



 , Giuseppe Pantò 1, Antonio Catalfamo 2, Alessia Filippone 2,3,*
, Giuseppe Pantò 1, Antonio Catalfamo 2, Alessia Filippone 2,3,* , Emanuela Esposito 2
, Emanuela Esposito 21 Department of Biochemical, Dental, Morphological and Functional Imaging, University of Messina, 98125 Messina, Italy
2 Department of Chemical, Biological, Pharmaceutical and Environmental Sciences, University of Messina, 31-98166 Messina, Italy
3 Now with Department of Medical Surgery, UniCamillus-Saint Camillus International University of Health Sciences, 00131 Rome, Italy
Abstract
Hepatocellular carcinoma (HCC) is a leading cause of cancer-related mortality worldwide, owing to its molecular complexity and limited therapeutic options. Three-dimensional (3D) in vitro models more accurately recapitulate in vivo conditions than traditional two-dimensional (2D) models, due to their ability to accurately reproduce the tumor microenvironment (TME). Among these models, HepG2 cell-derived spheroids have become an important tool for drug screening, toxicity assessment, and liver cancer research. This review highlights the advantages and limitations of currently available 3D culture systems. In particular, special attention is given to the multifaceted role of leucine-rich repeat kinase 2 (LRRK2), a gene traditionally associated with neurological disorders but increasingly implicated in cancer, a kinase, which emerges as a promising therapeutic target in HCC since it regulates oxidative stress, lipid metabolism, and treatment responses, all of which contribute to tumor progression. Finally, we explore future directions, including organ-on-chip technologies and co-culture systems, which hold considerable promise for improving precision medicine and translational research in HCC.
Keywords
- liver cancer
- hepatocellular carcinoma
- tumor microenvironment
- three-dimensional cell culture
- spheroids
- HepG2 cells
- leucine-rich repeat kinase 2
The third most common cause of cancer-related deaths worldwide and the sixth most prevalent malignancy overall is liver cancer, representing 7.8% of deaths and according to Cancer statistics (2024), it increases by about 2% per year [1]. Approximately 90% of primary liver malignancies are hepatocellular carcinomas (HCCs), representing a serious global health problem. The combination of chronic liver condition, the variety of diagnostic and treatment options, and the relative lack of solid scientific support for some standard procedures makes managing patients with HCC challenging. Clinical, radiological, and histological criteria are used to diagnose HCC. The gold standard for determining tumor grade, stage, and molecular features as well as validating the diagnosis of HCC is still histopathological analysis of liver tissue taken by biopsy [2]. Serum indicators include des-gamma-carboxy prothrombin (DCP), alpha-fetoprotein (AFP), and AFP-L3 are frequently used for HCC monitoring and diagnosis, despite their low sensitivity and specificity. Additionally, the lack of reliable biomarkers has prevented population-based treatments for HCC from producing the desired therapeutic outcomes [3, 4]. In recent years, in vitro models have been crucial to cancer research due to their ability to mimic the pathophysiological environment, drug toxicity evaluation, and the investigation of the molecular pathways underlying tumor formation [5]. Often employed as a model for genotoxic evaluation of chemicals [6], the highly differentiated liver features of the Hepatocellular carcinoma G2 (HepG2) cell line support it as one of the most used models for studying HCC. Moreover, the leucine-rich repeat kinase 2 (LRRK2) gene, which was first identified as being involved in neurological illnesses, is associated with numerous types of cancer. It has been also demonstrated to be relevant in oncology, particularly in HCC, because of its impact on lipid metabolism and oxidative stress. Advanced in vitro models, such as spheroids, microtissues, organoids, and organ-on-a-chip technologies are increasingly being used to study therapeutic effectiveness, safety, toxicity, and disease pathology [7]. Typically, spheroids develop a necrotic and hypoxic core, surrounded by an outer layer of actively proliferating cells (Fig. 1) [8]. This review would examine the different methods to generate HepG2 spheroids, especially in three-dimensional models and the multifaceted function of different kinases, such as LRRK2, in liver carcinogenesis and treatment response.
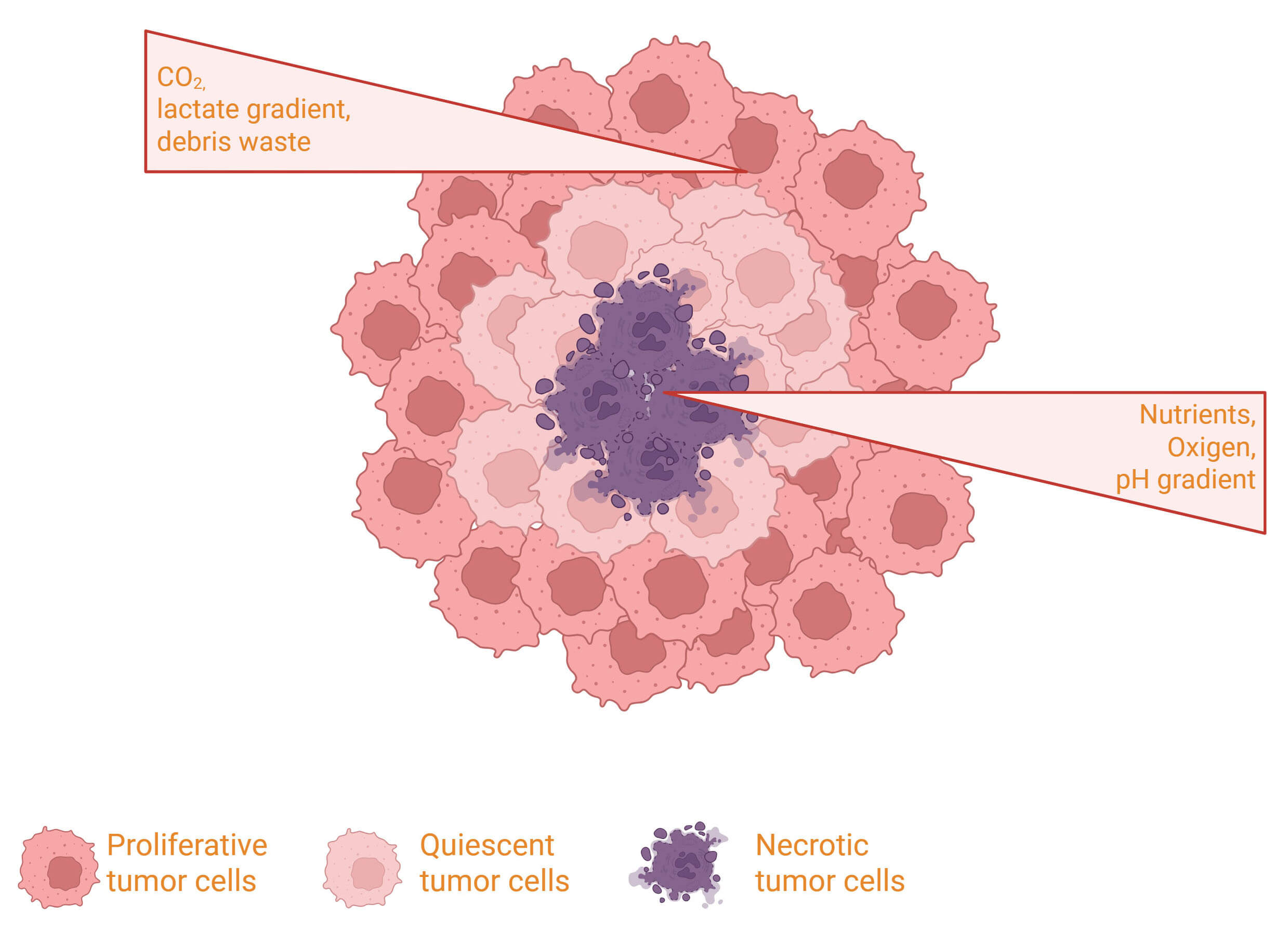 Fig. 1.
Fig. 1.
Schematic illustration of a tumor spheroid structure, highlighting the proliferating outer cell layer, the quiescent intermediate zone, and the necrotic core resulting from nutrient and oxygen gradients. Created in BioRender.com. Palermo, N. (2026) https://BioRender.com/dsxp9fj.
In vitro models are crucial resources for researching medication reactions, disease processes, and cell biology. Traditionally, cells are grown on flat, hard plastic surfaces in 2D monolayer systems in which cells develop as a monolayer adhered to a plastic surface in a flat petri dish or culture flask [9]. The intricacy of in vivo tumor microenvironment (TME) cannot be replicated by conventional 2D cell cultures, despite their widespread use, due to their ease of use and repeatability. On the other hand, 3D culture models, such as spheroids, enable cells to create more realistic nutrition and oxygen gradients as well as cell-matrix and cell-cell interactions, better simulating physiological settings. These characteristics have a big impact on cellular activity, including drug responsiveness, differentiation, and proliferation. In contrast to 2D monolayers, tumor spheroids frequently show greater resistance to chemotherapeutic treatments, which are consistent with observations made in clinical settings. Consequently, 3D models are becoming more widely acknowledged as more physiologically appropriate and predictive platforms for research on cancer biology and medication screening [10]. In 2D models, the absence of spatial and biochemical signals present in in vivo tissues, results in changes to cellular activity, gene expression, and polarity despite their affordability and convenience. While 3D culture allows cells to interact in all directions, generate extracellular matrix connections, and develop gradients of oxygen, nutrients, and metabolites, all of which more closely resemble the architecture of natural tissue. Particularly in cancer research, these circumstances result in more physiologically accurate models that accurately forecast in vivo reactions. Therefore, even though 3D systems are more complicated and technically difficult to use; they have become more popular for applications like drug screening, toxicity testing, and tumor modeling [11, 12]. Although the methods used may vary from laboratory to laboratory, we have summarized in Table 1 the common standard procedures for 3D culture models.
| Step | Aim | Activity | Critical parameters |
| Cell preparation and cell expansion | Ensure healthy cells are ready for three-dimensional (3D) culture | Two-dimensional (2D) expansion, viability check ( |
Cell type, initial density, cell viability |
| Cell seeding | Seed cells uniformly in the 3D system | Calculate seeding density, seed cells and incubate for cell growth | Attachment time, homogeneity, seeding density |
| 3D cell culture maintenance | Support growth and preservation of the 3D model | Regular media changes, viability monitoring | Culture time, incubator conditions, number of media changes |
| Treatment/Pharmacodynamic Experiment | Evaluate drug response in 3D cell culture | Define concentrations and time treatment, ensure drug penetration | Concentration, exposure time, drug dose |
| Endpoint and analysis | Drug effects evaluation | Measure cell viability, genes and protein expression | Data standardization, replicates and analysis methodology |
3D spheroids are small, spherical clusters of cells that can spontaneously self-assemble in non-adherent settings, closely resembling interactions between cells and biological processes that occur in an in vivo setting [13]. Spheroids can be generated using various techniques, each with specific advantages and limitations (Fig. 2). One of the most common approaches is spontaneous cell aggregation in non-adherent or ultra-low attachment (ULA) round-bottom plates, where cells naturally cluster into three-dimensional structures [11]. Another widely used method is the hanging drop technique, which involves placing small droplets containing cells on the underside of a culture plate lid, promoting aggregation through gravity [14]. More advanced strategies include the use of three-dimensional scaffolds, such as hydrogels or synthetic matrices, which provide structural support and facilitate tissue-like organization [15]. Additionally, dynamic systems such as rotating wall vessel bioreactors or spinner flasks enable spheroid formation under controlled flow conditions, improving size uniformity and reproducibility [16]. Generally, the ideal size range is between 0.5 mm3 and 1 mm3. But it must be considered that a 200 µm tight spheroid might already be hypoxic, whereas a 400 or 500 µm loosely spheroid might not be [17]. Moreover, it is important to consider that the stage of the spheroid affects both diffusion and toxicity, altering the response to the medication [18].
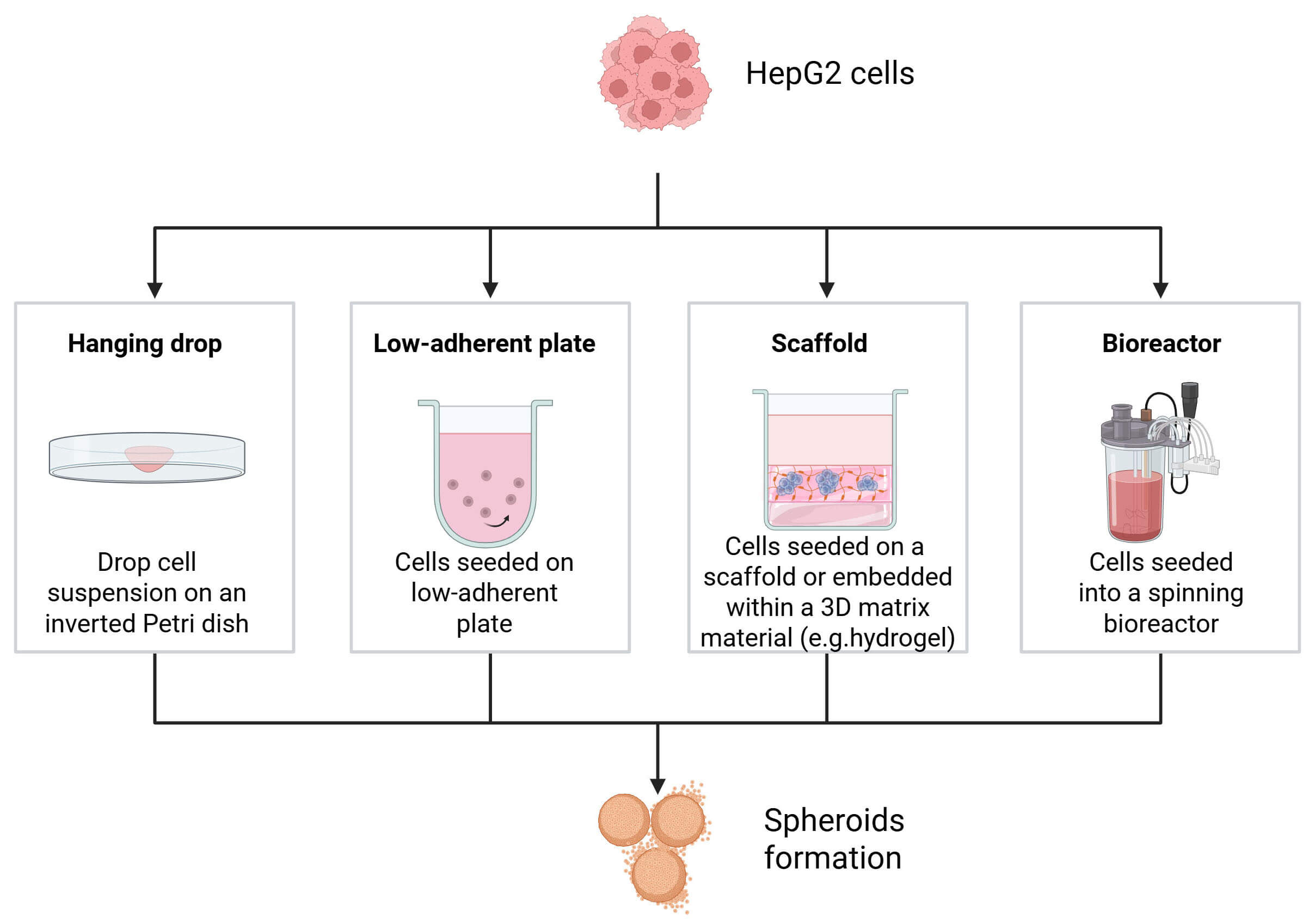 Fig. 2.
Fig. 2.
Overview of the main methods for generating HCC spheroids, including hanging drop, ultra-low attachment (ULA) plates, and hydrogel/Matrigel embedding, and bioreactors. Created in BioRender.com. Palermo, N. (2026) https://BioRender.com/t2v7ryk.
The choice of methods depends on factors such as cell type, desired application, and the need for control over spheroid size, shape, and microenvironment. 3D cell cultures have been developed and applied to preclinical screening as they can mimic the physiological and diseased states in vivo, while eliminating the discrepancies between animals and humans [19]. For instance, it has been observed that the hepatocyte cell lines, as HepG2 and Hepatoma-derived Progenitor cell line (HepaRG) can produce self-aggregating spheroids with a persistent hepatic phenotype that are appropriate for hepatotoxic and metabolic studies [20].
3D spheroids may be produced in several of ways, including ULA plates, which encourage spontaneous aggregation and inhibit cell adhesion. Spheroids cultivated with ULA plates exhibit a less distinct membrane surrounding their periphery, are less spherical, less compact, and are more irregular [21]. The oxygen, nutritional, and waste gradients that form as the spheroids enlarge closely mimic the circumstances found in avascular tumor areas. The cell type, the objectives of the experiment, and the requirement for scalability all influence the technique selection [11]. Human hepatocytes are frequently used to create bioartificial liver devices in hollow fiber formats; this configuration is likely among the best options for maintaining cultures of large numbers of hepatocytes for prolonged culture periods [22].
In hanging drops, cells develop 3D spheroids at the summit of the medium droplet. The hanging drop technique preserves a high local concentration of endogenous substances, hence sustaining tissue functions more effectively than monolayer cultures [23]. However, necrosis may result from limits in nutrition and gaseous diffusion as well as challenges with media exchange. These hypoxic conditions, in some cases, could be useful to study the growth and progression of tumors in cancer research [24]. Furthermore, modifications to the extracellular matrix, medium volume, and seeding density have led to the formation of spheroids with a variety of final properties [25]. Thus, the variable size between one spheroid and another, short-term cultures, evaporation loss of the culture medium and the limited volume (each spheroid is seeded in 20 µL/drop) of the culture are some of the disadvantages of this method [26].
Several natural and synthetic materials can be used to create 3D scaffolds.
Natural biocompatible biomaterials used as components of the extracellular matrix
(ECM) are collagen, fibrin, and hyaluronic acid. Biomaterials, ceramic-based
materials and self-assembled peptides are examples of synthetic materials
utilized in 3D scaffolds [27]. Synthetic scaffolds are more reproducible; they
can be non- biodegradable and inert, or they can be made with adjustable
degradability, which is not achievable with natural materials. Hydrogels and
solid scaffolds are the two main categories into which the many scaffold-based 3D
culturing techniques may be separated [15]. Hydrogels can be designed to either
allow cells to move from the gel surface into its core [25] or trap them in a
synthetic environment of extracellular matrix proteins to promote the growth and
function of specific cell types [28]. Biologically active substances can be added
to ECM microenvironment and cells can be encapsulated into the gels by radical
polymerizations by UV irradiation, ionic cross-linking, or self-assembly. The
capacity to encapsulate and release bioactive substances is one of the advantages
of the use of hydrogels. However, the diffusion of nutrients through hydrogel
could limit their ability to be cultivated for comparatively short periods [29].
Furthermore, the most scaffold-based technologies provide spheroid size control
through pore-size and density adjustments [29]. Oz et al. [30], studied
how the Huh7, Hep3B and HepG2 cell lines formed 3D structures, to do this they
seeded 1–2
Bioreactor systems have been designed to address some of the limitations associated with static culture conditions, aiming to improve both cell organization and tissue viability. These are typically employed to address scale-up. These systems often incorporate perfusion of the culture medium to better mimic the dynamic in vivo environment and enhance the diffusion of nutrients and gases near the cell surface. Industrial-scale bioreactors enable the cultivation of large volumes of cells and allow for precise monitoring of key in vitro parameters, such as oxygen concentration, glucose availability, and metabolite accumulation [31]. Hoyle et al. [32] developed a bioreactor system to improve the viability and function of cultured cells and tissues by introducing controlled medium perfusion. To validate the system, they used HepG2 cells to increase metabolic activity (albumin and urea secretion), and polarization under perfusion [32]. Chang and Hughes-Fulford [33] employed a rotating wall vessel bioreactor to produce spheroids up to 100 µm in diameter in 72 hours and up to 1 millimeter with long-term cultivation. Compared with animal studies, 3D cell culture systems cut the cost of drug screening since 90% of results obtained in vivo do not match the results of clinical trials, and they can drastically decrease the time of the screening process while providing input from more cell types and more physiological complexity. Most importantly, 3D cell culture as an in vitro platform allows high-throughput and high-content assays that can be integrated with imaging, automation, and computational tools to achieve next-generation drug screening for effective and specific drug discovery and development in pharmaceutical science and industry [19]. In Table 2 (Ref. [11, 15, 19, 22, 23, 24, 25, 26, 29, 31, 32, 33]) are reported the main characteristics of each method.
| Method | Principle | Advantages | Limitations | Typical morphology | References |
| Low adhesion plate (ULA) | The cells do not adhere to the surface and spontaneously aggregate in suspension. | Simple, reproducible, suitable for high-throughput assays. | Multiple spheroids or irregular aggregates are possible. | Compact and round spheroids, defined edge, internal heterogeneity with central necrosis. | [11, 15, 22] |
| Hanging Drop | Drops of cell suspension inverted; gravity promotes aggregation. | High control over initial cell number; single, uniform spheroids. | Not easily scalable; delicate handling. | Spherical, regular spheroids, more compact, radial cell distribution. | [23, 24, 25, 26] |
| Scaffold-based | The cells grow immersed in a 3D support that mimics the ECM (collagen, fibrin, hyaluronic acid). | Better reproduces the in vivo microenvironment; allows cell-matrix interactions. | Increased complexity; spheroids are difficult to recover. | Less spherical and more ‘tissue-like’ structure; polarized cells, epithelial cords similar to liver tissue. | [15, 25, 29] |
| Bioreactors | Shaking culture reduces adhesion and promotes self-aggregation. | Production of large amount of spheroids; good dynamic oxygenation. | Higher costs; mechanical stress can damage cells. | Variable, less compact spheroids, porous structure with central hypoxic zones. | [19, 31, 32, 33] |
One of the most used in vitro models in liver cancer research is the HepG2 cell line, which is derived from a human HCC. HepG2 cells are a useful tool for researching liver-specific processes like xenobiotic metabolism, drug-induced hepatotoxicity, and lipid metabolism because they maintain many differentiated hepatic functions, such as the expression of plasma proteins (e.g., albumin, transferrin), cytochrome P450 (CYP) enzymes, and different transporters. HepG2 cells offer a useful model from an oncological standpoint for examining the molecular processes of drug resistance, metabolic reprogramming, and tumor cell proliferation, especially in the setting of HCC. Though their poor tumorigenic potential and limited invasive ability must be considered, HepG2 cells are used in cancer research to examine mechanisms of tumor growth, treatment resistance, and metastatic behavior [34]. HepG2 cells are Hepatitis B Virus (HBV)-negative and genetically more stable than other HCC-derived cell lines such as Huh7 or Hep3B. This makes them appropriate for pharmacological and mechanistic research, but it also makes them less indicative of aggressive or metastatic HCC characteristics [35, 36]. Additionally, HepG2 cells that are cultivated as 3D spheroids show higher expression of drug- metabolizing enzymes and liver-specific markers, more closely resembling the in vivo hepatic milieu [37]. Thus, the HepG2 line remains a reliable and repeatable model for preclinical research focused on liver cancer biology, toxicity screening, and drug development, despite several drawbacks. Structurally the liver is highly vascularized, and its main function is to absorb and eliminate foreign substances from the body. Studies that use the intact liver have the advantage of maintaining the liver’s structural and functional organization through the maintenance of hemodynamic responses, hepatocyte polarity [38], the interaction and communication between various intrahepatic cell populations, intrahepatic micro- milieus-induced in situ signaling properties, and cellular and subcellular compartmentation [37]. Although there are now over 40 different hepatic tumor cell lines, the most often utilized ones are HepaRG, Huh7, SK-Hep-1, Hep3B, and HepG2, which are taken from different cancer types [39, 40]. The HepG2 cell line is one of them that has become well-known because of its numerous uses in scientific studies. HepG2 cells were the first hepatic cell lines to display the essential traits of hepatocytes. HCC was the name given to this line when it was identified in 1975 [39, 41]. HepG2 cells have been a part of the ATCC (American Type Culture Collection, Rockville, MD, USA) repository since 1980 as a human cell line [42]. One of the most significant differences between the HepG2 cell line and normal hepatocytes is the lack or poor expression of the cytochrome P450 (CYP) superfamily, which includes CYP3A4, CYP2C9, CYP2C19, CYP2A6, CYP2D6, and others [39]. HepG2 cells are the most similar tumor cells in hepatoblastoma based on their cytological properties, but they also have traits of normal hepatocytes. Hepatocytes are functionally polarized cells that absorb, process and release molecules and chemicals to the blood or bile with specific transporters on the basolateral or apical membrane [38, 43].
Approximately 20% of human hepatocytes are binucleated or polyploid [44, 45], and their nuclei are often anisokaryotic. In particular, HepG2 cells have between three and seven nuclei [46]. An increase in the number of nuclei is seen in tumor cells, which are also distinguished by an aberrant number of chromosomes. Smooth endoplasmic reticulum (SER) and mitochondria are abundant in hepatocytes, indicating their high levels of protein synthesis and energy consumption [47]. Compared to hepatocytes, HepG2 SER cells have half as many mitochondria and are underdeveloped. For the survival of HCC patients, new and improved diagnostic, prognostic and therapeutic approaches must be developed immediately due to the shortcomings of current methods. The identification of possible biomarkers to halt the disease’s pathophysiological processes may be a catalyst for the development of future diagnostic and more effective treatment approaches [48]. Hossen et al. [48] conducted a comprehensive review of the literature on 59 different studies, which collectively found 202 unique hub differentially expressed genes (HubGs) that contribute to HCC. They examined the most representative pharmacological targets and medicines for the therapy of HCC using an integrated bioinformatics approach [48]. An essential step is cell division, the cell cycle is regulated by Cyclin-dependent Kinase 1 (CDK1), Aurora Kinase A (AURKA), Cell Division Cycle protein 20 (CDC20), Cyclin B2 (CCNB2), polo-like kinase 1 (PLK1), and BUB1 mitotic checkpoint serine/threonine kinase B (BUB1B). One essential member of the serine/threonine protein kinase family, protein kinase CDK1, is often overexpressed in human malignancies and is essential for the progression into the mitotic phase. The disruption of this cycle is a common characteristic of cancer [49, 50].
Recently, it was discovered that 3D liver cell monoculture models fail to account for the direct or indirect signal exchange between heterotypic cells in liver tissue. Due to their numerous benefits, 3D co-culture liver models that combine 3D microenvironmental signaling and hepatic heterotypic cell interactions are particularly noteworthy. In drug-induced hepatotoxicity, hepatic drug metabolism, and drug interaction studies, several studies have shown that 3D cell co-culture liver models are more predictive than 2D and 3D monocultures and 2D co-cultures because the signaling regulation between heterotypic cells in 3D cell co-culture provides them with more relevant liver environments and phenotypes in vivo [43]. In 3D models, co-culturing HepG2 cells with non-parenchymal liver cells, like hepatic stellate cells (HSCs) or hepatic endothelial cells (HUVECs), has been demonstrated to improve cell polarization and long-term albumin expression. A 3D HepG2-NIH/3T3 co-culture system, for instance, increased albumin secretion, indicating that non-parenchymal cell contact can restore particular liver functions [51].
Prior research has demonstrated that HCC involves the dysregulation of several signaling pathways [52, 53], which have been explored as possible targets for treatment [54]. Important genes involved in cellular function are affected by the most common somatic mutations in HCC. According to Schulze et al. [55], mutations in the tumor protein p53 (TP53) and human telomerase reverse transcriptase (hTERT) promoter are found in roughly 35% and 60% of cases, respectively. The wingless-related integration site (WNT) signaling pathway includes other frequently altered genes; 30% of patients had mutations in CTNNB1, and 10% have mutations in AXIN1. Furthermore, 7% and 6% of cases, respectively, have mutations in chromatin remodeling genes such ARID1A and ARID2. Significant dysregulation of signaling pathways in HCC is supported by changes in gene expression in addition to genetic mutations [56]. Phosphatidylinositol 3-kinase (PI3K), protein kinase B (AKT), and the mammalian target of rapamycin (mTOR) are examples of oncogenic signal transduction pathways that enhance cancer cell proliferation and metastasis by promoting the Warburg effect in tumors [57, 58]. As a key metabolic organ, the liver is essential for regulating different metabolic pathways and preserving systemic metabolic homeostasis [59, 60]. The PI3K/AKT signaling cascade is thought to be a major risk factor for early recurrence and a poor prognosis in patients with HCC, and its activation has been linked to enhanced tumor aggressiveness [61]. Additionally, increased PI3K/AKT/mTOR pathway protein levels in PET/CT-positive HCC patients imply that this signaling axis may be crucial in determining the glycolytic phenotype that characterizes HCC cells [62].
Cell cycle progression, survival, metabolism, motility, and angiogenesis are
among the hallmarks of cancer that are largely regulated by the PI3K/AKT/mTOR
signaling pathway under normal settings [63]. PI3K dysregulation is one of the
most common occurrences in carcinogenesis. There are now three known classes of
PI3Ks in the human genome: Class I, Class II, and Class III [64]. Oncogenesis is
mostly linked to class I PI3Ks. Class I PI3Ks produce phosphatidylinositol
3,4,5-trisphosphate (PIP3) by phosphorylating phosphatidylinositol
4,5-bisphosphate (PIP2) when they are activated. The four catalytic subunits in
this class are encoded by PIK3CA, PIK3CB, PIK3CG, and
PIK3CD [65]. The most common mutation found in human malignancies is in
the p110
LRRK2 is a large, multi-domain protein that possesses both GTPase and kinase activities [76]. In recent years, various studies have explored LRRK2 expression across multiple tumor types, revealing its involvement in the pathogenesis of several cancers, including papillary thyroid carcinoma, colorectal, pancreatic, intrahepatic cholangiocarcinoma, as well as breast, lung and kidney cancer [77]. Inflammatory disorders, such as Crohn’s disease, have been connected to single nucleotide polymorphisms in the LRRK2 gene [78]. Interestingly, LRRK2 appears to influence tumor progression in a cancer-type-dependent manner, likely due to the tissue-specific nature of the underlying genetic networks. LRRK2 is also highly expressed in immune cells, where it contributes to cytokine production, modulating cell migration and antigen presentation.
Questions regarding the clinical use of LRRK2 inhibitors. According to data from
the Oncomine database, LRRK2 expression is reduced in cancers such as
colorectal, esophageal, ovarian, lung cancer, and leukemia, but elevated in head
and neck, kidney cancers, and lymphoma [77]. Looyenga et al. [79]
indicated that in thyroid and papillary renal carcinomas, the receptor tyrosine
kinase MET and LRRK2 cooperate to promote efficient tumor cell
proliferation and survival. Emerging evidence suggests that LRRK2 plays
a multifaceted role in the development and progression of HCC, involving
oxidative stress regulation, lipid metabolism, and therapeutic response
modulation (Fig. 3). In advanced HCC, a case-based clinical study found a
K1316N mutation in LRRK2, which was associated with increased
sensitivity to both sorafenib and radiotherapy, this effect was mediated by
elevated levels of reactive oxygen species (ROS), such as superoxide and hydrogen
peroxide, which enhanced DNA damage and cell death in tumor cells. LRRK2
mutations K1316N might indicate that HCC could respond better to
sorafenib and radiation therapy. They found that an overexpression of
LRRK2 K1316N significantly enhanced hydrogen peroxide levels and
increased sorafenib sensitivity in HCC cells [80]. Although encouraging, these
results are still preliminary and emphasize the need for additional preclinical
and clinical testing to confirm the role of LRRK2 as a predictive
biomarker. Additionally, LRRK2 silencing through siRNA further enhanced
the anti-tumor effects of sorafenib, indicating its regulatory role in therapy
response. In parallel, studies using HepG2 cells and models of Non-Alcoholic
Fatty Liver Disease (NAFLD)/Non-Alcoholic SteatoHepatitis (NASH)-related liver
damage have shown that LRRK2 promotes lipid
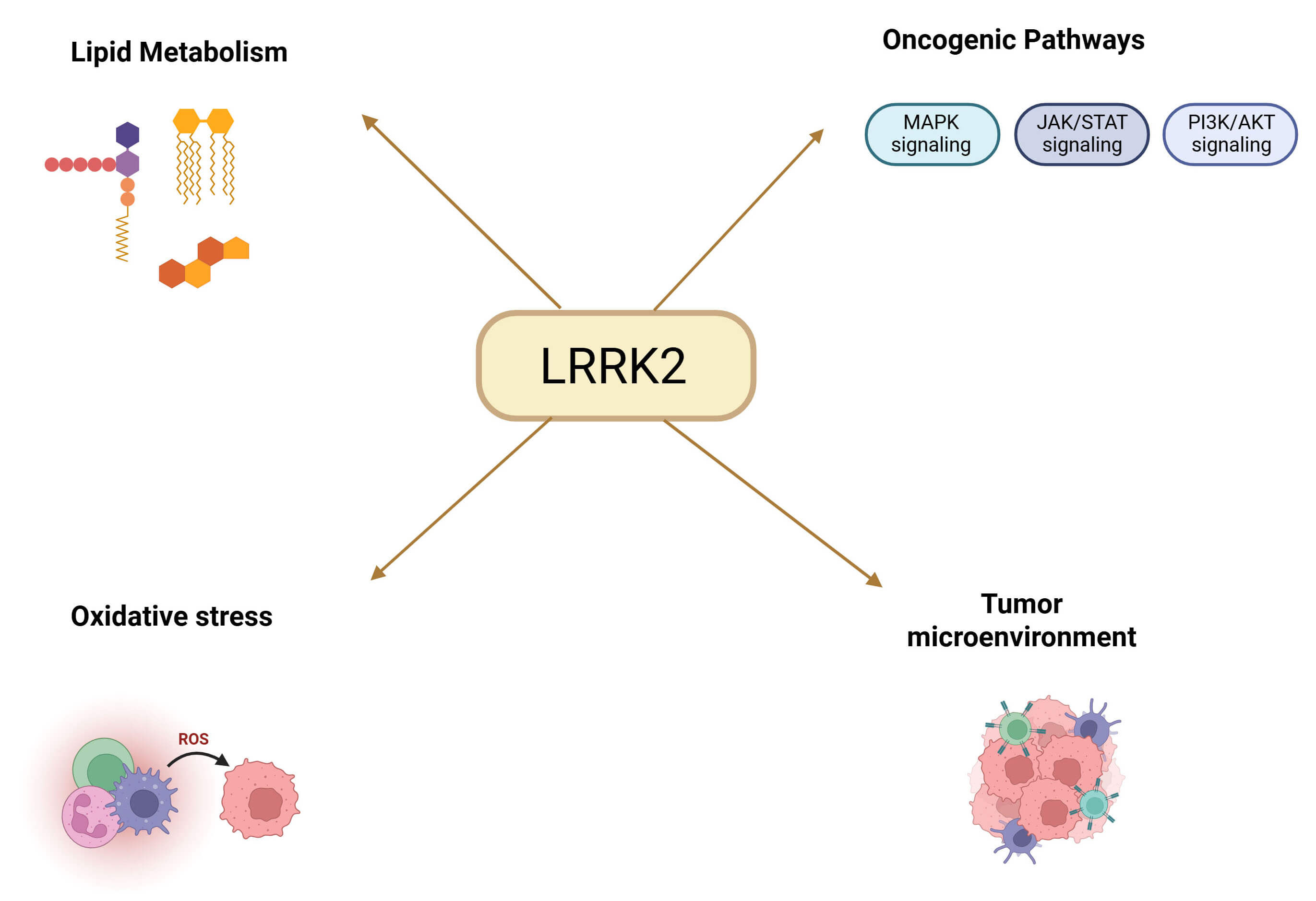 Fig. 3.
Fig. 3.
Schematic representation of the main biological processes and pathways regulated by LRRK2 in HCC, including lipid metabolism, oxidative stress, oncogenic signaling pathways (Mitogen-Activated Protein Kinase (MAPK), Janus kinase (JAK)/Signal Transducer and Activator of Transcription (STAT), phosphatidylinositol 3-kinase (PI3K)/Protein Kinase B (AKT)), and modulation of the tumor microenvironment. Created in BioRender.com. Palermo, N. (2026) https://BioRender.com/hktml7k.
3D models replicate in vivo and clinical features by representing a successful research tool, furthermore, advances in this field of technology could improve precision medicine, pre-clinical validation, and drug discovery [83]. In vivo, the 3D extracellular matrix offers structural support and also functions as a reservoir for various effector molecules, delivering essential chemical and physical signals that regulate cell behavior. To mimic migration, differentiation, survival, and development, innovative synthetic matrices are being engineered with specific features for cell adhesion, growth factor presentation, and controlled degradation [85]. To improve clinical translations and therapeutic predictability of 3D-models, recently, it has been studied the correlation between in vitro and in vivo screening intensively. Comparative studies of in vitro and in vivo models show that 3D systems more accurately simulate several aspects of solid tumors, including the release of soluble mediators, drug resistance mechanisms, gene expression patterns, and physiological responses, compared to 2D cultures. 3D systems offer a more complex microphysiological environment of human organs and allow for multiple dosing studies and drug safety evaluation [86, 87]. Nevertheless, other study demonstrated that concentration-dependent responses are often better reflected in 2D models, as 3D cultures exhibit greater variability, making it more challenging to obtain statistically meaningful results. Despite this, 3D systems may allow for better prediction of pharmacodynamic effects observed in vivo. However, further correlation studies are needed due to the variety and specificity of different organs and pathologies [88]. Literature lacks standardized protocols and operating procedures for creating 3D models, as cell seeding density, culture conditions and spheroid maturity vary from laboratory to laboratory and have a significant impact on drug susceptibility results. Indeed, scalability and standardization represent significant obstacles and could limit reproducibility [17]. Reliable preclinical models are necessary to identify novel treatment approaches for hepatocellular carcinoma. In this regard, the HepG2 cell line is used for its validity and adaptability for modeling the liver tumor environment, particularly when cultivated in three-dimensional structure. The scientific community is now working to define best practices, but the lack of common rules still makes it difficult to reproduce and translate results. Standardization of 3D culture protocols is a crucial missing step. Moreover, 3D cell co-culture liver models are expected to offer an accurate new platform since they can successfully increase the functional stability of hepatocytes in vitro and ensure the accuracy of in vitro drug prediction results. The future of translational research may integrate 3D models, co-cultures, and organ-on-chip technologies, which will make in vitro data closer to the clinical setting.
AFP, alpha-fetoprotein; AKT, Protein Kinase B; ATCC, American Type Culture Collection; AURKA, Aurora Kinase A; BUB1B, BUB1 mitotic checkpoint serine/threonine kinase B; CCNB2, Cyclin B2; CDC20, Cell Division Cycle protein 20; CDK1, Cyclin-dependent Kinase 1; CYP, cytochrome P450; DCP, des-gamma-carboxy prothrombin; ECM, Extracellular Matrix; HBV, Hepatitis B Virus; HCC, hepatocellular carcinomas; HepG2, Hepatocellular carcinoma-G2; HepaRG, Hepatoma-derived Progenitor cell line; HSC, Hepatic stellate cells; HUVECs, hepatic endothelial cells; hTERT, human telomerase reverse transcriptase; ICC, intrahepatic cholangiocarcinoma; JAK, Janus kinase; LRRK2, Leucine-Rich Repeat Kinase 2; MAPK, Mitogen-Activated Protein Kinase; mTOR, mammalian target of rapamycin; NAFLD, Non-Alcoholic Fatty Liver Disease; NASH, Non-Alcoholic SteatoHepatitis; PIP2, Phosphatidylinositol 4,5-bisphosphate; PIP3, Phosphatidylinositol 3,4,5-trisphosphate; PI3K, Phosphatidylinositol 3-kinase; PLK1, polo-like kinase 1; PTEN, phosphatase and tensin homolog; ROS, reactive oxygen species; SER, Smooth endoplasmic reticulum; SHARP, Sorafenib Hepatocellular Carcinoma Assessment Randomized Protocol; S6K1, ribosomal protein S6 kinase 1; STAT, Signal Transducer and Activator of Transcription; TME, Tumor Microenvironment; TP53, Tumor protein p53; ULA, Ultra Low Attachment; WNT, wingless-related integration site; 2D, Two-dimensional; 3D, Three-dimensional; 4E-BP1, factor 4E-binding protein 1.
AF and EE designed the research. NP developed the overall concept and wrote the draft. GP and AC integrated and refined the key highlights, created the image and searched the references. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This article was funded by PNRR - Missione 4, Componente 2, Investimento 1.5 - D.D. 3277 del 30.12.2021- Progetto SAMOTHRACE - Sicilian MicronanoTech Research And Innovation Center CUP J43C22000310006 - codice identificativo ECS00000022.
Given her role as the Guest Editor, Alessia Filippone had no involvement in the peer-review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process for this article was delegated to Qingping Dou.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.