

 , Jing Zhou 1, Yuge Gao 1, Guangyu Yang 1, Yu Zhang 1, Siyu Hao 1, Yuzhen Li 1,*
, Jing Zhou 1, Yuge Gao 1, Guangyu Yang 1, Yu Zhang 1, Siyu Hao 1, Yuzhen Li 1,*
1 Department of Dermatology and Venereology, The Second Affiliated Hospital of Harbin Medical University, 150086 Harbin, Heilongjiang, China
Abstract
This study aimed to investigate the role of endoplasmic reticulum stress (ERS) in atopic dermatitis (AD) and its comorbid mental disorders and evaluate the therapeutic potential of mesencephalic astrocyte-derived neurotrophic factor (MANF).
Clinical samples from patients with AD and comorbidities were analyzed for ERS and apoptosis markers using quantitative polymerase chain reaction and western blotting. Mouse models of depression were developed using 28 days of chronic unpredictable mild stress (CUMS), and dermatitis was induced using 0.2% 2,4-dinitrofluorobenzene. Adenovirus-mediated MANF overexpression was induced via local injection to develop comorbidities, comorbidities + negative control overexpression, and comorbidities + MANF overexpression groups. Further, the ERS inhibitors of 4-phenylbutyric acid were employed for the comorbidities + inhibitors group. Dermatitis scoring, the sucrose preference test for behavioral changes, and ERS and apoptosis assessments using transmission electron microscopy, terminal deoxynucleotidyl transferase dUTP Nick-End labeling staining, hematoxylin and eosin staining, and immunohistochemistry were conducted.
Clinical data demonstrated significant increases in ERS-related markers (GRP78, P-IRE1, and Caspase-12) and apoptosis marker of Bax, whereas Bcl-2 was decreased in comorbidities compared with those with AD alone. In mouse models, increased Bax and decreased Bcl-2 indicated apoptotic pathway activation in the comorbidities group. MANF overexpression reduced ERS-related factors and skin pathology while improving behavioral indicators. Further, MANF reduced skin cell apoptosis by downregulating Bax and upregulating Bcl-2. ERS inhibitors similarly alleviated cell apoptosis, thereby confirming the critical role of ERS in disease progression.
MANF overexpression reduces ERS and apoptosis, which improves AD symptoms and comorbid mental disorders, thereby highlighting its therapeutic target potential.
Graphical Abstract
Keywords
- endoplasmic reticulum stress
- atopic dermatitis
- psychiatric disorders
- apoptosis
- comorbidities
The endoplasmic reticulum (ER) is a crucial organelle that is connected to the nuclear envelope, which is key for lipid synthesis, Ca2+ regulation, and protein processing in cells. Properly folded proteins are crucial for cellular functions, making the ER vital for quality control [1]. However, conditions, such as gene mutations, nutrient deprivation, hypoxia, and oxidative stress, trigger ER stress (ERS), leading to protein misfolding in the ER, toxin stimulation, Ca2+ metabolic imbalance, and sustained oxidative stress stimulation [2]. Further, cells that undergo severe ERS frequently cause cell death through various pathways (e.g., apoptosis), causing significant harm to the organism [3]. To survive, cells trigger self-protection events, including upregulating ERS-related proteins PERK, IRE1, and ATF6 to refold misfolded or unfolded proteins and inhibiting ERS-induced pro-apoptosis proteins, including caspase and Bax proteins [4]. A substantial body of research indicates that ERS is closely associated with several diseases, particularly inflammation, neurodegenerative condition progression, and metabolic syndrome [5].
Atopic dermatitis (AD) is a chronic, recurring, non-infectious skin inflammation disorder characterized by continuous skin itching and eczema-like symptoms, including redness and papules, which differ in terms of age and skin dryness [6]. Chronic inflammation and scratching cause skin thickening and lichenification, and persistent itching disrupts daily activities and sleep, reduces quality of life, and leads to serious mental disorder complications (e.g., depression) and autoimmune diseases [7]. Atopic dermatitis is frequently comorbid with depression. The brain-skin axis mediates this comorbidity, and the combined chronic unpredictable mild stress (CUMS) + 2,4-dinitrofluorobenzene (DNFB) model is a validated one that recapitulates skin inflammation, pruritus, and depressive-like behaviors [8, 9]. AD involves complex interactions among genetic abnormalities, altered immune responses, epidermal barrier defects, and disrupted skin microbiome, all contributing to its complexity [10]. Recent studies have demonstrated that sebaceous glands produce Th2-related mediators via interleukin-4 receptors, a process regulated by galectin-12, which also inhibits ERS and promotes salivary gland enlargement and AD-like phenotype, thereby implying the crucial role of ERS in AD progression [11]. Previous studies have revealed that ERS plays a crucial role in regulating immune responses and skin barrier function. For instance, pathway activation during ERS leads to proteolytic cleavage of precursor ATF6 in the Golgi apparatus, thereby producing an active ATF6 fragment that translocates into the nucleus [12]. In murine models of psychiatric disorders, IRE1 pathway activation under ERS conditions promotes XBP1 mRNA splicing via its endoribonuclease activity, which generates the active transcription factor XBP1s, thereby upregulating the expression of inflammation-related genes [13]. This evidence indicated that ERS helped in AD development and progression and played a role in its mental disorder complications.
Mesencephalic astrocyte-derived neurotrophic factor (MANF), a neurotrophic factor that supports the survival of midbrain dopaminergic neurons, has been upregulated and secreted in response to ERS [14]. MANF is involved in ERS-related neurological disorders, and shows its potential as a novel research target for ERS-associated metabolic diseases [15]. Recent studies have identified key AD molecular targets and therapies, including natural ingredients regulating inflammatory/oxidative stress and endothelin receptor antagonists with immunomodulatory/anti-inflammatory effects [16, 17]. However, the role of MANF in AD and psychiatric disorder comorbidities remains largely unexplored, and its precise molecular mechanisms are unclear. This study investigates ERS in AD and mental comorbidities and evaluates the regulatory potential of MANF.
Clinical data of patients with AD alone or AD comorbid with mental disorders—including 6 patients in the AD-alone group and 6 patients in the AD-comorbid mental disorders group—were collected from the Second Affiliated Hospital of Harbin Medical University from July 2024 to November 2024 to investigate the features of ERS and inflammation via quantitative polymerase chain reaction (qPCR) and western blotting. The baseline clinical characteristics of all enrolled patients, including gender, age, AD disease duration, and disease severity scores (SCORAD, EASI, IGA, VAS) as well as HADS anxiety and depression scores, are summarized in Supplementary Table 1. This study was conducted following the Declaration of Helsinki, and all participants signed written informed consent.
Diagnostic Criteria for AD: Diagnosis was based on the Chinese Guidelines for the Diagnosis and Treatment of Atopic Dermatitis, requiring pruritus as a core symptom plus at least two of the following: flexural eczematous lesions, personal or familial history of atopic diseases (e.g., asthma, allergic rhinitis), increased serum total IgE or peripheral blood eosinophil percentage, and history of skin dryness.
Diagnostic Criteria for Comorbid Psychiatric Disorders: Two attending
psychiatrists made the diagnosis based on the International Classification of
Diseases, 11th Revision (ICD-11), including major depressive disorder (Hamilton
Depression Rating Scale [HAMD-17] score
Exclusion criteria included comorbidity with autoimmune skin diseases or infectious skin diseases; use of systemic glucocorticoids, immunosuppressants, or psychotropic drugs within 1 month before enrollment; presence of severe hepatic, renal, or cardiovascular diseases or malignancy; pregnancy or lactation; and inability to cooperate with sample collection.
Healthy controls were enrolled if they reported no history of skin or
psychiatric diseases, had not used immunomodulatory, psychotropic, or
glucocorticoid drugs in the past 3 months, and had normal indicators, including
liver and renal function. A 3 mm
The university ethics committee approved the in vivo protocol design and procedures. In this study, 4–6-week-old specific-pathogen-free BALB/c mice (Vital River, SPF level) of both sexes were randomly allocated, with 8 mice allocated to each experimental group to ensure statistical validity. Mice were randomly assigned to each experimental group using a random number table method to eliminate grouping bias. A depression model was developed via 28 days of chronic unpredictable mild stress (CUMS) [18], comprising a series of stressors applied randomly. The stressors were horizontal shaking, tilting of the cage, food and water deprivation, tail clipping, heat exposure, and cold water swimming. Each stressor was applied once per day, and the sequence was varied to prevent habituation to the stressors. The AD model was developed on day 14 of the depression model by applying a 0.5% 2,4-dinitrofluorobenzene (DNFB) (Sigma-Aldrich, D9136) solution for sensitization. This was followed by 0.2% DNFB applications on days 5, 8, and 14 to induce dermatitis. Adenovirus-mediated MANF overexpression (negative control overexpression [oeNC] and MANF overexpression [oeMANF]) was performed via subcutaneous injection of 100 µL viral solution at the dermatitis site in the established depression model, establishing comorbidities, comorbidities + oeNC, and comorbidities + oeMANF groups. Further, the 4-phenylbutyric acid (4-PBA) as an ERS inhibitor (500 mg/kg) was administered at two separate dosages for the AD, comorbidities, and comorbidities+inhibitors [18] groups. The animal experiment was conducted from November 2024 to January 2025, with a total duration of 3 months. The CUMS-induced depression model lasted 28 days, followed by 21 days of AD induction and intervention, with behavioral and pathological assessments conducted during the final 7 days. At the end of the animal experiment, all mice were euthanized via intraperitoneal injection of sodium pentobarbital (100 mg/kg, 50 mg/mL, Sigma-Aldrich). The skin and brain tissue were dissected within 10 min for tissue collection after euthanasia to maintain tissue integrity.
The total number of scratching bouts was counted and utilized to quantify pruritus severity. Dermatitis severity was assessed using a previously established scoring system, including five key parameters [19]: (1) edema/papules; (2) exudate/scabs; (3) epidermal erosion; (4) lichenoid change; and (5) dryness of the unaffected skin. Each item was scored on a scale (typically 0–3) according to severity, with the total dermatitis score obtained by summing the scores of all five parameters. Each parameter was scored on a scale from 0 to 3, indicating no signs or symptoms, mild manifestations, moderate severity, and severe manifestations, respectively.
The sucrose preference test (SPT) assesses an animal’s emotional state by measuring its preference for sweet water, including a sucrose solution. Before testing, mice were given free access to both 1% sucrose solution and tap water. After 12 h of food and water deprivation, mice were provided with preweighed bottles containing sucrose solution and water for a 2-h testing period [20]. Fluid consumption was recorded, and the SPT was calculated utilizing the following formula:
Total RNA was extracted from treated cells or tissues using TRIzol reagent
(Invitrogen, 5596026), and cDNA was synthesized with a Transcriptor First Strand
cDNA Synthesis Kit (Invitrogen, N8080234, CA, USA). qPCR was performed on an ABI
Step-One Plus real-time PCR system using LightCycler 480 SYBR Green I Master Mix
(azyme, SGD2203, Shanghai, China). Relative RNA expression levels of
GRP78, P-IRE1, Caspase-12, Bax,
Bcl-2, ATF4, CHOP, and P-EIF-2
Total proteins were extracted from cultured cells using RIPA lysis buffer
(Sigma-Aldrich, 20-188, St. Louis, MO, USA). Protein concentration was determined
using a BCA protein assay kit (Pierce™, Thermo Fisher Scientific,
Waltham, MA, USA). Equal amounts of protein (30 µg) were separated by
SDS-PAGE (Bio-Rad, Hercules, CA, USA) with protein markers (Beyotime, P0071,
Shanghai, China; Yeason, 20350ES72, Shanghai, China), then transferred onto
polyvinylidene difluoride (PVDF) membranes. After blocking with 5% nonfat milk
in TBST, membranes were incubated overnight at 4 °C with primary
antibodies: GRP78 (Proteintech, 10768-1-AP, Rosemont, IL, USA), P-IRE1 (CST,
3294S, Danvers, MA, USA), Bax (CST, 2772S, Danvers, MA, USA), Bcl-2 (CST, 2876S,
Danvers, MA, USA), ATF4 (CST, 11815S, Danvers, MA, USA), CHOP (CST, 2895S,
Danvers, MA, USA), p-eIF2
Mice tissue samples were washed in PBS (Beyotime, ST476, Shanghai, China) for 30 min and then dehydrated through graded ethanol (Sigma-Aldrich, 459836-100ML). The tissues were cleared in xylene (Sigma-Aldrich, 339056, St. Louis, MO, USA), and stored at –20 °C. After sectioning (4 µm), sections were treated with 3% hydrogen peroxide (Sigma-Aldrich, 88597, St. Louis, MO, USA) for 10 min, followed by antigen retrieval in citrate buffer (Sigma-Aldrich, C9999, St. Louis, MO, USA) and blocking with 3% bovine serum albumin (Servicebio, GC305010-50g, Wuhan, China). Primary antibodies against Caspase-12 (Abcam, ab23352, Cambridge, UK), GRP78 (Cell Signaling Technology, 3183, Danvers, MA, USA), and P-IRE1 (Abcam, ab124945, Cambridge, UK) were incubated overnight at 4 °C. After washing, sections were incubated with secondary antibody (Cell Signaling Technology, 7074, Danvers, MA, USA) for 30 min at 37 °C. Further, 3,3’-diaminobenzidine (Sigma-Aldrich, D4293, St. Louis, MO, USA) was used for color development, followed by counterstaining with Mayer’s hematoxylin (Abcam, ab220365, Cambridge, UK). Sections were dehydrated, cleared in xylene, and mounted with neutral balsam (Sigma-Aldrich, 36406, St. Louis, MO, USA).
The HE staining kit (Vectorlabs, H-3502, Burlingame, CA, USA) was used for HE staining. The fixative volume was 4–10 times the tissue volume. After fixation and dehydration, sections were trimmed, placed face down in labeled embedding cassettes, and redehydrated. Tissues were then embedded, cut into 5-µm sections, and stained with Harris HE.
The In Situ Cell Death Detection Kit, Fluorescein, which is TUNEL technology, POD (Roche, Basel, Switzerland), was performed on sections from three animals per group for tissue injury analysis in the dermatitis site. Tissues were directly analyzed under a fluorescence microscope with an excitation wavelength of 530 nm and a detection wavelength of 630 nm green.
Paraffin sections were successively immersed in xylene I and II (Selleck, 2650-17-1, Houston, TX, USA) for 10 min each. Dehydration was performed through a series of ethanol (Sigma-Aldrich, 459836-100ML) concentrations, washed with PBS (Beyotime, ST476, Shanghai, China), and stained in toluidine blue (Selleck, 92-31-9, Houston, TX, USA), followed by washing in tap water for 2 min to remove excess dye. Differentiation was performed in 95% ethanol, followed by graded ethanol dehydration, xylene clearing, and mounting.
The samples were fixed overnight at 4 °C using PBS (Beyotime, ST476, Shanghai, China) and then washed four times with PBS. Subsequently, the samples were fixed for 2 h at 4 °C using 1% osmium tetroxide solution (Sigma-Aldrich, 251755, St. Louis, MO, USA) and then washed four times with PBS. The samples were dehydrated using an ethanol gradient. After centrifugation at 2000 g for 10 min, bacterial pellets were fixed in 2% glutaraldehyde in 0.1 M sodium cacodylate buffer (pH 7.4), embedded in 2% agarose, postfixed in 1% osmium tetroxide at 25 °C overnight, dehydrated with gradient ethanol, and embedded in Durcupan resin (Sigma-Aldrich, St. Louis, MO, USA). 55-nm sections were examined using a JEM-1200 transmission electron microscope (JEOL, Tokyo, Japan) equipped with a 4-K Eagle digital camera (FEI, Hillsboro, OR, USA).
Mice were individually placed in a water tank (24
Mice were suspended vertically by the distal 1 cm of the tail for 5 min. The immobility time was quantified, defined as the absence of active struggling movements.
Mice were placed in the central zone of the EPM (50 cm height) facing a closed arm and allowed to explore freely for 5 min. The percentage of time spent in the open arms was calculated as the primary index of anxiety-like behavior.
Data are presented as mean
The ERS gene expression in clinical patients between the AD and comorbidities
groups was assessed, revealing significantly increased GRP78, P-IRE1,
and Caspase-12 expressions in the comorbidities group (p
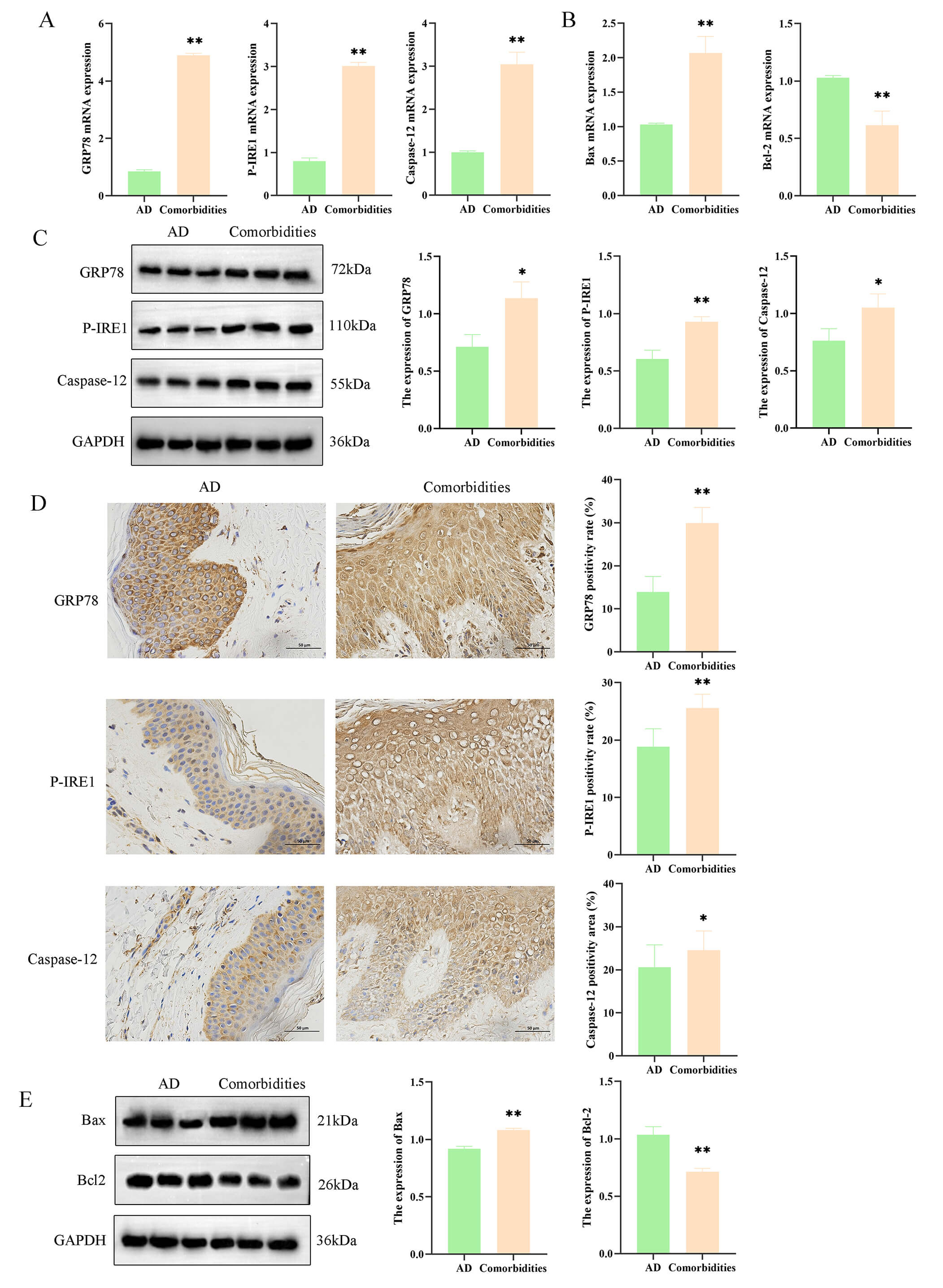 Fig. 1.
Fig. 1.
Endoplasmic reticulum stress and apoptosis are increased in
atopic dermatitis (AD) patients with psychiatric comorbidities. (A) mRNA levels of GRP78, P-IRE1, and Caspase-12 in skin
tissues from AD and comorbid AD patients. (B) mRNA levels of
Bax and Bcl-2 in the two patient groups. (C) Protein
levels of GRP78, P-IRE1, and Caspase-12 detected by Western blot. (D) Immunohistochemistry of ER stress markers (scale bar = 50 µm) and
quantitative analysis. (E) Protein levels of Bax and Bcl-2. Data are
mean
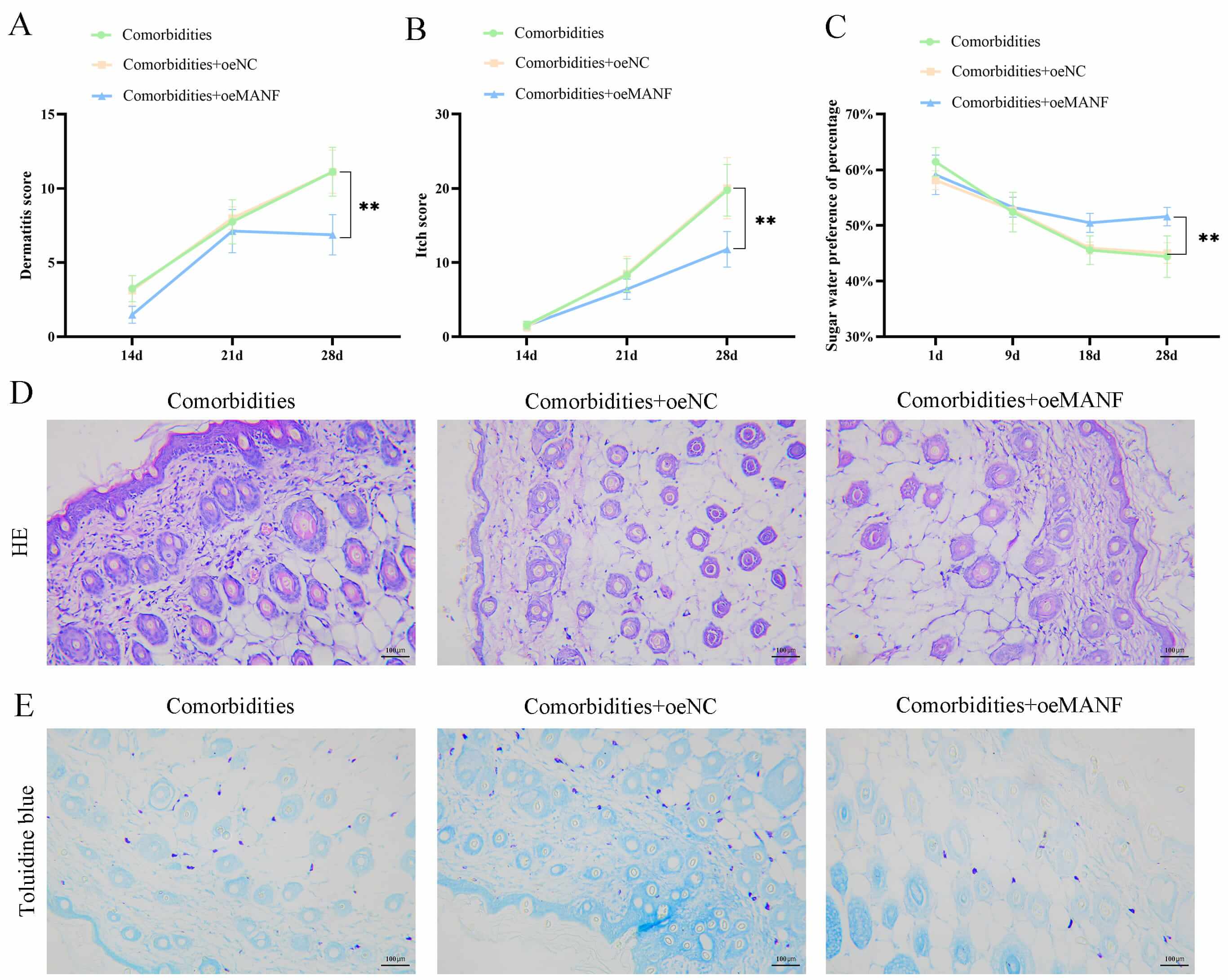
Subsequently, the behavioral and dermatopathological alterations were assessed
in the comorbidities, comorbidities + oeNC, and comorbidities + oeMANF groups.
The SPT revealed that, by day 28, the behavioral dermatitis scoring presented
that the comorbidities + oeMANF group demonstrated milder dermatitis symptoms,
with significantly lower dermatitis scores compared with the comorbidities group
(p
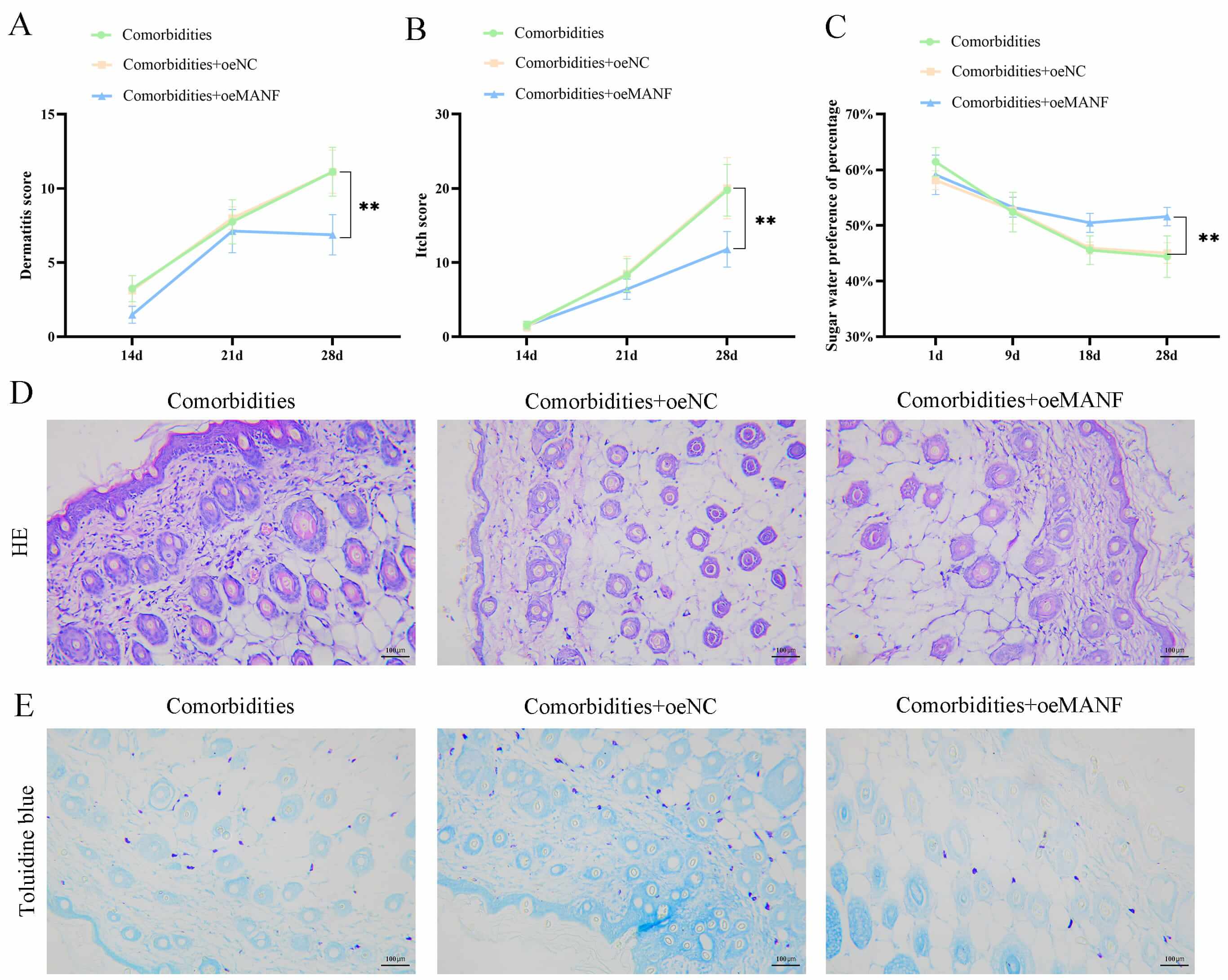 Fig. 2.
Fig. 2.
Mesencephalic astrocyte-derived neurotrophic factor (MANF)
overexpression improved behavior and skin lesions in comorbidities. (A) Dermatitis severity scores in comorbid, comorbid + negative control
overexpression (oeNC), and comorbid + MANF overexpression (oeMANF) mice.
(B) Scratching bouts in each group. (C) Sucrose preference test
for depressive-like behaviors. (D) Hematoxylin and eosin (HE)
staining of mouse skin tissues (scale bar = 100 µm). (E) Toluidine
blue staining for mast cell infiltration (scale bar = 100 µm). Data are
mean
| Condition | Edema/Papules | Exudate/Scabs | Epidermal erosion | Lichenoid change | Dryness (unaffected skin) |
| Comorbidity 1 | 2 | 3 | 3 | 2 | 3 |
| Comorbidity 2 | 2 | 3 | 3 | 2 | 2 |
| Comorbidity 3 | 2 | 3 | 2 | 2 | 2 |
| Comorbidity 4 | 2 | 2 | 2 | 2 | 2 |
| Comorbidity 5 | 3 | 3 | 2 | 3 | 2 |
| Comorbidity 6 | 2 | 2 | 2 | 3 | 2 |
| Comorbidity 7 | 2 | 2 | 3 | 2 | 2 |
| Comorbidity 8 | 2 | 2 | 2 | 1 | 1 |
| Comorbidity + oeNC1 | 2 | 2 | 3 | 2 | 2 |
| Comorbidity + oeNC2 | 2 | 3 | 3 | 3 | 2 |
| Comorbidity + oeNC3 | 2 | 3 | 3 | 3 | 2 |
| Comorbidity + oeNC4 | 2 | 2 | 3 | 2 | 2 |
| Comorbidity + oeNC5 | 2 | 2 | 2 | 2 | 2 |
| Comorbidity + oeNC6 | 2 | 2 | 2 | 1 | 2 |
| Comorbidity + oeNC7 | 2 | 3 | 3 | 2 | 2 |
| Comorbidity + oeNC8 | 2 | 2 | 2 | 2 | 2 |
| Comorbidity + oeMANF1 | 2 | 2 | 2 | 2 | 1 |
| Comorbidity + oeMANF2 | 1 | 2 | 2 | 1 | 1 |
| Comorbidity + oeMANF3 | 1 | 1 | 2 | 1 | 1 |
| Comorbidity + oeMANF4 | 1 | 2 | 2 | 2 | 1 |
| Comorbidity + oeMANF5 | 1 | 1 | 2 | 1 | 1 |
| Comorbidity + oeMANF6 | 2 | 2 | 2 | 1 | 1 |
| Comorbidity + oeMANF7 | 1 | 1 | 1 | 1 | 1 |
| Comorbidity + oeMANF8 | 1 | 1 | 2 | 1 | 1 |
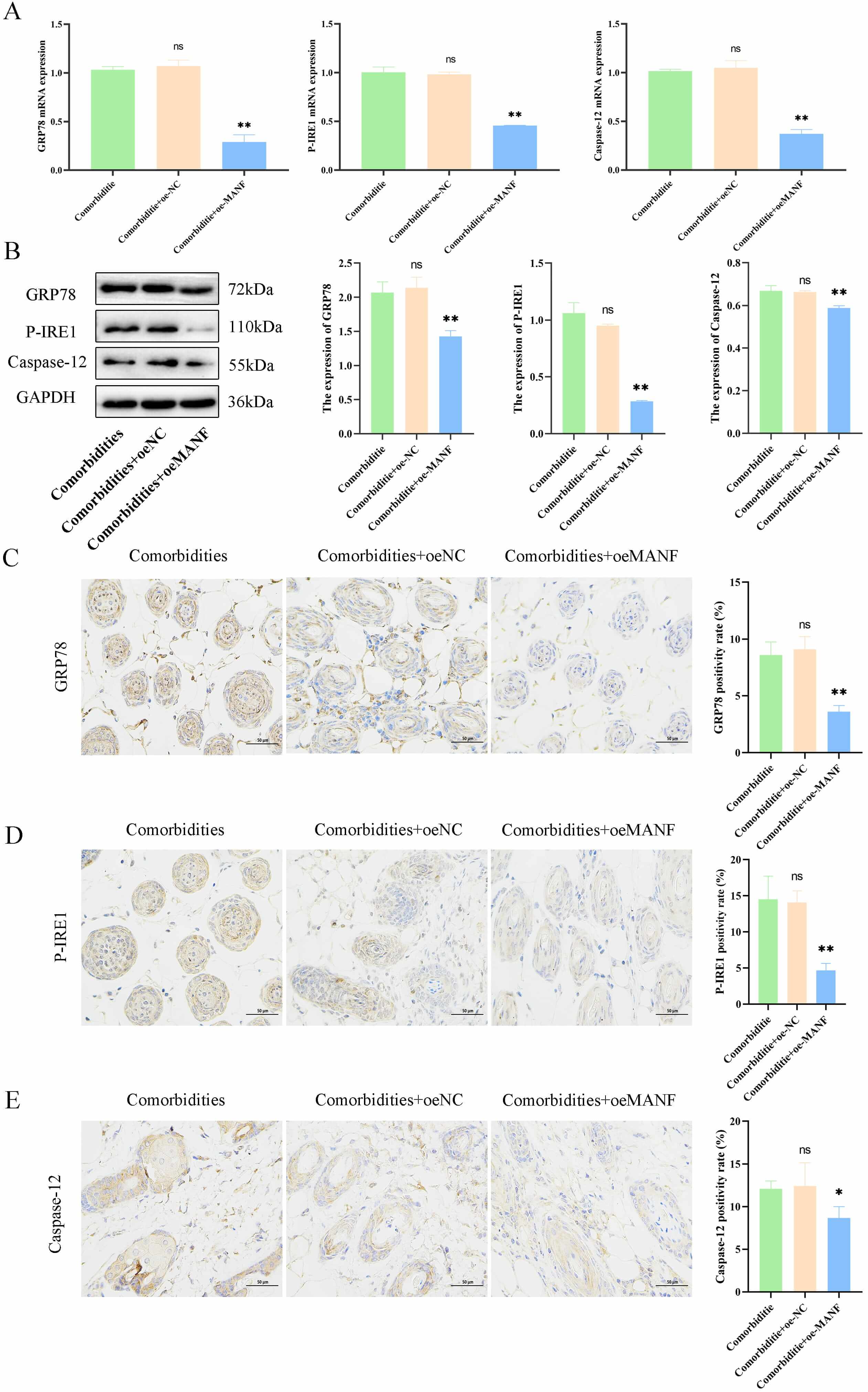
The underlying regulatory role of MANF in skin changes in comorbid mice under
ERS and its associated pathways were explored. The oeMANF significantly reduced
the expression of ERS-related factors GRP78, P-IRE1, and
Caspase-12 at both mRNA and protein levels in an AD mouse model with
comorbid mental disorders (comorbidities) (p
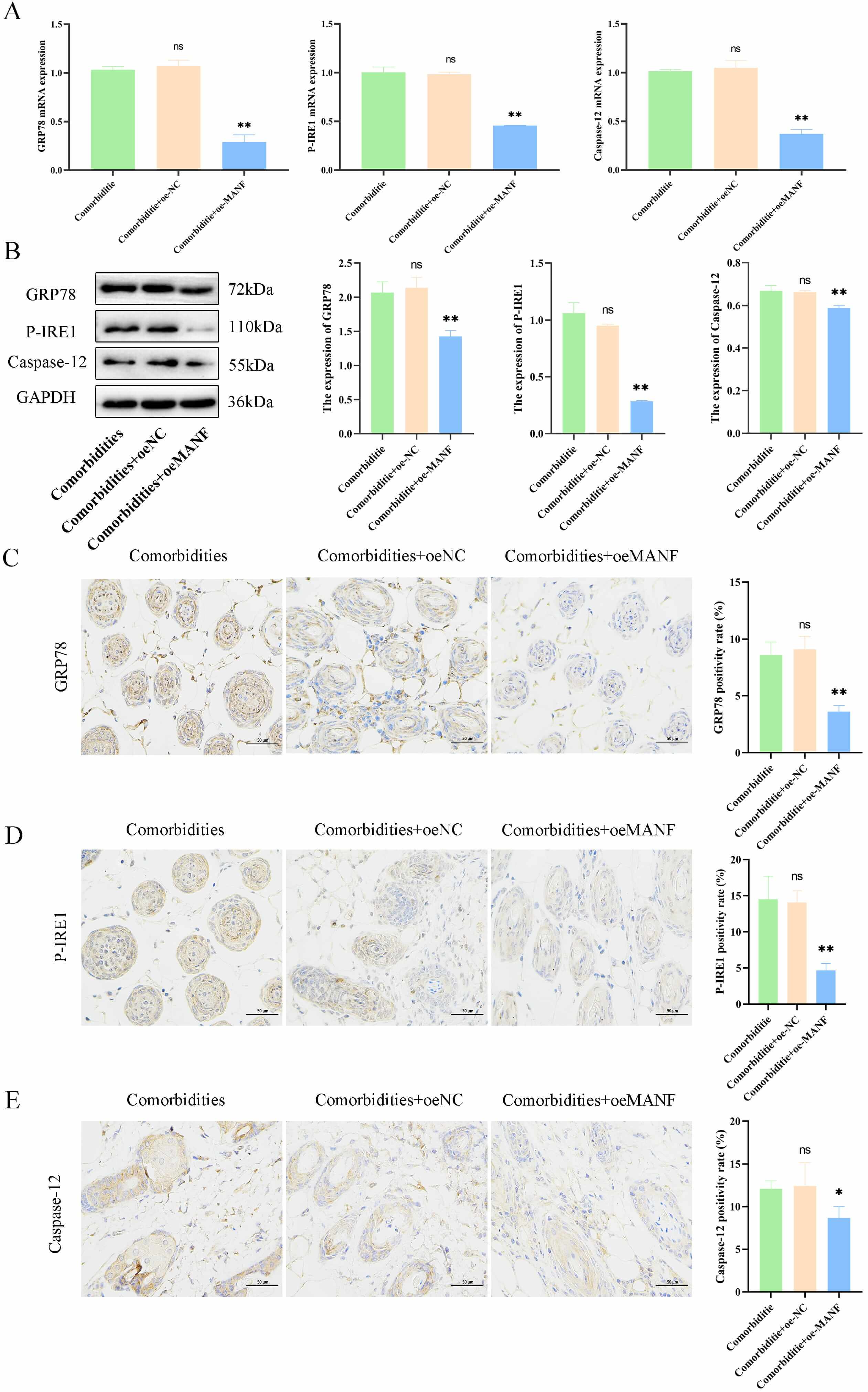 Fig. 3.
Fig. 3.
MANF overexpression alleviated endoplasmic reticulum stress
(ERS) and improved pathology in comorbidities models. (A) mRNA levels of
GRP78, P-IRE1, and Caspase-12 in mouse skin. (B)
Protein levels of ER stress-related markers. (C–E) Immunohistochemistry and
quantification of GRP78, P-IRE1, and Caspase-12 (scale bar = 50 µm). Data
are mean
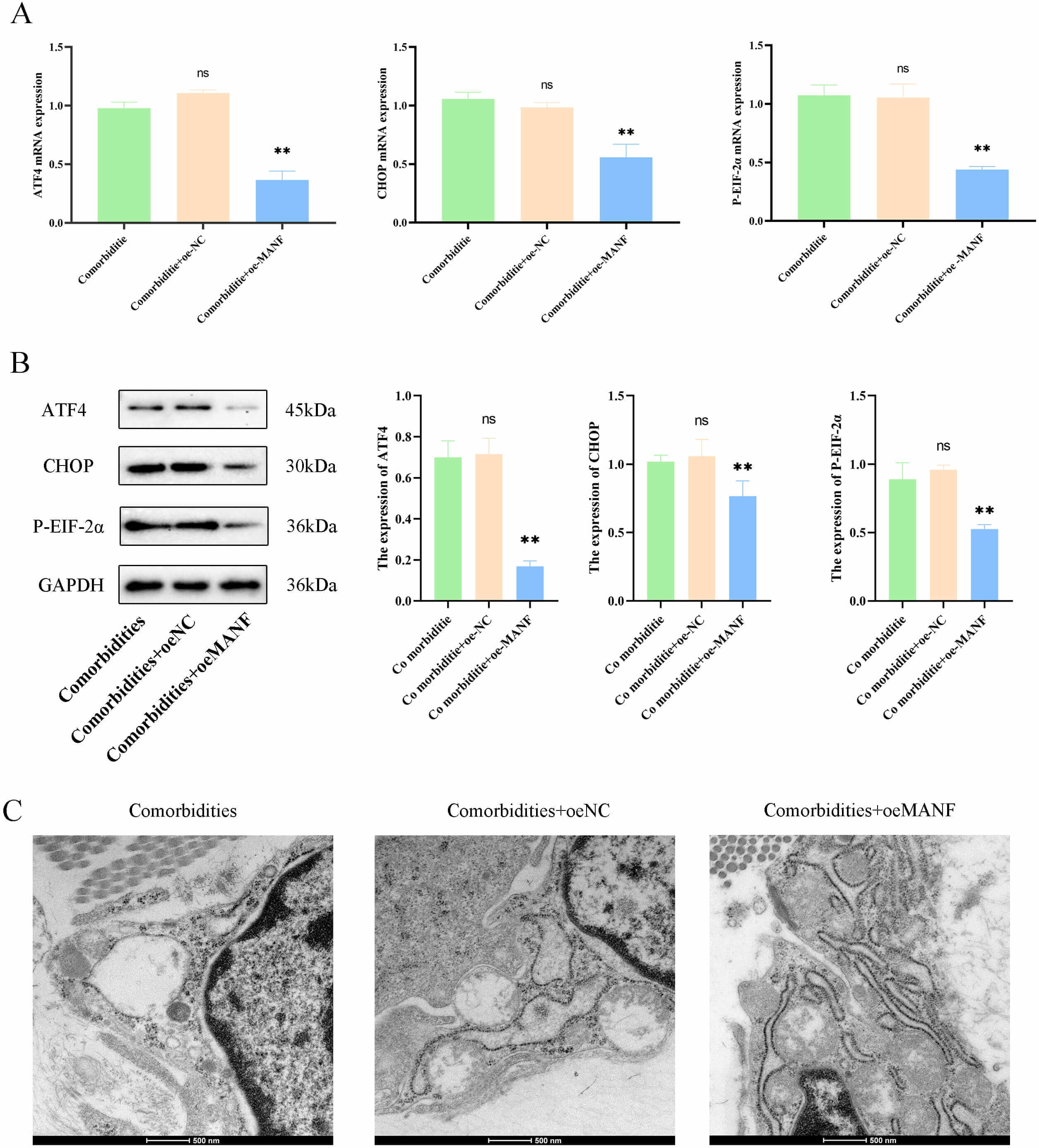
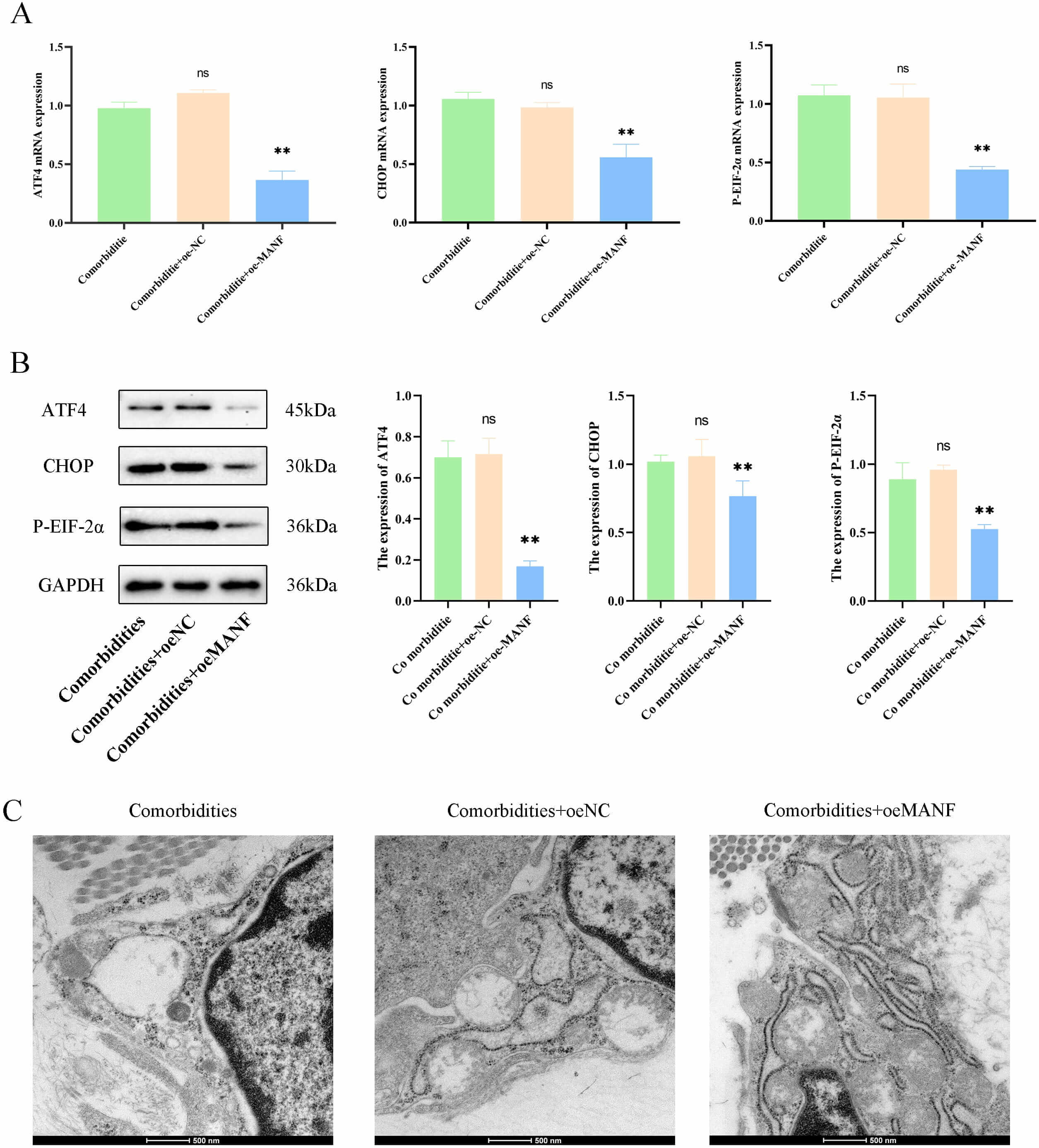 Fig. 4.
Fig. 4.
MANF overexpression alleviated ERS and improved pathology in
comorbidities models. (A) mRNA levels of ERS marker protein (ATF4,
CHOP, and P-eIF2
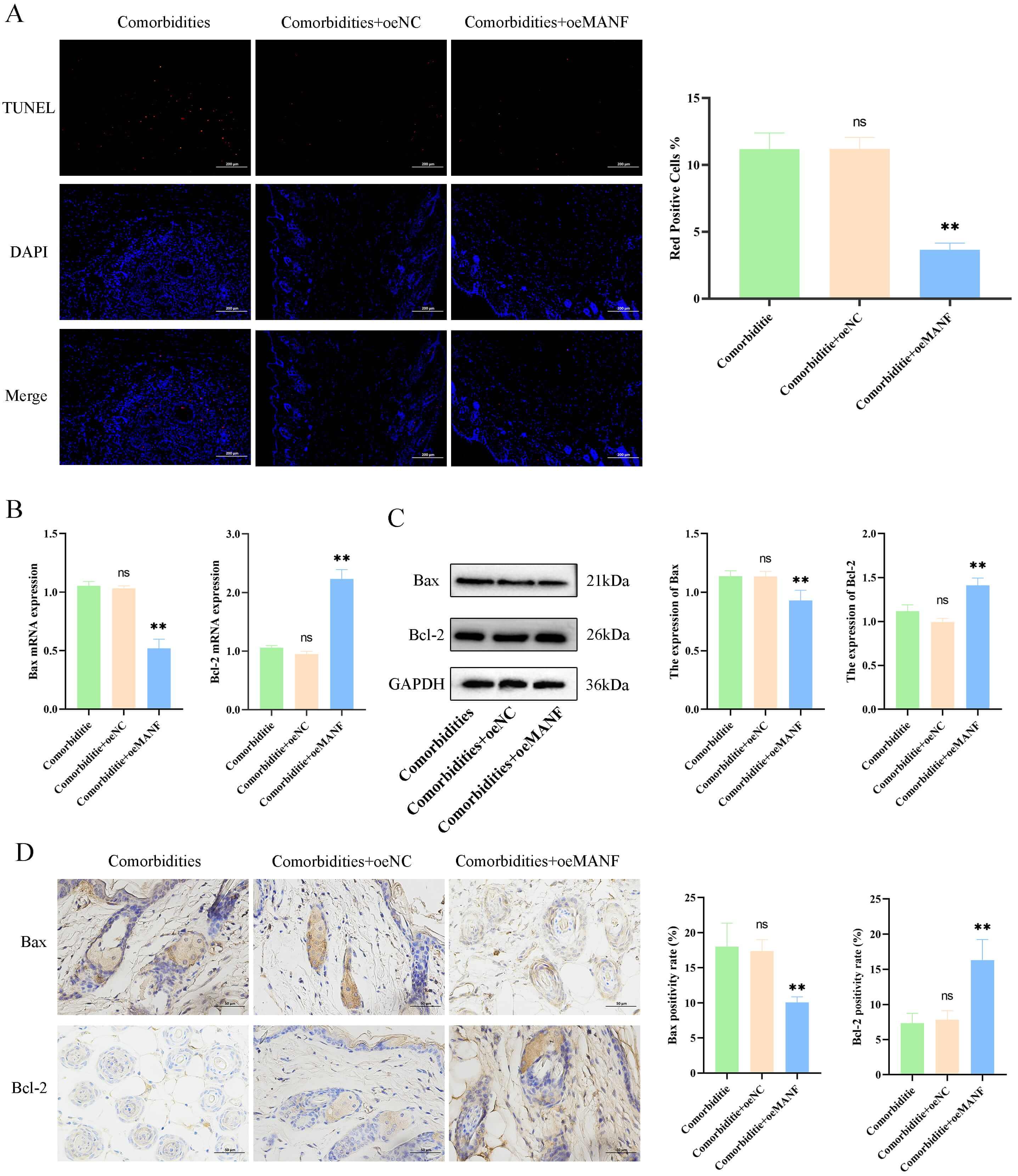
Further analysis of apoptosis in mouse skin tissue revealed significantly lower
skin apoptosis levels in the comorbidities + oeMANF group than in the
comorbidities group (p
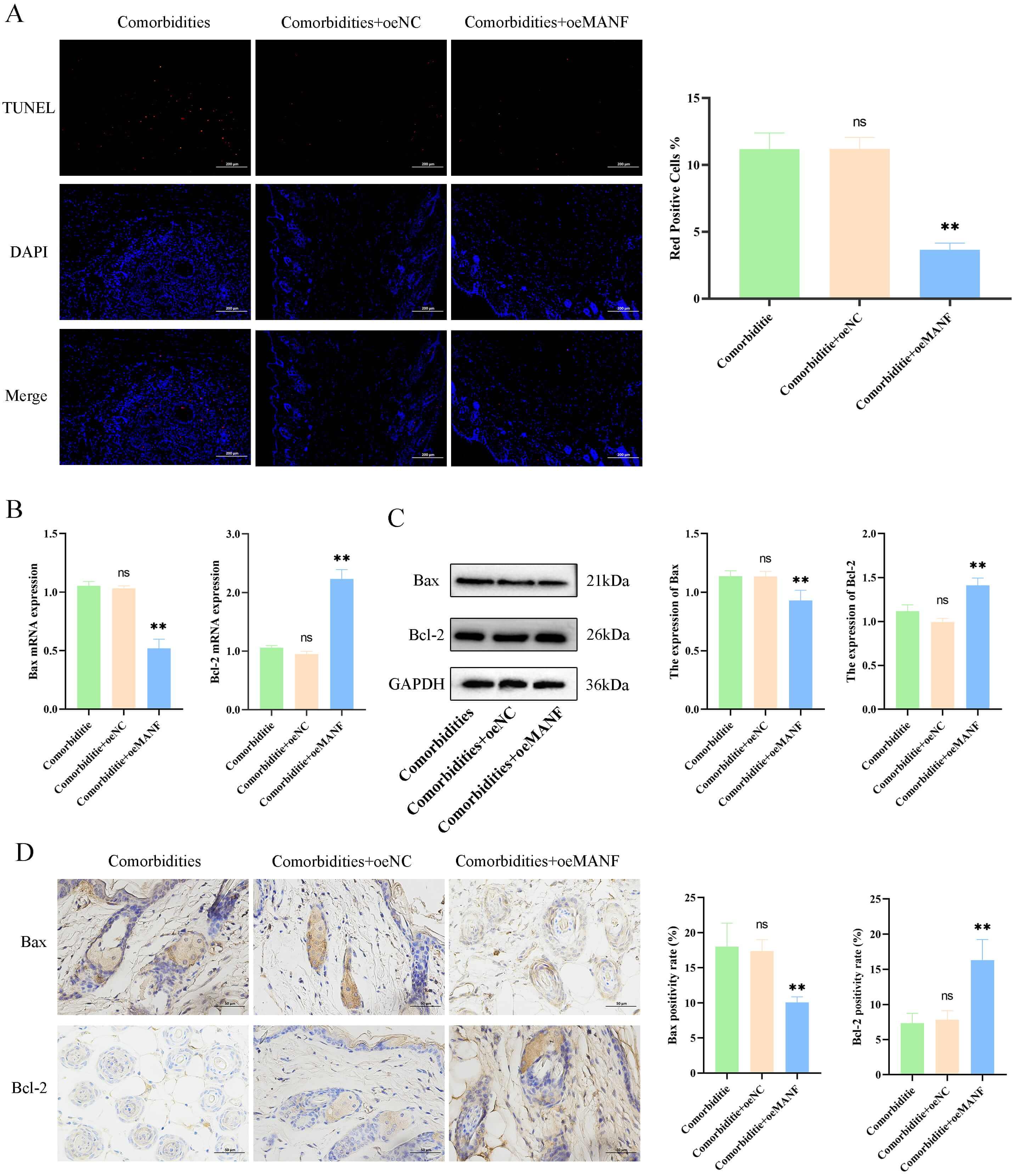 Fig. 5.
Fig. 5.
MANF overexpression reduces skin cell apoptosis in comorbid
mice. (A) Terminal deoxynucleotidyl transferase dUTP Nick-End labeling
(TUNEL) staining and apoptotic index (scale bar = 200 µm). (B) mRNA levels of Bax and Bcl-2. (C) Protein levels of
Bax and Bcl-2. (D) Immunohistochemistry and quantification of Bax and
Bcl-2 (scale bar = 50 µm). Data are mean
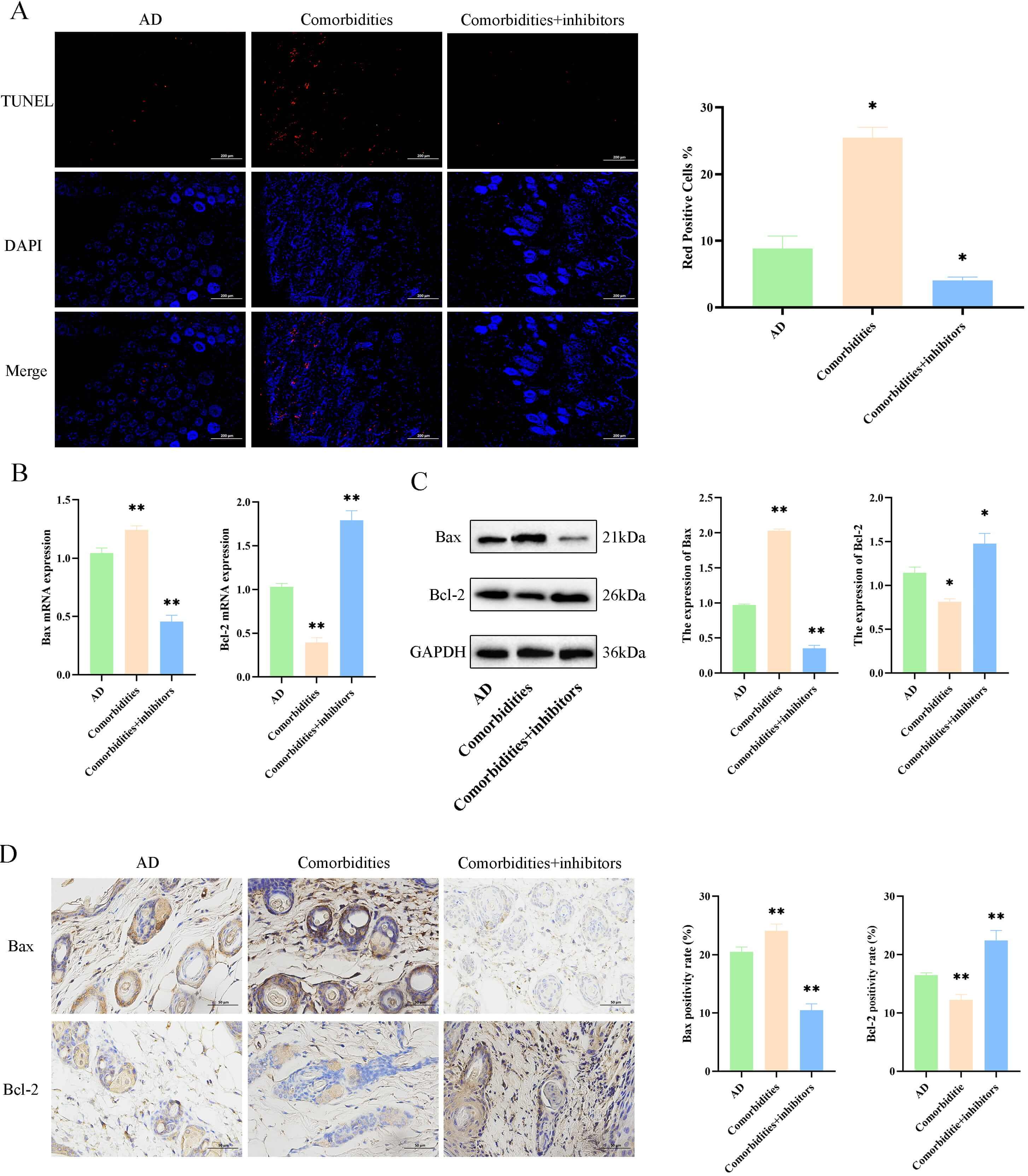
To investigate the regulatory effects of ERS on the apoptotic pathway proteins
in the skin tissues of comorbid mice, we first used the TUNEL staining to observe
skin tissue apoptosis. Skin apoptosis levels were higher in the comorbidities
group compared with the AD group (p
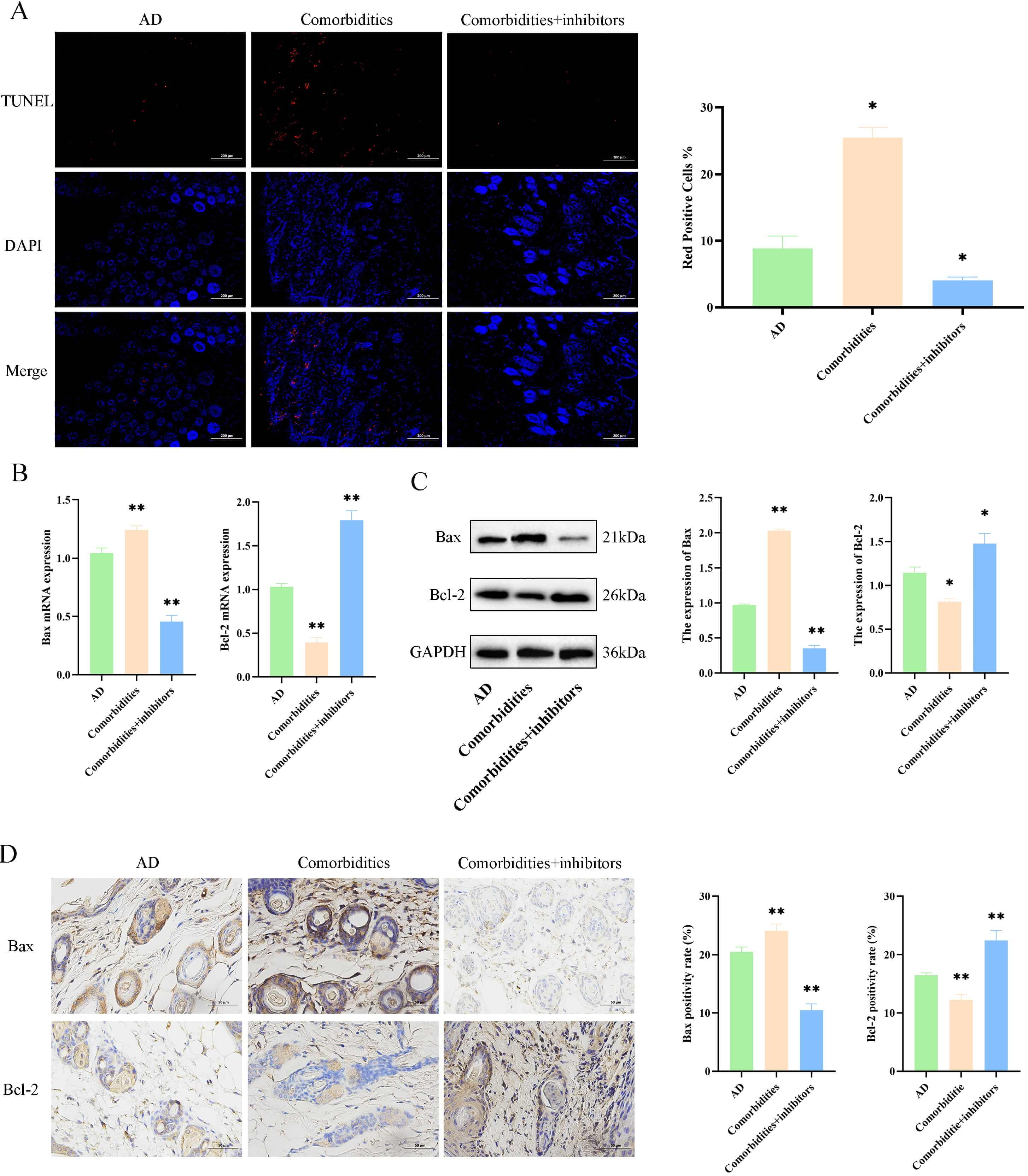 Fig. 6.
Fig. 6.
4-phenylbutyric acid (4-PBA)-mediated inhibition of ER stress
reduces apoptosis in comorbid mice. (A) TUNEL staining and apoptotic index
(scale bar = 200 µm). (B) mRNA levels of Bax and
Bcl-2. (C) Protein levels of Bax and Bcl-2. (D) Immunohistochemistry and quantification of Bax and Bcl-2 (scale bar = 50
µm). Data are mean
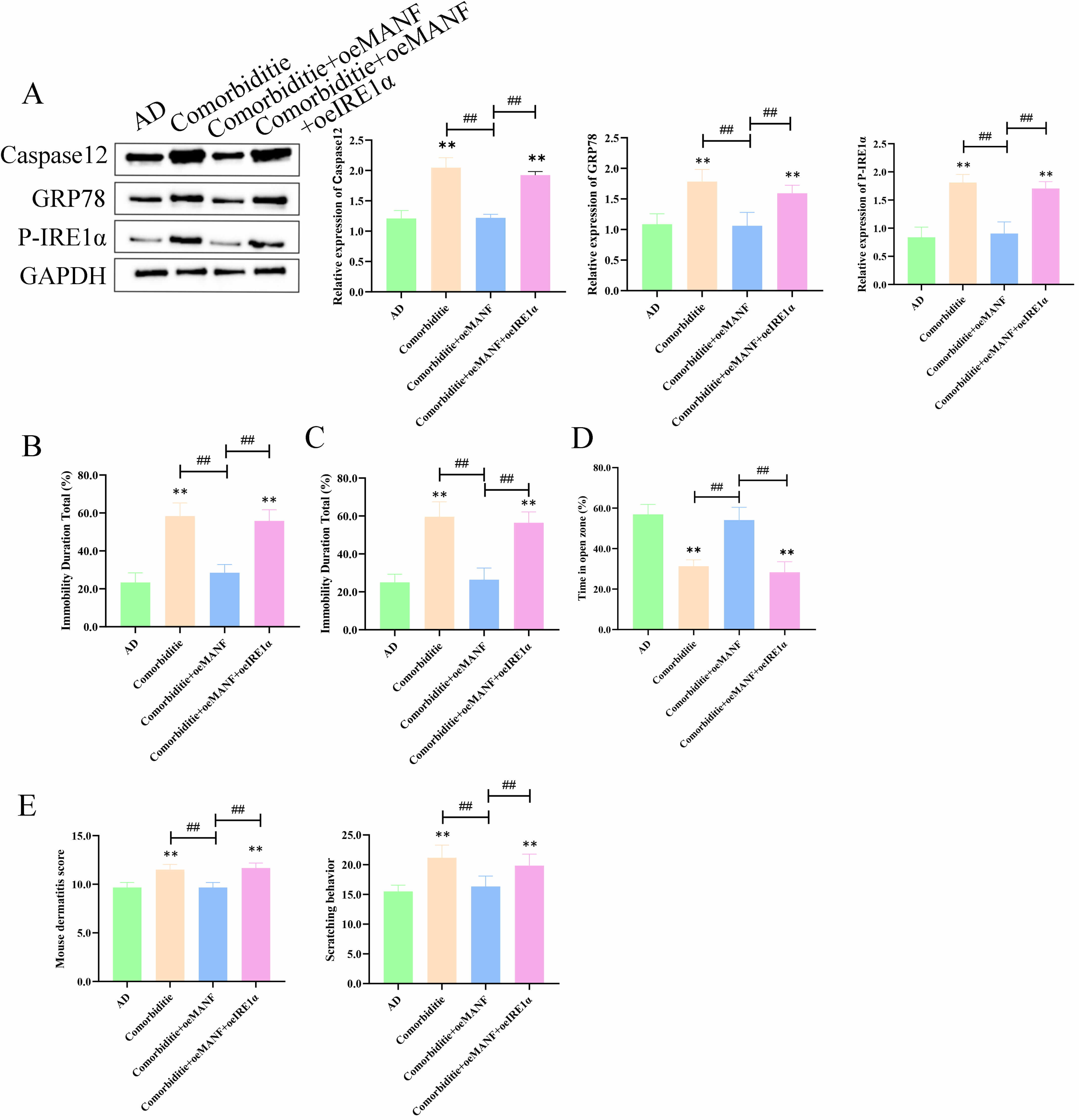
To explore how MANF regulates ER stress and apoptosis in skin and brain tissues,
rescue experiments were performed in four groups: AD, Comorbidity, Comorbidity +
oeMANF, and Comorbidity + oeMANF + oeIRE1
In skin tissues, the Comorbidity group showed markedly upregulated caspase‑12,
GRP78, and p-IRE1
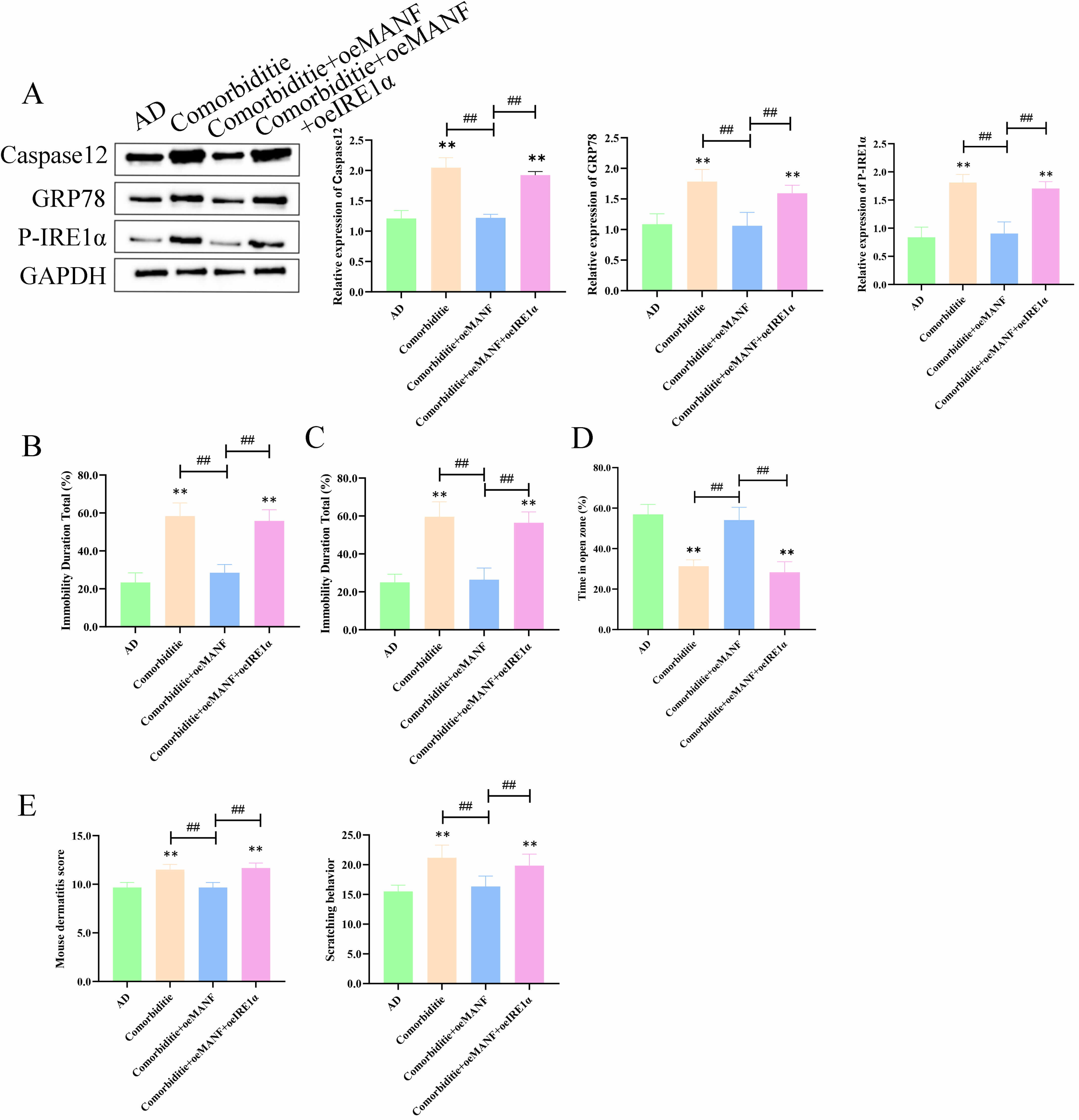 Fig. 7.
Fig. 7.
MANF suppresses IRE1
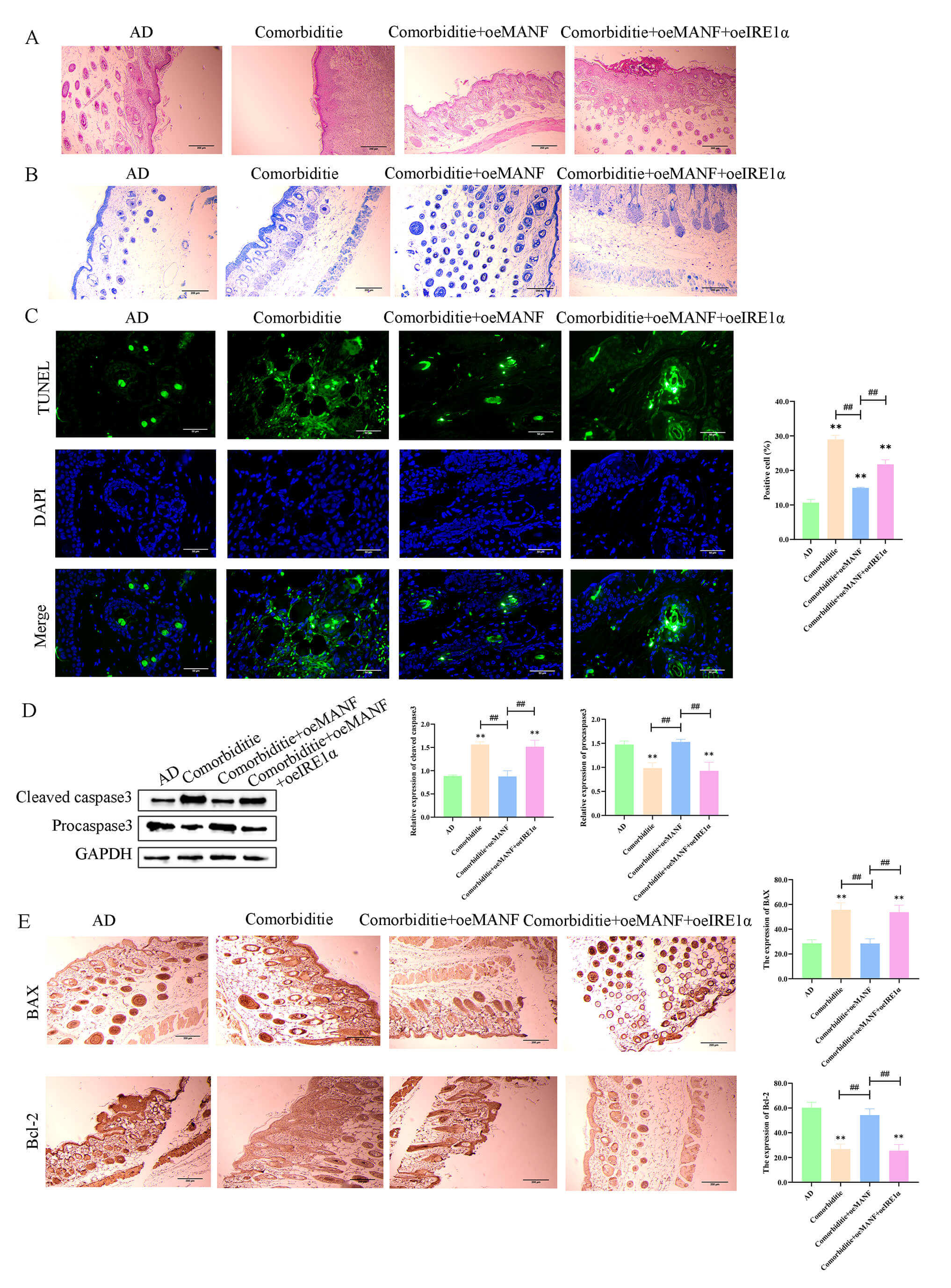
In skin tissues, the Comorbidity group displayed severe morphological injury,
increased mast cell accumulation, and elevated apoptosis (Fig. 8A–C). MANF
overexpression alleviated these pathological changes. Meanwhile, the ratios of
cleaved caspase‑3/pro caspase‑3 (Fig. 8D) and Bax/Bcl‑2 (Fig. 8E) were increased
in the Comorbidity group; MANF overexpression normalized these ratios, and these
effects were blocked by IRE1
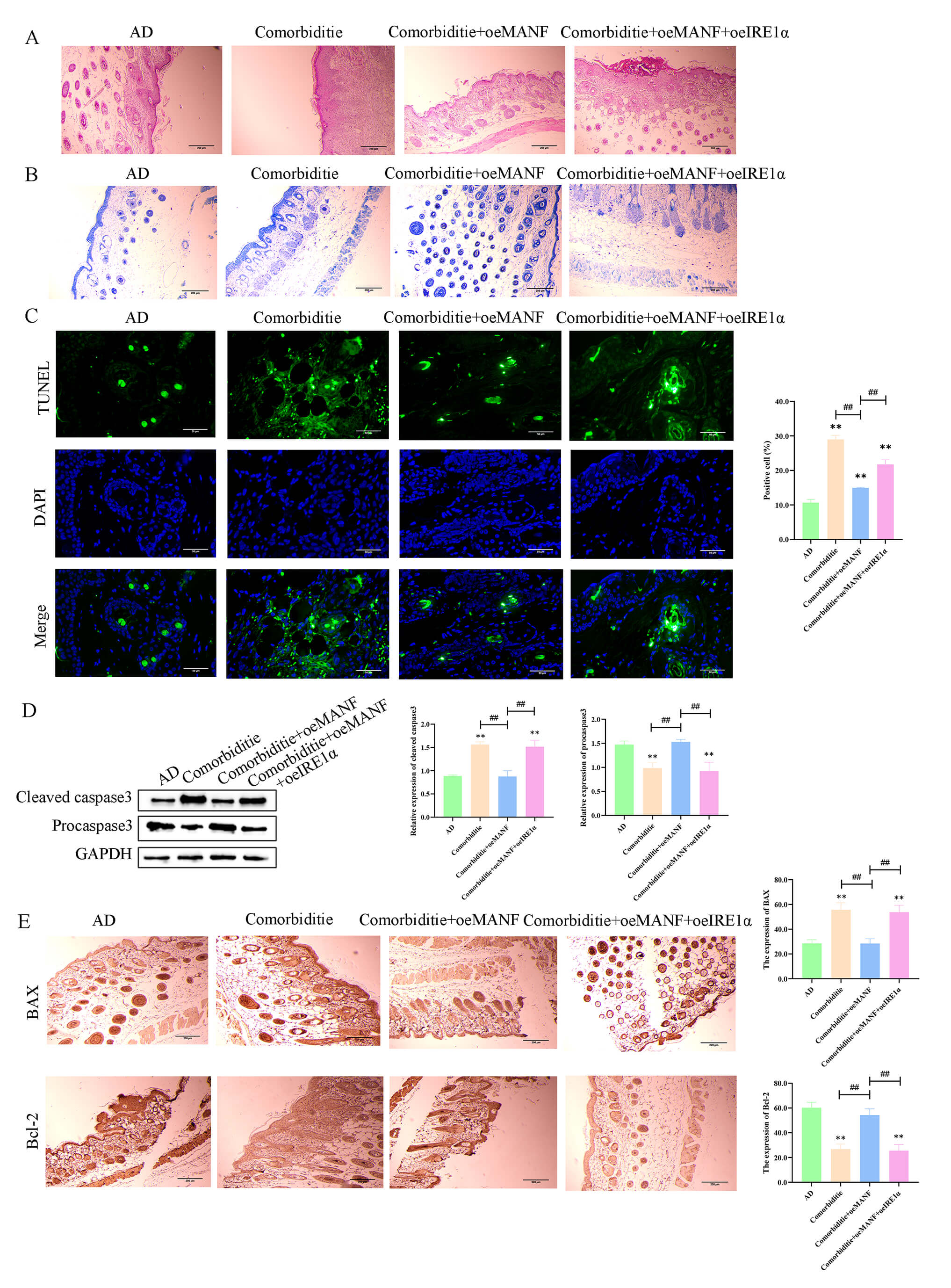 Fig. 8.
Fig. 8.
MANF alleviates skin pathology and inhibits apoptosis via
IRE1
In brain tissues, the Comorbidity group exhibited elevated caspase‑12, GRP78,
and p-IRE1
Collectively, MANF inhibits IRE1
AD pathogenesis is complex involving genetics, immunity, and environment, and recent studies indicate that ERS aggravates skin damage and promotes psychiatric symptoms by regulating immunity, apoptosis, and inflammation [21]. Grounded on this understanding, this study constructed a mouse AD model with comorbid psychiatric disorders to analyze the role of ERS in this process and explore the potential role of MANF in regulating ERS.
In this study, ERS-related proteins (GRP78, P-IRE1, Caspase-12) were
significantly upregulated, particularly in AD patients with psychiatric disorders
(p
ATF4, CHOP, and P-eIF2
ERS inhibitors (4-PBA) effectively regulate Bax/Bcl-2 expression to reduce skin
cell apoptosis, offering a novel strategy for ER-related skin diseases; Bax
promotes apoptosis while Bcl-2 exerts anti-apoptotic effects [31]. Our results
showed abnormal apoptosis-related protein expression in AD mice with psychiatric
disorders, characterized by significantly upregulated Bax and downregulated
Bcl-2. Previous studies confirmed that ERS promotes apoptosis via regulating
Bax/Bcl-2, thus exacerbating tissue injury [32]. This indicates that in the AD
mouse model, ERS may lead to excessive skin cell apoptosis by promoting Bax
activation, thereby exacerbating skin inflammation and barrier dysfunction.
Finally, this study confirmed that MANF significantly improved skin damage and
immune responses in the AD with psychiatric disorders mouse model. Specifically,
similar to the clinical sample results, GRP78, P-IRE1, and Caspase-12 expression
was significantly reduced, thereby alleviating ERS responses, with MANF playing
an intermediary regulatory role. Studies have demonstrated that P-IRE1 plays a
driving role in the pathological process of AD, particularly in exacerbating
local inflammation by regulating immune cell function [33]. Another study
revealed that MANF interacts with ER sensor IRE1
A limitation of this study is that 4-PBA has broad chaperone activity, and potential non-specific effects cannot be completely excluded despite its well-characterized role as an ER stress inhibitor. However, insufficient recruitment of eligible healthy volunteers precluded molecular marker testing and group comparisons, reducing conclusion specificity. Moreover, only 6 cases per clinical group (due to the difficulty in screening AD patients with psychiatric comorbidities) led to insufficient statistical power and limited generalizability. Future studies will expand recruitment, conduct multicenter research to supplement healthy control data and increase sample size, thereby enhancing study rigor and conclusion applicability. Further, expression changes of ERS-related factors in skin tissues were only evaluated at the whole-tissue level in this study, with no identification of their specific cellular sources via techniques. This precluded accurate cellular target localization for MANF-mediated ERS regulation and hindered in-depth clarification of MANF’s molecular mechanisms. Future studies are recommended to employ cell-specific detection techniques to address this gap.
In summary, our study indicates that changes in the ERS and apoptosis pathways in patients with AD and psychiatric disorders may play a crucial role in disease progression. MANF, as an ERS-related protein, helps improve skin damage and immune abnormalities in AD and comorbid psychiatric disorder mice by regulating these pathways.
The datasets generated and analyzed during the current study are not publicly available due to ethical restrictions, but will be made available on reasonable request from the corresponding author.
HC: Data curation, Formal analysis, Writing—original draft, Writing—review and editing; JZ, YG: Conceptualization; GY: Data curation; YZ: Obtaining funding, performed the research; SH and YL: Investigation, analyzed the data. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
The animal experiment was approved by the Second Affiliated Hospital of Harbin Medical University (YJSDW2024-001); The clinical research was approved by The Second Affiliated Hospital of Harbin Medical University (YJSKY2024-277). All animal procedures were strictly performed in accordance with the Guidelines for the Care and Use of Laboratory Animals issued by the Ministry of Science and Technology of China, as well as the ARRIVE guidelines and international animal welfare standards. All efforts were made to minimize animal suffering and the number of experimental animals used. The study was carried out in accordance with the guidelines of the Declaration of Helsinki, and written informed consent was obtained from all participants.
Not applicable.
Key Research and Development Program Project of Heilongjiang Province: Research on Comorbidity of Atopic Dermatitis and Mental Disorders (SC2022ZX06C0097).
The authors declare no conflicts of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBL47821.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.