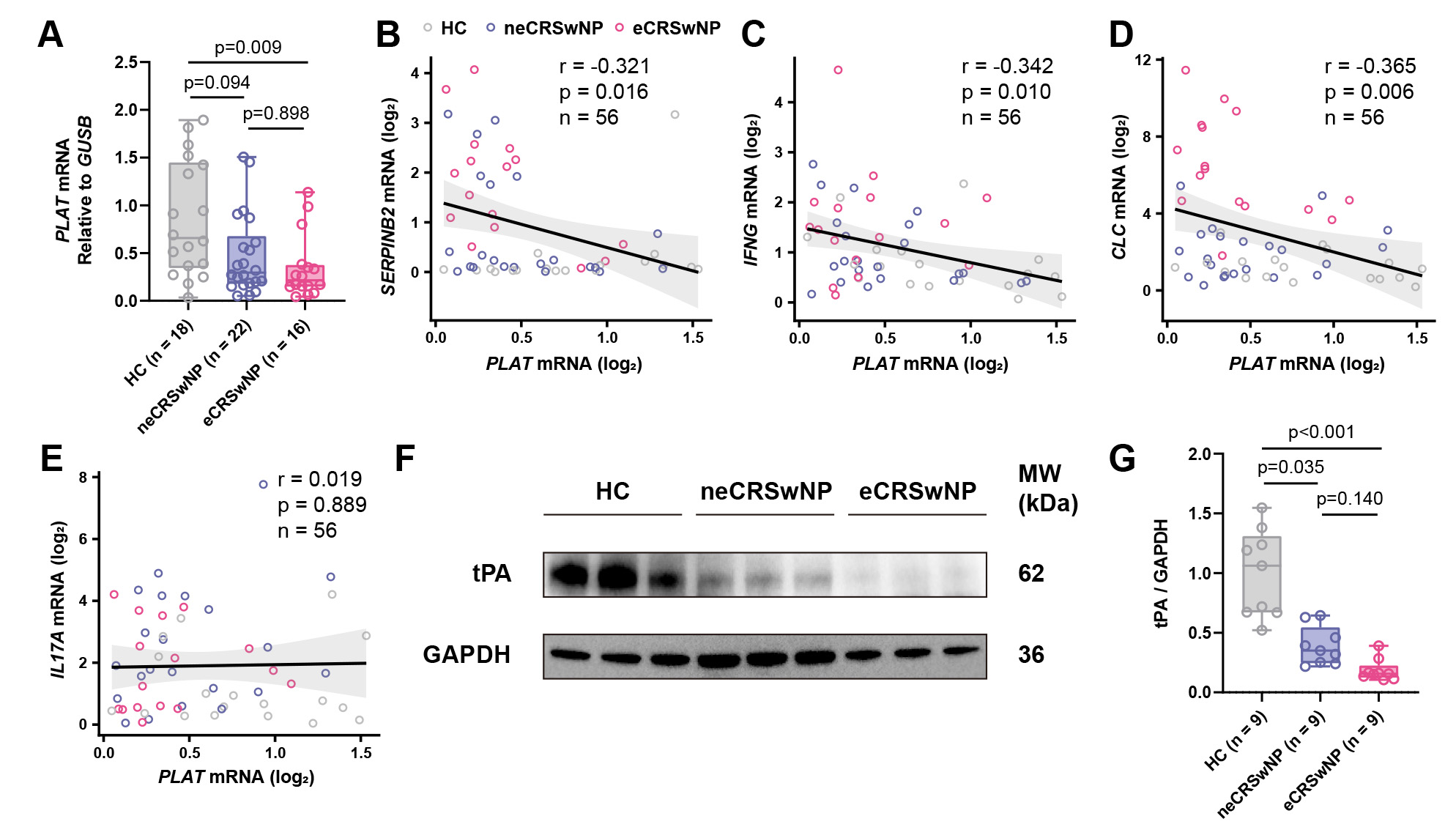
To investigate tPA expression in CRSwNP, we first examined PLAT mRNA levels in nasal polyp tissues from patients with eCRSwNP (n = 16) and neCRSwNP (n = 22), compared to HCs (n = 18). qRT-PCR analysis showed that PLAT mRNA levels were significantly reduced in eCRSwNP compared to control subjects, while neCRSwNP exhibited a trend toward reduction that did not reach statistical significance (Fig. 1A). Subgroup analysis showed no significant difference in PLAT expression between asthmatic and non-asthmatic patients within the eCRSwNP group (Supplementary Fig. 1A), or between non-asthmatic eCRSwNP and neCRSwNP patients (Supplementary Fig. 1B), suggesting that tPA downregulation in CRSwNP is not confounded by asthma comorbidity. Correlation analysis revealed inverse correlations between PLAT mRNA and SERPINB2 (Fig. 1B), IFNG (Fig. 1C), and CLC (Fig. 1D) expression in nasal polyp tissues, while no significant correlation was observed between PLAT and IL17A mRNA levels (Fig. 1E), suggesting that tPA downregulation is associate with both Th1 and Th2, but not Th17, inflammatory mediators. At the protein level, WB analysis confirmed significant downregulation of tPA in both eCRSwNP and neCRSwNP tissues compared to control subjects (Fig. 1F,G).
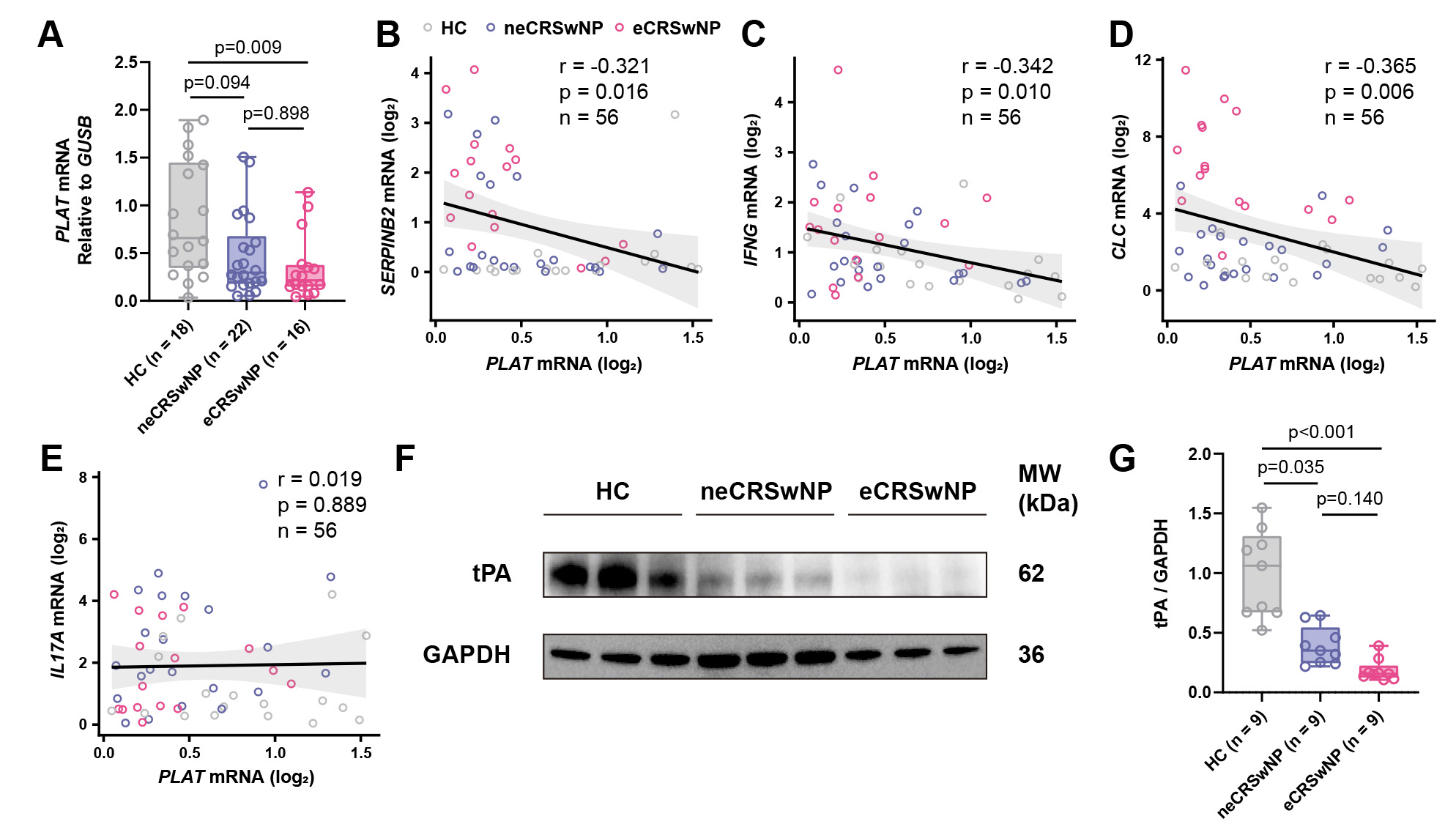 Fig. 1.
Fig. 1.
tPA expression is reduced in CRSwNP tissues and inversely correlates with SERPINB2. (A) RT-qPCR analysis of PLAT mRNA expression in inferior turbinate controls (n = 18), neCRSwNP (n = 22), and eCRSwNP (n = 16) tissues. (B–E) Spearman correlation analyses between PLAT mRNA levels and (B) SERPINB2, (C) IFNG, (D) CLC or (E) IL17A mRNA expression in all tissue samples (n = 56). (F) Representative Western blot images showing tPA protein levels in control, neCRSwNP, and eCRSwNP tissues. (G) Densitometric quantification of tPA protein levels normalized to GAPDH. HC, healthy control; neCRSwNP, non-eosinophilic CRSwNP; eCRSwNP, eosinophilic CRSwNP; SERPINB2, SERPIN family B member 2; tPA, tissue plasminogen activator; CRSwNP, chronic rhinosinusitis with nasal polyps.
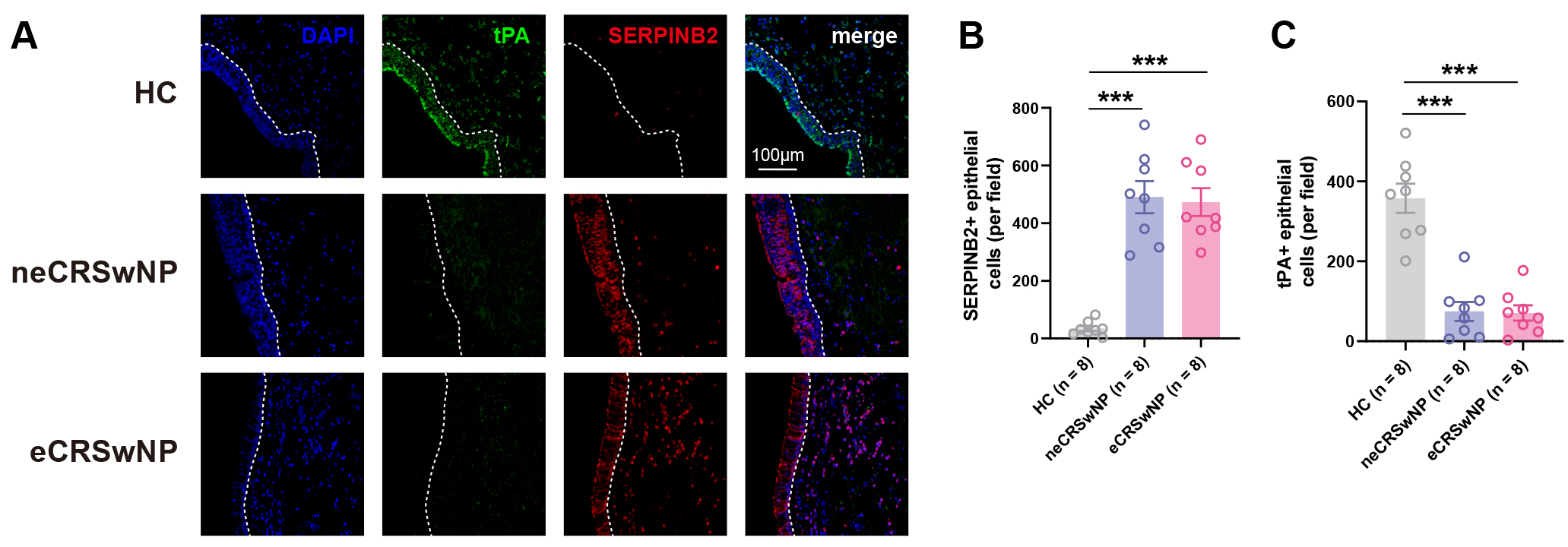
To further characterize tPA and SERPINB2 expression at the cellular level, we performed mIHC on nasal tissue sections. Consistent with the mRNA findings described above, mIHC analysis revealed a significant increase in SERPINB2+ epithelial cells in both eCRSwNP and neCRSwNP tissues compared to control subjects (Fig. 2A,B). Conversely, tPA+ epithelial cells were markedly reduced in both polyp endotypes relative to control subjects (Fig. 2A,C). No significant differences in either tPA+ or SERPINB2+ epithelial cell counts were observed between eCRSwNP and neCRSwNP groups (Fig. 2B,C). These findings confirm reciprocal changes in tPA and SERPINB2 expression within the nasal polyp epithelium, corroborating the inverse correlation observed at the mRNA level.
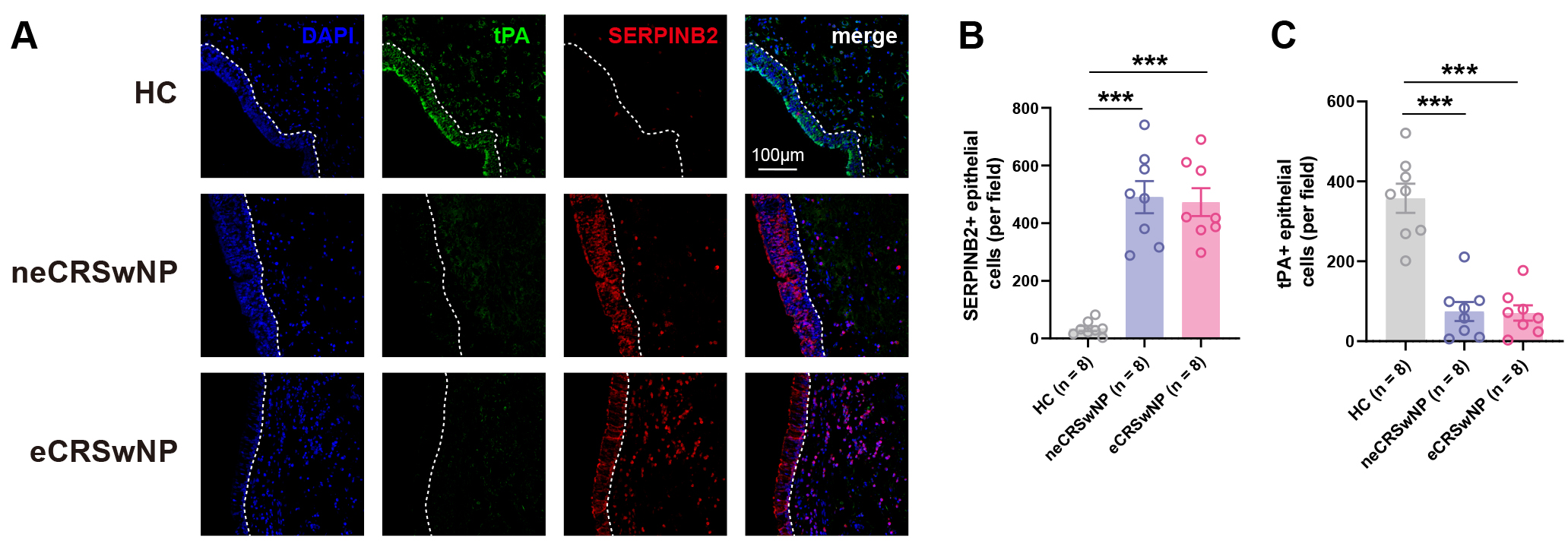 Fig. 2.
Fig. 2.
Multiplex immunohistochemistry confirms reciprocal tPA and
SERPINB2 expression in nasal polyp epithelium. (A) Representative multiplex
immunohistochemistry (mIHC) images showing tPA (green), SERPINB2 (red), and DAPI
(blue, nuclei) staining in healthy controls, neCRSwNP, and eCRSwNP tissues. Scale
bars: 100 µm. (B) Quantification of SERPINB2+ cells within the
epithelial compartment per field (200
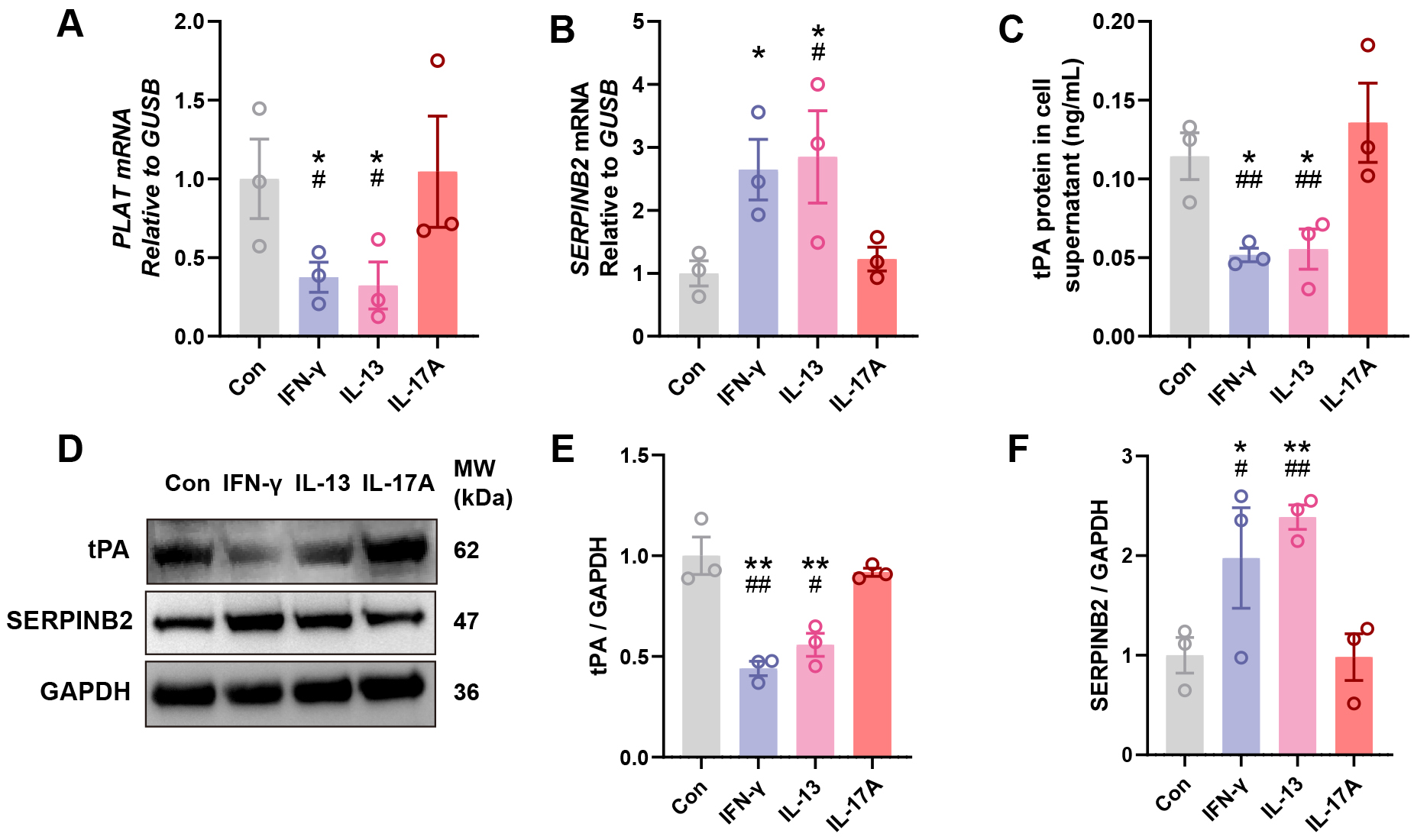
To investigate whether key inflammatory cytokines regulate tPA and SERPINB2
expression in nasal epithelium, we isolated and cultured primary human NECs from
healthy donors and stimulated them with Th1 (IFN-
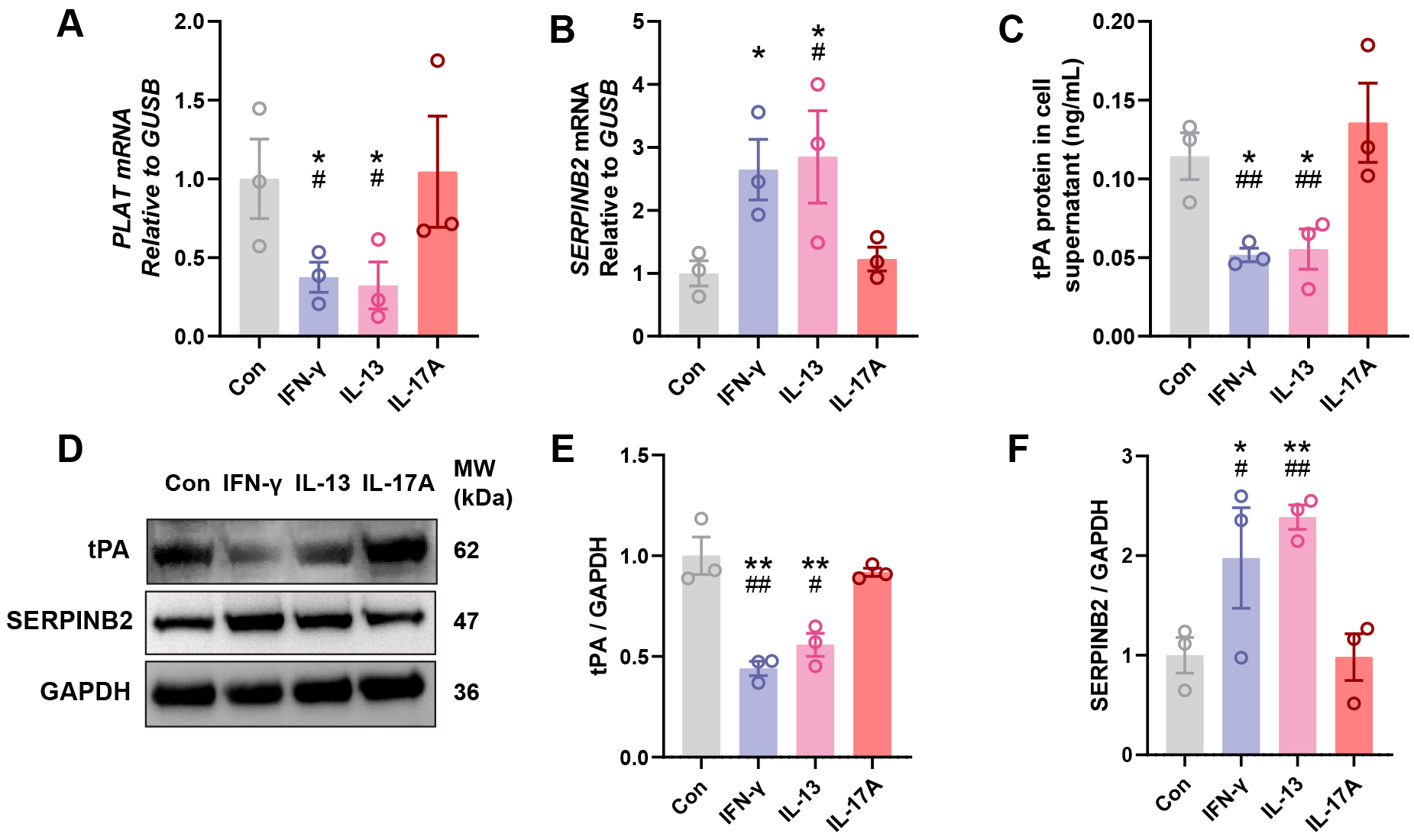 Fig. 3.
Fig. 3.
IFN-
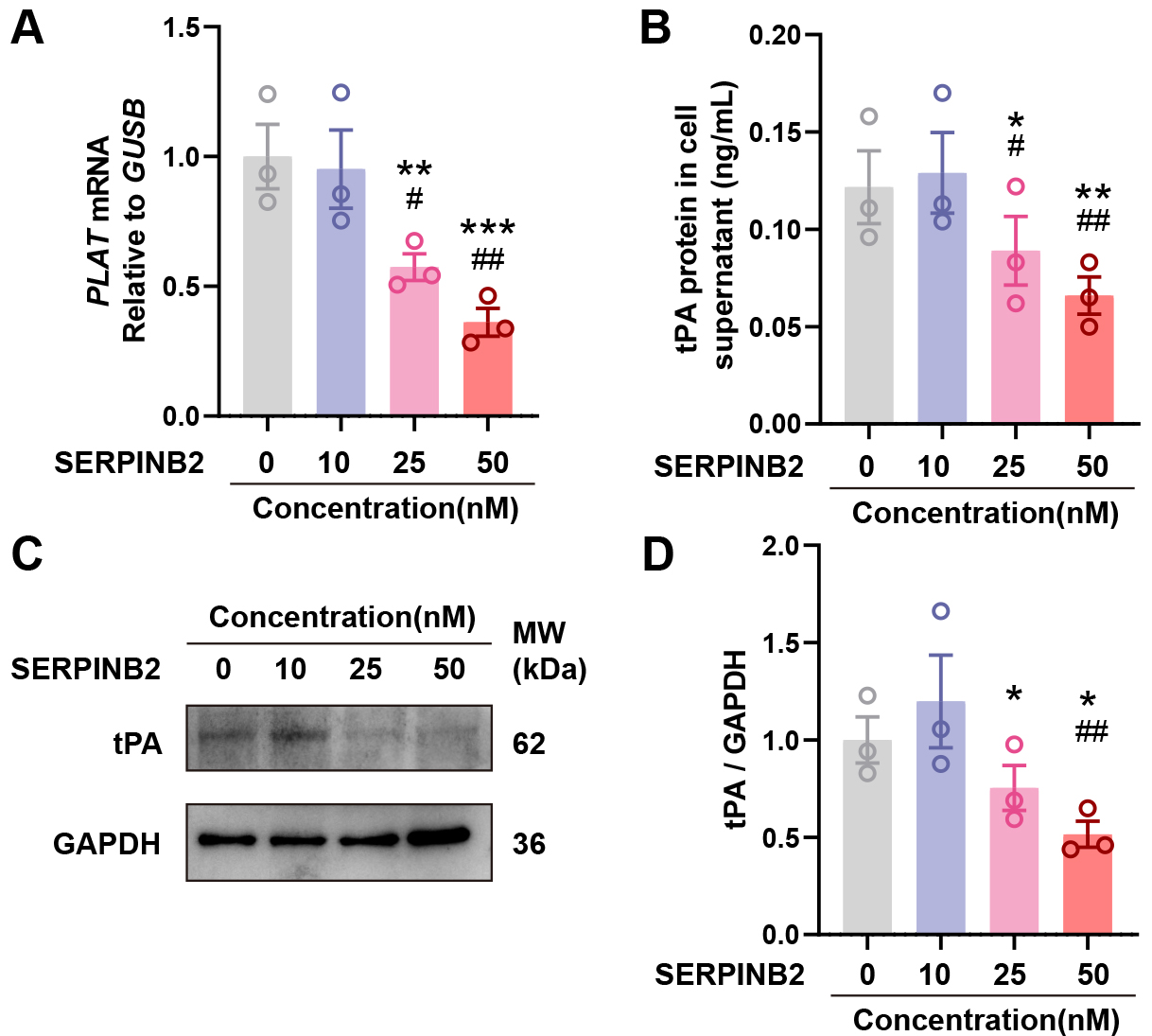
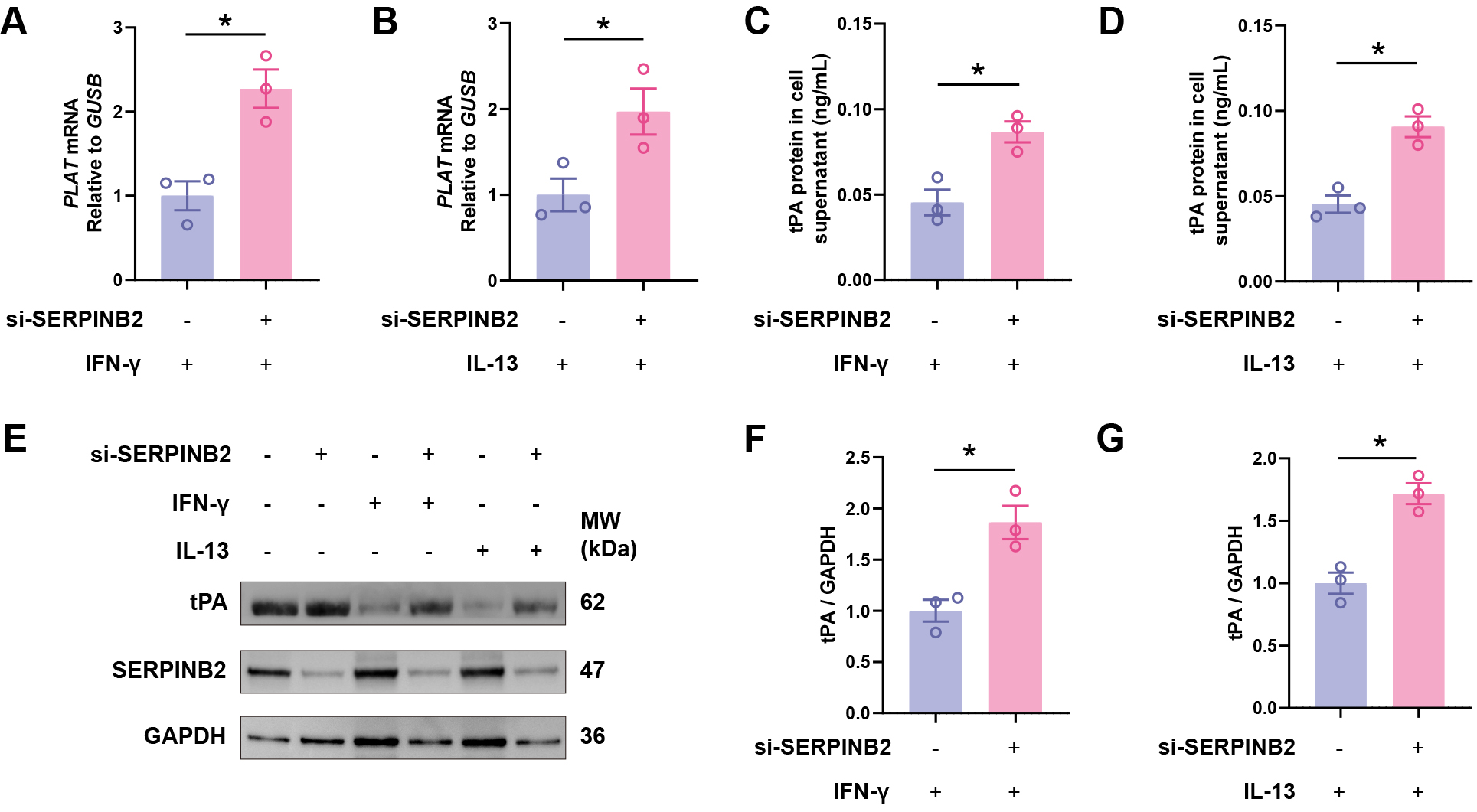
To investigate whether SERPINB2 directly regulates tPA expression, we treated primary NECs with recombinant human SERPINB2 protein (0, 10, 25, or 50 nM) for 48 hours. qRT-PCR showed dose-dependent suppression of PLAT mRNA with increasing SERPINB2 concentrations (Fig. 4A). ELISA revealed corresponding reductions in secreted tPA protein (Fig. 4B). Western blot confirmed decreased intracellular tPA protein in cell lysates (Fig. 4C,D). These results demonstrate that exogenous SERPINB2 dose-dependently suppresses tPA expression, suggesting a potential contribution to impaired fibrinolysis in CRSwNP.
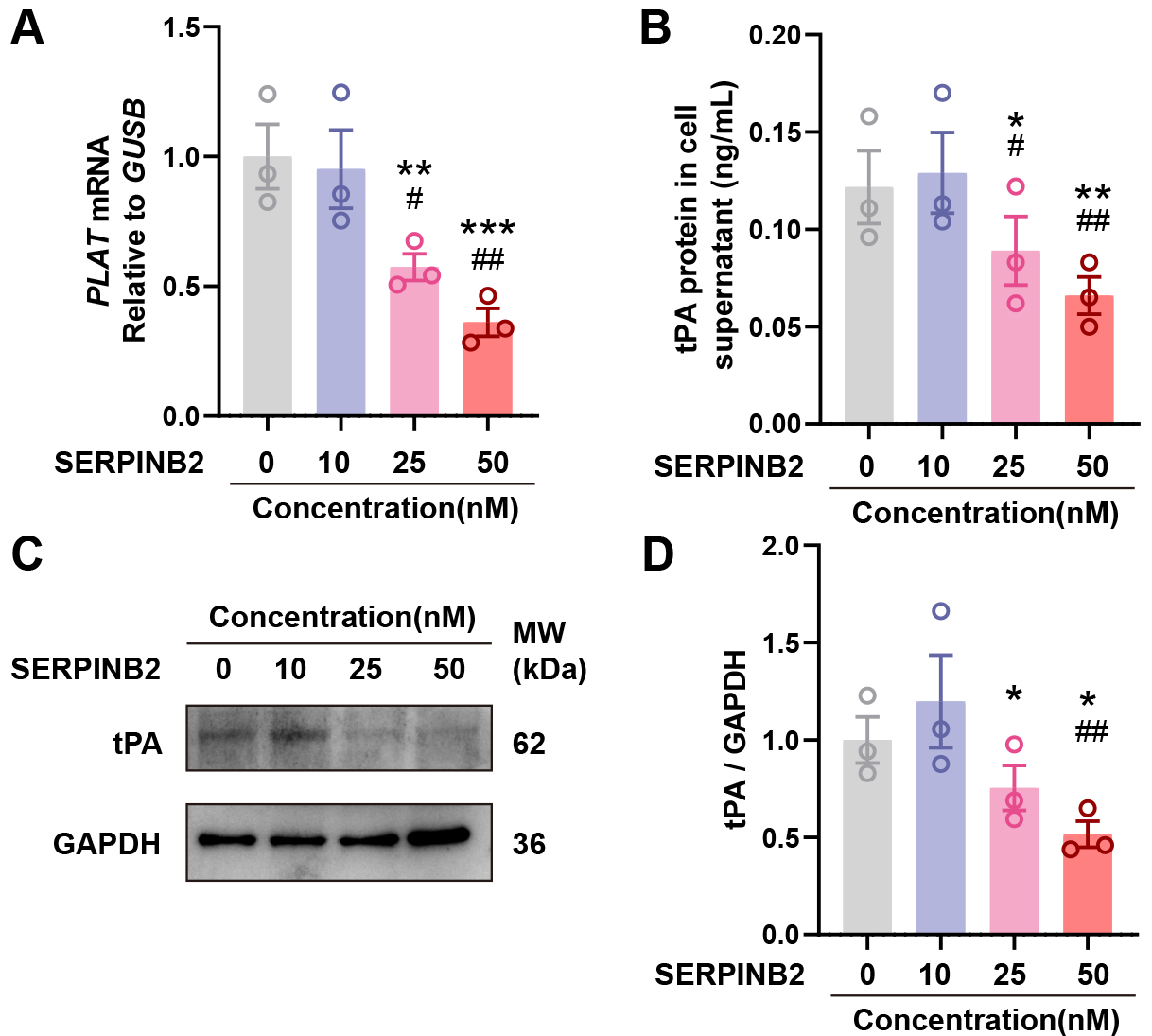 Fig. 4.
Fig. 4.
SERPINB2 dose-dependently suppresses tPA expression in primary
nasal epithelial cells. (A) RT-qPCR analysis of PLAT mRNA expression
following SERPINB2 treatment. (B) ELISA quantification of secreted tPA protein in
culture supernatants following SERPINB2 treatment. (C) Representative Western
blot images showing tPA protein levels in cell lysates from vehicle- or
SERPINB2-treated nasal epithelial cells. (D) Densitometric quantification of tPA
protein levels normalized to GAPDH. Data are presented as mean
To test whether SERPINB2 mediates cytokine-induced tPA suppression, we performed
siRNA-mediated SERPINB2 knockdown in primary nasal epithelial cells. Knockdown
efficiency was confirmed by Western blot analysis in three independent ALI
culture samples (Supplementary Fig. 2). Critically, SERPINB2 knockdown
rescued IFN-
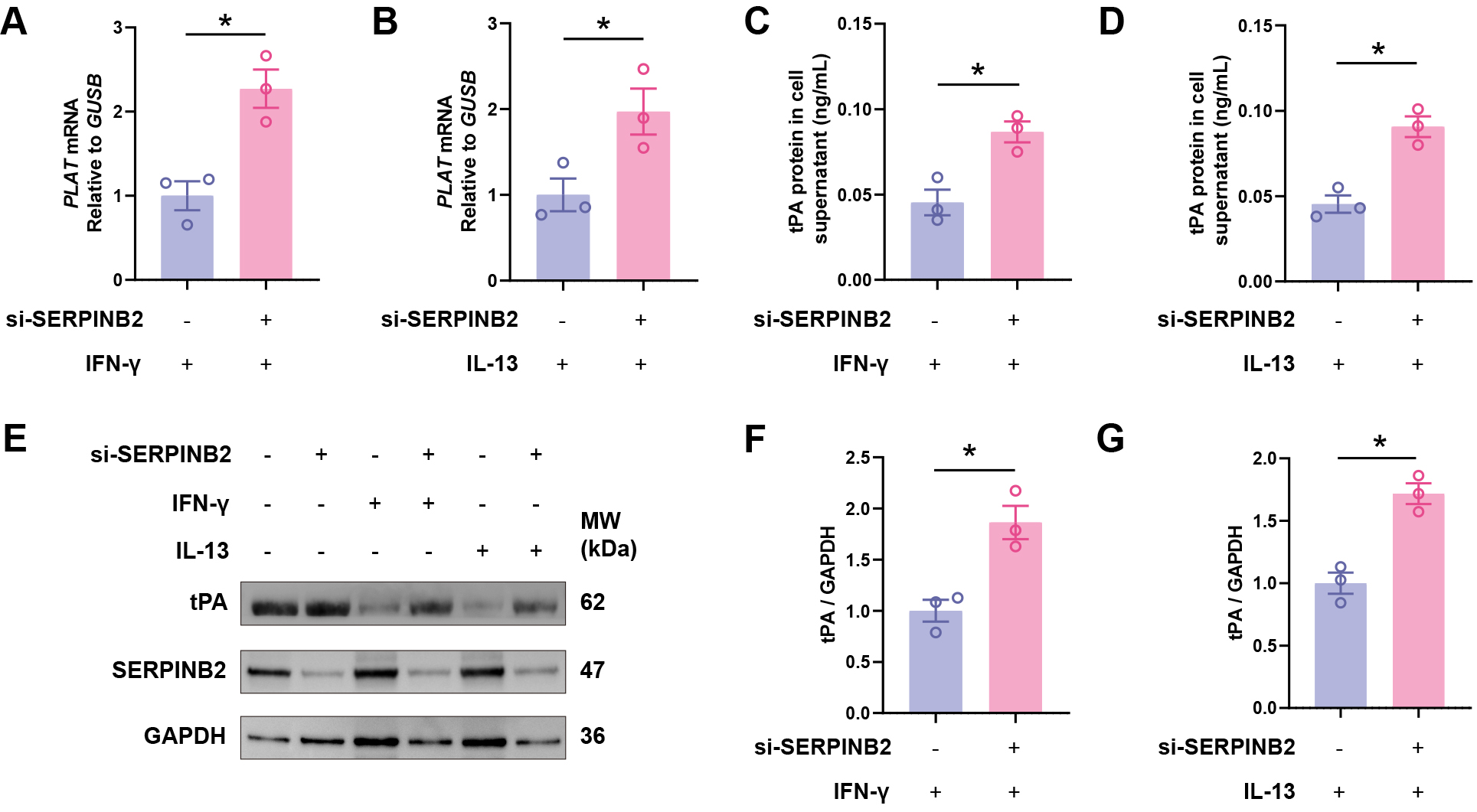 Fig. 5.
Fig. 5.
SERPINB2 knockdown rescues Th1 and Th2 cytokine-induced tPA
downregulation. (A,B) RT-qPCR analysis of PLAT mRNA expression in
IFN-
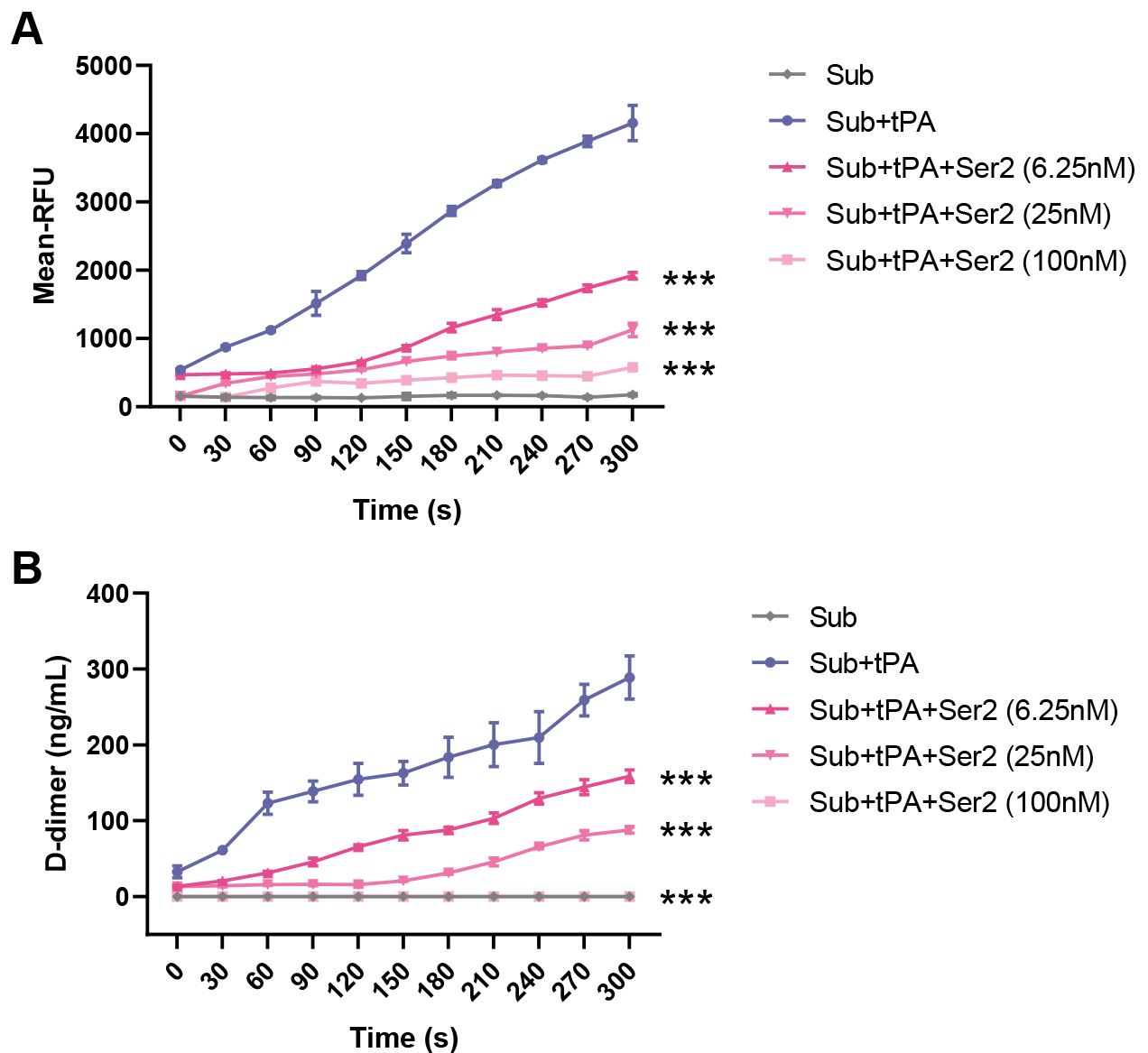
To validate the inhibitory effect of SERPINB2 on tPA activity and fibrinolytic function, we performed in vitro enzymatic activity and fibrinolysis assays. Using a fluorogenic substrate assay (Fig. 6A), we observed that tPA alone (Sub+tPA) produced robust enzymatic activity, with fluorescence signals reaching approximately 4000 RFU at 300 seconds. Co-incubation with SERPINB2 dose-dependently inhibited tPA activity. At concentrations of 6.25 nM, 25 nM, and 100 nM, SERPINB2 reduced fluorescence signals to approximately 1900, 1100, and 600 RFU, respectively, corresponding to inhibition rates of 55%, 74%, and 86%, respectively. Consistent with these findings, a D-dimer release assay (Fig. 6B) demonstrated that SERPINB2 significantly impaired fibrinolytic function. While tPA alone generated approximately 290 ng/mL D-dimer at 300 seconds, SERPINB2 at 6.25 nM, 25 nM, and 100 nM reduced D-dimer levels to approximately 160, 90, and 0 ng/mL, respectively, representing inhibition rates of 45–69%. These results demonstrate that SERPINB2 directly inhibits tPA enzymatic activity in a dose-dependent manner, thereby impairing fibrinolytic function and potentially contributing to edema formation and fibrin accumulation in CRSwNP.
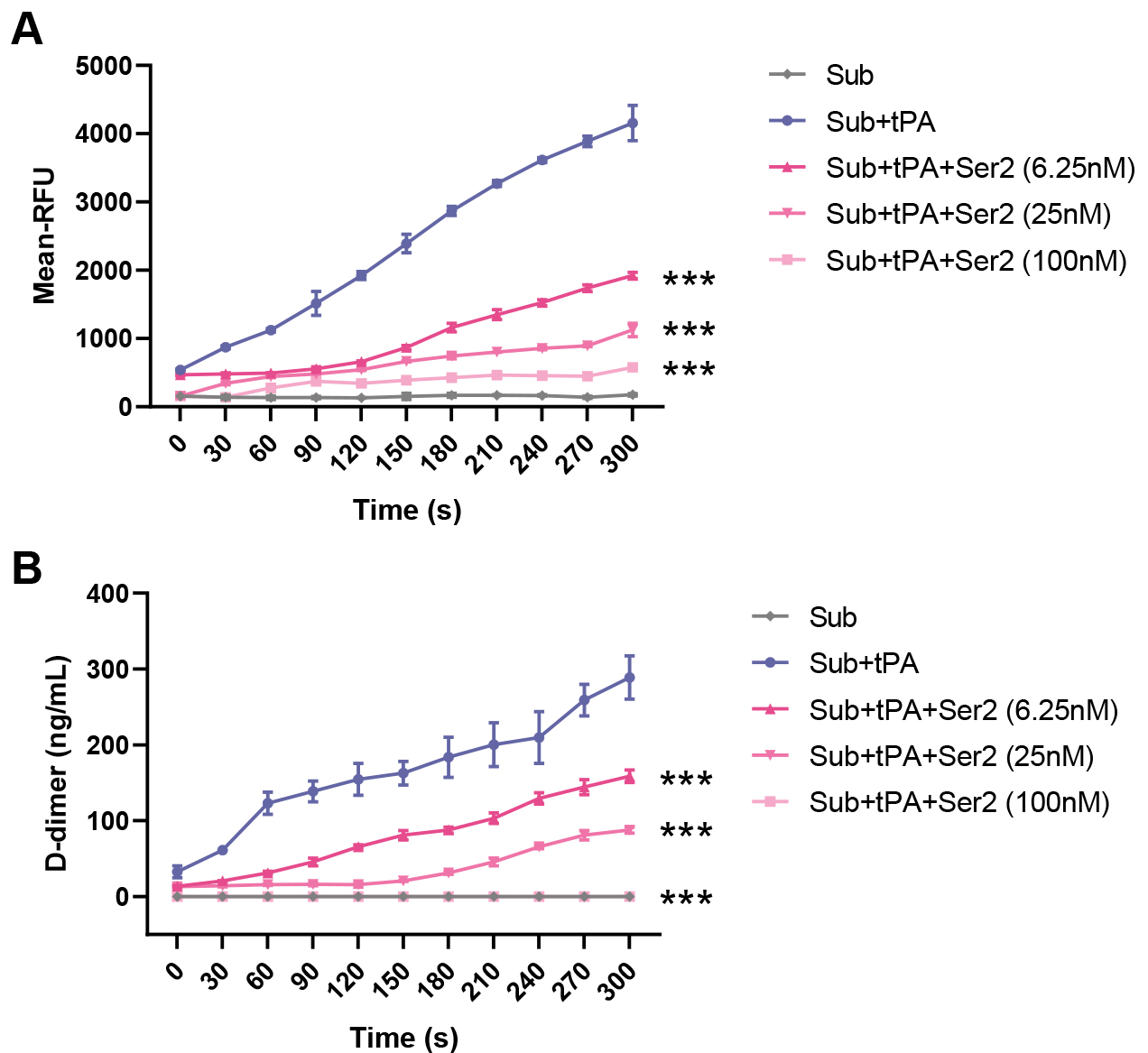 Fig. 6.
Fig. 6.
SERPINB2 inhibits tPA enzymatic activity and impairs
fibrinolytic function in vitro. (A) Time-course analysis of tPA
enzymatic activity measured by fluorometric substrate assay. (B) Fibrinolytic
function assessed by D-dimer release assay. ***p
In this study, we identified a novel SERPINB2-tPA regulatory axis in NECs that
contributes to fibrin deposition in CRSwNP. Our findings demonstrate that
SERPINB2 expression is significantly elevated in NP tissues and inversely
correlates with tPA levels, with epithelial cells serving as the primary cellular
source of this dysregulation. Importantly, we uncovered a dual inhibitory
mechanism whereby SERPINB2 not only directly inhibits tPA enzymatic activity
through classic SERPIN-mediated protease inhibition, but also suppresses tPA
expression at the transcriptional level. This regulation is driven by both Th1
(IFN-
The inverse relationship between SERPINB2 and tPA observed in nasal polyp tissues parallels findings in other fibroproliferative disorders [13, 14], suggesting a conserved pathogenic mechanism. Similar dysregulation has been reported in infectious pulmonary disease, chronic obstructive pulmonary disease, and allergic asthma [15, 16, 17], where impaired fibrinolysis contributes to excessive extracellular matrix deposition and progressive fibrosis. In the tumor microenvironment, elevated SERPINB2 has been associated with fibrin-rich matrices that promote cancer cell invasion and metastasis [18, 19]. However, our study reveals features unique to upper airway disease. Unlike PAI-1, SERPINB2 is highly expressed in nasal polyp epithelium, highlighting the central role of the airway epithelium in local hemostatic balance. The predominant epithelial localization also suggests that environmental triggers (allergens, pathogens, and pollutants) that directly stimulate epithelial cells may initiate the fibrinolytic dysfunction cascade [20]. Furthermore, the convergent regulation by both Th1 and Th2 cytokines distinguishes CRSwNP from other conditions where fibrinolytic impairment is typically associated with a specific inflammatory profile. This finding may explain why nasal polyps develop across different CRSwNP endotypes and why anti-inflammatory therapies targeting a single cytokine pathway show variable efficacy in preventing polyp recurrence.
Perhaps the most significant finding of our study is the discovery that SERPINB2 employs a dual mechanism to drive fibrinolytic dysfunction in CRSwNP. Excessive fibrin deposition is a pathological hallmark of CRSwNP, contributing to tissue remodeling and polyp recurrence [21, 22, 23]. The fibrinolytic system, primarily controlled by tPA and its inhibitors, normally maintains tissue homeostasis by clearing fibrin matrices [24]. However, the mechanisms underlying fibrinolytic impairment in nasal polyps have remained poorly understood. Our findings demonstrate that SERPINB2 disrupts this balance through two complementary pathways. Beyond its known function as a direct tPA inhibitor via SERPIN-mediated enzymatic inactivation, our data reveal a novel transcriptional regulatory role. Recombinant SERPINB2 dose-dependently suppressed tPA expression, while SERPINB2 knockdown rescued cytokine-induced tPA downregulation, providing strong evidence for this previously unrecognized mechanism. This dual inhibitory action—suppressing tPA gene expression while simultaneously inactivating residual tPA enzymatic activity—creates a synergistic anti-fibrinolytic effect that severely compromises local fibrin clearance in polyp tissues. Whether these two mechanisms interact through feedback loops remains an open question. SERPINB2-mediated inhibition of tPA activity may secondarily alter the local cytokine milieu and reinforce SERPINB2 expression in a feed-forward manner; conversely, tPA may reciprocally regulate SERPINB2 transcription through plasmin-dependent signaling. Elucidating the temporal hierarchy and potential interdependence of these two mechanisms represents an important direction for future investigation.
The molecular mechanisms underlying SERPINB2-mediated transcriptional
suppression of tPA warrant further investigation. Potential mechanisms include:
(i) SERPINB2 acting through cell surface receptor-mediated transduction signaling
to modulate downstream transcriptional activity, consistent with the demonstrated
GPER/SERPINB2 signaling axis [25]; (ii) intracellular SERPINB2 interacting with
transcription factors at the PLAT promoter, supported by evidence that
IRF1 functions as an upstream transcriptional regulator in type 1 inflammatory
environments [26]; and (iii) epigenetic modifications of the tPA gene locus,
which remain speculative and require dedicated investigation. These possibilities
align with the emerging recognition that SERPINs exert functions beyond classical
protease inhibition [27, 28]. The convergent suppression of tPA by IFN-
Our findings have important translational implications for the management of CRSwNP. First, the epithelial expression of SERPINB2 makes it potentially accessible as a disease biomarker through minimally invasive nasal brushing or lavage fluid sampling. Elevated SERPINB2 levels could serve as a surrogate marker of impaired fibrinolytic capacity and identify patients at high risk for polyp recurrence. Second, our knockdown experiments provide proof-of-concept that targeting SERPINB2 can restore tPA expression and fibrinolytic function. While therapeutic development would require extensive validation, potential strategies include neutralizing antibodies or small molecule inhibitors targeting the SERPINB2-tPA interaction. Ultimately, defective fibrin clearance and subsequent fibrin overload represent a shared pathogenic pathway underlying edematous changes across CRSwNP endotypes. Interventions targeting fibrinolytic dysfunction could provide treatment benefits independent of endotype by disrupting the self-reinforcing cascade linking fibrin deposition, tissue remodeling, and polyp formation.
Several limitations of this study should be acknowledged. First, this study relies primarily on in vitro ALI cultures and cross-sectional clinical tissue samples, and lacks in vivo functional validation. Future studies employing animal models with SERPINB2 knockout or epithelial-specific overexpression to assess fibrinolytic activity, fibrin deposition, and polyp formation would provide stronger causal evidence. Second, our in vitrostudies utilized healthy control-derived ALI cultures, which may not fully reflect the altered responsiveness of the polyp epithelium or the complex multicellular interactions in nasal polyp tissues. Third, our fibrinolysis assay utilized a serum-based system, which measures tPA-mediated plasminogen activation but does not fully recapitulate the tissue fibrin microenvironment of nasal polyps; future studies incorporating 3D fibrin gel models or ex vivo tissue explants would provide additional spatial and contextual information. Fourth, the clinical findings of this study are based on cross-sectional observations; longitudinal studies correlating SERPINB2 and tPA dynamics with disease progression and treatment response are needed to establish their clinical translational relevance.
This study identifies a novel SERPINB2-tPA regulatory axis that drives
fibrinolytic dysfunction in CRSwNP. We demonstrate that SERPINB2 employs a dual
inhibitory mechanism—suppressing tPA expression at the transcriptional level
while directly inhibiting its enzymatic activity—resulting in severely impaired
local fibrinolysis. The convergent upregulation of SERPINB2 by both the Th1
cytokine IFN-
The data presented in this study are available on request from the corresponding author.
Conceptualization, WZ, and ZL; methodology, YZ, JZ, SZ, and ZL; validation, YZ, JZ, and SZ; formal analysis, YZ, and ZL; investigation, YZ, JZ, SZ, and ZL; resources, WZ, and ZL; data curation, YZ, JZ, and SZ; writing—original draft preparation, YZ, and ZL; writing—review and editing, YZ, WZ, and ZL; visualization, YZ; supervision, WZ, and ZL; funding acquisition, WZ, and ZL. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
This study was approved by the Ethical Committee of Shanghai Sixth People’s Hospital Affiliated to Shanghai Jiao Tong University School of Medicine, with written informed consent obtained from all participants. Approval Number: 2022-069. The study was carried out in accordance with the guidelines of the Declaration of Helsinki.
We would like to express our sincere gratitude to all patients and healthy volunteers who contributed samples to this study. We also thank all peer reviewers for their constructive opinions and suggestions.
This work was supported by the National Natural Science Foundation of China [No. 82271137 and No. 82000951]; Shanghai Municipal Health Commission [SHWSRS(2023)62].
The authors declare no conflicts of interest.
During the preparation of this work, the authors used Claude to improve the grammar, readability, and organization of the manuscript text. After using this tool, the authors reviewed and edited the content as needed and took full responsibility for the content of the publication.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBL50406.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.