





 , Ruiqin Hu 1, Beiqi Yang 1, Shouwen Jiang 1, Liangbiao Chen 1,*
, Ruiqin Hu 1, Beiqi Yang 1, Shouwen Jiang 1, Liangbiao Chen 1,* , Zhong Chen 2,*
, Zhong Chen 2,*1 College of Fisheries and Life Science, Shanghai Ocean University, 201306 Shanghai, China
2 Department of Cardiology, Shanghai Sixth People’s Hospital Affiliated to Shanghai Jiao Tong University School of Medicine, 200233 Shanghai, China
Abstract
Cardiac hypertrophy is a major pathological feature of various cardiovascular diseases. Owing to the simple cardiac structure, rapid development, and genetic tractability, zebrafish are an ideal model for studying cardiac pathology. Zebrafish lacking Bone morphogenetic protein 10 (bmp10-/-) develop myocardial hypertrophy under normothermic conditions (28 °C). However, the long-term effects of chronic cold exposure on cardiac function and survival in this model remain unclear. Therefore, this study utilized the bmp10-/- zebrafish model to systematically evaluate the therapeutic potential of chronic hypothermia in attenuating myocardial hypertrophy and to elucidate the underlying molecular mechanisms.
This study was conducted in three sequential phases. Phase I: bmp10-/- (homozygous mutant, HO) zebrafish at 6 weeks post-fertilization (wpf) were maintained under normothermic (28 °C) or hypothermic (15 °C) conditions for 2 weeks to evaluate the effects of cold exposure on myocardial hypertrophy. Phase II: Building on the Phase I findings, bmp10-/- zebrafish were subjected to adaptive temperature-switching between normothermic and hypothermic conditions until 10 wpf to assess the temporal effects of cold exposure on cardiac remodeling. Phase III: Pharmacological validation experiments were performed to identify key regulatory genes mediating hypothermia-induced cardioprotection.
Long-term cold exposure significantly attenuated myocardial hypertrophy induced by bmp10 deficiency. Given that hypoxia-inducible factor-1 (HIF-1) is a canonical transcriptional regulator of cellular hypoxic responses and that the stability of HIF-1 is tightly controlled by prolyl hydroxylases such as Egl-9 family hypoxia-inducible factor 3 (EGLN3), the involvement of this pathway was further evaluated. These findings indicate that inhibition of EGLN3 activity stabilizes and activates HIF-1 signaling, thereby mediating cardioprotective effects under hypothermic conditions.
This study elucidates the functional interplay between the EGLN3 and HIF-1 signaling pathways. Under chronic hypothermia, EGLN3 modulates HIF-1 stability, contributing to the downregulation of BCL2-interacting protein 3 (BNIP3) and facilitating cardiomyocyte injury repair.
Keywords
- bone morphogenetic protein 10 (bmp10)
- hypothermia induced
- egl-9 family hypoxia-inducible factor 3 (egln3)
- hypoxia-inducible factor-1 (hif-1)
- cardiomegaly
Cardiac hypertrophy represents an adaptive response of the heart to various physiological and pathological stimuli. Physiological hypertrophy typically occurs in conditions of increased cardiac output demand and is characterized by preserved contractile function and structural integrity. In contrast, pathological hypertrophy—commonly observed in hypertension, myocardial ischemia, and cardiomyopathies—is associated with maladaptive remodeling; including ventricular wall thickening, chamber dilation, and progressive functional deterioration [1, 2, 3].
Bone morphogenetic protein 10 (bmp10) plays a central role in cardiovascular development and homeostasis [4, 5, 6]. It facilitates differentiation of human pluripotent stem cells into cardiovascular progenitors and supports arteriovenous network formation [7, 8, 9]. Loss of bmp10 disrupts normal cardiac morphogenesis [10, 11, 12], induces anemia and tissue hypoxia [13], and triggers compensatory enhancement of cardiac contractility; thereby exacerbating myocardial hypertrophy [14]. Moreover, bmp10 deficiency leads to endothelial hyperproliferation and arteriovenous malformations, further impairing oxygen circulation [15, 16]. Zebrafish share high conservation with humans in regulatory mechanisms of cardiac development, hypertrophy-related signaling pathways, and embryonic gene reactivation [17, 18, 19]. Their genetic manipulability and in vivo imaging advantages render them a valuable model for studying cardiac hypertrophy. Recent studies indicate that chronic cold exposure activates multiple protective signaling pathways [20, 21], including the hypoxia-inducible factor-1 (hif-1) and forkhead box O (foxo) pathways; which enhance vascular compliance and reduce oxidative stress and inflammatory responses [22], ultimately decreasing cardiac workload.
Therapeutic hypothermia (TH) is a clinically established intervention that
confers protection against ischemia-reperfusion injury, myocardial infarction,
and neurological disorders [23, 24]. TH exerts its protective effects by
modulating key cellular processes; including hypoxic signaling regulation,
apoptosis inhibition, and oxidative stress attenuation [25, 26, 27, 28]. Mild hypothermia
has been reported to suppress hypoxia-inducible factor-1α (hif-1
Therefore, this study focuses on the molecular mechanisms underlying hypothermia-induced reversal of cardiac hypertrophy in bmp10-deficient zebrafish, aiming to provide experimental evidence for environmental modulation of myocardial hypertrophy and to expand the investigative scope of the zebrafish model in this field.
The bmp10 gene was knocked out in zebrafish using CRISPR–Cas9 technology, generating bmp10+/- and bmp10-/- mutants. kdrl:EGFP transgenic zebrafish, including bmp10 wild type (bmp10+/+, WT) and bmp10-/- variants, were derived from the AB wild-type strain (Danio rerio), obtained from the National Zebrafish Resource Center (Wuhan, China). These gene-edited zebrafish served as parental fish for generating the bmp10-/- offspring. For convenience in selected figure legends, bmp10-/- zebrafish are termed HO, and bmp10+/+ zebrafish as WT. In this study, 15 °C is defined as hypothermic conditions, whereas 28 °C represents normothermic conditions.
The bmp10 gene is located on chromosome 5. Gene knockout was achieved using the CRISPR–Cas9 system targeting the sequence GGGTTGGATGATGGACATGG, resulting in a 72-bp deletion. gRNA was synthesized using the MAXIscript™ T7 in vitro transcription kit (AM1314, Ambion [Invitrogen], Waltham, MA, USA), and Cas9 protein (C120010, Sigma-Aldrich, St. Louis, MO, USA) (800 pg) with gRNA (100 pg) was co-injected into one-cell stage embryos. Genotyping of bmp10 was performed by PCR using the following primers: forward, ATTTTCATCTGCCACGTCTGC; reverse: TAGAGTTCCAGCATATACTCAG. PCR amplification was conducted using Takara rTaq DNA polymerase (RR901A, Takara Bio Inc., Kusatsu, Shiga, Japan). bmp10-/- mutants were identified by agarose gel electrophoresis using molecular biology-grade agarose (75510-019, Invitrogen, Carlsbad, CA, USA).
Offspring from bmp10+/- self-crosses were raised under normothermic conditions (28 °C) until 6 weeks post fertilization (wpf). Juvenile zebrafish (bmp10+/+, bmp10-/-, and bmp10+/-) were then assigned to either a hypothermic group 15 °C or a normothermic control group 28 °C.
The hypothermic exposure protocol consisted of three stages:
Stage 1: Two-week hypothermic exposure
At 6 wpf, zebrafish were exposed to hypothermic or normothermic conditions for 2 weeks (until 8 wpf; followed by analyses of body weight, cardiac morphology, gene expression, apoptosis, histology, and blood cell counts.
Stage 2: Temperature-switching experiments
(1) Zebrafish exposed to hypothermia from 6–8 wpf were transferred to normothermic conditions until 10 wpf.
(2) Zebrafish maintained under normothermic conditions from 6–8 wpf were transferred to hypothermic conditions until 10 wpf.
Stage 3: Long-term hypothermic exposure
Beginning at 6 wpf, zebrafish were continuously maintained under hypothermic conditions for 8 weeks (until 14 wpf).
Angiotensin-converting enzyme inhibitors (ACEIs), such as benazepril
hydrochloride (HY-B0093A, MedChemExpress, Monmouth Junction, NJ, USA), reduce
blood pressure and alleviate cardiac load by inhibiting the
renin–angiotensin–aldosterone system. The experiment utilized
bmp10+/- zebrafish offspring, which were raised at 28 °C
until they reached 6 wpf. At the start of the experiment, equal numbers of
juveniles (bmp10+/+; bmp10-/-) were assigned to
either the benazepril hydrochloride treatment group or the control group (which
received only the solvent without the drug) for a 2-week treatment period (until
8 wpf). Before the experiment, juveniles in both groups were allocated in equal
numbers, with each tank containing no more than 15 juveniles (ensuring a sample
size of n
Zebrafish were anesthetized, and hearts were rapidly dissected and fixed in 4% paraformaldehyde (E672002-0500, BBI Life Sciences, Shanghai, China) at 4 °C for 24 h. Following fixation, samples were subjected to graded ethanol (A500737-0500, BBI Life Sciences, Shanghai, China) dehydration, xylene (X103350, Sinopharm Chemical Reagent Co., Ltd., Beijing, China) clearing, and paraffin embedding. Paraffin-embedded heart tissues were sectioned into 4 µm-thick serial sections using Leica low-profile disposable blades 819 (14035838925, Leica Biosystems, Wetzlar, Germany) for subsequent histological analyses. Heart sections were stained with hematoxylin and eosin staining kit (E607318-0200, BBI Life Sciences, Shanghai, China) and masson’s trichrome staining kit (G1340, Solarbio, Beijing, China) in accordance with the manufacturers’ instructions to assess overall cardiac morphology and histological features. Images were acquired using a stemi 2000 stereomicroscope (Zeiss, Oberkochen, Germany).
Myocardial cell apoptosis in cardiac sections (4 µm) was examined using the TUNEL BrightGreen apoptosis detection kit (A112-02, Vazyme Biotech Co, Nanjing, China). The specimens were mounted with anti-fade mounting medium (C02-04003, Bioss Antibodies, Beijing, China) medium for fluorescence and sealed. Apoptotic signals were captured using a LSM710 confocal laser scanning microscope (Carl Zeiss AG, Oberkochen, Germany), and quantitative analysis of the apoptotic signals was performed using ImageJ software (version 1.53, National Institutes of Health, Bethesda, MD, USA).
Zebrafish were anesthetized with 0.02% tricaine (MS-222) (A5040, Sigma-Aldrich, St. Louis, MO, USA). Approximately 0.5 µL of peripheral blood was collected via tail vein transection and immediately diluted with 2 µL of ultrapure water. The diluted blood was dropped onto one end of a glass slide, and blood smears were prepared using the push–slide method. After air-drying, smears were fixed with absolute methanol (A601617, BBI Life Sciences, Shanghai, China) for 5 min. Blood smears were stained using a Wright–Giemsa staining kit (E607314, BBI Life Sciences, Shanghai, China), and blood cell morphology was analyzed under a light microscope.
Paraffin-embedded zebrafish heart sections were deparaffinized prior to staining. Cell membranes were stained with wheat germ agglutinin, alexa fluor 488 conjugate (G1730, Invitrogen, Carlsbad, CA, USA) for 15 min at room temperature in the dark, followed by three washes with phosphate-buffered saline (PBS) (E607008-0500, BBI Life Sciences, Shanghai, China). Nuclei were counterstained with Hoechst 33258 dye (33258, Solarbio Life Sciences, Beijing, China) for 5 min at room temperature in the dark, followed by PBS washes. After staining, sections were mounted using an anti-fade mounting medium (C02-04003, Bioss Antibodies, Beijing, China) and imaged using a confocal microscope.
Gene sequences for bmp10, bone morphogenetic protein 10-like (bmp10l), and Hypoxia-inducible factor 3 (egln3) were downloaded from the Ensembl website, and primers were designed based on the CDSs of these genes. The primer sequences for bmp10 were F: ATTTTCATCTGCCACGTCTGC and R: TAGAGTTCCAGCATATACTCAG. The primer sequences for bmp10l were F: AGCGGAGATGTACGTGGATT and R: CTGGACGATGGCGTGTTTAG. The primer sequences for egln3 were F: GCTTTGTTGGATCAGGGCTT and R: GATGTTCCTCCGGCAAACTC. First, RNA extraction was performed using the RNeasy Mini Kit (74104, Qiagen N.V, Hilden, Germany), followed by cDNA synthesis using TransScript Uni All-in-One First-Strand cDNA Synthesis SuperMix for qPCR (AU341-02-V2, TransGen Biotech Co, Beijing, China) with the extracted RNA. Finally, quantitative fluorescence analysis was performed using TB Green Premix Ex Taq II (Tli RNaseH Plus) (RR420A, Takara Bio Inc, Kusatsu, Shiga, Japan) on a CFX Opus 96 Real-Time PCR System (Bio-Rad Laboratories, Inc., Hercules, CA, USA) instrument.
All statistical analyses were performed using GraphPad Prism software (version
8.0.1, GraphPad Software Inc., San Diego, CA, USA). Data are presented as mean
Zebrafish larvae were immersed in 200 mg/L tricaine (MS-222; Sigma–Aldrich) for approximately 5 minutes until all movement and responses completely ceased. For euthanasia, zebrafish were exposed to an overdose of 400 mg/L tricaine for 15 minutes until they were completely unresponsive and cardiac activity had stopped. To ensure complete death, the fish were subsequently subjected to an ice-water bath at approximately 0 °C for 5 minutes following anesthesia. This procedure was conducted in accordance with the AVMA Guidelines for the Euthanasia of Animals (2020 edition). The tricaine stock was prepared in deionized water and diluted with system water to working concentrations of 200 or 400 mg/L. The pH of the working solutions was adjusted to 7.0–7.5 using sodium bicarbonate (NaHCO3) (A100865-0500, BBI Life Sciences, Shanghai, China).
Chronic hypothermic exposure markedly reversed cardiac hypertrophy in bmp10-/- zebrafish (Fig. 1A,B). Under normothermic conditions (28 °C), bmp10-/- zebrafish exhibited pronounced cardiac hypertrophy by 8 wpf (Fig. 1A,B). Following chronic hypothermic exposure (15 °C), heart size was reduced in both wild-type and bmp10-/- zebrafish, and the difference between the two groups was no longer detectable. These results indicate that hypothermic exposure attenuates myocardial hypertrophy induced by bmp10 deficiency, rather than producing a mutant-specific effect. Consistently, under normothermic conditions, body weight in 8-week-old bmp10-/- zebrafish differed significantly from that of control fish, and this difference was more pronounced than that observed at the early stage (6 weeks of age) (Fig. 1C). Notably, following hypothermic exposure, this difference was markedly abolished (Fig. 1D). Based on these findings, bmp10-/- zebrafish were selected as an appropriate experimental model for subsequent mechanistic investigations under hypothermic conditions. Downregulation of egln3 (Fig. 1G–I) was accompanied by activation of the hif-1 signaling pathway involved in cardiomyocyte hypoxic responses, together with significant downregulation of the pro-apoptotic genes bnip3 and bcl2l10 (Fig. 2F). This molecular profile is consistent with reduced cardiomyocyte size (Fig. 1B) and attenuation of myocardial injury. Although bmp10l exhibits partial functional redundancy with bmp10 during cardiac development, its expression did not fully compensate for the structural defects caused by bmp10 deficiency (Fig. 1E,F). In addition, hypothermic exposure significantly increased the expression of Kruppel-like factor 1 (klf1) in bmp10-/- zebrafish hearts (Fig. 2F), a gene implicated in myocardial injury repair and cardiomyocyte proliferation.
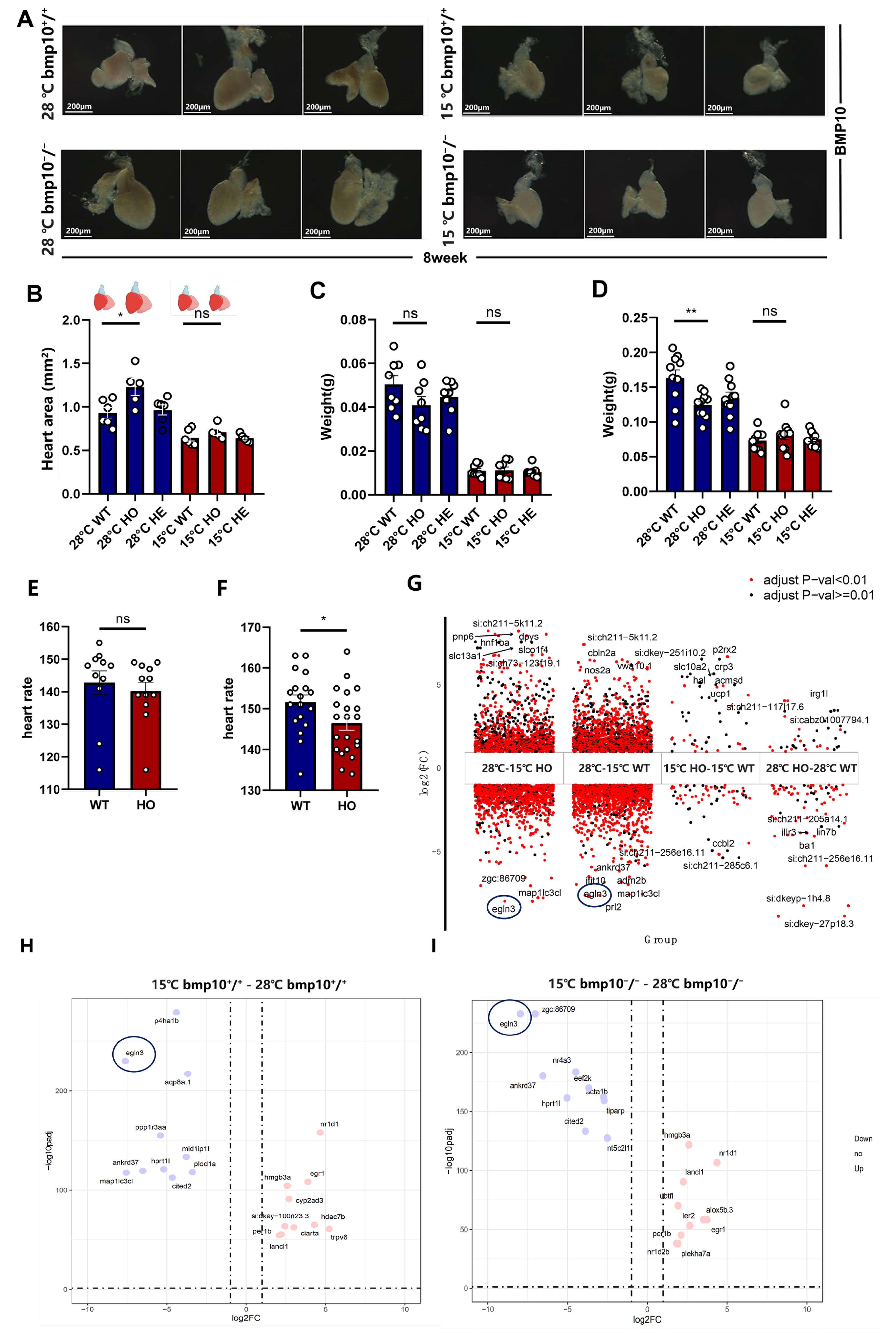 Fig. 1.
Fig. 1.
Comparison of zebrafish lacking bone morphogenetic protein 10 (bmp10-/-) treated at different temperatures. (A) Calculation of cardiac area
based on heart chambers (ventricle, atrium, and bulbus arteriosus). Scale bar =
200 µm. (B) The cardiac area of bmp10-/- zebrafish maintained
under normothermic conditions (28 °C) was significantly larger than that
of bmp10-/- zebrafish subjected to chronic cold exposure under
hypothermic conditions (15 °C). (C) Body weight of
bmp10-/- zebrafish at 6 weeks of age (4 weeks of age followed by 2
weeks of hypothermic treatment) under normothermic and hypothermic conditions.
(D) Body weight of bmp10-/- zebrafish at 8 weeks of age (6 weeks of
age followed by 2 weeks of hypothermic treatment) under normothermic and
hypothermic conditions. (E) Heart rate measurements of bmp10-/-
zebrafish at 72–84 hours post-fertilization (hpf) under normothermic conditions
(28 °C). (F) Heart rate measurements of bmp10-/- zebrafish
at 96–108 hpf under normothermic conditions (28 °C). (G) Gene
expression comparisons under different temperature conditions: 28 °C–15
°C HO (gene expression differences in bmp10-/- zebrafish
between 28 °C and 15 °C), 28 °C–15 °C WT
(gene expression differences in bmp10+/+ zebrafish between 28
°C and 15 °C), 15 °C HO–15 °C WT (gene
expression differences between bmp10-/- and bmp10+/+
zebrafish at 15 °C), and 28 °C HO–28 °C WT (gene
expression differences between bmp10-/- and bmp10+/+ zebrafish at 28 °C). (H) Analysis of Hypoxia-inducible factor-1 (hif-1) pathway factor
Egl-9 family hypoxia-inducible factor 3 (egln3) expression in bmp10+/+ zebrafish under hypothermic
(15 °C) and normothermic (28 °C) conditions. (I) Analysis of
hif-1 pathway factor egln3 expression in bmp10-/-
zebrafish under hypothermic (15 °C) and normothermic (28 °C)
conditions. Data are presented as mean
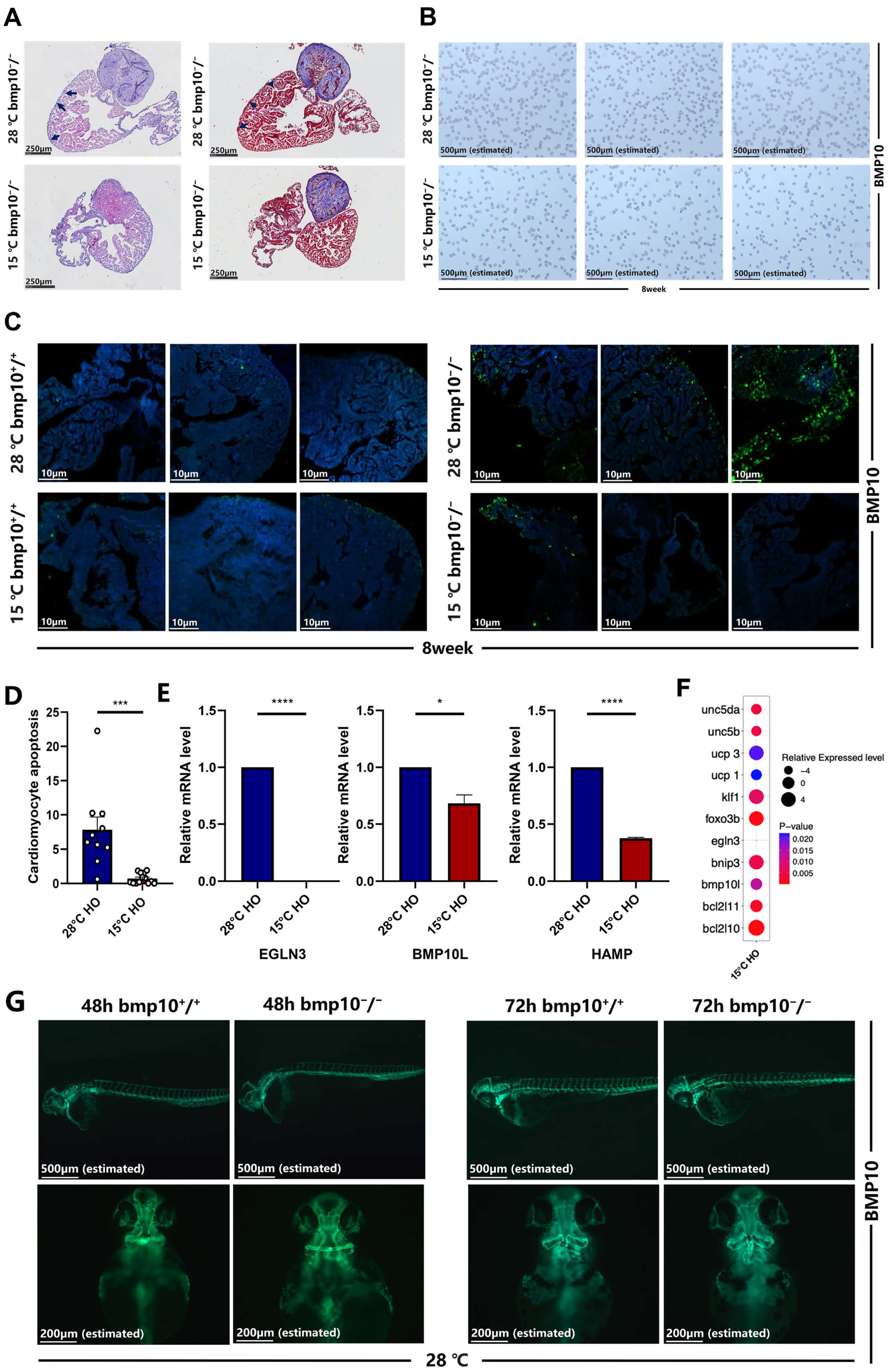 Fig. 2.
Fig. 2.
Chronic cold exposure alleviates cardiomyocyte hypertrophy. (A)
Morphological and fibrotic staining analysis of bmp10-/- zebrafish
hearts after 2 weeks of exposure to hypothermic (15 °C) or normothermic
(28 °C) conditions (from 6–8 wpf). Scale bar = 250 µm. (B) Red
blood cell density analysis of bmp10-/- zebrafish hearts after 2
weeks of exposure to hypothermic (15 °C) or normothermic (28
°C) conditions (from 6–8 wpf). Scale bar = 500 µm (estimated).
(C) Cardiomyocyte apoptosis comparison between bmp10-/- and
bmp10+/+ zebrafish under different temperature conditions. Scale
bar = 10 µm. (D) Quantitative analysis of cardiomyocyte apoptosis in panel
C. (E) Relative mRNA expression levels of egln3, bone morphogenetic protein 10-like (bmp10l), and hepcidin antimicrobial peptide (hamp) in
bmp10-/- zebrafish hearts after 2 weeks of exposure to hypothermic
(15 °C) or normothermic (28 °C) conditions (by 8 wpf). (F)
Relative mRNA expression levels of egln3, Bone morphogenetic protein 10-like (bmp10l), Kruppel-like factor 1 (klf1), BCL2 interacting protein 3 (bnip3), BCL2-like 10 (bcl2l10), BCL2-like 11 (bcl2l11),
forkhead box O (foxo3b), Uncoupling protein 1 (ucp1), Uncoupling protein 3 (ucp3), Unc-5 netrin receptor b (unc5b), and
Unc-5 netrin receptor da (unc5da) in bmp10-/- zebrafish cardiomyocytes under
hypothermic (15 °C) and normothermic (28 °C) conditions. For
individual gene expression levels, refer to Supplementary Fig. 1. (G) Imaging of major arteries in bmp10-/- and
bmp10+/+ zebrafish at 48 and 72 hpf under normothermic conditions
(28 °C). Scale bar = 200 µm (estimated). Data are presented as
mean
To ameliorate cardiac hypertrophy that had already by 8 wpf, bmp10-/- zebrafish were subjected to hypothermic exposure for 2 weeks starting at 6 wpf (Fig. 3Aa). Following hypothermic intervention (15 °C), bmp10-/- zebrafish exhibited substantial alterations in overall cardiac morphology (Fig. 2A). Moreover, erythrocyte density within the cardiac chambers showed substantial differences compared with those maintained under normothermic conditions (28 °C) (Fig. 2B). Hypothermic exposure significantly reduced cardiomyocyte apoptosis, as demonstrated by TUNEL staining and quantitative analysis (Fig. 2C,D). Gene expression analysis indicates that egln3 expression was significantly lower under hypothermic conditions than under normothermic conditions (Fig. 1G–I). These data indicate that egln3 downregulation may contribute to reduced cardiomyocyte apoptosis in bmp10-/- zebrafish.
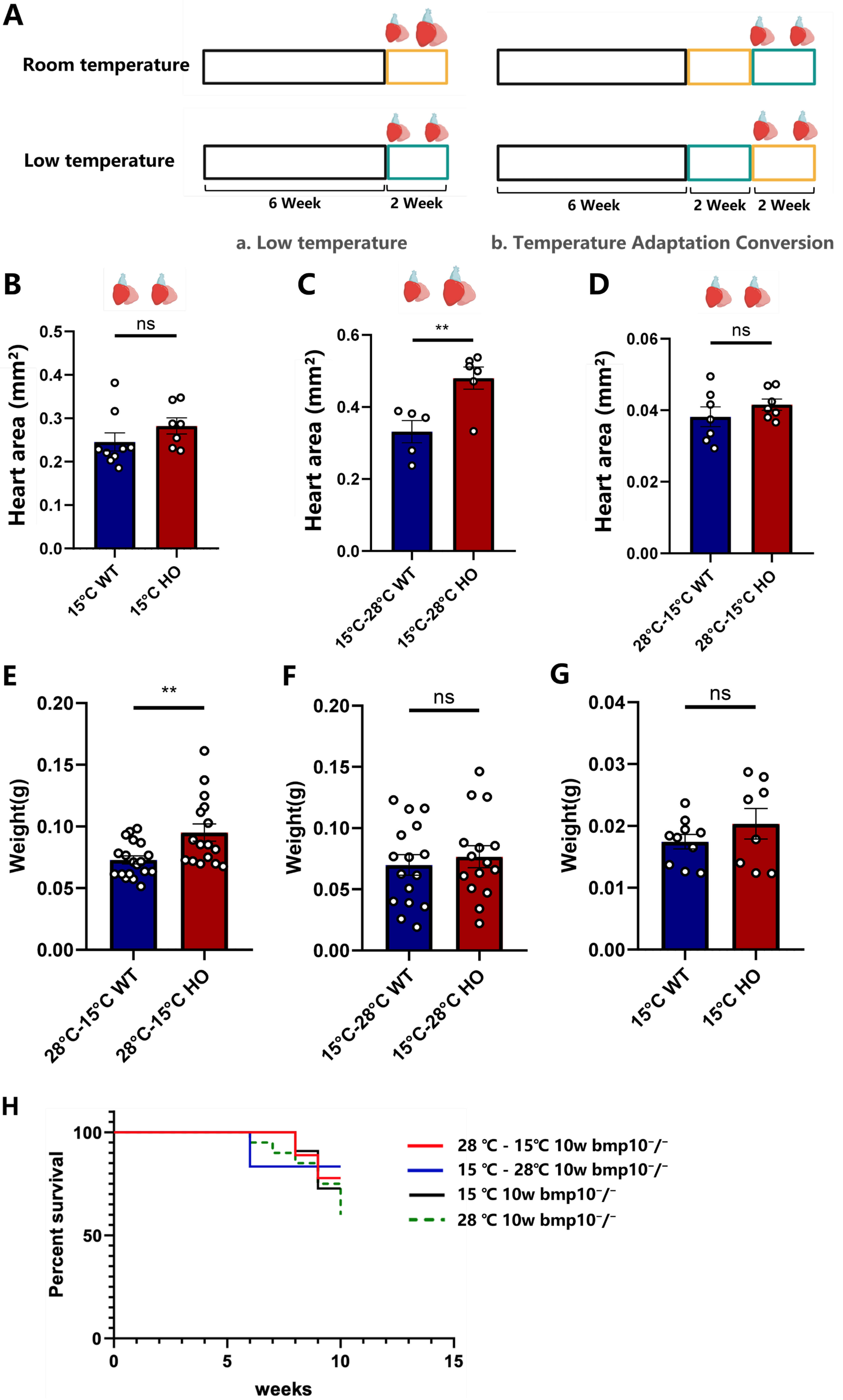 Fig. 3.
Fig. 3.
Cardiac and overall health status of bmp10-/-
zebrafish following temperature transitions. (A) Low temperature: Normothermic
group (Control group)—bmp10-/- zebrafish were maintained under
normothermic conditions (28 °C) until 8 wpf. Hypothermic group
(Experimental group)—bmp10-/- zebrafish were maintained under
normothermic conditions until 6 wpf and then transferred to hypothermic
conditions (15 °C) for 2 weeks (until 8 wpf, green box). Temperature
adaptation protocol: (i) bmp10-/- zebrafish were maintained
under normothermic conditions (28 °C) until 6 wpf, followed by an
additional 2 weeks at 28 °C (until 8 wpf; yellow box), and subsequently
transferred to hypothermic conditions (15 °C) for 2 weeks (until 10
wpf); (ii) bmp10-/- zebrafish were maintained at 28 °C
until 6 wpf, then exposed to 15 °C for 2 weeks (green box), and finally
returned to 28 °C for an additional 2 weeks (until 10 wpf). (B) Cardiac
area comparison between bmp10-/- and bmp10+/+
zebrafish following 8 weeks of chronic cold exposure (until 14 wpf). (C) Cardiac
area comparison between bmp10-/- and bmp10+/+
zebrafish after transitioning from hypothermic (15 °C) back to
normothermic (28 °C) conditions (until 10 wpf). (D) Cardiac area
comparison between bmp10-/- and bmp10+/+ zebrafish after transitioning from normothermic (28 °C) to hypothermic
(15 °C) conditions (until 10 wpf). (E) Body weight comparison between
bmp10-/– and bmp10+/+ zebrafish following
temperature transition from normothermic to hypothermic conditions (until 10
wpf). (F) Body weight comparison between bmp10-/- and
bmp10+/+ zebrafish after transitioning from hypothermic back to
normothermic conditions (until 10 wpf). (G) Body weight comparison between
bmp10-/- and bmp10+/+ zebrafish following 8 weeks of
chronic cold exposure (until 14 wpf). (H) Survival rates of
bmp10-/- zebrafish maintained under normothermic conditions (28
°C) or exposed to three different hypothermic treatment (until 10 wpf).
For individual survival curves at each stage, refer to Supplementary Fig. 2. Data are presented as mean
Forkhead box O (foxo) family members (including foxo3b, foxo1a, and
foxo1b) can modulate the stability and activity of
hif-1
Fertilized embryos transferred to 15 °C within 3 hours post-fertilization (hpf) resulted in embryonic lethality by 48 hpf. Comparative analysis of cardiac vascular morphology in bmp10-/- zebrafish maintained at 28 °C demonstrated substantial differences at 48 and 72 hpf relative to control embryos (Fig. 2G), indicating that bmp10 plays an essential role in vascular development. Differential gene expression analysis demonstrated significant upregulation of unc-5 netrin receptor b (unc5b) and unc-5 netrin receptor da (unc5da) in bmp10-/- zebrafish hearts under hypothermic conditions (Fig. 2F). These genes are involved in endothelial cell migration and proliferation and may influence vascular network formation and cardiac contractile function.
bmp10-/- zebrafish exposed to hypothermic conditions (15 °C) for 2 weeks starting at 6 wpf exhibited significant improvement in cardiac hypertrophy (Fig. 1B; Fig. 3Aa). In zebrafish transitioned from normothermic to hypothermic conditions at 8 wpf, morphological abnormalities associated with bmp10 deficiency were significantly alleviated after 2 weeks of hypothermic exposure (Fig. 3Ab–D; Fig. 3E; Fig. 3F; Fig. 4G,I,J). Survival was also significantly improved compared with zebrafish maintained under normothermic conditions (Fig. 3H).
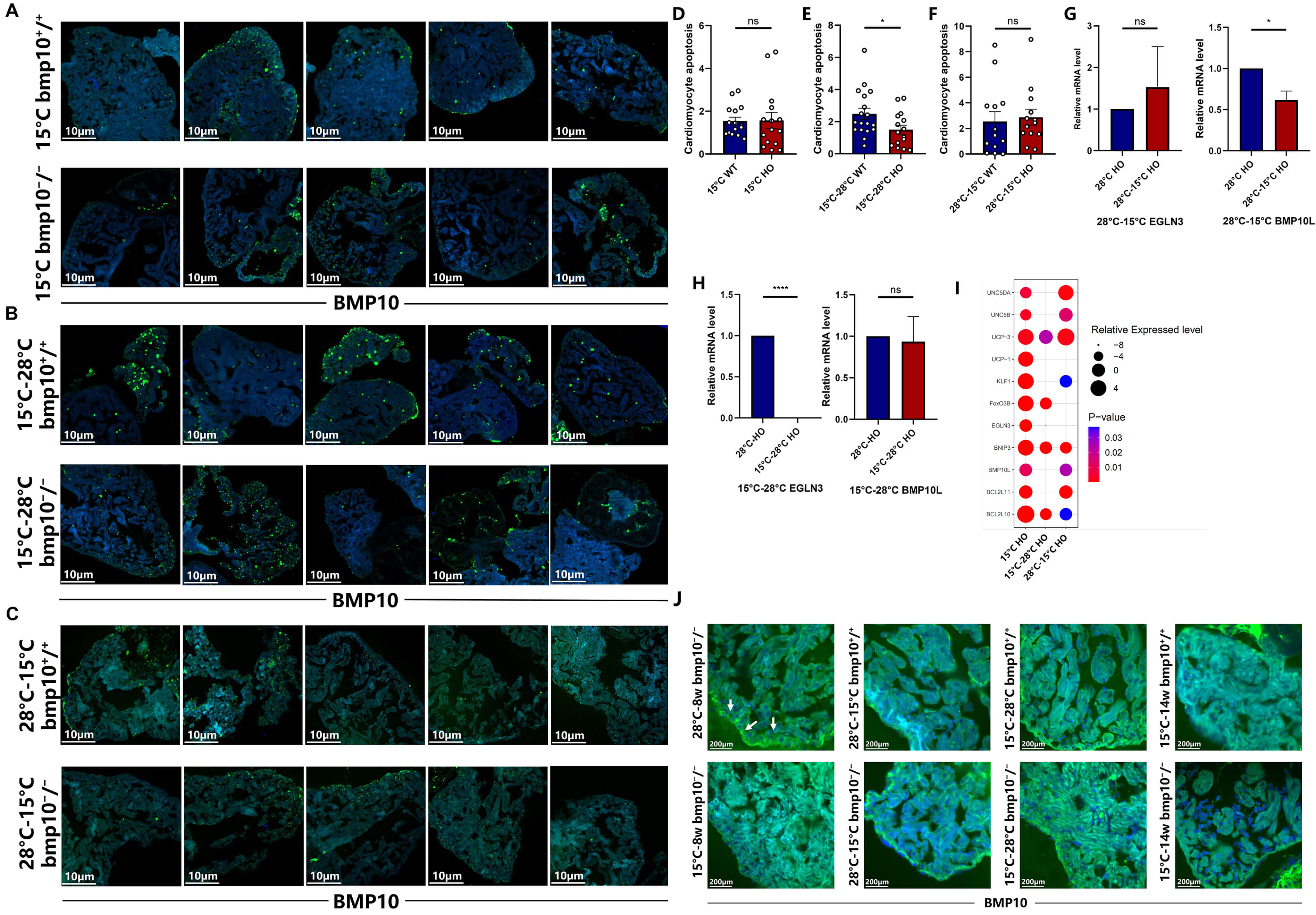 Fig. 4.
Fig. 4.
Sustained improvement in cardiac health following temperature
transitions. (A) Cardiomyocyte apoptosis in bmp10-/- and
bmp10+/+ zebrafish following 8 weeks of chronic cold exposure
(until 14 wpf). Scale bar = 10 µm. (B) Cardiomyocyte apoptosis in
bmp10-/- and bmp10+/+ zebrafish after
transitioning from hypothermic (15 °C) back to normothermic (28
°C) conditions (until 10 wpf). Scale bar = 10 µm. (C)
Cardiomyocyte apoptosis in bmp10-/- and bmp10+/+ zebrafish after transitioning from normothermic (28 °C) to
hypothermic (15 °C) conditions (until 10 wpf). Scale bar = 10 µm.
(D) Quantitative analysis of cardiomyocyte apoptosis in panel A. (E) Quantitative
analysis of cardiomyocyte apoptosis in panel B. (F) Quantitative analysis of
cardiomyocyte apoptosis in panel C. (G) Relative expression levels of
egln3 and bmp10l after transitioning from normothermic to
hypothermic conditions (until 10 wpf). (H) Relative expression levels of
egln3 and bmp10l after transitioning from hypothermic back to
normothermic conditions (until 10 wpf). (I) Relative expression levels of
egln3, bmp10l, klf1, bnip3, bcl2l10,
bcl2l11, foxo3b, ucp1, ucp3, unc5b, and
unc5da following three distinct chronic cold exposure paradigms. For
individual gene expression data, refer to Supplementary Figs. 1,3,4. (J) Morphological analysis of zebrafish cardiomyocytes under different
temperature transition paradigms: (i) 2 weeks of cold exposure at 15 °C
starting at 6 wpf (until 8 wpf); (ii) 2 weeks at 28 °C followed by 2
weeks at 15 °C (until 10 wpf); (iii) 2 weeks at 15 °C followed
by return to 28 °C for 2 weeks (until 10 wpf); and (iv) 8 weeks of
chronic cold exposure at 15 °C (until 14 wpf). Scale bar = 200 µm. The area at the edge of the tissue indicated by the white arrow is the ventricular wall of a zebrafish.
Data are presented as mean
To assess whether the cardioprotective effects of hypothermic exposure were transient, bmp10-/- zebrafish were maintained under hypothermic conditions until 8 wpf and then transferred back to normothermic conditions (Fig. 3Ab). The cardioprotective effects persisted following rewarming, with sustained improvements in cardiac morphology and survival (Fig. 3H; Fig. 4H,I,J). Growth delay induced by early-stage hypothermic exposure gradually normalized after return to normothermic conditions (Fig. 3F).
Under normothermic conditions, bmp10 deficiency significantly reduced survival after 6 wpf compared with wild-type controls (Fig. 3H). Continuous hypothermic exposure from 6–14 wpf significantly increased survival, reaching 72.7% compared with 45% under normothermic conditions (Fig. 3H). These data demonstrate that chronic hypothermic exposure improves both survival and cardiovascular health in bmp10-/- zebrafish (Fig. 4A–F; Fig. 3).
Continuous treatment of bmp10-/- zebrafish with benazepril hydrochloride (50 µM, 1 hour daily for two weeks), an ACEI, significantly ameliorated myocardial hypertrophy and reduced cardiomyocyte apoptosis (Fig. 5A–C). Concomitantly, erythrocyte density increased and ventricular wall thickness approached normal levels (Fig. 5D,E). These phenotypic improvements were accompanied by downregulation of egln3 expression. The molecular and phenotypic alterations induced by ACEI treatment closely resembled those observed under chronic hypothermic exposure (Fig. 5F).
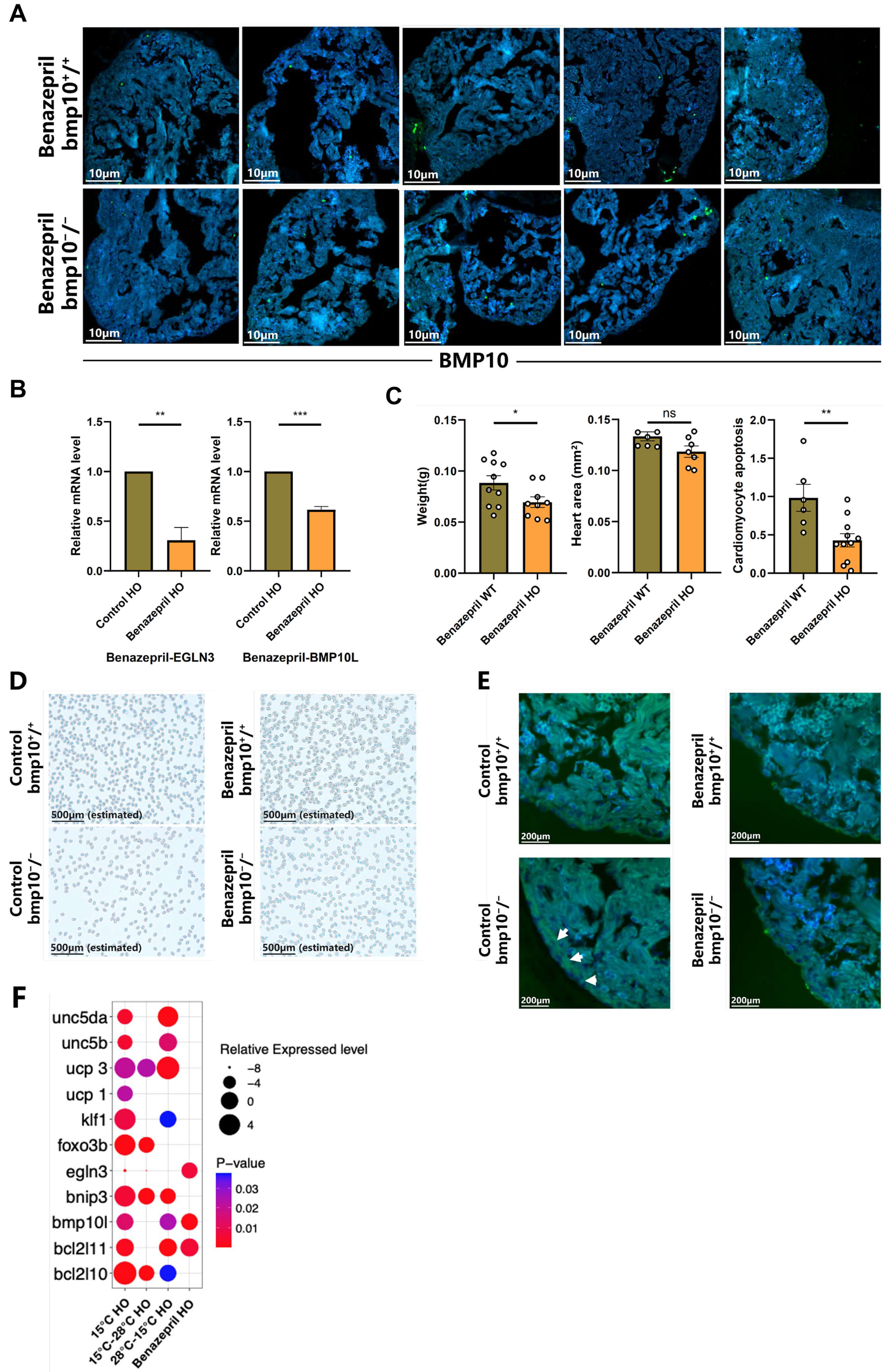 Fig. 5.
Fig. 5.
Treatment of bmp10-/- zebrafish with ACEIs. (A) Cardiomyocyte apoptosis in
bmp10-/- and bmp10+/+ zebrafish after 2 weeks of
ACEI treatment under normothermic conditions (28 °C) (until 10 wpf).
Scale bar = 10 µm. (B) Relative expression levels of egln3 and
bmp10l following 2 weeks of ACEI treatment (until 10 wpf). (C)
Quantitative analysis of body weight, heart area, and cardiomyocyte apoptosis in
bmp10-/- zebrafish following 2 weeks of ACEI treatment (until 10
wpf). (D) Erythrocyte density in bmp10-/- zebrafish following 2
weeks of ACEI treatment (until 10 wpf). Scale bar = 500 µm (estimated). (E)
Morphological analysis of myocardial structure in bmp10-/-
zebrafish following 2 weeks of ACEI treatment (until 10 wpf). Scale bar = 200
µm. The area at the edge of the tissue indicated by the white arrow is the ventricular wall of a zebrafish. (F) Expression levels of egln3, bmp10l, klf1, bnip3, bcl2l10, bcl2l11,
foxo3b, ucp1, ucp3, unc5b, and
unc5da following 2 weeks of ACEI treatment. For individual gene
expression data, refer to Supplementary Fig. 5. Data are
presented as mean
In this study, the effects of chronic cold exposure on pathological myocardial hypertrophy were systematically evaluated by using the bmp10-/- zebrafish model. Sustained hypothermia significantly improved cardiac structural remodeling, reduced cardiomyocyte apoptosis, and enhanced survival. At the molecular level, cold exposure was associated with downregulation of egln3, activation of the hif-1 signaling pathway, and reprogramming of anti-apoptotic gene expression profiles. Together, these changes suggest the establishment of a coordinated adaptive network that mitigates bmp10 deficiency–induced pathological remodeling.
bmp10 deficiency resulted in progressive ventricular wall thickening
and chamber dilation, whereas chronic hypothermia markedly attenuated these
structural abnormalities. Morphological and quantitative assessments indicate
that cold intervention delayed or partially reversed hypertrophic progression,
providing a phenotypic foundation for mechanistic studies. Reduced egln3
expression under hypothermic conditions coincided with enhanced hif-1
pathway activity. Given that egln family members suppress
hif-1
In the present model, egln3 downregulation was accompanied by increased
hif-1 pathway activity, suggesting a functional link between
temperature-dependent signaling and hypoxic adaptation in the
bmp10-deficient heart. Foxo family members (including
foxo3b, foxo1a, and foxo1b) are participants in
hypoxia and stress-responsive signaling networks. The present findings suggest a
potential association between foxo signaling and egln3
expression [37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49], possibly contributing to autophagy-related pathways and
structural remodeling [50, 51]. However, the precise regulatory relationship
between foxo factors and egln3 requires further mechanistic
investigation. Upregulation of hif-1
Transition from hypothermic to normothermic conditions partially preserved cardioprotective effects. Long-term hypothermia (6–14 wpf) significantly improved survival [59], with early intervention yielding superior outcomes, underscoring the importance of the therapeutic time window. Pharmacological ACEI treatment partially recapitulated the molecular and phenotypic effects of hypothermia, supporting the involvement of egln3-associated signaling networks, although causality remains to be definitively established. Consistent with previous reports, bmp10 deficiency induces progressive cardiac hypertrophy during early-stage development [60]. As remodeling advances, hypoxia-related signaling, oxidative stress, and cellular stress responses become increasingly pronounced. The present findings suggest that chronic hypothermia modulates these interconnected processes, promoting a more adaptive cardiac phenotype. Whether bmp10 deficiency results in chamber-specific hypertrophy or regional vulnerability requires further investigation.
Collectively, this study identifies hypothermic modulation as a significant environmental determinant of pathological cardiac remodeling. Unlike conventional pharmacological approaches, temperature modulation may exert cardioprotective effects through systemic network reprogramming, offering new insights into adaptive mechanisms under cardiac stress.
Although the present study demonstrates that chronic hypothermic exposure
attenuates myocardial hypertrophy in bmp10⁻/⁻ zebrafish through
suppression of cardiomyocyte apoptosis and modulation of the
egln3–hif-1
In conclusion, using a bmp10-/- zebrafish model, this study demonstrates that chronic cold exposure is associated with attenuation of pathological cardiac hypertrophy, reduced cardiomyocyte apoptosis, and improved survival. These beneficial effects coincide with coordinated modulation of hypoxia-related signaling, apoptotic pathways (including bnip3 and bcl2l10), oxidative stress responses (ucp1 and ucp3), and cardiac remodeling markers such as klf1. The present findings support the potential relevance of hypothermic modulation in influencing cardiac stress adaptation and pathological remodeling. Rather than establishing a definitive mechanistic pathway, this work provides a data-supported framework suggesting that egln3-associated signaling and downstream hypoxia-responsive networks may play important roles in mediating cardioprotective effects under low-temperature conditions. Taken together, this study offers a rationale for further investigation of hypothermia-associated signaling pathways in cardiac disease models. Future studies using mammalian systems and targeted mechanistic approaches will be required to validate these associations and to evaluate the translational potential of hypothermic strategies for chronic cardiac conditions, including hypertrophic cardiomyopathy.
bmp10, bone morphogenetic protein 10; bmp10l, bone morphogenetic protein 10-like; bmp10-/- (HO), bmp10 homozygous mutant; bmp10+/+ (WT), bmp10 wild type; wpf, weeks post fertilization; hpf, hours post-fertilization; egln3, egl-9 family hypoxia-inducible factor 3; hif-1, hypoxia-inducible factor 1; bnip3, BCL2 interacting protein 3; bcl2l10, BCL2-like 10; bcl2l11, BCL2-like 11; foxo, forkhead box O; ucp1, uncoupling protein 1; ucp3, uncoupling protein 3; unc5b, Unc-5 netrin receptor B; unc5da, Unc-5 netrin receptor da; ACE, angiotensin-converting enzyme; ACEI, angiotensin-converting enzyme inhibitor; TH, therapeutic hypothermia.
Further information and requests should be directed to the lead contact, Ph.D, Professor Zhong Chen (zhongchen7498@hotmail.com).
FL, ZC, and LBC designed the study and contributed to the conceptualization of the manuscript. FL drafted the original manuscript and contributed to writing, editing, visualization, validation, software, resources, methodology, investigation, and data management. ZC and LBC reviewed and edited the manuscript, and contributed to methodology, supervision, validation, resources, investigation, formal analysis, and funding acquisition. RQH participated in manuscript review and editing, and contributed to resources, investigation, formal analysis, supervision, validation, data acquisition, and analysis. BQY contributed to manuscript review and editing, visualization, software, validation, data acquisition and analysis, and supervision. SWJ participated in manuscript review and editing, provided resources, and contributed to formal analysis, data acquisition and analysis, investigation, validation, and supervision. All authors contributed to manuscript revision and editing, have read and approved the final version, and agree to be accountable for all aspects of the work.
All animal experiments were conducted in accordance with the institutional guidelines of Shanghai Ocean University and complied with the European Union Directive 2010/63/EU on the protection of animals used for scientific purposes, as well as the ARRIVE 2.0 guidelines. The experimental protocol was reviewed and approved by the Animal Welfare and Ethical Review Body (AWERB) of Shanghai Ocean University (Approval No. SHOU-DW-2021-068). Ethical assessment of animal use was performed in accordance with Directive 2010/63/EU, which provides a regulatory framework for ethical review and risk assessment in scientific research involving animals. The study strictly adhered to the principles of Replacement, Reduction, and Refinement (3R). Experimental necessity was carefully evaluated during study design, procedures were optimized, and appropriate measures were implemented to minimize animal numbers as well as potential pain and distress.
The authors thank all individuals who contributed to the preparation of this manuscript. The authors also appreciate the constructive comments and valuable suggestions provided by the peer reviewers, which substantially improved the quality and clarity of this work.
This study was funded by National Natural Science Foundation of China Key Project (32130109) and Funds of Shanghai Sixth People’s Hospital (2017006).
The authors declare no conflict of interest.
Please refer to the supplementary materials (S1, S2, S3, S4, S5) for additional information. Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBL49658.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.