
 , Yunlong Xiao 1, Junjie Chen 4, GuoYou Wang 1,3,*
, Yunlong Xiao 1, Junjie Chen 4, GuoYou Wang 1,3,*
1 Department of Joint Surgery, The Affiliated Traditional Chinese Medicine Hospital of Southwest Medical University, 646000 Luzhou, Sichuan, China
2 The First School of Clinical Medicine, Beijing University of Chinese Medicine, 100029 Beijing, China
3 Center for Orthopedic Diseases Research, The Affiliated Traditional Chinese Medicine Hospital, Southwest Medical University, 646000 Luzhou, Sichuan, China
4 Department of Integrated Traditional Chinese and Western Medicine, Chengdu University of Traditional Chinese Medicine, 611137 Chengdu, Sichuan, China
†These authors contributed equally.
Abstract
The gut–bone axis represents a paradigm shift in our understanding of skeletal biology, revealing how the gastrointestinal system and its microbial inhabitants profoundly influence bone homeostasis through complex bidirectional communication. This comprehensive review synthesizes recent high-impact research to elucidate the multifaceted mechanisms underlying gut–bone crosstalk, including microbial metabolite signaling, gut barrier integrity, immune modulation, and endocrine regulation. We examine how short-chain fatty acids (SCFAs), tryptophan metabolites, and bile acid derivatives modulate bone remodeling through epigenetic, immunologic, and hormonal pathways. The translational implications of these findings for the management of osteoporosis, osteoarthritis, and inflammatory bone disorders are critically evaluated, with particular emphasis on microbiome-targeted interventions, gut hormone-based therapies, and innovative approaches such as engineered microbial therapeutics. Furthermore, we explore the roles of farnesoid X receptor (FXR) signaling, G protein–coupled receptor activation, and mitochondrial function in bone cells as modulated by gut-derived factors. This review provides a framework for developing novel diagnostic and therapeutic strategies that target the gut–bone axis, highlighting the transition from traditional calcium-centric bone health paradigms toward integrated microbiome-targeted approaches that address the systemic nature of bone metabolism regulation.
Keywords
- gastrointestinal microbiome
- osteoporosis
- short-chain fatty acids
- bone remodeling
- metabolic bone diseases
- homeostasis
- metabolome
Bone remodeling is an exquisitely regulated process that maintains skeletal integrity through the balanced activities of osteoblasts and osteoclasts [1]. Traditional paradigms of bone homeostasis have emphasized hormonal regulation, mechanical loading, and nutritional status as primary determinants of bone mass [2]. However, the discovery that germ-free mice exhibit significant alterations in bone density first suggested that microbial colonization influences skeletal development [3]. Subsequent research has established the gut-bone axis as a critical regulatory system integrating gastrointestinal and skeletal health through complex bidirectional communication [4].
The clinical relevance of this axis is underscored by the high prevalence of osteoporosis and fractures in patients with inflammatory bowel disease (IBD), celiac disease, and other gastrointestinal disorders [5]. Meta-analyses have quantified these associations, revealing that IBD patients have a 40% increased risk of osteoporosis (relative risk [RR] 1.40, 95% CI 1.25–1.56) and a 60% higher overall fracture risk (RR 1.60, 95% CI 1.30–1.97) compared to matched controls. The risk is particularly elevated for vertebral fractures (RR 2.48, 95% CI 1.97–3.12). Similarly, individuals with celiac disease exhibit a 2.1-fold increased odds of osteoporosis (odds ratio [OR] 2.1, 95% CI 1.7–2.6) and a 1.4-fold higher risk of any fracture (hazard ratio [HR] 1.4, 95% CI 1.2–1.7). These associations cannot be fully explained by malabsorption or nutritional deficiencies alone, suggesting direct mechanistic links between gut health and bone metabolism [6]. Furthermore, age-related changes in gut microbiota composition correlate with bone loss, implicating microbial dysbiosis in the pathogenesis of senile osteoporosis [7].
However, these classical frameworks fail to fully account for the skeletal manifestations observed in various gastrointestinal and metabolic disorders, nor can they adequately explain the significant bone density alterations identified in germ-free animal models. This conceptual gap has been bridged by the emerging paradigm of the gut-bone axis, which reveals how extra-skeletal factors—particularly the gut microbiota and its diverse metabolic output—profoundly influence bone remodeling through integrated immune, endocrine, and neural pathways. The gastrointestinal system, far from being merely a digestive organ, thus emerges as a critical regulator of skeletal homeostasis, orchestrating systemic responses that complement traditional calcium-centric and mechanical loading paradigms.
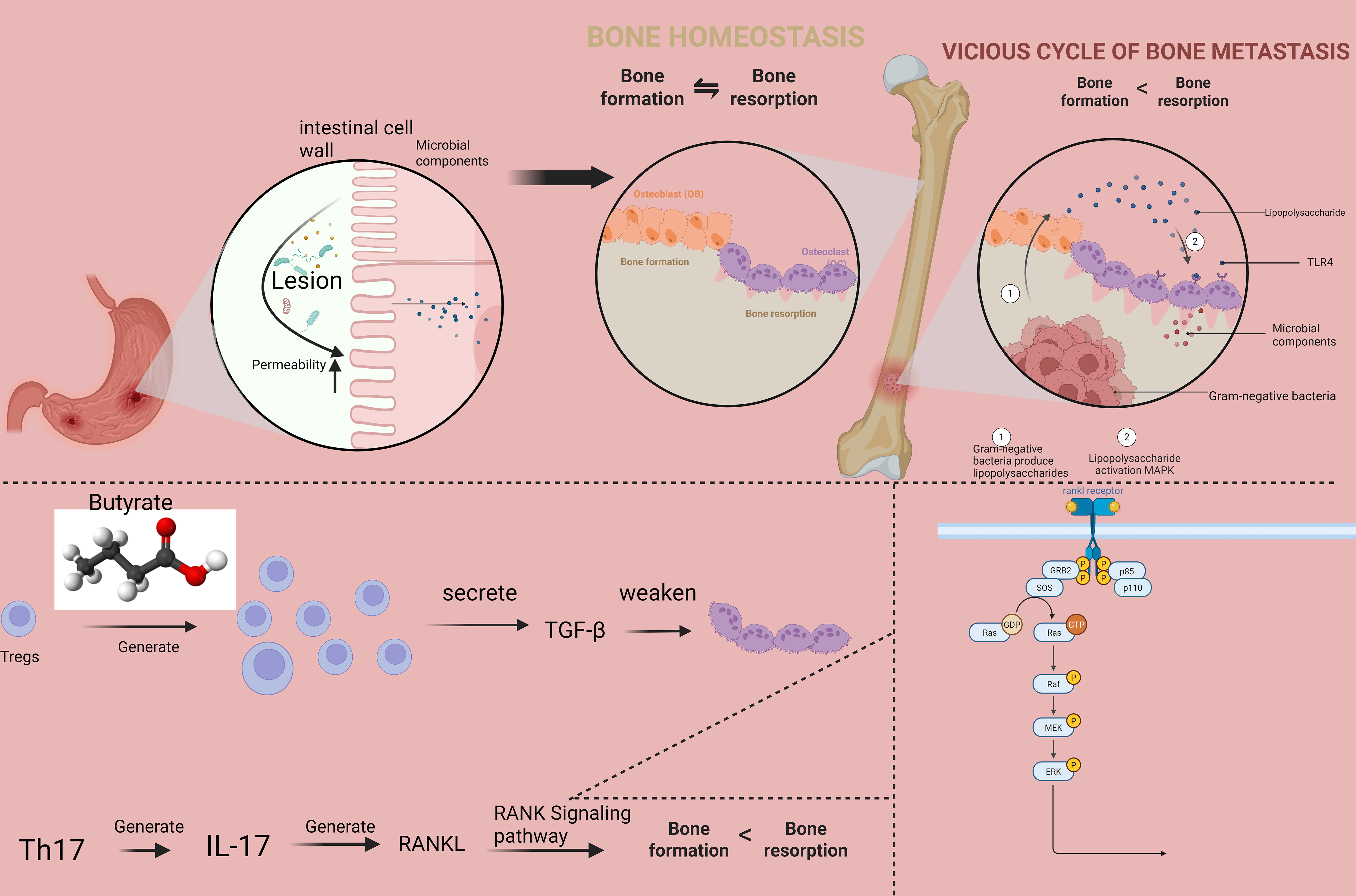
This manuscript provides a comprehensive analysis of the molecular mechanisms underlying gut-bone communication, with particular emphasis on recent advances published in high-impact journals. We examine how microbial metabolites influence bone cell function through receptor-mediated signaling and epigenetic modifications, how gut barrier integrity affects skeletal health, and how immune cells serve as intermediaries between intestinal and bone compartments (Fig. 1). Therapeutic implications for microbiome-modulating interventions are discussed in depth, with a critical appraisal of current evidence and future directions for research and clinical translation. These interventions encompass several promising approaches, including specific probiotic strains (e.g., Lactobacillus and Bifidobacterium species), prebiotic supplements (e.g., inulin, fructooligosaccharides), targeted dietary modifications (e.g., high-fiber, Mediterranean diets), fecal microbiota transplantation (FMT), and emerging synthetic biology approaches such as engineered microbial therapeutics. Accumulating evidence from preclinical models and preliminary clinical studies has demonstrated the efficacy of these strategies in ameliorating bone loss, improving bone microarchitecture, and reducing fracture risk, highlighting their translational potential for managing metabolic bone disorders [6, 7].
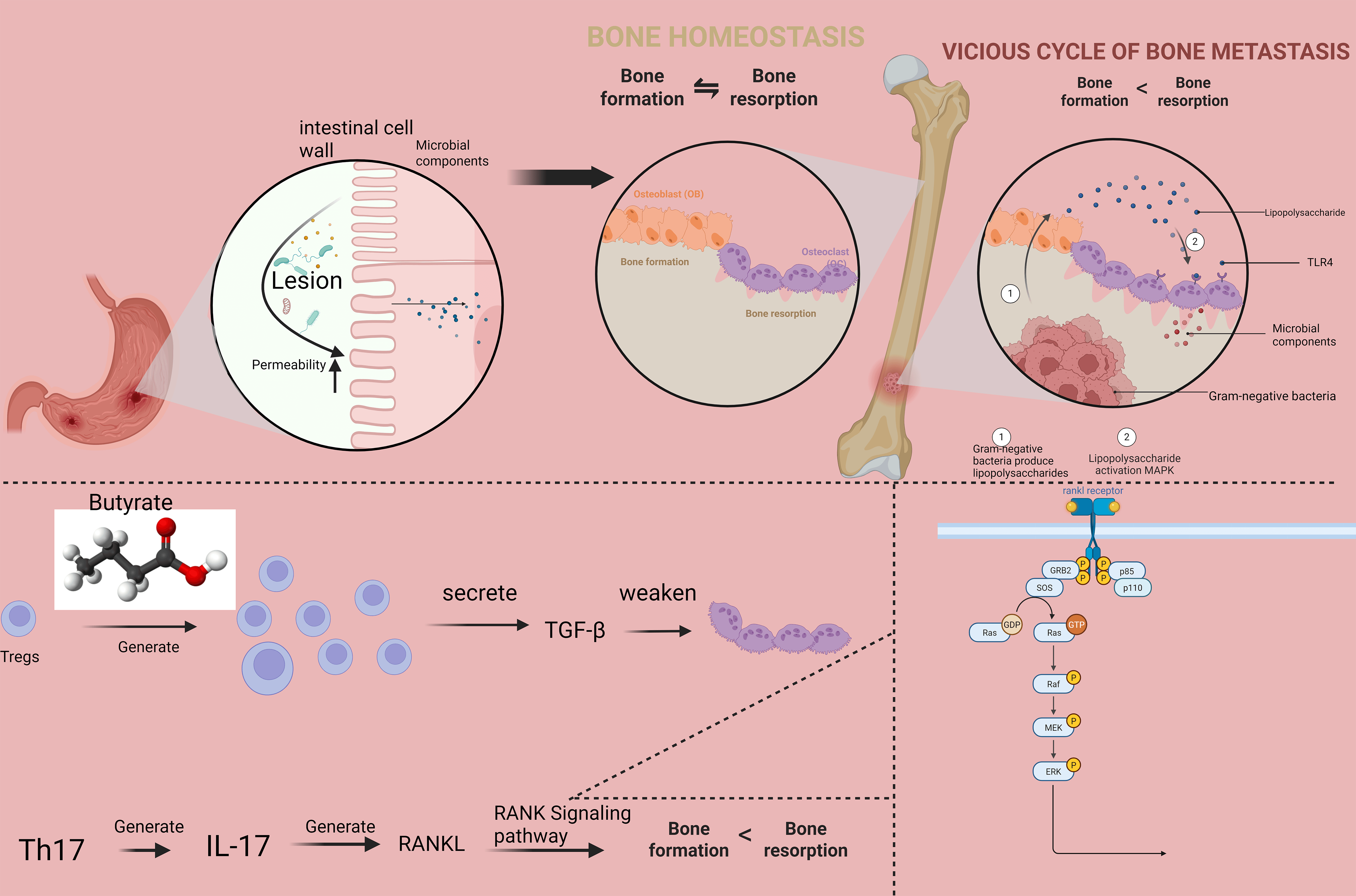 Fig. 1.
Fig. 1.
The molecular interplay within the gut-bone axis
linking compromised intestinal integrity to disrupted bone homeostasis. The
model illustrates how altered circulating cytokine levels differentially regulate
osteoblastic and osteoclastic activity, shifting the bone remodeling balance
toward net resorption. TGF-
Short-chain fatty acids (SCFAs), including acetate, propionate, and butyrate,
are produced by microbial fermentation of dietary fiber and represent key
mediators of gut-bone communication [8]. Acetate, the most abundant SCFA in
circulation, exerts multifaceted effects on bone homeostasis primarily through
activation of its cognate receptors G protein-coupled receptor 43 (GPR43) and
GPR41. In osteoblasts, acetate binding to GPR43 stimulates the
Wnt/
Tryptophan metabolism by gut microorganisms generates numerous bioactive
compounds, including indole derivatives, serotonin, and kynurenine pathway
metabolites [15]. Indole-3-aldehyde and indole-3-propionic acid activate the aryl
hydrocarbon receptor (AhR) in osteoblasts, promoting expression of Cytochrome
P450 family 1 subfamily A member 1 (CYP1A1) and Cytochrome P450 family 1
subfamily B member 1 (CYP1B1) while enhancing osteogenic differentiation [16].
AhR activation also suppresses osteoclastogenesis by inhibiting NF-
Primary bile acids undergo microbial transformation into secondary bile acids, which function as signaling molecules through activation of both nuclear and membrane-bound receptors [22]. The G protein-coupled bile acid receptor TGR5 (GPBAR1) has emerged as a crucial mediator of bile acid actions on bone metabolism. Lithocholic acid (LCA) and deoxycholic acid (DCA) activate Takeda G protein-coupled Receptor 5 (TGR5) on osteoblasts, triggering intracellular Cyclic Adenosine Monophosphate (cAMP) accumulation and subsequent protein kinase A (PKA) activation, which phosphorylates the transcription factor cAMP response element-binding protein (CREB) [22, 23]. This signaling cascade promotes osteoblast proliferation, differentiation, and mineralization by upregulating key osteogenic factors including RUNX2, Osterix, and alkaline phosphatase [23]. Beyond direct effects on bone cells, TGR5 signaling exerts systemic influence on bone homeostasis through multiple pathways. TGR5 activation in intestinal enteroendocrine L-cells stimulates glucagon-like peptide-1 (GLP-1) secretion, which contributes to the postprandial suppression of bone resorption [24]. Additionally, TGR5 expression in immune cells modulates inflammatory responses, potentially influencing osteoclast differentiation and activity in inflammatory bone diseases [25]. Preclinical evidence from TGR5-deficient mouse models demonstrates accelerated age-related bone loss and impaired fracture healing, underscoring the physiological importance of this receptor in skeletal maintenance [26]. The interplay between TGR5 and farnesoid X receptor (FXR) signaling creates a sophisticated regulatory network for bile acid-mediated bone homeostasis. While TGR5 activation generally promotes bone formation through both direct and indirect mechanisms, FXR signaling exerts more complex, tissue-specific effects [23]. Intestinal-specific FXR activation appears to negatively regulate bone mass, as evidenced by the 24% increase in trabecular bone volume observed in intestinal epithelial cell-specific FXR knockout mice [27]. This effect is associated with reduced sclerostin expression and enhanced Wnt signaling in bone tissue [27]. Ursodeoxycholic acid (UDCA) and its taurine-conjugated derivative Tauroursodeoxycholic acid (TUDCA) function as dual modulators of both TGR5 and FXR signaling pathways, contributing to their osteoprotective effects [28]. In postmenopausal women with osteopenia, UDCA supplementation (15 mg/kg/day) for 12 months increased lumbar spine Bone Mineral Density (BMD) by 2.8% compared to placebo [29]. Mechanistic study indicate that UDCA reduces oxidative stress in bone marrow mesenchymal stem cells and enhances their osteogenic potential through complementary activation of both TGR5-mediated signaling and Nrf2 antioxidant pathways [30].
Trimethylamine N-oxide (TMAO), a gut microbiota-derived metabolite generated
from dietary precursors such as choline, L-carnitine, and betaine, has been
extensively studied in relation to bone metabolism, though its role remains
complex and context-dependent [29]. TMAO is produced via a two-step process: gut
microbial conversion of dietary nutrients to trimethylamine (TMA), followed by
hepatic oxidation via flavin-containing monooxygenase 3 (FMO3) [30]. Current
evidence reveals a concentration-dependent duality in TMAO’s skeletal effects.
Moderate TMAO levels have demonstrated osteoprotective properties in preclinical
models. In ovariectomized rats, TMAO supplementation (120 mg/kg/day) attenuated
trabecular bone loss by approximately 30%, suppressing osteoclast
differentiation through inhibition of NF-
Beyond the well-characterized SCFAs, tryptophan metabolites, and bile acids, gut
microbiota-derived polyamines and vitamins significantly contribute to bone
homeostasis [29]. Polyamines, including spermidine and spermine, are produced by
bacteria such as Bifidobacterium and Lactobacillus through enzymatic
decarboxylation of amino acids [30]. These compounds enhance osteoblast
differentiation and mineralization by modulating autophagy and reducing oxidative
stress [29, 31]. In aged murine models, dietary spermidine supplementation (3 mM
in drinking water) increased trabecular bone volume by 27% and improved bone
strength through upregulation of Nrf2-dependent antioxidant pathways [32].
Microbiota-synthesized vitamin K2 (menaquinone) is essential for
| Metabolite class | Representative molecules | Primary microbial producers | Molecular targets & receptors | Signaling pathways & mechanisms | Observed effects on bone in vivo | Clinical & translational insights |
| Short-chain fatty acids (SCFAs) | Butyrate | Faecalibacterium prausnitzii, Roseburia spp., Lachnospiraceae | HDACs, GPR41, GPR43 | HDAC inhibition |
Collagen-induced arthritis: |
Circulating butyrate levels correlate positively with BMD in postmenopausal women. Butyrate-producing capacity of gut microbiome is reduced in osteoporosis. |
| Propionate | Bacteroides spp., Dialister spp. | GPR41, GPR43 | GPR43 activation on osteoblasts |
GPR43⁻/⁻ mice show abolished osteogenic response. Propionate ester supplementation prevents OVX-induced bone loss. | - | |
| Tryptophan derivatives | Indole-3-propionic acid (IPA), Indole-3-aldehyde (IAld) | Lactobacillus spp., Clostridium sporogenes | Aryl Hydrocarbon Receptor (AhR) | AhR ligand binding |
Engineered L. reuteri (tryptophan overproducer) prevents OVX-induced bone loss by 78%. AhR antagonism blocks protective effects. | Serum IPA levels are inversely correlated with bone turnover markers in IBD patients. |
| Bile acids | Ursodeoxycholic Acid (UDCA), Lithocholic Acid (LCA) | Clostridium scindens, C. hiranonis (7 |
TGR5, FXR, VDR, Nrf2 | Nrf2 TGR5 (LCA, UDCA): Activation |
Nrf2 TGR5 (LCA, UDCA): Activation |
UDCA clinical trial (15 mg/kg/d): |
| Microbial polyamines | Spermidine, Spermine | Bifidobacterium spp., Lactobacillus spp. | Autophagy pathways, Nrf2 | Enhancement of autophagy flux |
Enhancement of autophagy flux |
Fecal spermidine levels correlate with BMD in elderly cohorts. |
| Bacterial vitamins | Menaquinone (Vitamin K2) | Bacteroides spp., Escherichia coli, Bacillus subtilis | Pregnane X Receptor (PXR), GGCX enzyme | PXR activation: Promotes osteoblast gene expression. Cofactor for GGCX: |
Vitamin K-deficient diets |
High circulating MK-7 |
| Reference | [8, 9, 10, 11, 12, 13, 23, 24, 25, 26, 27, 28, 29, 44, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69] | [8, 9, 10, 11, 21, 22, 23, 24, 25, 26, 27, 42, 43, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 69] | [10, 28, 29, 30, 31, 32, 41, 47, 70] | [8, 9, 10, 11, 21, 22, 23, 24, 25, 26, 27, 42, 43, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 69] | [36, 39, 40, 45, 46, 48, 58] | [11, 12, 13, 14, 45, 46, 48, 58, 69] |
Increased intestinal permeability permits translocation of microbial components
into systemic circulation, triggering immune activation that adversely affects
bone remodeling [36]. Lipopolysaccharide (LPS) from Gram-negative bacteria
promotes osteoclast formation through Toll-like receptor 4 (TLR4)-mediated
activation of NF-
The gut microbiota profoundly influences the development and function of immune
cells that participate in bone remodeling [76]. Regulatory T cells (Tregs)
expanded by microbial metabolites such as butyrate suppress osteoclastogenesis
through production of IL-10, TGF-
The connection between IBD and osteoporosis exemplifies the clinical significance of the gut-bone axis [83]. Approximately 67% of IBD patients present with osteopenia and 57.6% with osteoporosis, indicating profound skeletal complications beyond traditional malabsorption explanations [42, 83]. Recent research has identified novel epigenetic mechanisms underlying IBD-induced bone loss [42]. A groundbreaking study revealed that Fat Mass and Obesity-Associated protein (FTO) SUMOylation at sites K216, K357, and K365 regulates the differentiation of bone marrow mesenchymal stromal cells (BMSCs) in IBD-induced bone loss 4 [43, 84]. SUMOylation of FTO impairs its N6-methyladenosine (m6A) RNA demethylase activity, shifting BMSC differentiation toward adipogenesis rather than osteogenesis [43]. This epigenetic modification represents a critical mechanism through which intestinal inflammation affects skeletal homeostasis [43]. Therapeutic targeting of this pathway has shown promise. Interleukin-6 receptor monoclonal antibody (tocilizumab) combined with Adeno-Associated Virus (AAV)-FTO-3KR (a mutant resistant to SUMOylation) attenuated bone loss and enhanced bone formation in IBD mice [85, 86]. This approach highlights the potential of targeting specific epigenetic modifications in the treatment of inflammation-induced osteoporosis [86]. The role of microbial metabolites in IBD-related bone loss extends beyond epigenetic regulation. Butyrate deficiency in IBD patients correlates with impaired bone healing and reduced osteoblast activity [87]. Rectal administration of butyrate enemas in a dextran sulfate sodium (DSS)-induced colitis model restored osteoblast function and reduced trabecular bone loss, highlighting the therapeutic potential of metabolite replacement [88]. Additionally, microbiota-derived hydrogen sulfide (H2S) from sulfate-reducing bacteria exacerbates bone loss in IBD by inducing osteocyte apoptosis through mitochondrial dysfunction [89, 90]. Inhibition of H2S production with tungstate treatment preserved bone mass in colitis models by reducing caspase-3 activation in osteocytes [90] (Table 2, Ref. [9, 10, 11, 12, 13, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 37, 38, 39, 40, 41, 42, 43, 64, 71, 72, 79, 80, 81, 82, 84, 85, 86, 87, 88, 89, 91, 92, 93, 94]).
| Model category | Induction method | Key gut microbiome phenotype | Key bone phenotype | Primary mechanistic insights | Successful interventions |
| Germ-free (GF) | Raised in sterile isolators | Complete absence of microbial colonization. | Reduced bone mass ( |
Lack of microbial stimuli |
Monocolonization studies reveal specific bacterial functions (e.g., SFB |
| Ovariectomy (OVX) | Surgical removal of ovaries | Reduced diversity, |
High-turnover osteoporosis: |
Estrogen deficiency |
Probiotics (e.g., L. reuteri), prebiotics (inulin), SCFA supplements significantly ameliorate bone loss. |
| Inflammatory bowel disease (IBD) | DSS in drinking water; IL-10 | Severe dysbiosis, loss of mucosal-associated bacteria, expansion of E. coli. | Systemic bone loss, inhibited bone formation, increased marrow adiposity. | Chronic intestinal inflammation |
Anti-TNF- |
| Aging model | Natural aging (18–24 month) | Loss of diversity, |
Age-related bone loss: reduced bone formation, decreased bone quality. | Inflammaging ( |
Caloric restriction; supplementation with A. muciniphila or butyrate; senolytics. |
| High-fat diet (HFD) | 45–60% fat diet | Increased bone fragility and decreased bone mass despite higher body weight. | Gut barrier dysfunction |
FXR antagonism; Prebiotics/probiotics to restore barrier function and SCFA production. | |
| References | [9, 10, 11, 12, 29, 39, 40, 71, 72, 81, 82] | [21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 37, 38, 81, 82, 88, 89, 91] | [13, 29, 34, 37, 38, 42, 43, 84, 85, 86, 87, 93, 94] | [21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 37, 38, 64, 88, 89, 91, 92] | [12, 37, 38, 41, 43, 79, 80, 84, 85] |
Emerging research highlights innate immune cells as critical intermediaries in
the gut-bone axis. Group 3 innate lymphoid cells (ILC3s) reside in the intestinal
lamina propria and are educated by the local microbial milieu [95]. ILC3s produce
IL-22, a cytokine that enhances gut barrier integrity by stimulating mucin
production and epithelial cell regeneration [44]. Furthermore, IL-22 exerts
direct osteoanabolic effects by activating Signal Transducer and Activator of
Transcription 3 (STAT3) signaling in osteoblasts, increasing their proliferation
and differentiation capacity [91]. Microbial dysbiosis, particularly a reduction
in Lactobacillus species, diminishes IL-22 production, leading to a compromised
barrier and reduced bone formation [96]. Conversely, mucosal-associated invariant
T (MAIT) cells, activated by microbial vitamin B2 derivatives, can exacerbate
bone erosion in inflammatory conditions by producing pro-osteoclastic cytokines
like TNF-
Enteroendocrine hormones play a pivotal role in postprandial bone turnover regulation [99]. Glucose-dependent insulinotropic polypeptide (GIP), glucagon-like peptide 1 (GLP-1), and glucagon-like peptide 2 (GLP-2) are secreted by intestinal endocrine cells in response to nutrient intake and have demonstrated significant effects on bone remodeling processes [99, 100]. At physiological concentrations, GIP appears to be the primary contributor to postprandial bone resorption suppression, while supraphysiological concentrations of gut hormones induce more potent anti-resorptive effects [100]. Notably, research has revealed that GIP infusion significantly reduced bone resorption (as measured by CTX levels) by 14% at 30 minutes, while GLP-1 showed no such effect [45]. This suggests that GIP’s osteoprotective actions remain intact even in conditions where its insulinotropic effects are impaired, highlighting its potential as a therapeutic target for bone disorders independent of its glycemic functions [45, 99]. GLP-2 has emerged as another important regulator of bone metabolism, primarily through its effects on reducing parathyroid hormone (PTH) and enhancing intestinal calcium absorption [101]. The combined application of GIP and GLP-2 in preclinical studies has shown synergistic benefits for bone quality, suggesting potential for combination therapies targeting multiple gut hormone pathways [102].
Emerging evidence suggests that the autonomic nervous system (ANS) serves as an important conduit in gut-bone communication [103]. A Mendelian randomization and mediation analysis study revealed that specific gut microbiota taxa can influence bone density through heart rate variability (HRV), a measure of autonomic function, with mediation effects reaching up to 40% [104]. This suggests that gut microbes may modulate bone metabolism through neural pathways in addition to endocrine and immune mechanisms. The proposed “gut-nerve-bone axis” represents a novel dimension of skeletal regulation that expands our understanding of how gastrointestinal health influences bone homeostasis [103, 105]. Vagal nerve stimulation and other neuromodulatory techniques may offer future therapeutic approaches for osteoporosis that leverage this neural connectivity [106] (Table 3, Ref. [8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 36, 39, 40, 41, 44, 64, 70, 77, 79, 80, 81, 82, 98, 107, 108]).
| Immune cell | Regulatory T cells (Tregs) | T helper 17 cells (Th17) | Regulatory B cells (Bregs) | Group 3 innate lymphoid cells (ILC3s) | Macrophages |
| Microbial cues | SCFAs (Butyrate via HDACi) | Segmented Filamentous Bacteria (SFB); Bile acids | B. fragilis PSA via TLR2 | AhR ligands (Tryptophan metabolites) | LPS (TLR4); SCFAs (GPR43) |
| Functional change | Expansion, enhanced suppressive function, stability (FoxP3+CTLA-4+) | Differentiation, expansion, production of pro-inflammatory cytokines | Differentiation into IL-10 and OPG-producing cells | Production of IL-22. Maintenance of epithelial barrier integrity | Altered polarization: M1 (pro-inflammatory) vs. M2 (anti-inflammatory) |
| Net effect on bone | Potent inhibition of osteoclastogenesis. Potential indirect promotion of bone formation | Potent stimulation of osteoclastogenesis, leading to inflammatory bone erosion | Inhibition of osteoclastogenesis via OPG production. Maintenance of bone homeostasis | Indirect (barrier protection) and direct (STAT3 activation in osteoblasts) promotion of bone formation | M1: Pro-osteoclastogenic via TNF- |
| Key molecular mediators | IL-10, TGF- |
IL-17, IL-22, RANKL | IL-10, OPG, TGF- |
IL-22, STAT3 | TNF- |
| Therapeutic targetability | Butyrate/probiotic supplementation; Low-dose IL-2; Treg adoptive transfer | Anti-IL-17/IL-23 biologics; Depletion of SFB (e.g., via antibiotics/phages) | PSA or synthetic TLR2 agonists as immunomodulators | AhR agonists (FICZ); Recombinant IL-22 | Prebiotics |
| Reference | [8, 9, 10, 11, 14, 64, 107] | [8, 12, 41, 79, 80] | [70, 80, 81, 82, 98] | [15, 16, 17, 18, 19, 44, 77] | [8, 9, 10, 11, 12, 13, 36, 39, 40, 108] |
PSA, Polysaccharide A; RANKL, Receptor activator of nuclear factor kappa-B ligand; CTLA-4, Cytotoxic T-Lymphocyte-Associated protein 4.
Gut microbiota significantly modulate calcium and magnesium absorption through
acidification of the colonic lumen via SCFA production [107]. This acidic
environment enhances solubility and passive absorption of minerals critical for
bone mineralization [92]. Lactobacillus and Bifidobacterium strains increase
expression of transient receptor potential vanilloid type 6 (TRPV6) and
calbindin-D9k in intestinal epithelial cells, facilitating active calcium
transport [92, 107]. In ovariectomized rats, probiotic supplementation with
Lactobacillus helveticus increased calcium absorption efficiency by 38% and
prevented femoral bone loss by maintaining serum ionized calcium levels [46].
Magnesium absorption is similarly enhanced by microbial action. Akkermansia
muciniphila promotes magnesium absorption through upregulation of Transient
Receptor Potential Melastatin6 (TRPM6) and TRPM7 channels in the colon [47, 109].
Magnesium deficiency alters bone crystal structure and increases osteoclast
activity, while supplementation restores bone quality by reducing IL-1
Gut dysbiosis can activate the hypothalamic-pituitary-adrenal (HPA) axis,
leading to elevated systemic glucocorticoid levels that are detrimental to bone
[112]. Pathobionts such as Enterococcus faecalis stimulate immune cells to produce
IL-1
| Study type | Population/cohort | Key findings related to gut-bone axis | Implications | References |
| Epidemiological & observational studies | Elderly populations | • Inverse correlation between circulating LPS levels and BMD; highest quartile had 3.2x greater risk of vertebral fracture | • Systemic microbial antigen exposure is a novel risk factor for osteoporosis | [32, 47, 48, 110, 111, 116, 117, 118] |
| • Higher serum vitamin K2 (menaquinone) levels associated with 34% reduction in hip fracture risk | • Microbiota-derived vitamin K2 is crucial for bone mineralization | |||
| Inflammatory Bowel Disease (IBD) patients | • ~67% prevalence of osteopenia; ~58% prevalence of osteoporosis | Bone loss in IBD is multifactorial, involving inflammation, malabsorption, and gut barrier dysfunction | [32, 34, 85, 86, 87, 88, 89, 90] | |
| • Elevated serum zonulin (marker of gut permeability) correlated with |
||||
| Interventional clinical trials | Postmenopausal women with osteopenia | UDCA supplementation (15 mg/kg/day, 12 months) increased lumbar spine BMD by 2.8% vs. placebo | Repurposing bile acid drugs for osteoporosis is a viable therapeutic strategy | [27, 28, 29, 55, 58] |
| Postmenopausal women with osteoporosis | Vitamin K2 supplementation (45 mg/day, 24 weeks) improved ucOC levels and increased lumbar spine BMD by 1.7% | Supplemental microbial vitamins can improve bone quality | [49, 50, 51, 52, 53, 114] | |
| Patients with Celiac Disease | Larazotide acetate (zonulin antagonist) reduced CTX by 34% | Restoring gut barrier integrity is a valid target for reducing bone resorption | [45, 71, 72, 110, 115, 119] | |
| Microbiome association studies | Osteoporosis vs. Healthy Controls | Distinct gut microbiota composition (dysbiosis) characterized by reduced microbial diversity, altered SCFA-producing taxa, and enrichment of pro-inflammatory species | Microbiome signatures could serve as diagnostic or prognostic biomarkers for bone disease | [7, 41, 79, 96, 120] |
| Osteoarthritis (OA) patients | • Reduction of C. bolteae correlated with OA severity | A “gut-bile acid-joint axis” exists, suggesting novel therapeutic targets for OA | [27, 28, 29, 54, 55, 76] | |
| • GUDCA levels inversely correlated with joint degeneration via intestinal FXR inhibition |
GUDCA, glycoursodeoxycholic acid; TBS, Trabecular Bone Score.
Targeted manipulation of the gut microbiota through probiotic supplementation and prebiotic substrates represents a promising strategy for improving bone health [101]. Specific bacterial strains have demonstrated significant osteoprotective effects in clinical and preclinical studies [102]. Lactobacillus reuteri DSM 17938 has been shown to increase annual growth velocity by 7 cm in 1-6-year-old children, while Bifidobacterium longum enhances Insulin-like Growth Factor-1(IGF-1) activity through upregulation of Insulin-like Growth Factor-Binding Protein 3(IGFBP3) [114, 115]. Similarly, Akkermansia muciniphila maintains chondrocyte morphology and improves bone microarchitecture [108]. Prebiotic approaches have also shown promise in supporting bone health. Goat milk oligosaccharides (GMO) enrich Bacteroidota populations and enhance IGF-1 receptor expression, while microbiome-directed complementary food (MDCF-2) formulations containing chickpea flour improved linear growth in malnourished children [48, 116]. These findings suggest that targeted nutritional interventions can effectively modulate the gut-bone axis for therapeutic benefit [116]. However, the efficacy of probiotic interventions demonstrates considerable inter-individual variability. This heterogeneity is primarily attributed to the host’s baseline gut microbiota composition, genetic makeup, dietary habits, and medication use, which collectively influence the colonization and functional expression of administered strains. Future research should therefore focus on identifying predictive biomarkers for treatment response to facilitate the development of personalized probiotic regimens, moving beyond a one-size-fits-all approach.
Fecal microbiota transplantation (FMT) from young donor mice to aged recipients restored bone mass and microarchitecture to levels comparable to young animals [117]. These effects were mediated through restoration of IGF-1 signaling and reduction of oxidative stress in bone tissue [118]. FMT from osteoporotic patients to germ-free mice recapitulated the bone phenotype, demonstrating the causal role of microbiota in bone pathology [119, 121]. Clinical applications of FMT for bone disorders are currently under investigation [120]. A pilot study of FMT in patients with corticosteroid-induced osteoporosis showed trends toward improved bone turnover markers and reduced bone loss, though larger studies are needed to confirm efficacy [122]. Safety concerns regarding long-term effects of FMT necessitate careful donor screening and monitoring [119]. The clinical application of FMT for skeletal disorders is still in its infancy, and its standardization is critical for ensuring consistent efficacy and safety. Key challenges include the establishment of rigorous donor screening protocols to exclude pathogens and undesirable metabolic phenotypes, the standardization of fecal material processing (e.g., microbial concentration, viability), and the determination of optimal delivery routes and dosages. Developing universally accepted industry standards and implementing long-term follow-up protocols are essential steps toward the rational and safe application of FMT in managing bone diseases.
The growing understanding of gut hormones in bone metabolism has spurred interest in hormone-based therapeutics for skeletal disorders [49]. GIP receptor agonists and GLP-2 analogs show particular promise for conditions characterized by increased bone resorption [49, 50]. The finding that GIP retains its bone-protective effects in pancreatic-insufficient cystic fibrosis (PI-CF) patients even when its insulinotropic actions are impaired suggests that GIP-based therapies could benefit patients with various forms of bone loss [50]. Dual- and multi-receptor agonists represent an advanced approach in this domain [51]. Tirzepatide (GIP/GLP-1 receptor agonist) and CagriSema (GLP-1/amylin analogue) may offer synergistic benefits for bone health through combined activation of complementary signaling pathways [51, 52]. However, concerns remain about potential over-suppression of bone turnover, which could lead to increased bone fragility despite improved density metrics [52]. Prolonged and profound suppression of bone remodeling presents several potential long-term consequences for bone quality and fracture risk. While increased bone mineral density (BMD) is typically associated with reduced fracture risk, excessively suppressed turnover impairs the repair of microdamage that accumulates during normal mechanical loading. This can lead to increased tissue age, advanced glycation end-product accumulation, and altered collagen cross-linking patterns, ultimately compromising bone toughness and increasing susceptibility to atypical fractures [45, 54]. Clinical evidence from long-term antiresorptive therapies demonstrates that excessive suppression of bone turnover (evidenced by very low levels of bone turnover markers such as CTX) is associated with increased risk of atypical femoral fractures, though the specific risk profile for gut hormone-based therapies requires further investigation [53, 108]. Therefore, careful monitoring of bone turnover markers and periodic assessment of treatment duration may be necessary to balance the benefits of reduced bone resorption against the potential risks of oversuppression when using dual- and multi-receptor agonists for bone disorders [54].
Recent research has revealed a novel “gut-bile acid-joint axis” with implications for bone health [54]. The study demonstrated that glycoursodeoxycholic acid (GUDCA) levels inversely correlate with osteoarthritis severity through selective inhibition of the FXR in the intestine [55]. This discovery suggests that existing bile acid-based medications, such as UDCA, might be repurposed for bone disorders through FXR modulation [56]. The reduction of C. bolteae in osteoarthritis patients’ guts and its association with disease severity further supports the therapeutic potential of microbiome modulation targeting specific bacterial taxa [57]. This approach may be particularly relevant for age-related bone loss, given the significant shifts in gut microbiota composition that occur with aging [54, 55].
Synthetic biology approaches offer innovative strategies for targeting the gut-bone axis [58]. The development of engineered Trp CB has demonstrated impressive results in animal models of osteoporosis and intestinal dysfunction [59]. This intervention not only repaired intestinal barrier function but also increased P1NP by 1.7-fold while reducing osteoclast numbers by 41.8% [60]. Notably, this approach demonstrated efficacy across age groups, increasing trabecular bone number by 35% and cortical bone density by 18.4% in 22-month-old aged mice [8]. The cross-age applicability suggests that microbial therapeutics might address both inflammatory and age-related bone loss through shared mechanisms involving barrier protection and inflammation reduction [61]. Despite the promising therapeutic potential of engineered bacteria, their long-term safety remains a paramount concern for clinical translation. Potential risks include the horizontal gene transfer of engineered constructs to host cells or other members of the indigenous microbiota, unpredictable behavior and evolution of engineered strains within the complex gut ecosystem, and unintended immunogenic reactions. Consequently, the implementation of robust biocontainment strategies and comprehensive long-term toxicological studies are imperative prerequisites before these innovative therapies can be widely applied in clinical settings.
Advancements in precision nutrition have enabled targeted dietary interventions based on individual microbiome compositions [62]. High-throughput sequencing identifies microbial signatures associated with bone health, allowing for personalized prebiotic and probiotic recommendations [63]. For instance, individuals with low Prevotella abundance benefit more from arabinoxylan oligosaccharides, which increase SCFA production and improve calcium absorption [64]. Machine learning algorithms integrating microbiome data, genetic polymorphisms, and clinical parameters predict responsiveness to specific interventions [65]. A recent clinical trial demonstrated that personalized nutrition plans based on gut microbiota composition increased lumbar spine BMD in osteopenic patients [66]. This approach represents a paradigm shift toward precision medicine in bone health management [66] (Table 5, Ref. [9, 10, 11, 12, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 37, 38, 39, 40, 64, 71, 72, 81, 88, 89, 91, 92]).
| Biomarker category | Specific biomarker | Biological significance | Assay/measurement | Association with bone health |
| Microbial metabolites | Fecal/Serum SCFAs (Butyrate, Propionate) | Direct mediators of microbial influence on bone (HDAC inhibition, GPCR activation) | GC-MS LC-MS/MS | Low levels associated with increased bone resorption and inflammation. |
| Serum Ursodeoxycholic Acid (UDCA)/GUDCA | FXR modulators with osteoprotective and chondroprotective potential | LC-MS/MS | UDCA supplementation increased BMD. | |
| Low GUDCA correlates with OA severity. | ||||
| Urinary/Serum Indole Derivatives (I3A, IPA) | Indicators of bacterial tryptophan metabolism and AhR ligand availability | Levels may predict responsiveness to pre/probiotics targeting tryptophan metabolism. | ||
| Gut barrier function | Serum Zonulin | Regulator of intestinal tight junctions; marker of “leaky gut” | ELISA | Elevated levels correlate with increased bone resorption markers (CTX) in celiac disease. |
| Circulating LPS (Endotoxemia) | Marker of bacterial translocation and systemic inflammation | LAL assay ELISA | Inverse correlation with BMD; high levels confer 3.2 | |
| Lipopolysaccharide-Binding Protein (LBP) | Acute-phase reactant that binds LPS, amplifying immune response | ELISA | Elevated LBP indicates chronic, low-grade endotoxemia and inflammation. | |
| Bone turnover & systemic inflammation | Citrullinated Proteins (ACPA) | Autoantibodies generated due to loss of gut barrier function and dysbiosis | ELISA | Link gut dysbiosis, RA, and associated inflammatory bone loss. |
| Ratio of ucOC to cOC | Indicator of vitamin K status and osteocalcin activation | ELISA | High ratio indicates vitamin K deficiency and impaired bone mineralization. | |
| Reference | [21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 37, 38, 81, 88, 89, 91] | [9, 10, 11, 12, 29, 39, 40, 71, 72, 81] | N/A | [21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 37, 38, 64, 88, 89, 91, 92] |
GPCR, G-Protein-Coupled Receptor; GC-MS, Gas Chromatography-Mass Spectrometry; LC-MS/MS, Liquid Chromatography-Tandem Mass Spectrometry; LAL, Limulus Amebocyte Lysate assay; cOC, circulating Osteocalcin; RA, Rheumatoid Arthritis.
Bacteriophages offer targeted manipulation of gut microbiota to promote bone health [67]. Phages specifically targeting osteoclast-promoting bacteria (e.g., Clostridium histolyticum) reduce bacterial load and subsequent bone resorption [67, 68]. In a proof-of-concept study, oral administration of C. histolyticum-specific phages (109 PFU/day) reduced serum CTX levels and increased trabecular bone volume in osteoporotic mice [69]. Phage therapy also minimizes disruption to beneficial microbiota, avoiding off-target effects common with broad-spectrum antibiotics. Phage engineering further enhances specificity and efficacy: modular phage platforms equipped with CRISPR-Cas systems can selectively deplete pathobionts while delivering osteogenic genes to gut epithelial cells. Safety studies indicate minimal off-target effects or immune activation, supporting phage therapy as a precision tool for gut-bone axis modulation [68, 123].
Future research should employ single-cell multi-omics to decipher cell-type-specific responses to microbial signals in bone marrow niches. Spatial transcriptomics of gut-bone organoids will elucidate how microbial metabolites directly influence osteocyte-osteoblast crosstalk [124]. Advanced organoid and microphysiological systems (e.g., gut-bone organ-on-a-chip) could model bidirectional crosstalk in a human-relevant context. Advanced imaging techniques can track metabolite distribution in bone tissue, while gnotobiotic models with humanized microbiota will provide physiologic relevance. Key unanswered questions include: How do microbial signals integrate with mechanical loading? Do osteocytes directly sense gut-derived metabolites? Resolving these mechanisms will unlock new therapeutic targets [94].
Precision interventions using metabolite analogs or receptor-specific modulators hold promise. AhR agonists (e.g., FICZ) and FXR antagonists (e.g., guggulsterone) show efficacy in preclinical models [125]. Synthetic biology approaches may engineer Lactobacillus strains to secrete PTH (1–34) or OPG in response to inflammation, creating “smart probiotics” for dynamic bone protection. Nanoparticle-based delivery of SCFAs to bone marrow via intestinal targeting could overcome bioavailability challenges [70].
Integration of multi-omics data with clinical parameters using machine learning algorithms will facilitate patient stratification and predictive biomarker identification [126]. Validation of microbial signatures or metabolite profiles (e.g., serum zonulin, fecal butyrate) could guide personalized nutritional or probiotic interventions [127].
Bacteriophage Therapy: Targeted depletion of pro-osteoclastic bacteria (e.g., C. bolteae, C. histolyticum) may reduce bone resorption with minimal microbiota disruption [128]. FMT: Standardized protocols and long-term outcomes in osteoporotic patients need evaluation in randomized controlled trials [120]. Gene and Cell Therapy: CRISPR-based editing of microbial or host genes (e.g., FTO SUMOylation sites) may correct aberrant bone remodeling in inflammatory settings [129].
Well-designed human trials are essential to validate efficacy and safety of microbiome-targeting interventions. Neurobiome-modulating approaches may synergize with existing therapies to address neurogenic components of bone loss. Harmonization of microbial analysis methods, intervention formulations, and outcome measures will facilitate cross-study comparisons and meta-analyses.
The gut-brain-bone axis, particularly autonomic nervous system mediation, represents an underexplored area. Vagal nerve stimulation or neurobiome-modulating approaches may offer novel therapeutic opportunities for metabolic bone diseases.
While promising, long-term consequences of profound microbiome alteration (e.g., via FMT or engineered bacteria) remain unknown. Potential risks include off-target immune effects, horizontal gene transfer, and metabolic disturbances [130]. Rigorous post-marketing surveillance and long-term animal studies are imperative to monitor for adverse effects such as abnormal bone remodeling patterns or hepatic steatosis.
The gut-bone axis must be understood within the broader framework of systemic immunometabolism [131]. Future studies should employ systems biology approaches to construct predictive models that integrate microbiome data with immune cell profiles, serum metabolomes, and bone turnover markers [132]. This will help identify master regulators of the axis and develop multi-target therapies that simultaneously address gut permeability, immune dysregulation, and metabolic dysfunction to achieve superior osteoprotection [132].
The gut-bone axis represents a sophisticated multidimensional system that integrates microbial, endocrine, immune, and neural signals to maintain skeletal homeostasis [103]. Rather than being a simple linear relationship, this axis comprises numerous feedback loops and cross-regulatory mechanisms that respond to nutritional status, inflammatory signals, and metabolic demands. The evidence summarized in this review supports a paradigm shift in which bone health is understood as a systemic outcome influenced significantly by gastrointestinal processes. Recent discoveries have illuminated several promising therapeutic directions. The identification of FTO SUMOylation as a key epigenetic mechanism in IBD-induced bone loss provides a novel target for intervention [85]. Similarly, the development of engineered tryptophan-producing bacteria offers an innovative approach to simultaneously address intestinal barrier dysfunction and bone loss [61]. The differential effects of gut hormones on bone metabolism—with GIP but not GLP-1 reducing bone resorption in PI-CF patients—suggests that precision approaches targeting specific hormonal pathways may yield better outcomes than broad-spectrum interventions [99]. Despite significant progress, important challenges remain in translating these findings into clinical practice. The strain-specific effects of probiotics, individual variability in microbiome composition, and complex dose-response relationships present obstacles to developing universally effective therapies [133]. Future research should focus on identifying specific microbial strains and consortia with optimal osteoprotective properties, developing targeted delivery systems for microbial metabolites, and conducting large-scale human trials to validate preclinical findings. While traditional calcium and vitamin D supplementation remains relevant, the evidence suggests that addressing microbial dysbiosis, intestinal inflammation, and gut barrier integrity may be equally important for preventing and treating bone disorders. As research in this field advances, targeting the gut-bone axis offers promise for developing more effective and comprehensive strategies to maintain skeletal health throughout the lifespan.
ANOVA, analysis of variance; SEM, structural equation modeling; GDP, gross domestic product; HLM, hierarchical linear modeling; HDAC, histone deacetylases; GPR, G protein-coupled receptor; RUNX2, Runt-related transcription factor 2; OVX, Ovariectomy; BV/TV, Bone volume per tissue volume; BMD, Bone Mineral Density; FXR, farnesoid X receptor; PKA, protein kinase A; IBD, inflammatory bowel disease; HR, hazard ratio; CYP1A1, Cytochrome P450 family 1 subfamily A member 1; TGR5, Takeda G protein-coupled Receptor 5; NFATc1, Nuclear Factor of Activated T-cells, Cytoplasmic 1; VDR, Vitamin D Receptor; cAMP, Cyclic Adenosine Monophosphate; FGF, Fibroblast Growth Factor; MK, Midkine; ucOC, Under-carboxylated Osteocalcin; Nrf2, nuclear factor erythroid 2–related factor 2; HO-1, Heme Oxygenase-1; NQO1, NAD(P)H: quinone oxidoreductase 1; PI-CF, Pancreatic Insufficient Cystic Fibrosis.
ZBS, ZJY: Conceptualization, Writing – Original Draft Preparation, Supervision. DJY: Data Curation, Formal Analysis, Investigation, Visualization. JJC, YLX: Validation, Resources, Writing – Review & Editing. GYW, Project Administration, Funding Acquisition, Supervision, Figures Production, References Collection. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work
Not applicable.
Not applicable.
We received fund from: The Affiliated Traditional Chinese Medicine Hospital of Southwest Medical University, Biomechanical research on cartilage transplantation based on the theory of “equal emphasis on muscle and bone”, 2023ZYQJ02.
The authors declare no conflict of interest.
During the preparation of this work the authors used ChatGPT-3.5 in order to check spell and grammar. After using this tool, the authors reviewed and edited the content as needed and takes full responsibility for the content of the publication.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.