








 , Honggang Liu 1,†, Xue Yan 1, Cheng Xu 1, Qin Wang 1, Yifan Xu 1, Junwei Li 1,2,3,*
, Honggang Liu 1,†, Xue Yan 1, Cheng Xu 1, Qin Wang 1, Yifan Xu 1, Junwei Li 1,2,3,*
1 Department of Infectious Disease, The Second Hospital of Nanjing, Affiliated with Nanjing University of Chinese Medicine, 210003 Nanjing, Jiangsu, China
2 Medical Innovation Center for Infectious Disease of Jiangsu Province, 210003 Nanjing, Jiangsu, China
3 College of Veterinary Medicine, Qingdao Agricultural University, 266109 Qingdao, Shandong, China
†These authors contributed equally.
Abstract
In recent years, drug-resistant influenza viruses have emerged frequently, making influenza a persistent and serious public health burden. Therefore, potential anti-influenza virus drugs are urgently needed. Nanobodies, variable domains of heavy-chain antibodies (VHHs), have the advantages of easy preparation, excellent solubility, deep tissue penetration, and weak immunogenicity; thus, they have broad application prospects in the fields of basic research and drug development. However, its short half-life and low stability limit its clinical therapeutic application. Fenobody is an engineered display platform with the ability to present multimerized nanobodies on the surface of ferritin to overcome these disadvantages and increase its potency.
In this study, we engineered a fenobody displaying multimerized VHH against haemagglutinin (HA) of influenza virus (A/California/07/2009(H1N1), pdm09) on the surface of ferritin by using the property of SpyTag to spontaneously bind to SpyCatcher, named ferritin-NP-VHH.
Compared with VHH alone, ferritin-NP-VHH improved the cross-neutralizing activity, stability and affinity for influenza virus in vitro and prolonged its half-life in vivo.
These results suggest that the implementation of genetic engineering technology to construct multimerized anti-influenza virus nanoparticles provides new tools to control infection with influenza virus.
Keywords
- fenobody
- nanobody
- influenza virus
- stability
- affinity
- protection
Influenza is an important human respiratory infectious disease caused by seasonal or emerging influenza viruses. Statistically, infection by seasonal influenza viruses kills 250,000–500,000 people worldwide each year [1, 2, 3]. In recent years, drug-resistant strains and new subtypes of influenza viruses that infect humans have emerged frequently, making infection with influenza virus a persistent and serious public health problem [4, 5, 6, 7]. Although vaccination is the primary method for preventing infection by the influenza virus, its implementation poses some technical challenges. These include (i) difficulty in predicting which strains will emerge and infect humans in the coming season, (ii) the lag period between the emergence of new strains and the availability of clinically approved vaccines, and (iii) poor immune response in aged and immunocompromised people [8, 9]. Therefore, given the increase in drug resistance, reduced vaccination effectiveness in some populations, and the short therapeutic window of existing antiviral drugs, a new type of anti-influenza virus is urgently needed [10, 11].
Nanobodies, also known as VHHs, are the variable domain of heavy-chain immunoglobulins (HcAbs) found in the family Camelidae. Despite their small size (12–15 kDa), VHHs are superior to conventional antibodies in terms of affinity and specificity [12, 13, 14]. Owing to their exceptional stability over a wide temperature range, resistance to the action of various detergents, and proteolytic cleavage, single-domain antibodies can be delivered into the body orally and by inhalation [15, 16, 17, 18, 19]. However, the samll size render nanobodies with short serum half-life and low affinity, that limits its therapeutic application in vivo.
Ferritin [20, 21, 22] is an important functional protein that regulates the storage and release of iron. Naturally, synthesized ferritin mostly presents a hollow spherical nanocage structure with an outer diameter of 12 nm and an inner diameter of 8 nm. Its spherical structure consists of an inner core and an outer shell. The inner core is mainly full of minerals. When ferritin is used as a nanocarrier, the target molecule can be wrapped inside the cage to be released slowly, or the target molecule can be anchored on the outer surface of the cage to stabilize the structure and efficiently deliver the target protein. Moreover, ferritin is easily produced in E. coli at high yield and resistant to high temperature and various detergents because of its high stability and exhibits excellent biocompatibility, without affecting its natural structure [23]. Therefore, self-assembled ferritin nanoparticles have recently been widely applied in drug delivery [24, 25, 26].
In the current study, we rationally designed a ferritin particle-based multimeric VHH, named fenobody. We linked VHH, an anti-haemagglutinin of an influenza virus nanobody, to the C-terminus of the ferritin monomer subunit for fusion expression, thereby anchoring the VHHs on the surface of the self-assembled ferritin nanocages. This ferritin particle-based multimeric nanobody was named ferritin-NP-VHH. Our in vitro and in vivo results suggest that compared with monovalent VHH, ferritin-NP-VHH has impressive prevention and therapeutic effects against the infection by influenza virus.
A/Puerto Rico/8/1934 (PR8, H1N1) was preserved by the Laboratory of Preventive Veterinary Medicine, Qingdao Agricultural University. CHO cells (CHO-2H6) lacking the GS gene and MDCK cells (ATCC, CRL-2936) were preserved in the Second Hospital of Nanjing affiliated the University of Chinese Medicine. CHO-2H6 cells were cultured in CHOGrow®302 medium purchased from Yuanpei (Shanghai) Biotechology Co., Ltd. MDCK cells were cultured in DMEM medium supplemented with 10% fetal bovine serum (FBS) and 10 units/mL of penicillin and 10 µg/mL of streptomycin. All cells were maintained at 37 °C and 5% CO2 atmosphere. All cell lines were validated by short tandem repeats (STR) profiling and tested negative for mycoplasma. 6 to 8 week-old female specific pathogen-free (SPF) BALB/c mice were purchased from Jinan Pengyue (Jinan) Experimental Animal Breeding Co., Ltd., and raised at the Animal Breeding Center of Qingdao Agricultural University with a germ-free environment.
The genes encoding SpyTag-ferritin and SpyCatcher-VHH were synthesized by GenScript (Nanjing) and optimized with CHO preferred codons. Ferritin is adopted from residues 5–174 of Pf ferritin. VHH (R1a-G6) was reported by Simon E. Hufton and colleagues [9]. Three G3S linkers were inserted between SpyTag and Ferritin and between SpyCatcher and VHH. His-tag was added at the carboxy terminus of SpyCatcher-VHH. The amino terminus of the SpyTag-ferritin and SpyCatcher-VHH genes followed after a murine IgG signal peptide sequence. The DNA sequence of the signal peptide is 5′-GAATTCATGGGTTGGAGTTGCATCATCCTATTTCTAGTGGCCACCGCTACCGGCGTG. Subsequently, the SpyTag–ferritin and SpyCatcher-VHH gene segments were amplified and inserted into the P3 plasmid containing the GS gene between the XbaI and Hind III sites. The successfully cloned P3 plasmid was introduced into GS gene-deficient CHO cells by electroporation. The procedure is set as follows. After being digested, the eukaryotic expression plasmids, 20 µg P3-SpyTag-Ferritin and 20 µg P3-Spy Catcher-VHH respectively, were added into the resuspended CHO-2H6 suspension cell culture and gently mixed, then the cells were incubated at room temperature for 10 minutes. Electroporator (ECM830) parameter (Holliston, MA, USA) is voltage 280 V, time 20 ms. Mixture of plasmids and CHO cells were added into a precooled 4 mm cuvette and electroporation was conducted, then the mixture was transferred into a culture flask preheated to 37 °C, with serum-free CHOGrow®302 medium containing L-alpha-aminoglutamic acid. After 24-hour static culture, the cells were transferred into a 125 mL cell culture flask, and suspended in vertical shaking incubator at 37 °C, 5% CO2, with speed of 110 rpm. During cell culture, number counting and viability test are performed every 24 hours, then viable cells were frozen for further use.
The CHO cells stably carrying P3-SpyTag-Ferritin or P3-SpyCatcher-VHH were screened by using medium without L-glutamine. After the screening was completed, the CHO cells were continuously cultured, and supernatant was collected for protein purification when the cell viability reached 70%. SpyTag-ferritin were purified using a Superose 6 Increase (GE Healthcare, Wauwatosa, WI, USA) size exclusion column according to the manufacturer’s protocol. SpyCatcher-VHH was purified using a Ni-NTA agarose column (Thermo Fisher Scientific, Shanghai, China) according to the manufacturer’s protocol.
SpyTag-ferritin and SpyCatcher-VHH were mixed at a 1:1 molar ratio and incubated overnight at 4 °C. Ferritin-NP-VHH was then purified using a Superose 6 Increase size exclusion column. Impurities were removed by using a 220 nm filter prior to purify ferritin-NP-VHH.
5 µL purified sample was dropped on a 300-mesh carbon-supported membrane copper grid and stained with 2% (w/v) uranyl acetate. Excessive uranyl acetate was removed until the copper mesh was dry. The sample was then observed under a transmission electron microscope (Hitachi, Tokyo, Japan) at a scale of 100 nm.
Ferritin-NP-VHH and monovalent VHH were tested for their avidity with H1N1 influenza virus by indirect ELISA. ELISA plates were coated with H1N1 influenza virus at 200 ng/well and incubated overnight at 4 °C. Afterwards, the plates were washed three times with 300 µL PBST and blocked in 300 µL of 2.5% nonfat milk at 37 °C for 2 hours. After the plate was washed three times with 300 µL PBST, 400 ng of ferritin-NP-VHH or monovalent VHH was added into each well, and the plate was incubated at 37 °C for 1 hour. After three-times wash with 300 µL PBST in each well, the plates were incubated at 37 °C for 1 hour and 100 µL of mouse anti-His polyclonal antibody (66005-1-lg, Proteintech, Wuhan, China) diluted 1:2000 was added into each well. After three-times wash, 100 µL of 1:5000 diluted goat anti-mouse-HRP-labelled antibody (SA00001-1, Proteintech) was added into each well, and the plate was incubated at 37 °C for 1 hour. After being washed, the plate was incubated with 100 µL of chromogenic solution per well for 15 minutes at room temperature in the dark. Finally, the reaction was terminated with 50 µL of 2 M H2SO4 per well, and the OD450nm was read in an automatic microplate reader (BioTek, Winooski, VT, USA).
For comparing the affinity of ferritin-NP-VHH or monovalent VHH with H1N1 influenza virions, plates were coated with 800 ng of ferritin-NP-VHH or monovalent VHH per well and incubated at 4 °C overnight. After being washed with 300 µL PBST, plates were blocked with 300 µL/well of 2.5% non-fat milk at 37 °C for 2 hours, the plates were washed three times with 300 µL PBST again. Then, 100 µL of H1N1 influenza virions diluted in gradient dilutions was added into each well, and the plates were incubated at 37 °C for 2 hours. After the plate was washed three times, 100 µL of 1:2000 diluted H1N1 HA mAb (11085-MM01, SinoBiological, Beijing, China) was added into each well, and the plates were incubated at 37 °C for 1 hour. After the plate was washed three times, 100 µL of 1:5000 diluted goat anti-mouse-HRP was added into each well, and the plate was incubated at 37 °C for 1 hour. After the plate was washed three times with 100 µL of chromogenic solution per well, the plates were incubated at room temperature for 15 minutes in the dark. Finally, the reaction was stopped with 50 µL of 2 M H2SO4 in each well, and the plates were read at an OD450nm in a plate reader. The binding constant Kd of the binding reaction was accurately calculated by using GraphPad Prism software (GraphPad Software, Boston, MA, USA, version 5.0).
5
Ferritin-NP-VHH and monovalent VHH were placed in dialysis bags containing carbonate buffer (CBS) and dialyzed at 4 °C for 24 hours. Then, 200 nM FITC and 50 nM Ferritin-NP-VHH or monovalent VHH were incubated overnight at 4 °C with rotation to label Ferritin-NP-VHH and VHH, then the mixture was dialyzed in CBS at 4 °C with stirring for 24 hours to remove the surplus FITC. The dialysate was transferred from the dialysis bag into an ultrafiltration tube and centrifuged at 4000 rpm at 4 °C. Subsequently, equal amounts (50 µg) of FITC-labelled ferritin-NP-VHH or monovalent VHH were injected into female BALB/c mice via the tail vein. Blood was collected from the submandibular vein at different time points, and the fluorescence value of the FITC-labelled protein in the blood was measured at an excitation wavelength of 485 nm and an emission wavelength of 535 nm on a VICTOR™ X series multi-label microplate reader (Waltham, MA, USA).
All experiments were approved by the Animal Ethics Committee of Qingdao
Agricultural University and performed according to the committee’s guidelines.
Female BALB/c mice (6–8 week-old) were randomly divided into groups of each
group with 8 mice and housed under designated pathogen-free conditions with food
and water ad libitum. Before administration and challenge, mice were
anaesthetized with isoflurane. All steps are performed in a biosafety hood
equipped with a charcoal filter, including (1) prepare mixture of 20%v/v
isoflurane in propylene glycol, (2) soak cotton pad in the mixed isoflurane, (3)
retrieve mouse nose close to the soaked cotton with isoflurane mixture. The depth
of anesthesia can be adjusted by moving the nostrils closer to or farther from
the cotton pad, and then intranasally administered 50 µg ferritin-NP-VHH or
monovalent VHH at 4 hours before and 24 hours after challenge with
2
Lung specimens were collected, fixed in 10% neutral buffered formalin, and embedded in paraffin wax. Five-micrometer-thick sections were stained with hematoxylin and eosin (H&E) and evaluated for histopathological alterations.
Lung samples of mice were collected and total RNA was extracted by using an RNeasy RNA extraction kit (Qiagen, Düsseldorf, Germany) according to the manufacturer’s protocol. The concentration of viral RNAs (vRNAs) was quantified by real-time RT-PCR with primers specific for the NP gene which was conducted in two steps by using a SuperScript III Platinum two-step qRT-PCR kit with SYBR green (Invitrogen, Carlsbad, CA, USA) as per the manufacturer’s instructions in an iCycler iQ-Multicolor real-time PCR detection system (Bio-Rad, Hercules, CA, USA). Specifically, vRNA was reverse-transcribed by using the Uni-12 primer and amplified by PCR using F5′-GATTGGTGGAATTGGACGAT and B5′-AGAGCACCATTCTCTCTATT as primers. The concentrations of vRNA were obtained by comparison with serially diluted plasmid carrying NP gene segment. All RNA determinations were assayed in duplicate and repeated three times.
Comparisons between different groups were performed by using nonparametric
one-way ANOVA with the Tukey multiple comparison test and Fisher’s exact test.
The survival data were analysed by using log-rank test. All analyses were
performed by using GraphPad Prism version 5.0 for Windows (GraphPad Software).
p values
In this study, ferritin-NP-VHH was generated using a modified SpyTag/SpyCatcher technique in which VHH was linked at the C-terminal of SpyCatcher convalently, as shown in Fig. 1 [27, 28, 29, 30]. Both SpyTag-ferritin and SpyCatcher-VHH were expressed in glutamine synthetase (GS)-deficient CHO cells [31, 32, 33, 34, 35]. The self-assembled nanoparticles were confirmed by transmission electron microscopy (TEM), and the particle size of Ferritin–NP-VHH was slightly larger than that of SpyTag-ferritin, as shown in Fig. 2.
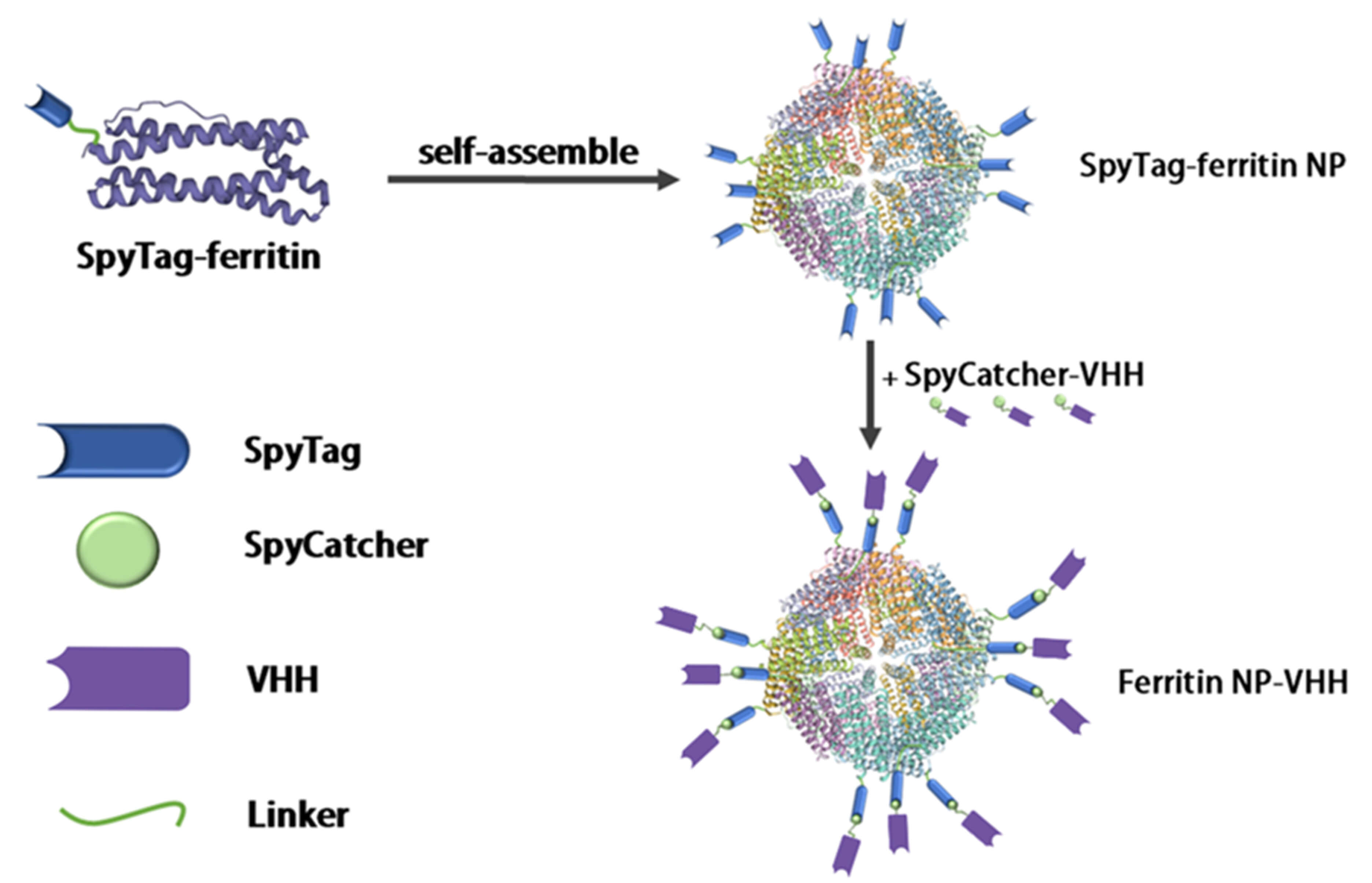 Fig. 1.
Fig. 1.
Design of the multimerization of nanobodies. Using modified SpyTag/SpyCatcher technology, the VHH was exposed to the outer surface of the ferritin shell. Each ferritin is composed of 24 ferritin monomers, and 24 VHHs can be displayed on the surface of each ferritin particle to form anti-influenza virus nanoparticles, thereby realizing the multimerization of nanobodies. VHH, variable domains of heavy-chain antibody.
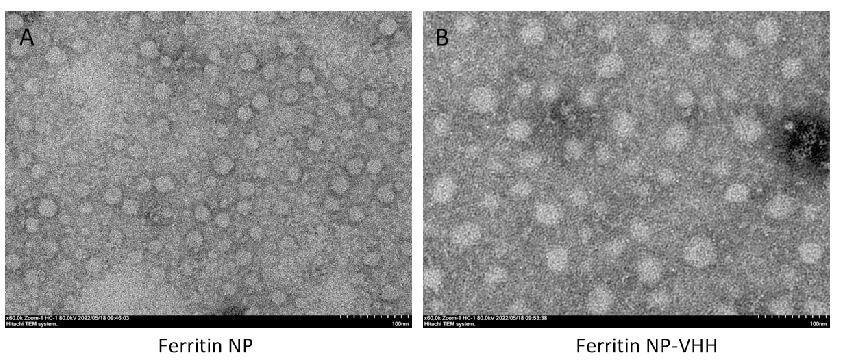 Fig. 2.
Fig. 2.
Images of ferritin particle (A) and ferritin-NP-VHH (B) were taken by transmission electron microscopy. Under the same magnification conditions, the particle size of ferritin-NP-VHH is significantly larger than that of ferritin particle. NP, nanoparticle. Scale bar = 100 nm.
To evaluate the specific binding activity of ferritin-NP-VHH, monovalent VHH with influenza virus, ELISA plates were coated with H1N1 influenza virus overnight, after which equivalent amounts (0.03 µM each) of ferritin-NP-VHH or monovalent VHH were added to each well. The plates were read in a plate reader at an absorbance of 450 nm. Finally, the data were analysed by using GraphPad Prism software (version 5.0), and the binding constant Kd was calculated. The results showed that both ferritin-NP-VHH and monovalent VHH are efficiently bound to the influenza virus. However, compared with monovalent VHH, ferritin-NP-VHH was more sensitive to influenza virus capture by 7.18-fold, as shown in Fig. 3.
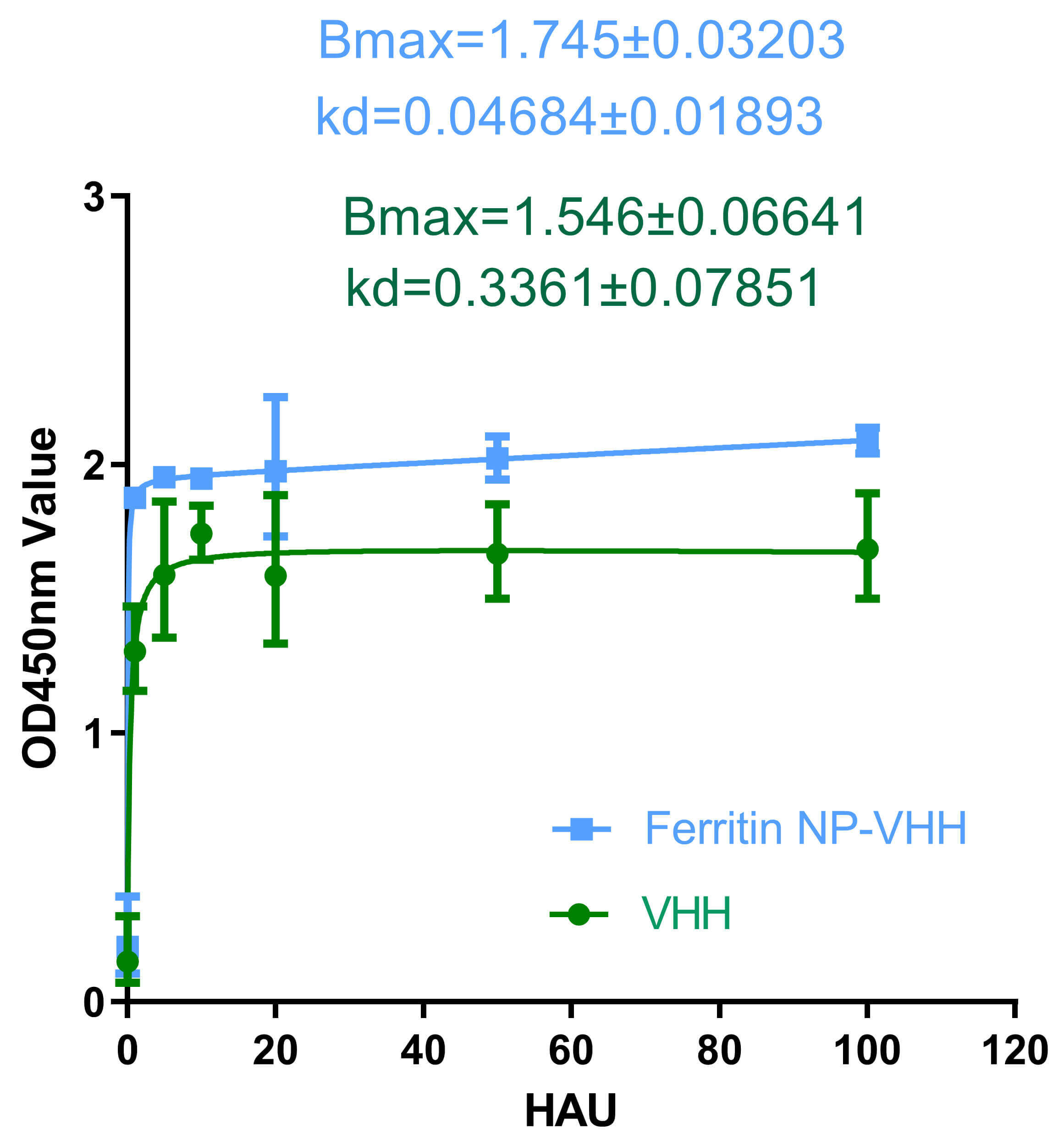 Fig. 3.
Fig. 3.
Binding of ferritin-NP-VHH or monomer VHH with
influenza virus. Both ferritin-NP-VHH and VHH can specifically bind to influenza
virus, but the capture affinity of ferritin-NP-VHH (Kd = 0.4684
Next, we determined the specific neutralizing activity of ferritin-NP-VHH or monovalent VHH with influenza virus by using the standard plaque assay method. The neutralization and plaque size results show that ferritin-NP-VHH conferred higher neutralizing activity than monovalent VHH against the influenza virus, as shown in Fig. 4A,B.
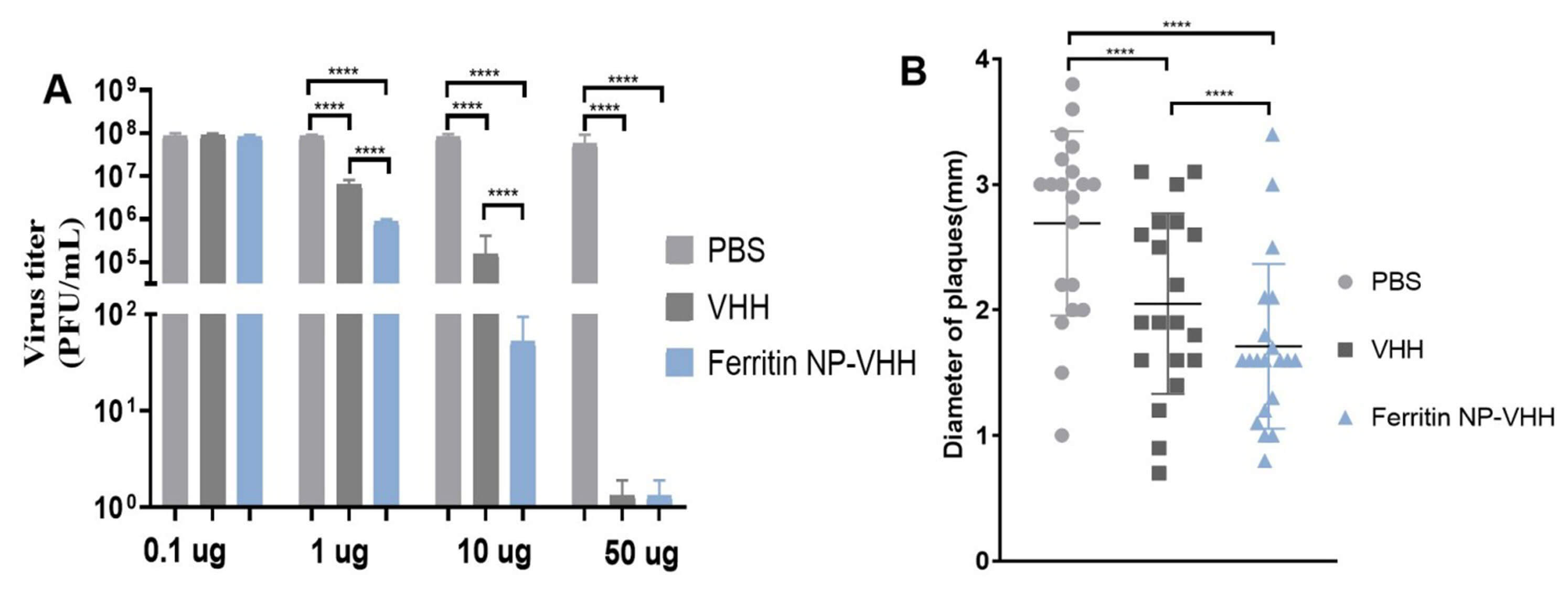 Fig. 4.
Fig. 4.
Neutralization of ferritin-NP-VHH or monomer VHH with
influenza virus. Both ferritin-NP-VHH and VHH showed potent neutralizing
activity against influenza virus. However, neutralization (A) and plaque size (B)
suggested ferritin-NP-VHH conferred higher neutralizing activity against
influenza virus than monomeric VHH (SD was specified as error bars and
****p value
To compare the half-life of ferritin-NP-VHH with that of monovalent VHH in vivo, the fluorescence values in the blood of BABL/c mice injected with ferritin-NP-VHH-FITC or monovalent VHH-FITC by tail vein were measured. The results show that the half-life of ferritin-NP-VHH in the blood is 2.44 times longer than that of monovalent VHH, as shown in Fig. 5.
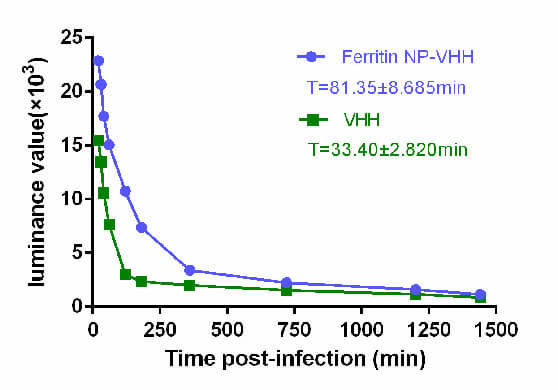 Fig. 5.
Fig. 5.
Ferritin-NP-VHH had a longer half-life than mononer VHH did in mice. To compare the half-lives of ferritin-NP-VHH and VHH, the fluorescence values in the blood of BABL/c mice injected with ferritin-NP-VHH/VHH-FITC were measured. The results show that the half-life of ferritin-NP-VHH in the blood is approximately 2.44 times longer than that of VHH. FITC, fluorescin isothiocyanate.
We subsequently investigated the protective potential of ferritin-NP-VHH against
lethal infection with influenza A virus in vivo. Eight-week-old female
BALB/c mice were inhaled intranasally with 50 µg ferritin-NP-VHH or 50
µg monovalent VHH at 4 hours before and 24 hours after challenge with the
influenza A/Puerto Rico/8/1934 (H1N1) virus at a dose of 2
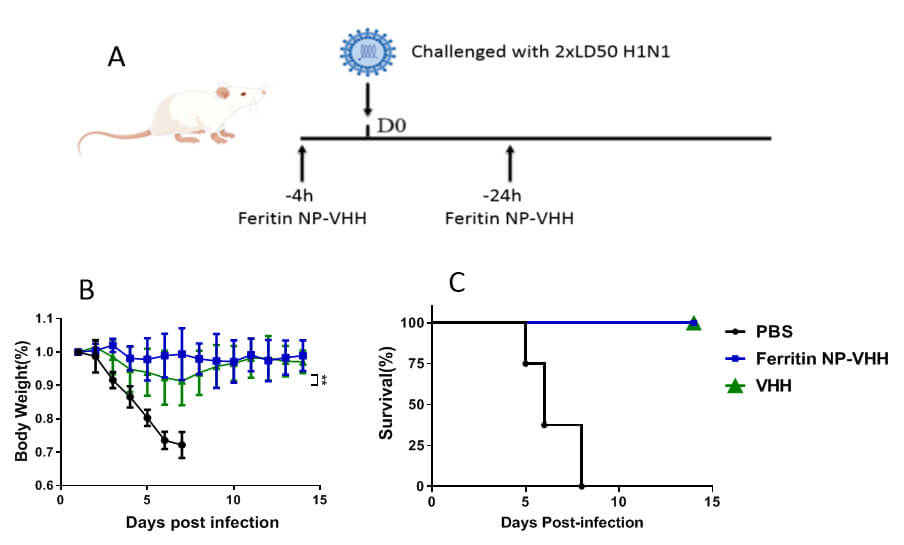 Fig. 6.
Fig. 6.
Ferritin-NP-VHH and monomeric VHH provided protection
for mice against challenge with lethal influenza virus. Compared with PBS
control mice, mice treated with ferritin-NP-VHH were significantly better
protected, as indicated by weight loss and lethality caused by influenza virus
infection. Furthermore, compared with mice in the VHH group, mice in the
ferritin-NP-VHH group had significantly less body weights after the challenge (n
= 8 SD was specified as error bars, **p
Additionally, pathological changes were detected in the lungs of mice challenged with the influenza A/Puerto Rico/8/1934 (H1N1) virus. Significant histological changes, such as diffuse inflammation of the interalveolar septa accompanied by intra-alveolar oedema, were observed in the lung sections of mice in the PBS group; otherwise, few pathological changes and less viral replication were observed in the lung tissues of mice in the ferritin-NP-VHH group (Fig. 7A–C). The NP gene segment was presented in a very low level (Fig. 7D). These findings indicated that ferritin-NP-VHH protected mice from infection by the influenza A/Puerto Rico/8/1934 (H1N1) virus.
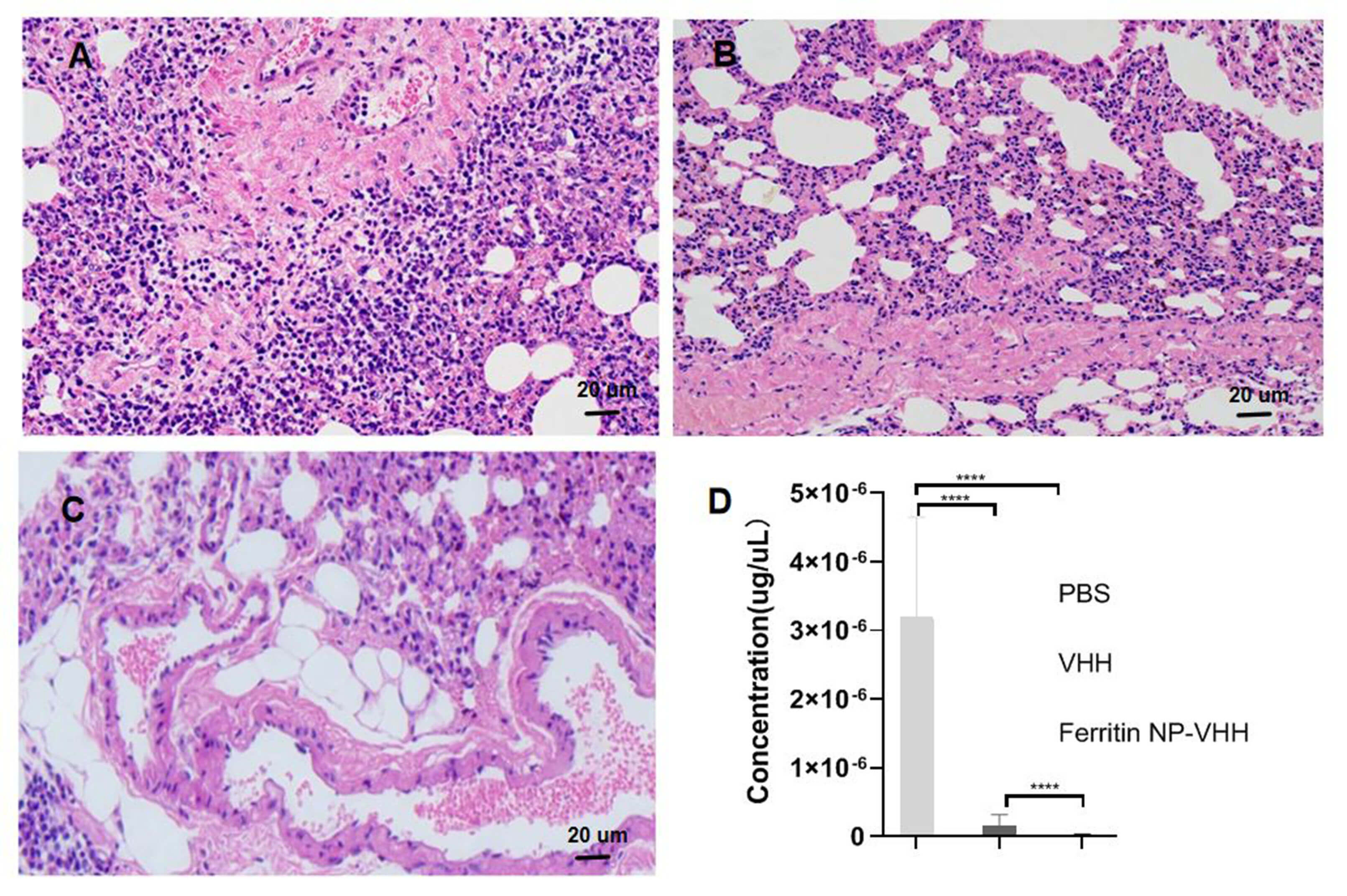 Fig. 7.
Fig. 7.
Pathological alterations in the lungs of mice.
Pathological alterations in each tissue section were examined via microscopy at
an original magnification of 40
Engineered VHH fragments have enhanced specificity, activity, half-life, and cross-protection for the prevention or treatment of infectious diseases [36, 37]. Here, we displayed VHH fragments against the HA of the influenza virus (A/California/07/2009(H1N1) pdm09) on the surface of ferritin particles to obtain multimeric nanoparticles and tested whether compared with VHH monomers, VHH improved the activity of anti-influenza virus. To explore this possibility, both SpyTag-ferritin and SpyCatcher-VHH were expressed in GS-deficient CHO cells by using modified SpyTag/SpyCatcher technology. The two proteins were then linked by using the automatic linking property of SpyTag and SpyCatcher to obtain multimeric nanoparticles on the basis of the ferritin platform.
The assembled ferritin-NP-VHH particles and SpyTag-ferritin particles were observed under the same conditions by TEM. The particles of ferritin-NP-VHH were significantly larger than those of SpyTag-ferritin particles were, indicating that SpyTag-ferritin and monovalent VHH were assembled successfully to fix multimerized nanoparticles. We first tested the half-life of ferritin-NP-VHH and monovalent VHH in mice. The half-life of ferritin-NP-VHH was longer than that of monovalent VHH. We evaluated the specific binding activity and sensitivity of ferritin-NP-VHH and monovalent VHH to A/Puerto Rico/8/1934 (H1N1) influenza virus. We found that the specific binding activity and sensitivity of ferritin-NP-VHH to influenza virus were significantly greater than those of monovalent VHH. Similar results have been presented by other researchers. Wang et al. [38] reported the construction of a Salmonella-specific fenobody through the fusion of a nanobody to ferritin, resulting in a self-assembled 24-valent nanocage-like structure. Compared with the conventional nanobody, their fenobody exhibited a 35-fold increase in avidity while retaining good thermostability and specificity. Next, we verified the neutralizing activity of ferritin-NP-VHH and monovalent VHH with influenza, and both had neutralizing activity with A/Puerto Rico/8/1934 (H1N1) influenza virus. Fan and colleagues [39] also reported that the half-life of fenobody was 10-fold longer than that of its monomer counterpart in a murine model and that antibody binding increased approximately 360-fold when H5N1 fenobody was used. Fenobodies can also be self-assembled in silkworm baculovirus expression systems, and ferritin self-assembly technology can effectively increase the neutralization activity of nanobodies against SARS-CoV-2 [40, 41]. Wang and colleagues [38] reported that fenobody increased the specific neutralizing activity against hepatitis E virus capsid and provided broad protection.
Finally, we compared ferritin-NP-VHH and monovalent VHH in protecting mice against potentially lethal influenza A virus challenge. Mice treated with ferritin-NP-VHH particles conferred significant protection against lethal infection by A/Puerto Rico/8/1934 (H1N1) influenza virus, as indicated by weight loss and survival rate. These findings suggested that the preventive and therapeutic function of ferritin-NP-VHH particles was significantly better than that of monovalent VHH.
In this study, our results showed that ferritin-NP-VHH provided superior neutralization and protection against A/Puerto Rico/8/1934 (H1N1) influenza virus although the original VHH (R1a-G6) was selected against influenza virus A/California/7/09 (H1N1) (CA09) [9]. Theoretically, VHH (R1a-G6) functionally blocks attachment of HA to receptors on cell surface which was mediated by the globular head domain. Here, these data suggested that multimerization of VHH improved the binding affinity and breath of the nanobodies and prolonged the blood half-life in vivo in line with published research [39]. It is better structurally identified the epitope recognized by VHH to lay the basis for heterologous protection between VHH and different influeza virus strains. The multimerization of a nanobody, fenobody, is not only superior to that of a monovalent nanobody but also enhances sensitivity and specificity and elongates the half-life in vivo. This study may lay a foundation for the establishment of a broad range of neutralizing drugs for the prevention and treatment of influenza viruses. In this study, the limitation is only A/Puerto Rico/8/1934 (H1N1) influenza virus was employed to test the neutralization and protection of ferritin-NP-VHH and monovalent VHH. In the future work, an array of influenza viruses should be used to tested for cross-protection conferred by ferritin-NP-VHH, such as H3N2, H5N1 and H7N9 influenza viruses. In this study, ferritin cage (SpyTag-ferritin) control group is not included in the animal experiments. It is possible that the SpyTag/SpyCatcher components or ferritin cage could induce a non-specific innate immune response that contributes to virus and nanoparticle clearance. It is inevitable to test the innate immunty stimulation by SpyTag/SpyCatcher components or ferritin cage that could limit the repeated administration with SpyTag-ferritin on behalf of the clinical application of fenobody.
Despite these advantages, ferritin is highly conserved from prokaryote to mammal species and has immunogenicity [42]. Challenges still lie in the clinical application of fenobodies, such as possible anti-ferritin antibodies due to repeated administration, safety concern, comparably high production costs, and poor payload delivery performance. However, with the obvious advantages of high stability and biocompatibility, fenobodies are still promising platforms for the development of nanotecnology-based drugs.
The ferritin-NP-VHH based on ferritin nanoparticles not only displays the advantages of traditional VHH but also has the advantages of high sensitivity and long half-life. Thus, it is of great significance and potential to the prevention and treatment of infection by influenza A (H1N1) viruses and other subtypes.
The data underlying this article are available in the article and are available upon request from corresponding author.
QWeng and JL designed this research study. QWeng, HL, XY, and CX performed the research and made figures. QWang and YX collected the samples and search references. JL wrote and revised the manuscript. All authors read and approved the final manuscript. All authors contributed to editorial changes in the manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
In this study, the animal experimental protocol was approved by the Institutional Animal Care and Use Committee at Qingdao Agricultural University (approval reference number # 20200018), and animal experiments were conducted at Qingdao Agricultural University. All animal experimental tests were carried out in accordance with the 2016 laboratory animal standards in China and other related regulations in the Animal Welfare Act.
We appreciate all the funders for supporting this study. As well, we appreciate Dr. Aidong Gu’s advice and suggestion on this study.
This study was supported by Nanjing Important Science & Technology Specific Projects (2021-11005) and the Innovation Center for Infectious Disease of Jiangsu Province (NO. CXZX202232).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.