







 , Mi Cheng 1,2,3,†, Quan-Long Wu 1,2, Ya-Ya Zhang 1,2, Yu Wang 1,2,4, Yan-Jiao Chen 1,2, Lei-Miao Yin 2,5, Yong-Qing Yang 1,2,*
, Mi Cheng 1,2,3,†, Quan-Long Wu 1,2, Ya-Ya Zhang 1,2, Yu Wang 1,2,4, Yan-Jiao Chen 1,2, Lei-Miao Yin 2,5, Yong-Qing Yang 1,2,* , Yu-Dong Xu 1,2,*
, Yu-Dong Xu 1,2,*1 Acupuncture Neurobiology Research Department, Yueyang Hospital of Integrated Chinese and Western Medicine, Shanghai University of Traditional Chinese Medicine, 200437 Shanghai, China
2 Shanghai Research Institute of Acupuncture and Meridian, Key Laboratory of Acupuncture and Molecular Biology, Shanghai University of Traditional Chinese Medicine, 200030 Shanghai, China
3 Department of Traditional Chinese Medicine, Zhabei Centre Hospital of Shanghai Jing’an District, 200070 Shanghai, China
4 Experimental Center for Science and Technology, Shanghai University of Traditional Chinese Medicine, 201203 Shanghai, China
5 Department of Acupuncture and Moxibustion, Shanghai Municipal Hospital of Traditional Chinese Medicine, Shanghai University of Traditional Chinese Medicine, 200071 Shanghai, China
†These authors contributed equally.
Abstract
Acupuncture has emerged as a promising complementary therapy for allergic asthma, but the molecular mechanisms underlying its therapeutic effects remain unclear. This study aimed to investigate whether Clara cell 10-kDa protein (CC10), an airway epithelial immunoregulatory protein, mediates the anti-inflammatory actions of acupuncture through the modulation of pulmonary dendritic cells (DCs).
We conducted a clinical study in patients with allergic asthma who underwent acupuncture at GV14 (Dazhui), BL12 (Fengmen), and BL13 (Feishu) acupoints to evaluate changes in asthma control, leukocyte counts, and serum CC10 levels. Parallelly, a house dust mite (HDM)-induced murine asthma model was used to examine the effects of acupuncture on airway hyperresponsiveness, pulmonary inflammation, T helper type 2 (Th2) cytokine production, and CC10 expression. To confirm the role of CC10, wild-type and CC10-deficient mice were compared, focusing on DC subsets analyzed by flow cytometry.
Clinically, acupuncture significantly improved Asthma Control Test (ACT) scores, reduced peripheral leukocyte counts, and elevated serum CC10 concentrations. Consistent with these clinical findings, acupuncture attenuated airway hyperresponsiveness, suppressed eosinophilic infiltration, downregulated Th2 cytokines, and restored pulmonary CC10 expression in HDM-challenged mice. Notably, these protective effects were largely abrogated in CC10-deficient mice, which displayed heightened airway inflammation, mucus hypersecretion, and enhanced Th2 responses. Mechanistic studies revealed that acupuncture reduced pro-inflammatory CD11b+ DCs in a CC10-dependent manner, thereby contributing to airway immune homeostasis.
Acupuncture attenuates allergic airway inflammation by upregulating CC10 and subsequently modulating pulmonary DC subsets, specifically CD11b+ DCs. These findings elucidate a novel mechanistic basis for the therapeutic efficacy of acupuncture and underscore CC10 as a viable therapeutic target for the management of allergic asthma.
NCT01931696, https://clinicaltrials.gov/study/NCT01931696.
Keywords
- asthma
- acupuncture therapy
- uteroglobin
- dendritic cells
- inflammation
Asthma is a chronic inflammatory disorder characterized by airway hyperresponsiveness (AHR), reversible airflow obstruction, and persistent airway inflammation predominantly driven by T helper type 2 (Th2) immune responses [1, 2]. Despite substantial advancements in pharmacological interventions, approximately 30–50% of patients continue to experience suboptimal symptom control and recurrent exacerbations, underscoring an urgent need for more effective and sustainable therapeutic modalities [3, 4].
Acupuncture, a prominent modality of traditional Chinese medicine, has attracted increasing clinical interest as a complementary approach for the management of allergic disorders, including asthma [5, 6]. Accumulating clinical and preclinical evidence indicates that acupuncture can enhance respiratory function and modulate immune responses [7, 8, 9]. Our previous study demonstrated that acupuncture at the GV14 (Dazhui), BL12 (Fengmen), and BL13 (Feishu) acupoints effectively suppressed Th2-driven airway inflammation by modulating pulmonary CD11b+ dendritic cells (DCs), a key cell subset involved in the initiation of Th2 immune responses [10]. These findings highlight DCs as critical cellular mediators of acupuncture-induced immunomodulation and raise important questions regarding the upstream molecular pathways that govern these effects.
Clara cell 10-kDa protein (CC10), also known as SCGB1A1 or uteroglobin, is an immunomodulatory secretory protein predominantly expressed by airway epithelial club cells [11]. CC10 exerts potent anti-inflammatory effects by suppressing Th2 cytokine secretion, reducing leukocyte infiltration, and maintaining epithelial integrity [12, 13, 14]. Notably, CC10 expression is significantly decreased in allergic asthma, suggesting that its deficiency may contribute to asthma pathogenesis [15, 16, 17]. Furthermore, our previous investigations have demonstrated that recombinant CC10 administration attenuates allergic airway inflammation through the modulation of DC function and inhibition of Th2 immune responses [13], thereby establishing a mechanistic link between CC10 and DC-mediated immunoregulation in asthma.
Building on this evidence, we hypothesized that CC10 acts as a critical molecular mediator through which acupuncture exerts its anti-inflammatory effects, particularly by modulating pulmonary DC activity. To test this hypothesis, we first evaluated the effects of acupuncture on serum CC10 levels and clinical outcomes in patients with allergic asthma. We then employed a house dust mite (HDM)-induced murine asthma model to investigate the effects of acupuncture on airway inflammation and CC10 expression. Finally, using CC10 knockout mice, we directly examined the indispensable role of endogenous CC10 in mediating the therapeutic efficacy of acupuncture, with a focus on Th2 immune responses and pulmonary DC modulation. Together, this study provides novel mechanistic insights into acupuncture-mediated regulation of airway immunity, identifying CC10 as a key molecular effector underlying its therapeutic benefits in allergic asthma.
A total of forty patients with a diagnosis allergic asthma were enrolled in this study. The clinical study was conducted in accordance with the protocol previously published by our team [18]. Briefly, patients received acupuncture treatment every other day over a 6-week period, comprising a total of 20 treatment sessions. The selected acupoints included GV14, bilateral BL12, and bilateral BL13. Following needle insertion, standardized manual stimulation was consistently applied, consisting of twisting and lifting–thrusting manipulations for 20 seconds every 10 minutes. Each acupuncture session lasted 30 minutes, after which the needles were carefully withdrawn. In the current mechanistic study, peripheral blood samples were collected from a subset of patients assigned to the acupuncture treatment arm both before and after the intervention. Collected blood samples were centrifuged, and the resulting serum was aliquoted and stored at –80 °C until further analysis of inflammatory molecules, including CC10. Asthma control status was assessed using the validated Asthma Control Test (ACT) questionnaire, while peripheral blood leukocyte counts were measured by routine hematological analyses. This clinical study was prospectively registered (ClinicalTrials.gov identifier: NCT01931696, registered on 26 August 2013) and approved by Yueyang Hospital, affiliated with Shanghai University of Traditional Chinese Medicine (approval number: 2013–041). All participants provided written informed consent prior to study initiation.
Female C57BL/6 wild-type (Cc10WT) mice and CC10 knockout (Cc10-/-) mice, generated using CRISPR/Cas9 gene-editing technology by the Shanghai Model Organisms Center on a C57BL/6 background, aged 6–8 weeks, were used in this study. All animal procedures were reviewed and approved by the Institutional Animal Care and Use Committee of Shanghai University of Traditional Chinese Medicine (Approval no. PZSHUTCM2309080009). All animal experiments were performed and reported in accordance with the ARRIVE 2.0 guidelines. Animals were maintained under specific pathogen-free conditions with a controlled 12-hour light-dark cycle, and were provided ad libitum access to standard rodent chow and water.
Mice were randomly assigned to four groups: PBS group (control mice sensitized and challenged with PBS), HDM group (asthmatic mice sensitized and challenged with HDM), HDM+Acu (asthmatic mice treated with acupuncture), and PBS+Acu (control mice treated with acupuncture). A murine model of allergic asthma was established as previously described [13]. Briefly, mice were intranasally sensitized with 10 µg HDM extract (Cat. no. 322781, Greer Laboratories, Lenoir, NC, USA) on days 0 and 7. Subsequently, from day 14 to day 18, mice underwent daily intranasal challenges with 20 µg HDM. Control animals received an equivalent volume of PBS at corresponding time points.
All acupuncture procedures were performed under inhalational anesthesia using 2% isoflurane to minimize animal stress. Beginning immediately after the sensitization phase, acupuncture was applied every other day for two consecutive weeks. Mice in the HDM+Acu and PBS+Acu groups received manual acupuncture at acupoints GV14, bilateral BL12, and bilateral BL13, localized according to murine anatomical analogs of the human World Health Organization (WHO) Standard Acupuncture Point Locations. Disposable stainless-steel needles (0.30 mm in diameter and 13 mm in length; Suzhou Medical Appliance Factory, Suzhou, China) were inserted perpendicularly to a depth of approximately 2.5–3 mm. Immediately upon insertion, needles were subjected to standardized manual manipulation, consisting of bidirectional rotations (360°, 200 rotations per minute) for 30 seconds continuously. This manipulation was repeated every 3 minutes, with a total needle retention time of 30 minutes per session. In contrast, animals in the PBS and HDM groups underwent identical anesthesia and handling without needle insertion. All acupuncture procedures were performed by the same licensed acupuncturist, who was extensively trained in this protocol and blinded to the group assignments.
AHR was assessed 24 hours after the final HDM challenge (day 20) using the FinePointe RC system (Data Sciences International, St. Paul, MN, USA). Mice were anesthetized by intraperitoneal injection of 1% pentobarbital sodium (10 mL/kg, Cat. no. P3761, Sigma-Aldrich, St. Louis, MO, USA). Following surgical tracheotomy, an 18-gauge catheter was inserted into the trachea and connected to a computer-controlled mechanical ventilator set at 140 breaths/min, a tidal volume of 0.2 mL, and a positive end-expiratory pressure (PEEP) of 2 cm H₂O.
After a 5-minute stabilization period with aerosolized PBS, animals were
sequentially exposed to increasing concentrations of aerosolized methacholine
(MCh; 0, 1.5, 3, 6, and 12 mg/mL; Cat. no. A6625, Sigma-Aldrich, St. Louis, MO,
USA) using an integrated ultrasonic nebulizer (Data Sciences International, St.
Paul, MN, USA). Airway resistance (RL) was continuously recorded
for 3 minutes following MCh administration at each concentration.
RL values, expressed as cmH₂O
Following AHR assessment, mice were humanely euthanized by intraperitoneal
overdose of pentobarbital sodium (
Lung tissues were harvested, fixed in 4% paraformaldehyde, paraffin-embedded, and sectioned at a thickness of 4 µm. To assess airway inflammation, sections were stained with hematoxylin and eosin (H&E). The following standardized semi-quantitative scoring system, based on the number of cell layers surrounding the bronchi and blood vessels was used to evaluate inflammatory cell infiltration: 0 = no inflammatory cells; 1 = few scattered cells; 2 = 1–2 cell layers; 3 = 3–4 cell layers; and 4 = more than 4 cell layers. Goblet cell metaplasia in the airway epithelia was evaluated using periodic acid-Schiff (PAS) staining. The percentage of PAS-positive goblet cells relative to the total number of epithelial cells was quantified in 5–10 randomly selected airway regions per mouse. All histological evaluations were independently performed by two experienced pathologists blinded to group allocation to minimize observer bias. Microscopic images were captured using a Nikon 80i light microscope (Nikon Corporation, Tokyo, Japan).
Lung tissue sections were deparaffinized, rehydrated, and subjected to antigen retrieval in 10 mM sodium citrate buffer (pH 6.0) at 95 °C for 20 minutes. Endogenous peroxidase activity was quenched with 3% hydrogen peroxide for 10 minutes, followed blocking with 5% normal goat serum to prevent non-specific binding. Sections were subsequently incubated overnight at 4 °C with a rabbit anti-CC10 primary antibody (Cat. no. ab40873, Abcam, Cambridge, UK; diluted 1:1000 in PBS supplemented with 1% BSA). After washing, sections were incubated with HRP-conjugated secondary antibody (Cat. no. sc-2004, Santa Cruz Biotechnology, Dallas, TX, USA) for 30 minutes at room temperature. Immunoreactivity was visualized using DAB substrate (Cat. no. P0203, Beyotime Biotechnology, Shanghai, China) and counterstained with hematoxylin. Images were acquired using a Nikon 80i light microscope (Nikon Corporation, Tokyo, Japan), and quantification of CC10-positive areas was performed using ImageJ software (version 1.53, National Institutes of Health, Bethesda, MD, USA).
Following euthanasia, spleens were excised and immediately weighed. The spleen index was determined by calculating the ratio of spleen weight (mg) to body weight (g).
Serum levels of HDM-specific IgE were determined by enzyme-linked immunosorbent assay (ELISA) using a modified protocol based on a commercial kit (Cat. no. 432404, BioLegend, San Diego, CA, USA). Briefly, 96-well plates were coated overnight at 4 °C with a capture antibody, followed by blocking with assay buffer. Diluted serum samples were incubated for 2 hours at room temperature. Subsequently, biotinylated HDM extract (Cat. no. 02.01.88, CITEQ, Quebec City, QC, Canada) was applied for 1 h, followed by incubation with streptavidin-HRP conjugate. Colorimetric development was initiated using TMB substrate, and the reaction was terminated with 1 M H₂SO₄. Absorbance was measured at 450 nm using a microplate reader (Synergy H1, BioTek Instruments, Winooski, VT, USA). HDM-specific IgE levels were expressed as optical density (OD) values.
Levels of interleukin (IL)-4, IL-5, and IL-13 in BALF and mediastinal lymph node (MLN) cell culture supernatants were measured using commercial ELISA kits. Specifically, IL-4 and IL-5 levels were assessed using BioLegend kits (Cat. nos. 431101 and 431201, respectively; BioLegend, San Diego, CA, USA), while IL-13 was quantified using an Invitrogen kit (Cat. no. 88-7137-88, Thermo Fisher Scientific, Waltham, MA, USA). All assays were performed strictly in accordance with the manufacturers’ instructions.
Total RNA was extracted from lung tissues using TRIzol reagent (Cat. no. 15596026, Thermo Fisher Scientific, Waltham, MA, USA) in accordance with the manufacturer’s guidelines. Subsequently, 1 µg of total RNA was reverse-transcribed into complementary DNA (cDNA) using a reverse transcription kit (Cat. no. K1622, Thermo Fisher Scientific, Waltham, MA, USA). qRT-PCR was performed on a LightCycler 96 System (Roche, Switzerland) using SYBR Green PCR Master Mix (Cat. no. QPK-201, TOYOBO, Osaka, Japan). The relative mRNA expression of the target genes, Cc10 and Muc5ac, was normalized to the housekeeping gene Gapdh using the 2-ΔΔCt method. Primer sequences are detailed in Supplementary Table 1.
Lung tissues were homogenized in RIPA lysis buffer (Cat. no. P0039, Beyotime
Biotechnology, Shanghai, China) supplemented with protease and phosphatase
inhibitors (Cat. no. 04693132001, Roche Diagnostics, Mannheim, Germany). Protein
concentrations were quantified using a BCA assay, and equal amounts of total
protein (20 µg) were separated by SDS-PAGE and transferred onto PVDF
membranes. Membranes were blocked for 1 hour at room temperature with 5% non-fat
milk in Tris-buffered saline containing 0.1% Tween-20 (TBST). Membranes were
incubated overnight at 4 °C with primary antibodies against CC10
(1:1000, Cat. no. ab40873, Abcam, Cambridge, UK) and
Lung single-cell suspensions were prepared by mechanical dissociation followed by enzymatic digestion in RPMI 1640 medium supplemented with 1 mg/mL collagenase IV and 200 U/mL DNase I at 37 °C for 45 minutes. The digested tissues were filtered through a 70 µm cell strainer, and red blood cells were removed using an RBC lysis buffer. Cells were then stained with fluorochrome-conjugated antibodies against CD45, CD11c, MHC II, CD11b, and CD103 (BioLegend and BD Biosciences). Dead cells were excluded using a fixable viability dye (Cat. no. 65-0865-18, Thermo Fisher Scientific, San Diego, CA, USA). Detailed information on all antibodies used for flow cytometry is provided in Supplementary Table 2. Data were acquired on an Attune NxT flow cytometer (Thermo Fisher Scientific, Waltham, MA, USA), and analyzed using FlowJo software (version 10.9, BD Biosciences, Ashland, OR, USA). DCs were identified as live CD45+CD11c+MHC II+ cells, and DC subsets were defined based on differential expression of CD11b and CD103. Dimensionality reduction and subset visualization were performed using t-distributed stochastic neighbor embedding (t-SNE).
All data are presented as mean
To investigate the clinical relevance of CC10 and the effects of acupuncture in
patients with allergic asthma, we first measured serum CC10 concentrations,
asthma symptom control, and peripheral blood leukocyte counts before and after
acupuncture treatment. As shown in Fig. 1A,B, acupuncture significantly increased
serum CC10 levels from 16.90
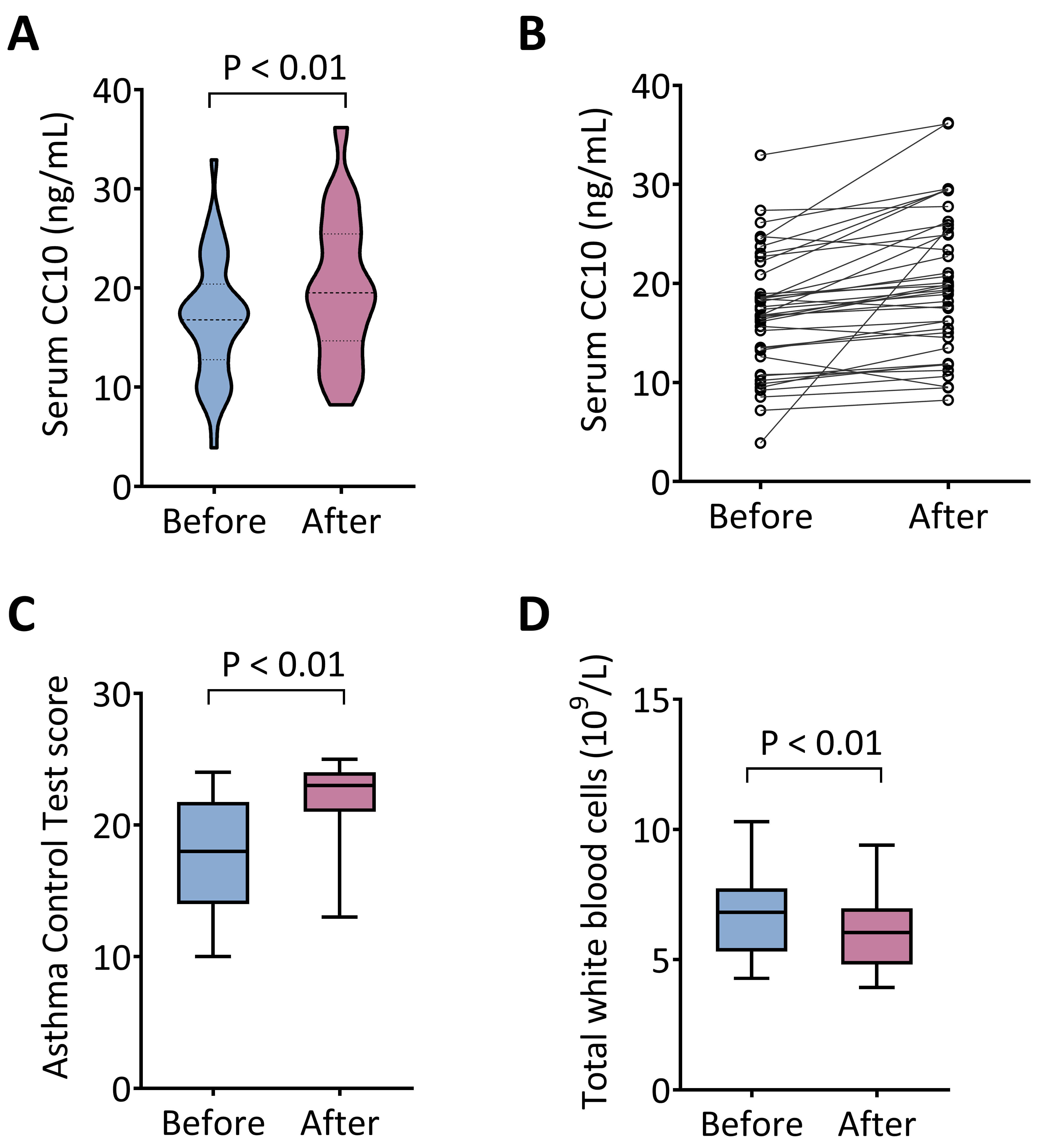 Fig. 1.
Fig. 1.
Acupuncture elevates serum CC10 levels, improves asthma control,
and reduces peripheral leukocyte counts in allergic asthma patients. (A,B) Serum
CC10 concentrations were significantly elevated post-acupuncture treatment (n =
40). (C) Asthma symptom control, measured by Asthma Control Test (ACT) scores,
significantly improved following acupuncture. (D) Total peripheral white blood
cell counts significantly decreased after acupuncture treatment. Data are
expressed as mean
To further elucidate the mechanisms underlying the therapeutic effects of acupuncture, we employed a murine model of HDM-induced allergic asthma (Fig. 2A). As expected, HDM sensitization and subsequent challenge significantly enhanced AHR, evidenced by increased RL in response to increasing doses of MCh compared with PBS-treated controls (Fig. 2B,C). Notably, acupuncture treatment markedly ameliorated HDM-induced AHR, resulting in a pronounced reduction in RL toward baseline levels.
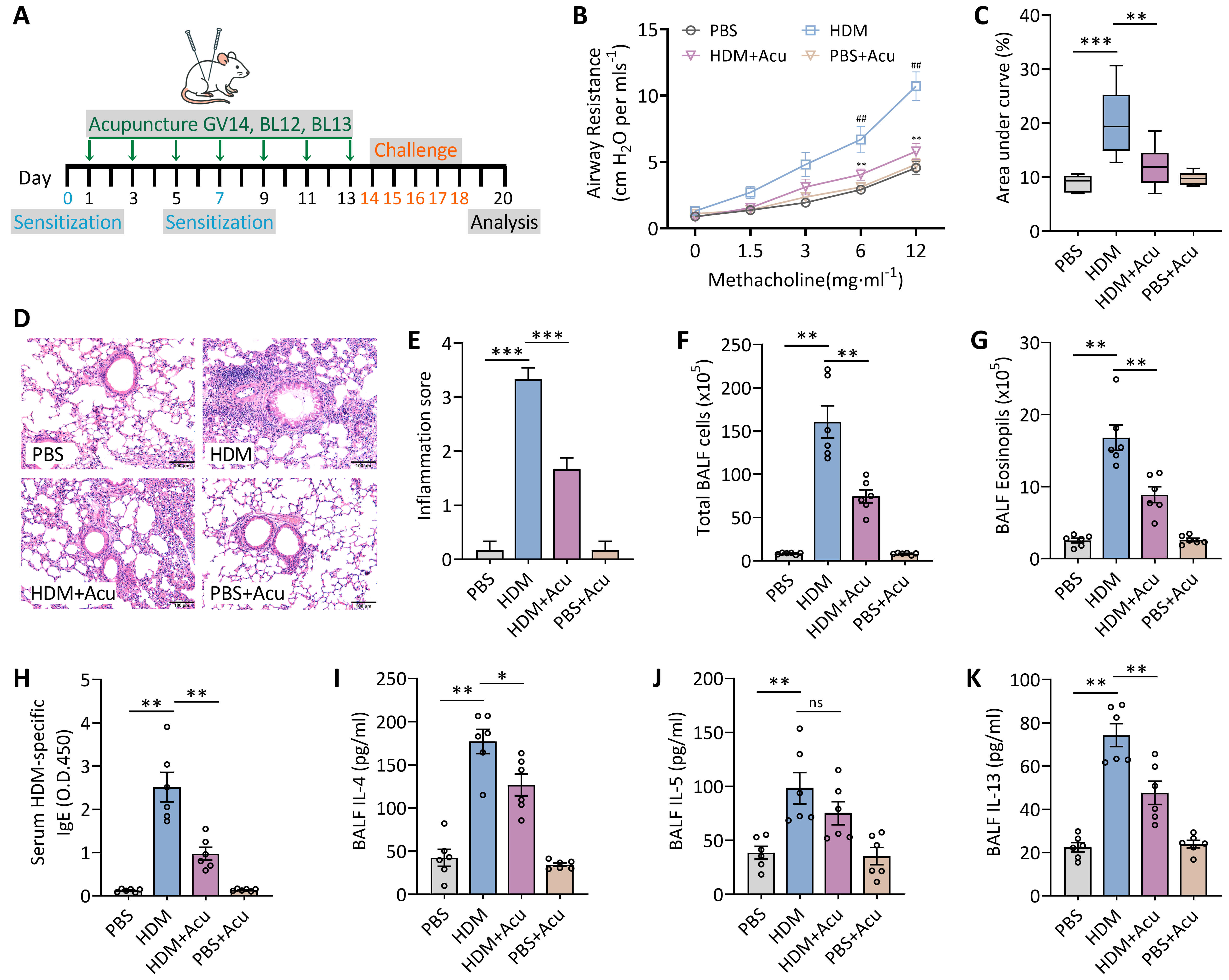 Fig. 2.
Fig. 2.
Acupuncture reduces airway hyperresponsiveness and allergic
airway inflammation in an HDM-induced asthma mouse model. (A) Experimental
timeline illustrating sensitization and challenge with HDM or PBS, with
concurrent acupuncture treatments at GV14, BL12, and BL13 on the indicated days.
(B) Airway resistance (RL) measured in response to increasing doses
of methacholine (0–12 mg/mL), normalized to baseline values. (C) Area under the
curve (AUC) analysis derived from RL data in panel (B). (D)
Representative hematoxylin and eosin (H&E) staining of lung sections. Scale bars = 100 µm. (E) Quantitative inflammation scores from
H&E-stained lung sections. (F) Total leukocyte counts in BALF. (G) Eosinophil
counts in BALF. (H) Serum levels of HDM-specific IgE. (I–K) Concentrations of
Th2 cytokines IL-4 (I), IL-5 (J), and IL-13 (K) in BALF. Data represent mean
Histopathological examination using H&E staining revealed extensive peribronchial and perivascular inflammatory cell infiltration, accompanied by pronounced airway epithelial damage in HDM-challenged mice. Acupuncture treatment markedly alleviated inflammatory cell infiltration and facilitated epithelial repair, thereby preserving airway epithelial integrity (Fig. 2D,E). Consistent with these histological findings, acupuncture significantly decreased both the elevated total leukocyte and eosinophil counts in BALF induced by HDM exposure (Fig. 2F,G). Additionally, serum HDM-specific IgE levels, a hallmark of allergic sensitization, were substantially increased following HDM challenge but were significantly reduced after acupuncture intervention (Fig. 2H).
Further analysis demonstrated that HDM exposure markedly increased the levels of Th2-associated cytokines, including IL-4, IL-5, and IL-13, in BALF. Acupuncture treatment robustly suppressed the production of these cytokines (Fig. 2I–K). In summary, these results demonstrate that acupuncture effectively attenuates airway hyperresponsiveness and suppresses allergic airway inflammation in the murine model of HDM-induced asthma.
Considering the clinically observed elevation in serum CC10 levels following acupuncture treatment in allergic asthma patients, we next investigated whether acupuncture similarly modulates pulmonary CC10 expression in the murine model of HDM-induced asthma. Immunohistochemical staining revealed a marked reduction in CC10 protein expression within airway epithelial cells of HDM-challenged mice compared with PBS-treated control mice. Importantly, acupuncture intervention effectively restored CC10 expression, approaching levels observed in control animals (Fig. 3A). Consistent with these histological findings, qRT-PCR analysis demonstrated a significant downregulation of pulmonary Cc10 mRNA expression in HDM-induced asthmatic mice, which was robustly reversed following acupuncture treatment (Fig. 3B). Moreover, Western blot analysis further confirmed a pronounced decrease in CC10 protein levels in lung homogenates from asthmatic mice, whereas acupuncture effectively normalized CC10 expression (Fig. 3C,D). These findings suggest that HDM-induced asthma suppresses CC10 expression in the lung, and that acupuncture exerts a protective effect, at least in part, by restoring CC10 production within the airway epithelium.
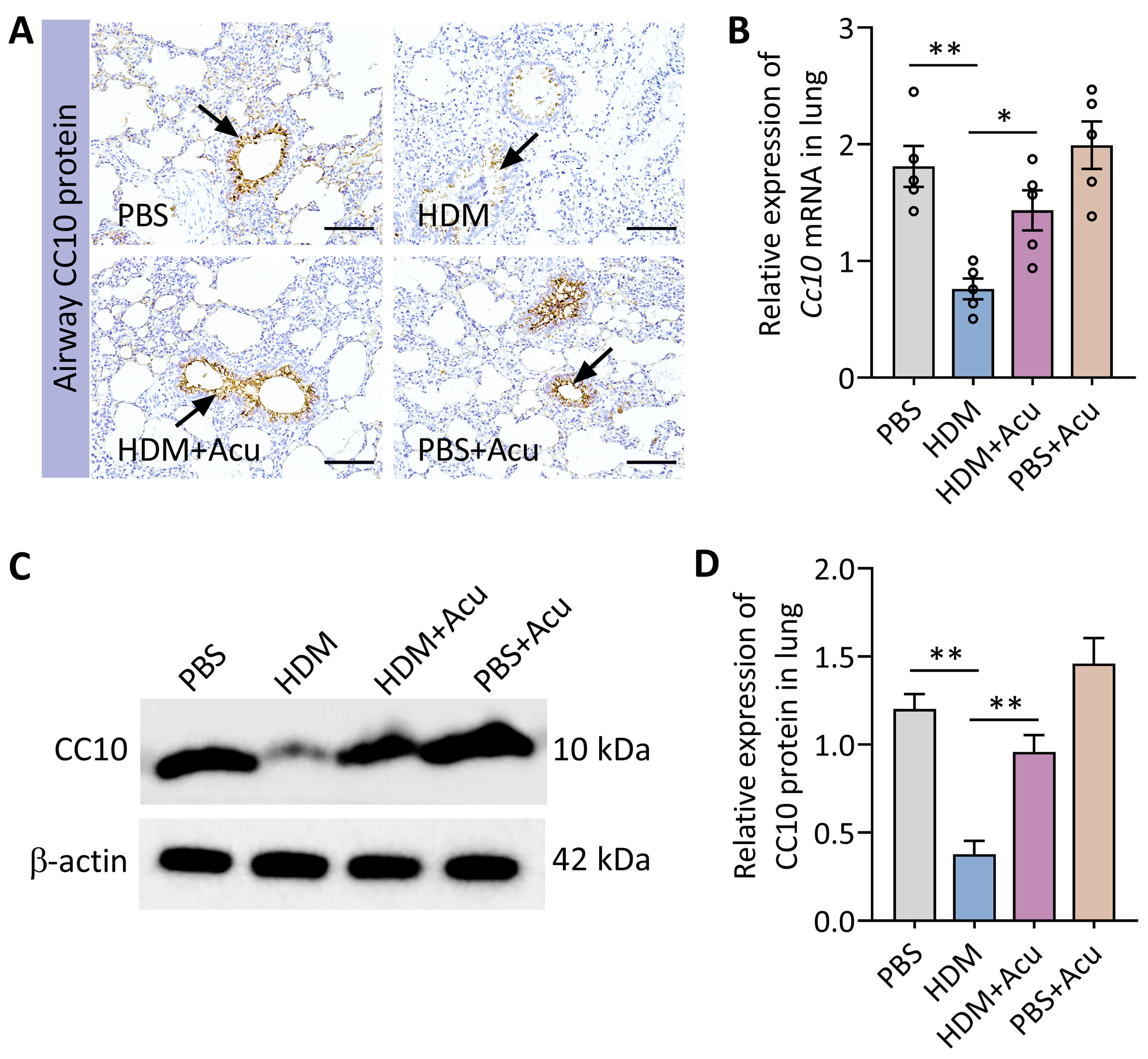 Fig. 3.
Fig. 3.
Acupuncture restores pulmonary CC10 expression in HDM-induced
asthmatic mice. (A) Immunohistochemical staining illustrating CC10 protein
expression (arrows) in airway epithelial cells. Scale bars = 50 µm. (B)
Relative pulmonary Cc10 mRNA expression (n = 5 per group) quantified by
quantitative Real-Time PCR (qRT-PCR). (C) Representative Western blot depicting
CC10 protein expression in lung homogenates. (D) Densitometric quantification of
CC10 protein normalized to
To define the indispensable role of CC10 in mediating the anti-asthmatic effects of acupuncture, we employed Cc10 knockout mice (Cc10-/-) and their wild-type littermates (Cc10WT) in an HDM-induced allergic asthma model, with or without acupuncture intervention. As shown in Fig. 4A,B, Cc10-/- mice exhibited significantly aggravated AHR following HDM challenge, as indicated by increased RL and elevated AUC values compared with HDM-challenged Cc10WT mice. Although acupuncture robustly attenuated AHR in Cc10WT mice, this protective effect was substantially blunted in Cc10-/- mice.
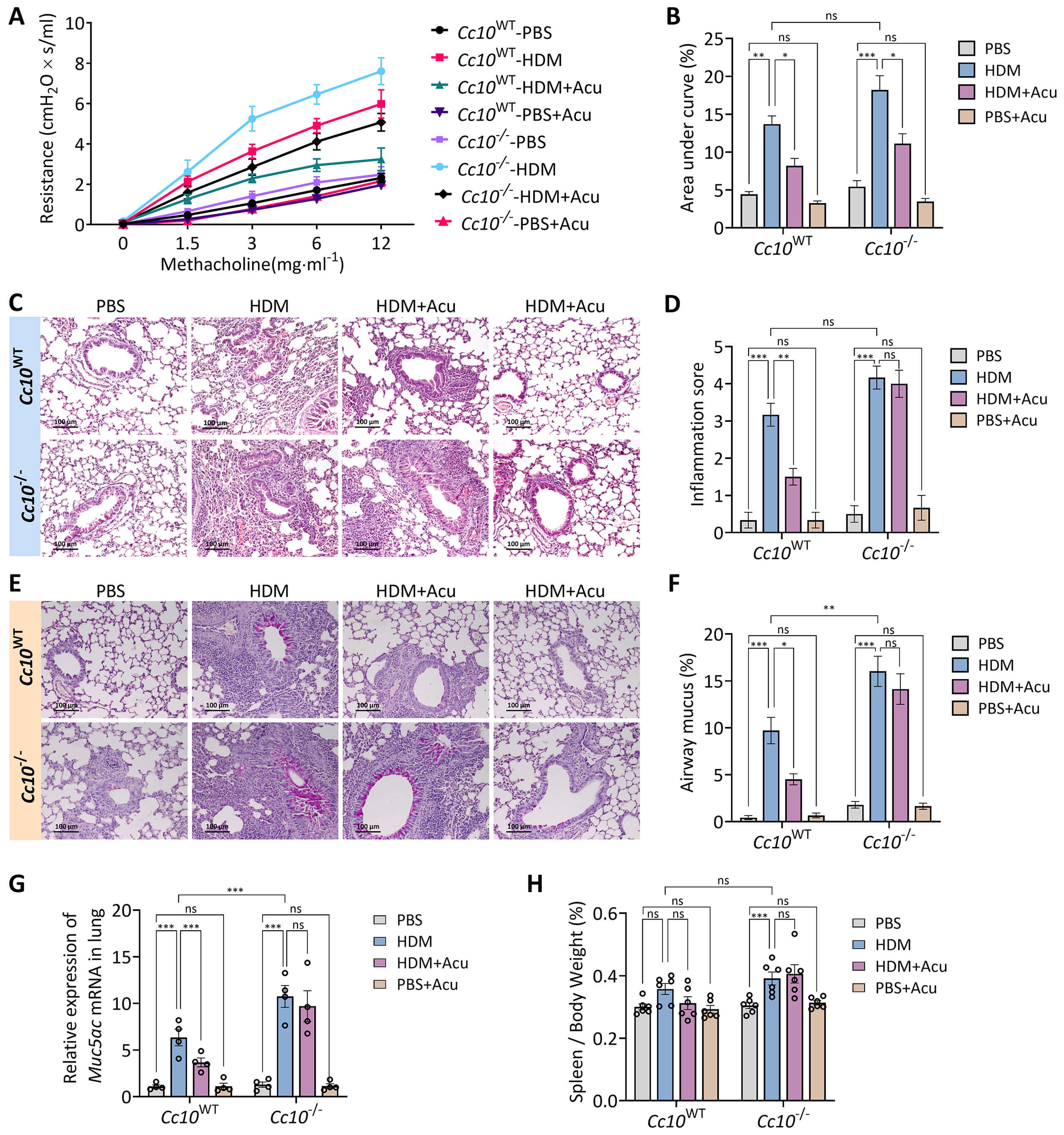 Fig. 4.
Fig. 4.
CC10 deficiency exacerbates asthma severity and reduces the
therapeutic efficacy of acupuncture. (A) Airway resistance (RL)
response curves to escalating methacholine doses (0–12 mg/mL). (B) Corresponding
area under the curve (AUC) analysis of RL data from panel (A). (C)
Representative H&E staining of lung sections illustrating airway inflammation.
Scale bars = 100 µm. (D) Quantitative inflammation scores derived from
H&E-stained sections. (E) Representative periodic acid-Schiff (PAS) staining of
lung sections depicting goblet cell metaplasia. Scale bars = 100 µm. (F)
Quantification of PAS-positive goblet cells as a percentage of total airway
epithelial cells. (G) Relative expression of Muc5ac mRNA in lung tissues
measured by qRT-PCR. (H) Spleen index expressed as spleen-to-body weight ratio.
Data represent mean
Histopathological examination further revealed that CC10 deficiency substantially intensified pulmonary inflammation (Fig. 4C,D) as well as airway goblet cell metaplasia, as demonstrated by PAS staining and subsequent quantification (Fig. 4E,F). Correspondingly, pulmonary Muc5ac mRNA expression, indicative of mucus hypersecretion, was significantly increased in asthmatic Cc10-/- mice compared with Cc10WT asthmatic mice (Fig. 4G). Notably, the acupuncture-mediated attenuation of inflammation and mucus hypersecretion observed in Cc10WT mice was markedly diminished or completely abolished in Cc10-/- mice. Additionally, the spleen index, reflecting systemic inflammatory burden, was significantly elevated in Cc10-/- asthmatic mice, with acupuncture’s protective effect substantially diminished (Fig. 4H). Taken together, these results underscore the indispensable role of CC10 in mediating the protective effects of acupuncture against airway hyperresponsiveness, inflammation, and mucus hypersecretion.
We next assessed the impact of CC10 deficiency on allergic airway inflammation, with particular focus on Th2 immune responses and the immunomodulatory effects of acupuncture. Analysis of BALF revealed markedly increased total leukocyte counts and eosinophil proportions in asthmatic Cc10-/- mice compared with their asthmatic Cc10WT counterparts. Importantly, the inhibitory effect of acupuncture on inflammatory cell infiltration was substantially diminished in the absence of CC10 (Fig. 5A).
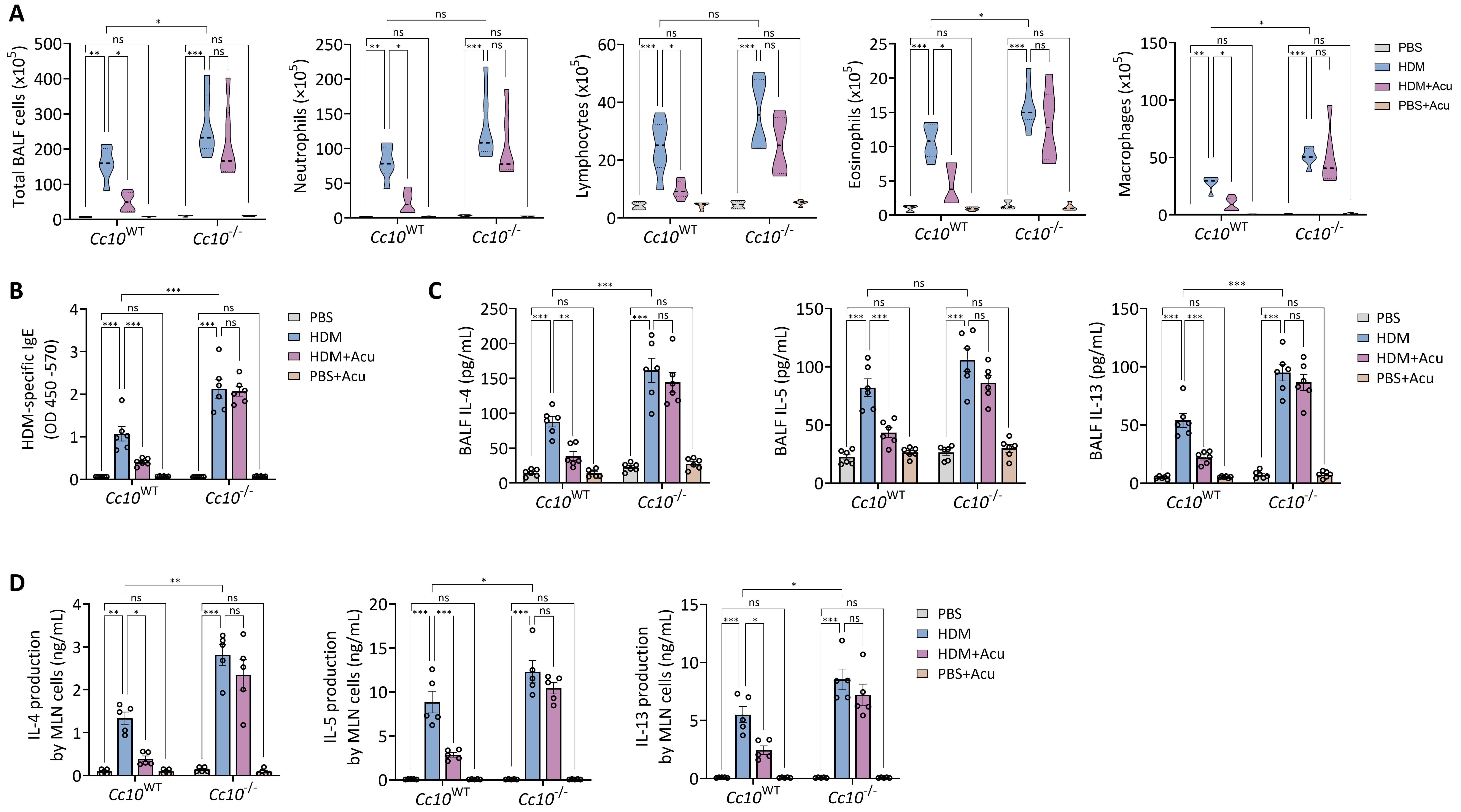 Fig. 5.
Fig. 5.
CC10 deficiency impairs acupuncture-mediated suppression of
airway Th2 inflammation. (A) Total leukocyte counts and differential analysis of
neutrophils, lymphocytes, eosinophils, and macrophages in BALF. (B) Serum
HDM-specific IgE levels. (C) Th2 cytokines (IL-4, IL-5, IL-13) quantified in
BALF. (D) Th2 cytokine levels (IL-4, IL-5, IL-13) measured in culture
supernatants from mediastinal lymph node (MLN) cells stimulated ex vivo with HDM
for 72 hours. Data represent mean
In parallel, serum levels of HDM-specific IgE were significantly elevated in Cc10-/- asthmatic mice. While acupuncture treatment effectively reduced HDM-specific IgE concentrations in Cc10WT mice, this therapeutic benefit was largely impaired in Cc10-/- mice (Fig. 5B). Furthermore, levels of the Th2-associated cytokines IL-4, IL-5, and IL-13 in BALF were profoundly increased in Cc10-/- asthmatic mice, indicative of exacerbated Th2-driven inflammation. Importantly, acupuncture-mediated suppression of these cytokines was significantly weakened or completely abolished in the absence of CC10 (Fig. 5C). Consistent with these in vivo findings, ex vivo stimulation of MLN cells with HDM extract resulted in enhanced Th2 cytokine production in cells derived from Cc10-/- mice, and acupuncture treatment exhibited limited capacity to modulate this exaggerated response (Fig. 5D). Together, these results strongly support a pivotal role of CC10 in restraining Th2-mediated inflammation and highlight its indispensability for the full immunomodulatory effects of acupuncture.
Building on our previous findings that acupuncture suppresses airway Th2 inflammation through modulation of pulmonary CD11b+ DCs, we further investigated whether CC10 serves as the molecular mediator of these regulatory effects. Flow cytometric analysis was performed using a refined gating strategy to identify total DC populations (CD45+CD11c+MHCII+ live cells) and their subsets (Fig. 6A). Although the overall frequency of pulmonary DCs remained unaffected by either CC10 deficiency or acupuncture treatment (Fig. 6B), distinct phenotypic shifts were observed within specific DC subpopulations.
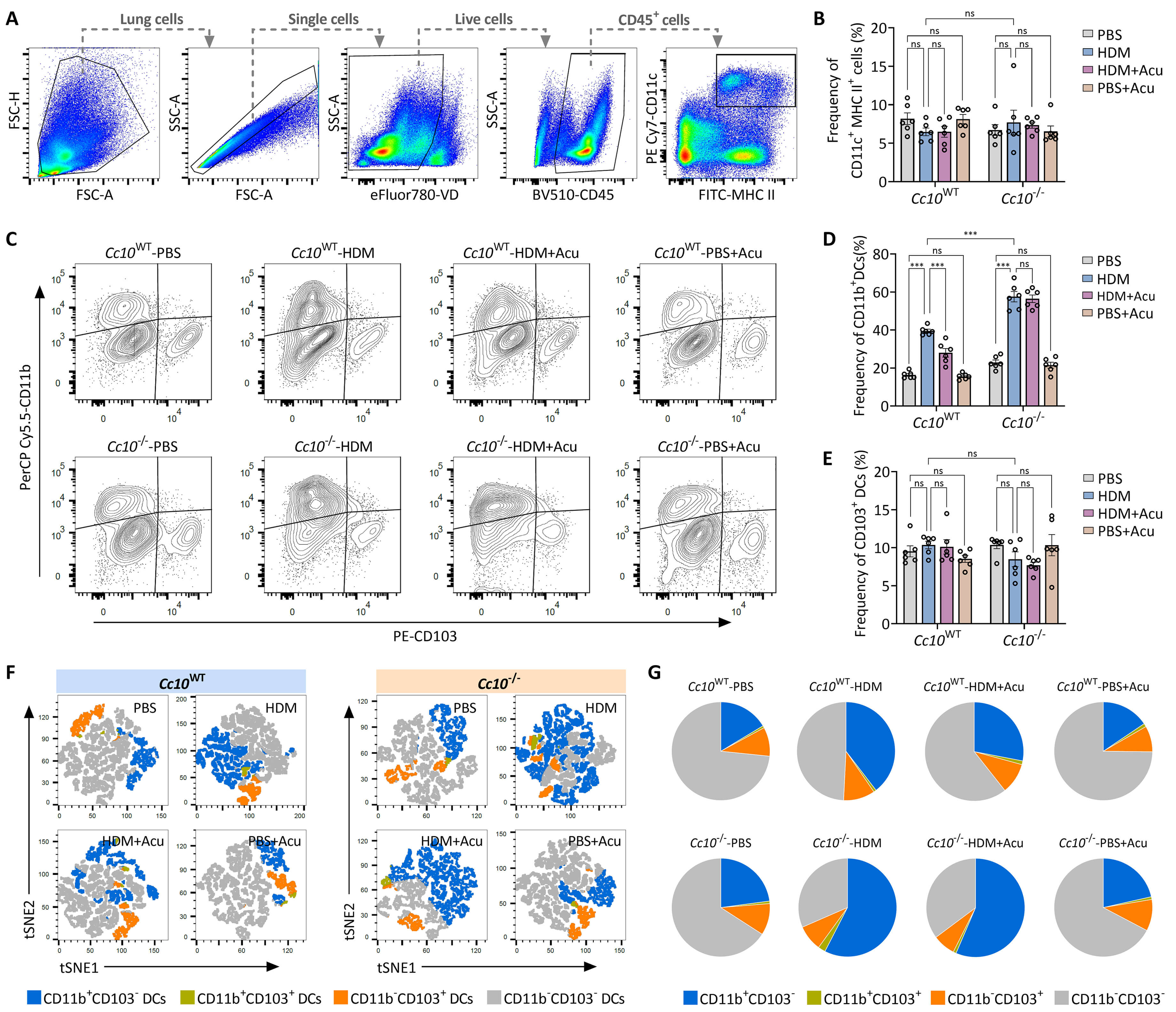 Fig. 6.
Fig. 6.
Acupuncture modulates pulmonary CD11b+ dendritic cells via
CC10. (A) Flow cytometric gating strategy identifying pulmonary DCs. Cells were
gated sequentially as total lung cells, single cells, live cells, CD45+ cells,
and further defined as CD11c+MHCII+ DCs. (B) Frequency of total pulmonary DCs
(CD11c+MHCII+CD45+ live cells). (C) Representative flow cytometry plots showing
frequencies of pulmonary CD11b+ DCs (CD11b+CD103⁻) and CD103+ DCs (CD11b⁻CD103+),
gated on live CD45+CD11c+MHCII+ cells. (D,E) Quantification of frequencies of
CD11b+ DCs (D) and CD103+ DC subsets (E). (F) t-Distributed Stochastic Neighbor
Embedding (t-SNE) visualization of pulmonary DC subset distributions,
illustrating distinct clusters identified by flow cytometry. Each color indicates
a unique DC subset. (G) Pie charts summarizing compositional proportions of
pulmonary DC subsets. Data are expressed as mean
Notably, the CD11b+ DC subset (CD11b+CD103–), a key driver of Th2-type immunity, was significantly expanded in asthmatic Cc10-/- mice compared to their Cc10WT counterparts. Acupuncture significantly reduced the frequency of CD11b+ DCs in Cc10WT mice; however, this suppressive effect was markedly diminished in CC10-deficient mice (Fig. 6C,D). In contrast, the CD103+ DC subset (CD11b–CD103+), typically associated with Th1 immune responses, exhibited differential patterns (Fig. 6E).
Further dimensionality reduction using t-SNE analysis provided an integrated visualization of pulmonary DC subset distributions. Acupuncture substantially contracted the CD11b+ DC clusters (blue) in Cc10WT mice (Fig. 6F); conversely, this subset rebalancing was notably impaired in Cc10-/- mice. This observation was further substantiated by pie chart analysis (Fig. 6G), which revealed a dominant expansion of CD11b+ DCs in Cc10-/- mice that remained refractory to acupuncture treatment. Overall, these results indicate that CC10 is indispensable for acupuncture-induced immunoregulation via the modulation of CD11b+ DC expansion, thereby providing a clear mechanistic link between acupuncture and airway immune homeostasis.
This study provides integrated clinical and experimental evidence that acupuncture alleviates allergic airway inflammation through a CC10-dependent mechanism involving the modulation of the pulmonary CD11b+ DCs. By utilizing both clinical subjects and a murine of HDM-induced asthma model, we showed that acupuncture enhances CC10 expression, dampens Th2-dominated inflammation, and reconfigures pulmonary DC subset composition. Crucially, we found that these protective effects of acupuncture were largely diminished in CC10-deficient mice, thus identifying CC10 as an indispensable molecular mediator linking acupuncture stimulation to immune regulation in allergic asthma. Our findings elucidate a critical mechanistic pathway, bridging the gap between the established clinical efficacy of acupuncture and its underlying molecular and cellular mechanisms.
The acupoint combination of GV14, BL12, and BL13 was selected based on established traditional Chinese medicine principles [19] and supported by contemporary biomedical evidence demonstrating immunoregulatory effects in well-validated asthma models, with anatomical distribution along the spinal and paraspinal regions associated with pulmonary innervation and sympathetic outflow [20, 21]. In the present study, the primary aim was not to evaluate acupoint specificity, but to characterize downstream molecular and cellular events engaged by a clinically validated acupuncture regimen [7]. To partially account for potential nonspecific effects of needling, a normal acupuncture control group (PBS+Acu) was included to assess acupuncture responses under non-inflammatory conditions. The finding that no significant inflammatory alterations occurred in this control group, compared with PBS controls, suggests that the immunomodulatory effects observed in asthmatic mice are largely disease-context dependent, although contributions from generalized needling-induced activation cannot be fully excluded. We further acknowledge that the absence of sham or non-acupoint stimulation controls limits a direct evaluation of acupoint-specific effects, a recognized challenge in acupuncture research given the lack of consensus on a biologically inert sham control. Accordingly, the observed immunomodulatory effects should be interpreted as demonstrating a CC10-dependent immunomodulatory mechanism within an established acupoint regimen rather than definitive evidence of acupoint specificity. Future investigations incorporating alternative-site stimulation or rigorously defined sham controls will be necessary to more conclusively address this question.
Our clinical observations align closely with previous reports of documenting the therapeutic efficacy of acupuncture in asthma management [22, 23]. Acupuncture-treated patients who exhibited improved asthma control, reduced peripheral leukocyte counts, and notably elevated serum CC10 levels. CC10, a club cell-derived epithelial protein characterized by its potent anti-inflammatory [24], immunosuppressive [25], and epithelial-protective properties [14, 26], is consistently downregulated in asthma conditions [15, 27]. Extending these observations, our experimental data demonstrate that acupuncture effectively restore CC10 expression in the airway epithelium of asthmatic mice, thereby facilitating inflammation resolution and the restoration of airway homeostasis.
At the cellular level, our data indicate that CC10 contributes to the regulation of pulmonary DC subsets during acupuncture treatment. Acupuncture significantly decreased the frequency of pulmonary CD11b+ DCs in wild-type asthmatic mice, whereas this suppressive effect was substantially attenuated in CC10 deficient animals. Given that CD11b+ DCs are recognized as a predominant subset driving allergen-induced Th2 polarization [28, 29, 30, 31] and that CC10 can modulate their phenotype profile and function [13], our findings suggest that acupuncture-mediated CC10 upregulation promotes a shift in pulmonary DC composition toward a less proinflammatory state. This CC10-dependent modulation of CD11b+ DCs provides a cellular framework through which acupuncture attenuates Th2-mediated airway inflammation in asthma.
Furthermore, genomic deletion of CC10 exacerbated the hallmark pathological features of allergic asthma, including AHR, eosinophilic inflammation, mucus hypersecretion, elevated Th2 cytokines, and increased HDM-specific IgE production. Notably, CC10 deficiency significantly impaired the protective effects of acupuncture, thus supporting the conclusion that CC10 is functionally required for achieving optimal immunomodulatory outcomes. However, residual reductions in eosinophil counts and Th2 cytokine levels were still observed in CC10-deficient mice receiving acupuncture intervention, thereby suggesting the involvement of additional CC10-independent mechanisms. This observation aligns with the prevailing concept that acupuncture engages a multifaceted regulatory network, potentially involving neural signaling and broader systemic immune modulation [32].
Finally, the acupuncture-induced upregulation of CC10 expression is likely related to the preservation of airway epithelial integrity. Environmental allergens typically disrupt epithelial barrier functions, triggering the release of epithelial-derived alarmins, such as IL-25, IL-33, and thymic stromal lymphopoietin (TSLP), which orchestrate downstream type 2 inflammatory cascades [33]. Consistent with our previous findings that acupuncture suppresses alarmin expression and reduces the recruitment of type 2 innate lymphoid cells (ILC2s) [10], the present data support a model in which acupuncture contributes to the integrity of epithelial structure and promotes club cell recovery. This, in turn, augments CC10 production and restores epithelial-immune homeostasis within the allergic airway environment.
Our study identifies CC10 as a key molecular mediator essential for the therapeutic efficacy of acupuncture in allergic asthma. Acupuncture augments CC10 expression and modulates pulmonary DC subsets, subsequently leads to the marked suppression of Th2-driven airway inflammation and the restoration of airway immune homeostasis. These findings provide mechanistic insight into the immunoregulatory actions of acupuncture in asthma and highlight CC10 as a potential therapeutic target for the management of allergic airway diseases.
The data that support the findings of this study are available from the corresponding author upon reasonable request.
YDX and YQY contributed to the study conception and design; JL, MC, QLW, and YYZ designed and carried out the experiments; JL, MC, QLW, and YDX collected and analyzed the data; YW, YJC, and LMY contributed to the interpretation of the data; YDX wrote the manuscript. YW, YJC, and LMY critically revised the manuscript for important intellectual content. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
The study was carried out in accordance with the guidelines of the Declaration of Helsinki. The human study protocol was reviewed and approved by the Human Research Ethics Committee of Yueyang Hospital of Integrated Chinese and Western Medicine, Shanghai University of Traditional Chinese Medicine (Approval No. 2013-041). Patients with asthma were enrolled from the clinical trial NCT 01931696, and all provided written informed consent. All animal procedures were performed in accordance with the Guide for the Care and Use of Laboratory Animals and received approval from the Institutional Animal Care and Use Committee of Shanghai University of Traditional Chinese Medicine (Approval no.PZSHUTCM2309080009).
Not applicable.
This work was supported by grants from the Natural Science Foundation of China (82374583, and 81973952) and the Natural Science Foundation of Shanghai (23ZR1459900).
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBL48140.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.