






 , Yuanyuan Yang 1, Zhixia Zhang 1, Yiqin Ouyang 2,*
, Yuanyuan Yang 1, Zhixia Zhang 1, Yiqin Ouyang 2,* , Xiaowen Tong 1,*
, Xiaowen Tong 1,*1 Department of Obstetrics and Gynaecology, Shanghai Tongji Hospital of Tongji University, Tongji University School of Medicine, 200065 Shanghai, China
2 Department of Obstetrics and Gynaecology, Shanghai Dongfang Hospital of Tongji University, Tongji University School of Medicine, 200065 Shanghai, China
Abstract
Cervical cancer (CC) is one of the most prevalent gynecological malignancies. The expression and functional role of the long non-coding RNA (lncRNA) Ras-related protein Rab-11B antisense RNA 1 (RAB11B-AS1) in CC remain poorly understood.
The expression profile of lncRNA RAB11B-AS1 across multiple cancer types was initially assessed using data from The Cancer Genome Atlas. Its expression in CC tissues and lesions of varying pathological grades was subsequently validated via RNA in situ hybridization. To investigate its functional role in CC, a combination of transcriptomic, proteomic, and functional assays was employed to delineate the molecular role of RAB11B-AS1. The effects of alterations in RAB11B-AS1 expression on cervical cancer growth were ultimately validated in vivo.
LncRNA RAB11B-AS1 was downregulated in CC and associated with a favorable patient prognosis. Functionally, RAB11B-AS1 promoted apoptosis while suppressing proliferation, migration, and invasion of CC cells in vitro, and inhibited tumor growth in vivo. Mechanistically, RAB11B-AS1 upregulated ribosomal protein L26 (RPL26) expression. Notably, RAB11B-AS1 suppressed cervical cancer progression by activating the p53 pathway via RPL26. Critically, in vitro and in vivo experiments confirmed that RPL26 knockdown abrogates the tumor-suppressive functions of RAB11B-AS1, establishing RPL26 as a pivotal downstream effector of RAB11B-AS1 in CC.
Our findings demonstrate that lncRNA RAB11B-AS1 suppresses cervical cancer progression primarily through upregulation of RPL26 and suggest that RAB11B-AS1 may serve as a potential biomarker and therapeutic target in cervical cancer.
Keywords
- RAB11B-AS1
- cervical cancer
- cell proliferation
- metastasis
- apoptosis
- ribosomal protein L26
Cervical cancer (CC) is among the most prevalent malignant neoplasms that pose a threat to global women’s health [1]. Globally, CC is the fourth most common cancer in women, accounting for an incidence of 6.6% and a mortality rate of 7.5% [2]. Both its morbidity and mortality rates have been on the rise [3]. Despite the comprehensive and diversified diagnostic and treatment approaches for CC, the prognosis for patients with locally advanced or metastatic CC remains very poor [4]. Consequently, foundational molecular research into CC progression may lead to the identification of novel therapeutic targets.
Long non-coding RNAs (lncRNAs) have gained increasing recognition in cancer biology as pivotal regulators of gene expression. Numerous studies have established that lncRNAs play fundamental roles as oncogenes or tumor suppressors, governing key aspects of tumorigenesis such as cell proliferation, apoptosis, invasion, and metastasis, through their involvement in chromatin remodeling, transcription, and post-transcriptional modifications [5]. In cervical cancer, several lncRNAs (e.g., MALAT1, PVT1) have been mechanistically implicated in proliferation, metastasis, therapy resistance and immune modulation, underscoring the central role of lncRNAs in disease biology and their potential as diagnostic or therapeutic targets [6].
The lnc-RNA Ras-related protein Rab-11B antisense RNA 1 (RAB11B-AS1) is transcribed from the opposite strand of the RAB11B gene transcription, which contains 1034 nucleotides (three exons) and is located at chromosomal band 19p13.2 [7]. Existing studies have reported its involvement in tumorigenesis across various cancer types, yet its functions are highly context-dependent. RAB11B-AS1 is downregulated in osteosarcoma but shows elevated expression in lung and breast cancers [8, 9, 10]. Notably, in breast cancer, it enhances the expression of pro-angiogenic factors VEGFA and ANGPTL4, thereby promoting cell migration and invasion [10]. Furthermore, a recent study revealed that METTL16 diminishes RAB11B-AS1 stability via m6A modification, leading to its downregulation, which subsequently suppresses proliferation, migration, and invasion while promoting apoptosis in hepatocellular carcinoma (HCC) cells [11]. However, the functional role and underlying molecular mechanism of RAB11B-AS1 in CC have not been fully investigated. Because lncRNAs often function as upstream modulators of oncogenic signaling and exhibit high tissue specificity, dissecting the mechanistic role of RAB11B-AS1 may not only fill an important gap in cervical cancer biology but also reveal new avenues for therapeutic development.
In this study, tumorigenesis tests were conducted in vitro and in vivo to analyze the effect of RAB11B-AS1 on CC. The results revealed that RAB11B-AS1 is negatively associated with pathological grade in human CC and inhibits the growth of CC cells via the ribosomal protein L26 (RPL26)-p53 axis. These results shed light on a new prospect for CC therapy development.
The expression of lnc-RNA RAB11B-AS1 was analyzed using data from the TCGA database (https://cancergenome.nih.gov/) and data regarding RAB11B-AS1 copy number alterations were sourced from cBioPortal (https://www.cbioportal.org/). The Kaplan-Meier survival analysis was employed to assess the overall survival of patients diagnosed with cervical cancer, with a p-value of less than 0.05 being defined as statistically significant.
A total of 135 formalin-fixed, paraffin-embedded (FFPE) cervical tissue specimens were included in this study, which were collected from patients in our hospital between January 2019 and June 2024. The cohort consisted of two groups: 35 adjacent non-cancerous tissue specimens and 100 cervical carcinoma tissue specimens. The adjacent normal tissues were obtained from the non-cancerous periphery of surgical resections in patients with early-stage cervical cancer, all of which were confirmed to be histologically normal by pathological examination. The carcinoma tissues were collected from patients who underwent biopsy or surgical procedures in our Department of Gynecology, with a confirmed diagnosis of cervical cancer. All tissue specimens were processed according to standard protocols within 30 minutes after resection, fixed in 10% neutral buffered formalin, routinely embedded in paraffin, and stored in our hospital’s pathology department biobank for subsequent sectioning and analysis. All cervical cancer patients enrolled in this study were treated at the Department of Obstetrics and Gynecology, Shanghai Tongji Hospital. None of the included patients had a history of other malignant tumors, and 135 patients not undergone any radio or chemotherapy prior to this study. This study was conducted in accordance with the guiding principles of the Declaration of Helsinki. The experimental protocol was approved by the Ethics Committee of Shanghai Tongji Hospital (Approval Number: K-W-2025-011), and prior written informed consent was acquired from all enrolled patients.
The human cervical carcinoma cell line SiHa was commercially purchased from the American Type Culture Collection (ATCC) (Catalog No. HTB-35) and was cultured strictly according to the provider’s guidelines. SiHa was cultured at 37 ℃ in 90% humidity and 5% carbon dioxide (CO2) in DMEM medium (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) with 10% fetal bovine serum (FBS) (Gibco, USA) and penicillin-streptomycin (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) in a CO2 incubator. All cell lines were validated by short tandem repeat analysis and tested negative for mycoplasma.
The expression of RAB11B-AS1 was detected using RNAscope, a novel RNA in situ hybridization assay. Specific RNAscope probes targeting RAB11B-AS1 (Probe-Hs-RAB11B-AS1, Cat # 838261, Advanced Cell Diagnostics, Newark, CA, USA) were employed. RAB11B-AS1 expression levels were quantified using the semiquantitative histopathological ACD scoring system, which is outlined below: Zero = no staining or fewer than one red punctate dot per ten cells; one = one to three dots per cell; two = four to nine dots per cell with no or rare clustering; three = ten to fifteen dots per cell and/or fewer than ten per cent of dots in clusters; four = at least fifteen dots per cell and/or at least ten per cent of dots in clusters.
Total RNA extraction was carried out using Trizol reagent (Cat. No. F419KA1560, Sangon Biotech, Shanghai, China) according to the manual. The reverse transcription process was carried out utilising the PrimeScript RT Reagent Kit (Cat. No. RR047A, Takara Bio, Tokyo, Japan). Quantitative PCR (qPCR) was conducted using PrimeSTAR Max DNA Polymerase with SYBR Green (Cat. No. R045A, Takara Bio, Tokyo, Japan) on an ABI7500 instrument (This product was manufactured by Applied Biosystems, a company based in Carlsbad, CA, USA).
All primer sequences were synthesized by Shanghai Sangon Biological Co., LTD. The specific sequences are as follows:
RAB11B-AS1 forward (5′-3′): TAATCCCAGCCATTTGTG,
RAB11B-AS1 reverse (5′-3′): GAATCTCGCTCTGTTGCC.
RPL26 forward (5′-3′): GACAGAAGTACAACGTGCGA,
RPL26 reverse (5′-3′): TTTTGCGGTCTTTGTCCAGT.
The full-length cDNA of human RAB11B-AS1 (NR_038237.1) was synthesized by Viral Therapy Technologies (Wuhan, China) and was cloned into lentiviral vector pLVX-ZsGreen-Puro. rLV-CON and rLV-RAB11B-AS1 were prepared with pLVX-ZsGreen-Puro and pLVX-RAB11B-AS1-ZsGreen-Puro, respectively (Viral Therapy Technologies, Wuhan, China).
The RAB11B-AS1 sgRNA targeting RAB11B-AS1 (5′-GGCTAGGATGCGCGGCTATACGG-3′) was cloned into the lentiviral vector pLVX-U6-CMV-Cas9-P2A-ZsGreen to construct recombinant plasmid pLVX-U6-RAB11B-AS1 sgRNA-Cas9-ZsGreen (pLVX-Cas9-RAB11B-AS1). The resulting plasmid, which was used to produce the lentiviruses rLV-Cas9-RAB11B-AS1, rLV-Cas9-CON and rLV-Cas9-RAB11B-AS1, was prepared by Viral Therapy Technologies (Wuhan, China). The infection of SiHa cells with rLV-Cas9-CON and rLV-Cas9-RAB11B-AS1 was conducted in strict accordance with the manufacturer’s guidelines.
The shRPL26 (5′-GAAGTACAACGTGCGATCCAT-3′) was cloned into the pLVX-shRNA-mCherry-hygro (lentiviral vector) to construct a plasmid pLVX-shRNA-mCherry-hygro to construct a recombinant plasmid pLVX-shRNA-mCherry-hygro-hRPL26 (pLVX-RAB11B-AS1-siRPL26) which was then packaged into a lentivirus rLV-RAB11B-AS1+ rLV-shRNA-RPL26. Next, the infection of SiHa cells with rLV-CON, rLV-RAB11B-AS1, rLV-RAB11B-AS1+ rLV-shRNA-RPL26.
The SiHa cells were seeded 2
A total 600 cells were seeded in a 6-well culture dish and subsequently cultured for a period of two weeks. The colonies were fixed with methanol for a period of 15 minutes and subsequently stained with 0.5% Giemsa for a further 10 minutes. The colonies were observed and enumerated.
The SiHa cell apoptosis was detected using the AnnexinV-APC/7-AAD Apoptosis Detection Kit (Tianjin Sungene Biotech Co., Ltd., Tianjin, China). Briefly, cells were stained with 7-AAD (5 µL) and Annexin V-APC 1 µL in the dark for 15 min. Then the cells were detected using a flow cytometer (CytoFLEX, Beckman Coulter Life Sciences, Brea, CA, USA).
Cells were seeded into upper transwell chambers (Lot: 353097; Corning Inc.,
Corning, NY, USA) coated with or without Matrigel at a density of 6
1.2
SiHa cells were seeded in a 24-well plate at a density of 5
Total cellular RNA was extracted in accordance with the manufacturer’s instructions. Construction of the sequencing library and RNA-sequencing (RNA-seq) were performed by Beijing Novogene Company.
The total proteins were isolated. In brief, the samples were lysed with a 0.5 mL lysis buffer, which contains 6 M Urea, 0.1 M ammonium bicarbonate, and 0.2% SDS. Extracts were reduced with 10 mM DTT for 1 h at 56 ℃ and subsequently alkylated with sufficient iodoacetamide for 1 h at room temperature shielded from light. Then samples were mixed with 4 times the volume of precooled acetone by vortexing and incubated at –20 ℃ for at least 2 h. Subsequently, the samples were subjected to a process of centrifugation, resulting in the collection of the precipitation. Following two cycles of washing with cold acetone, dissolution of the pellet was achieved by means of dissolution buffer. This buffer contains 0.1 M triethylammonium bicarbonate (TEAB, pH 8.5) and 6 M urea. LC–MS/MS analyses were performed in Beijing Novogene Company.
A total of 3
Female BALB/c nude mice aged between four and six weeks were procured from Shanghai JieSiJie Laboratory Animal Co., Ltd. The housing of the mice was conducted in accordance with the guidelines stipulated by the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals (NIH Publication No. 8023, revised 1978). It is imperative to note that all procedures conducted in an animal experimental context were meticulously reviewed and formally endorsed by the esteemed Animal Ethics Committee of Tongji University (Approval Number: TJAB04225102).
SiHa cells in the logarithmic growth phase were digested with trypsin,
resuspended in phosphate-buffered saline (PBS), and counted to achieve a final
concentration of 5
Statistical analysis was performed using SPSS 22.0 software (IBM Corp, SPSS,
Chicago, IL, USA) and GraphPad Prism software (Version 8.0.2, GraphPad Software,
Inc., San Diego, CA, USA). The measurement data were expressed as the mean
In order to investigate the association between RAB11B-AS1 and cancers, the TCGA data was analysed initially. The study revealed a substantial decrease in RAB11B-AS1 expression levels in tumor tissues compared to normal tissues across the majority of cancer types (Fig. 1A). Concurrent analysis using cBioPortal indicated copy number loss of RAB11B-AS1 in 82% of CC cases (Fig. 1B). Consequently, we assessed RAB11B-AS1 expression in normal cervical tissues and CC tissues using RNA in situ hybridization. The results demonstrated significant downregulation of RAB11B-AS1 in cervical cancer tissues, with markedly lower RNAscope scores compared to normal tissues (Fig. 1C,D). Further analyses across different histological grades revealed a progressive decline in RAB11B-AS1 expression with increasing tumor grade, reaching its lowest level in grade 3 (G3) CC (Fig. 1E,F). In conclusion, the analysis of online survival curves demonstrated that patients exhibiting elevated levels of RAB11B-AS1 expression demonstrated a more favorable prognosis (Fig. 1G).
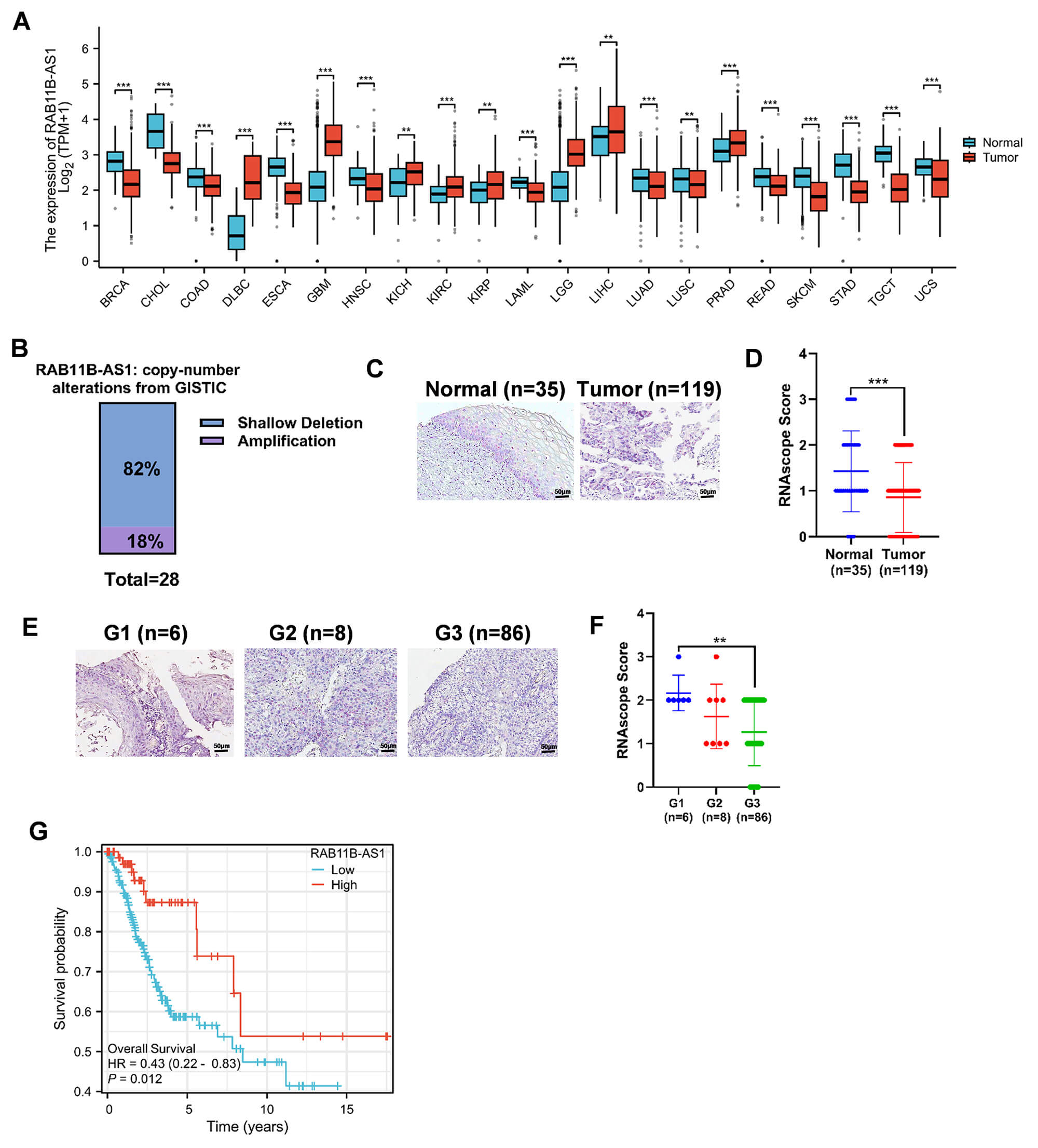 Fig. 1.
Fig. 1.
RAB11B-AS1 upregulation correlates with better CC prognosis.
(A) RAB11B-AS1 expression in normal and cancer tissues of multiple cancers via
TCGA data. (B) Copy number variation of RAB11B-AS1 in CC in the cBioPortal
database (https://www.cbioportal.org/). (C) Expression level of RAB11B-AS1 in 35
normal cervical tissues and 100 CC tissues. All micrographs (20
To explore the functional role of RAB11B-AS1 in cervical cancer, we generated SiHa cell lines with stable overexpression of RAB11B-AS1. As shown in Fig. 2A, RAB11B-AS1 expression was significantly higher in rLV-RAB11B-AS1 cells than in rLV-CON control cells. Subsequent CCK-8 assays revealed that overexpression of RAB11B-AS1 significantly inhibited the proliferation of SiHa cells (Fig. 2B). Consistent with this, colony formation assays revealed that RAB11B-AS1-overexpressing cells formed significantly fewer colonies than controls (Fig. 2C). Further analysis using Transwell assays showed that SiHa cells with RAB11B-AS1 overexpression exhibited significantly impaired migratory and invasive capacities (Fig. 2D,E). To corroborate the anti-proliferative effect, EdU incorporation assays were performed, confirming significantly attenuated proliferation in rLV-RAB11B-AS1 cells versus rLV-CON cells (Fig. 2F). Finally, flow cytometric analysis indicated that RAB11B-AS1 overexpression significantly increased the apoptosis rate in SiHa cells (Fig. 2G).
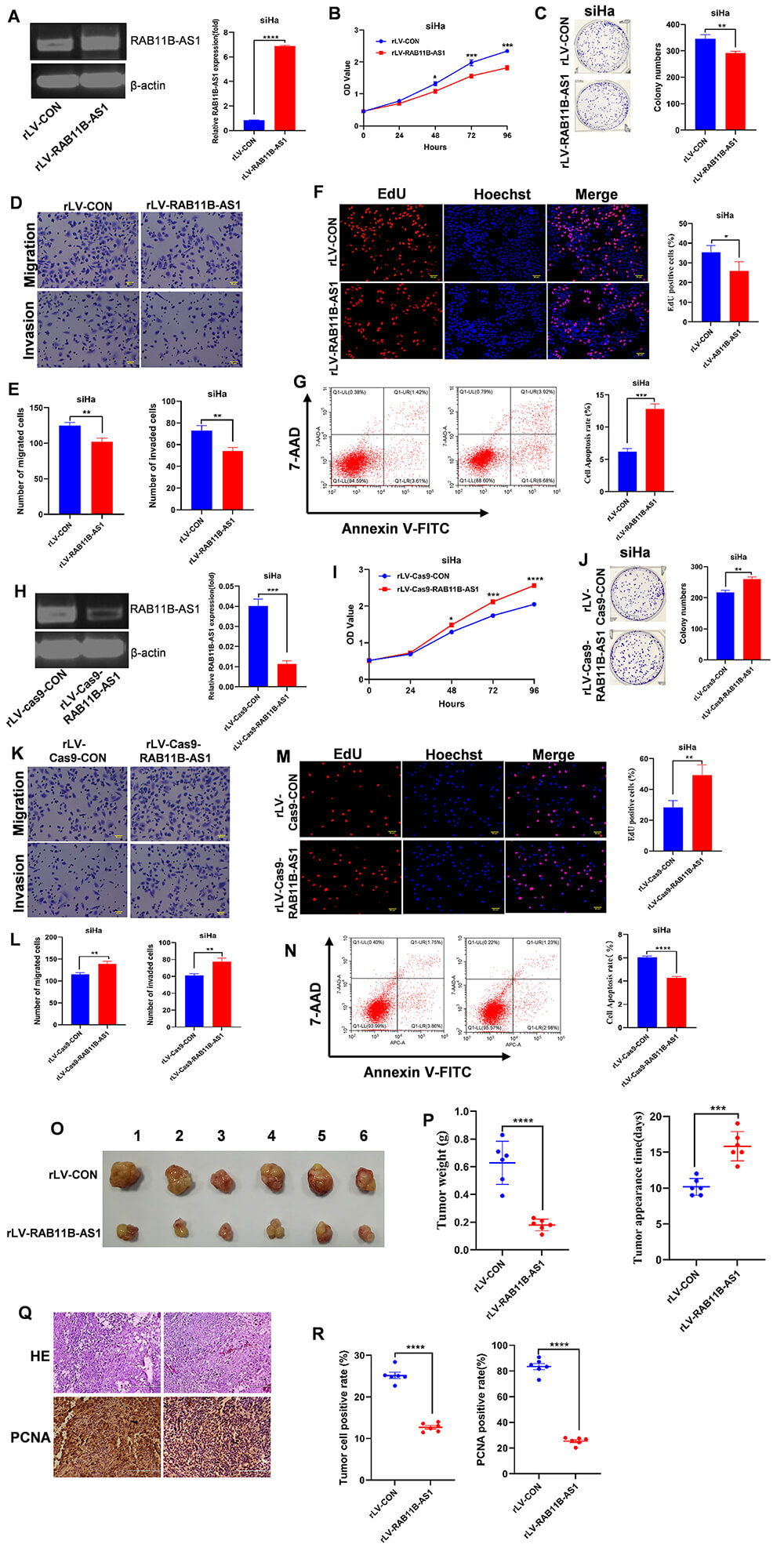 Fig. 2.
Fig. 2.
RAB11B-AS1 inhibits the tumorigenic cervical
cancer cells in vitro. (A) RT-qPCR detected RAB11B-AS1 in SiHa
cells infected with either rLV-CON or rLV-RAB11B-AS1. (B) CCK-8 assay
was used to determine the impact of RAB11B-AS1 overexpression on SiHa
cell proliferation. (C) The ability of cell colonies to form was compared between
the rLV-RAB11B-AS1 group and the rLV-CON group. Representative images
from three independent biological replicates are shown. (D,E) Pictures show cell
migration and invasion in SiHa cells infected with rLV-CON and
rLV-RAB11B-AS1. Cell migration is shown in the upper panel and cell
invasion in the lower panel. All micrographs are
To further validate the inhibitory role of RAB11B-AS1 in CC cells, we used CRISPR-Cas9 technology to generate RAB11B-AS1 knockout SiHa cells. The efficiency of the knockout was confirmed by qRT-PCR and PCR gel electrophoresis (Fig. 2H). Subsequent functional assays revealed that RAB11B-AS1 depletion significantly enhanced the proliferative capacity of SiHa cells and increased colony formation compared to control cells (Fig. 2I,J). Furthermore, RAB11B-AS1 knockout markedly promoted cell migration and invasion relative to the rLV-Cas9-CON control group (Fig. 2K,L). EdU incorporation assays corroborated the enhanced proliferation upon RAB11B-AS1 loss (Fig. 2M). Finally, flow cytometry demonstrated a significant reduction in apoptosis in RAB11B-AS1 knockout cells compared to controls (Fig. 2N). Taken together, these results demonstrate that RAB11B-AS1 suppresses the proliferation, migration and invasion of cervical cancer cells in vitro, while promoting apoptosis.
Consequently, we evaluated its function in vivo using a xenograft tumor model in nude mice. Tumors derived from rLV-RAB11B-AS1 cells exhibited significantly smaller volumes, reduced average weight, and delayed onset compared to tumors from rLV-CON control cells (Fig. 2O,P). Histological analysis (H&E staining) revealed a significant reduction in the number of poorly differentiated cells in rLV-RAB11B-AS1 tumors compared to the control group (Fig. 2Q). Further immunohistochemical analysis revealed significantly lower PCNA positivity in the rLV-RAB11B-AS1 group than in the rLV-CON group (Fig. 2R). Overall, these results suggest that RAB11B-AS1 inhibits cervical tumor growth.
As RAB11B-AS1 has been shown to inhibit the growth of human cervical cancer cells in vitro and in vivo, it is important to identify the genes and signaling pathways affected by RAB11B-AS1 in cervical tumor development. First, we performed chromatin immunoprecipitation sequencing (ChIP-seq) with anti-H3K9Ac in the two stable cell lines (Fig. 3A). Heat map analysis showed 16496 up peaks and 475 down peaks, such as RPL26, S100A14, echinoderm microtubule-associated protein like 2 (EML2), cellular retinoic acid-binding protein 2 (CRABP2), microtubule-associated protein 2 (MAP2), RPL35, RPL37, RPL28, and CDC28 protein kinase regulatory subunit 1B (CKS1B) (Fig. 3B). KEGG enrichment analysis of differentially expressed genes revealed the primary cellular pathways affected by the overexpression of RAB11B-AS1. Genes found to be downregulated following this overexpression—specifically those identified via ChIP-seq peak analysis—were significantly associated with several key biological processes, particularly ribosome, regulation of actin cytoskeleton, neuroactive ligand-receptor interaction, focal adhesion, and axon guidance (Fig. 3C). Further analyses revealed a difference in the modification ability of H3K9Ac on chromosome 17: the rLV-CON group contained 1500 peaks, whereas the rLV-RAB11B-AS1 group contained 3311 peaks. Moreover, the modification ability of H3K9Ac in the promoter regions of several genes was different. For example, six peaks appeared in the promoter region of RPL26 in the rLV-RAB11B-AS1 group and three peaks appeared in the rLV-CON group (Fig. 3D). Compared with the rLV-CON group, the scores of peaks in RPL26 were higher than those in the rLV-RAB11B-AS1 group (Fig. 3E). The pGL3-RPL26-promoter-LUC luciferase reporter gene with the RPL26 promoter sequence was transfected into two groups of cells (rLV-CON and rLV-RAB11B-AS1). The results showed that, compared with the rLV-CON group, luciferase reporter gene activity was significantly higher in the rLV-RAB11B-AS1 group (Fig. 3F). Taken together, these results suggest that RAB11B-AS1 increases H3K9Ac modification in the RPL26 promoter region.
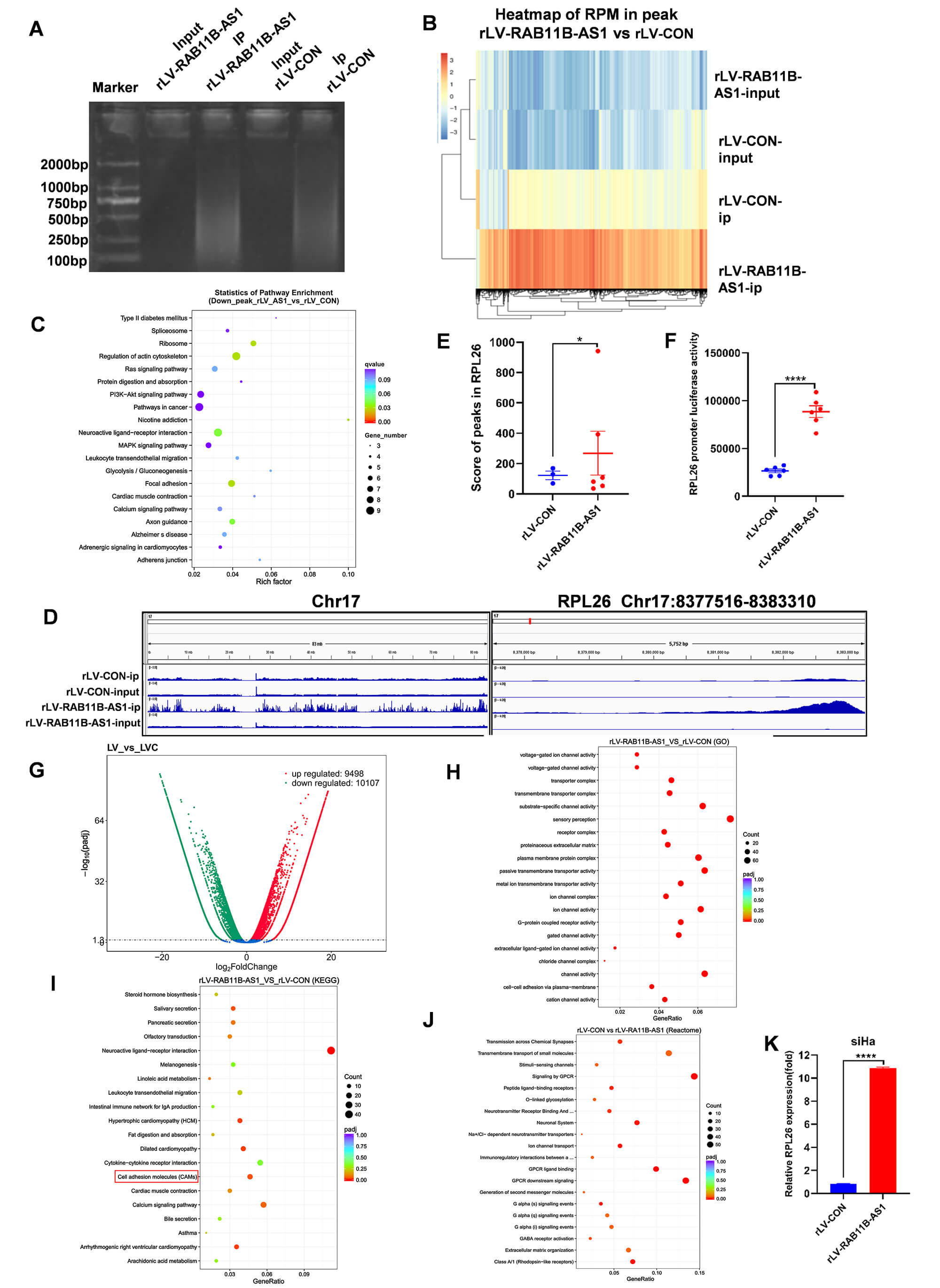 Fig. 3.
Fig. 3.
RAB11B-AS1 enhances the transcriptional activity of
RPL26 by the modification of H3K9ac. (A) IP enriched
RAB11B-AS1 content in the rLV-CON and rLV-RAB11B-AS1 groups.
(B) Heat map of RPM in peaks in the rLV-RAB11B-AS1 and rLV-CON groups.
(C) KEGG enrichment analysis of down-regulated genes after RAB11B-AS1
overexpression in ChIP-seq data. (D) Different modification ability of H3K9ac on
chromosome 17 was visually browsed using the IGV browser. H3K9ac data indicated
enhanced transcriptional activity of RPL26 in the rLV-RAB11B-AS1
and rLV-CON groups. (E) Analysis of the score of peaks in RPL26 between
the rLV-RAB11B-AS1 and rLV-CON groups. (F) Analysis of RPL26
promoter luciferase activity between the rLV-RAB11B-AS1 and rLV-CON
group. (G) Volcano plot of differential transcripts. Abscissa presents log2FC and
ordinate presents-log10 (padj). (H) The transcriptomics GO enrichment analysis is
illustrated in a scatter plot, where the GeneRatio is plotted against GO term
annotations. Dot color represents the adjusted p-value (padj), and dot
size corresponds to the number of enriched transcripts. (I) KEGG pathway
enrichment analysis of up-regulated differential genes after RAB11B-AS1
overexpression in RNA-seq results. (J) Analysis of reactome pathway enrichment.
Log10 (padj) and ordinate present reactome pathway enrichment annotation and the
number of enriched transcripts. (K) qRT-PCR showed that RPL26 was
significantly up-regulated after RAB11B-AS1 overexpression. For (E,F,K)
Data represent the mean
We sought to identify how RAB11B-AS1 affected on some genes and related signaling pathways involved in cervical tumorigenesis. Firstly, we analyzed transcriptome by RNA sequencing in the two stable cell lines. The results show that 19,605 differentially expressed transcripts were identified among the 133,924 transcripts analysed, including 9498 transcripts that were up-regulated and 10,107 transcripts that were down-regulated in the rLV-RAB11B-AS1 group compared to the rLV-CON group (Fig. 3G). In particular, the expression of RPL26 was significantly higher in the rLV-RAB11B-AS1 group than in the rLV-CON group (Fig. 3G). Transcriptome GO analysis revealed that, compared to the rLV-CON group, the rLV-RAB11B-AS1 group showed significant enrichment of gene sets in pathways related to chipmaker perception, substrate-specific channel activity, plasma membrane protein complex, passive transmembrane transporter activity, ion channel activity, and channel activity (Fig. 3H). KEGG analysis showed that the differential transcripts were highly enriched in pathways related to the neuroactive ligand-receptor interaction, calcium signaling pathway, and cell adhesion molecules (CAMs) (Fig. 3I). Reactome analysis revealed the substantial enrichment of differential transcripts in the rLV-RAB11B-AS1 group than in rLV-CON group, particularly in pathways such as G-protein coupled receptor (GPCR) downstream signaling and signaling by GPCR (Fig. 3J). Finally, the expression level of RPL26 was significantly increased in the rLV-RAB11B-AS1 group compared with rLV-CON group (Fig. 3K). Collectively, these observations suggest that RAB11B-AS1 enhances the transcriptional ability of RPL26.
Considering that RAB11B-AS1 can alter cancer related-genes at the level of transcription, we determined whether RAB11B-AS1 regulates cancer related genes at the protein level. Proteins were isolated from cells infected by rLV-RAB11B-AS1 and rLV-CON, followed by quantitative analysis. A volcano plot revealed 97 differentially expressed proteins (DEPs) out of a total of 6536 identified proteins. This subset, included 51 upregulated and 46 downregulated proteins in the rLV-RAB11B-AS1 group compared with the rLV-CON group, such as RPL26, S100A14, EML2, CRABP2, TMEM168, MAP2, RPS23, and RPL37 (Fig. 4A). Subcellular localization analysis revealed that the DEPs were mainly enriched in the nucleus (34.92%) (Fig. 4B). GO analysis showed that the DEPs were highly enriched in the rLV-RAB11B-AS1 group compared to in rLV-CON group, including protein phosphorylation (n = 199), metabolic process (n = 126), nucleus (n = 307), integral component of membrane (n = 199), protein binding (n = 1201), ATP binding (n = 527), and zinc ion binding (n = 339) (Fig. 4C). Proteomics KEGG analysis showed that the DEPs were highly enriched in rLV-RAB11B-AS1 group, including systemic lupus erythematosus, cardiac muscle contraction and hypertrophic cardiomyopathy. In particular, the differentially expressed transcripts were highly enriched in the ribosome in the rLV-RAB11B-AS1 group compared with the rLV-CON group (Fig. 4D). Protein-protein interactions analysis showed that 103 upregulated proteins (e.g., RPL26, MAP2, RPL28, RPL37) and 53 downregulated proteins involved in the protein were interaction network (e.g., S100A14, EML2, CRABP2, TMEM168) (Fig. 4E–G). Among these, the most pronounced variation was observed in RPL26 expression. Subsequent validation by western blotting revealed a significant reduction in RPL26 protein levels following RAB11B-AS1 knockout (Fig. 4H).
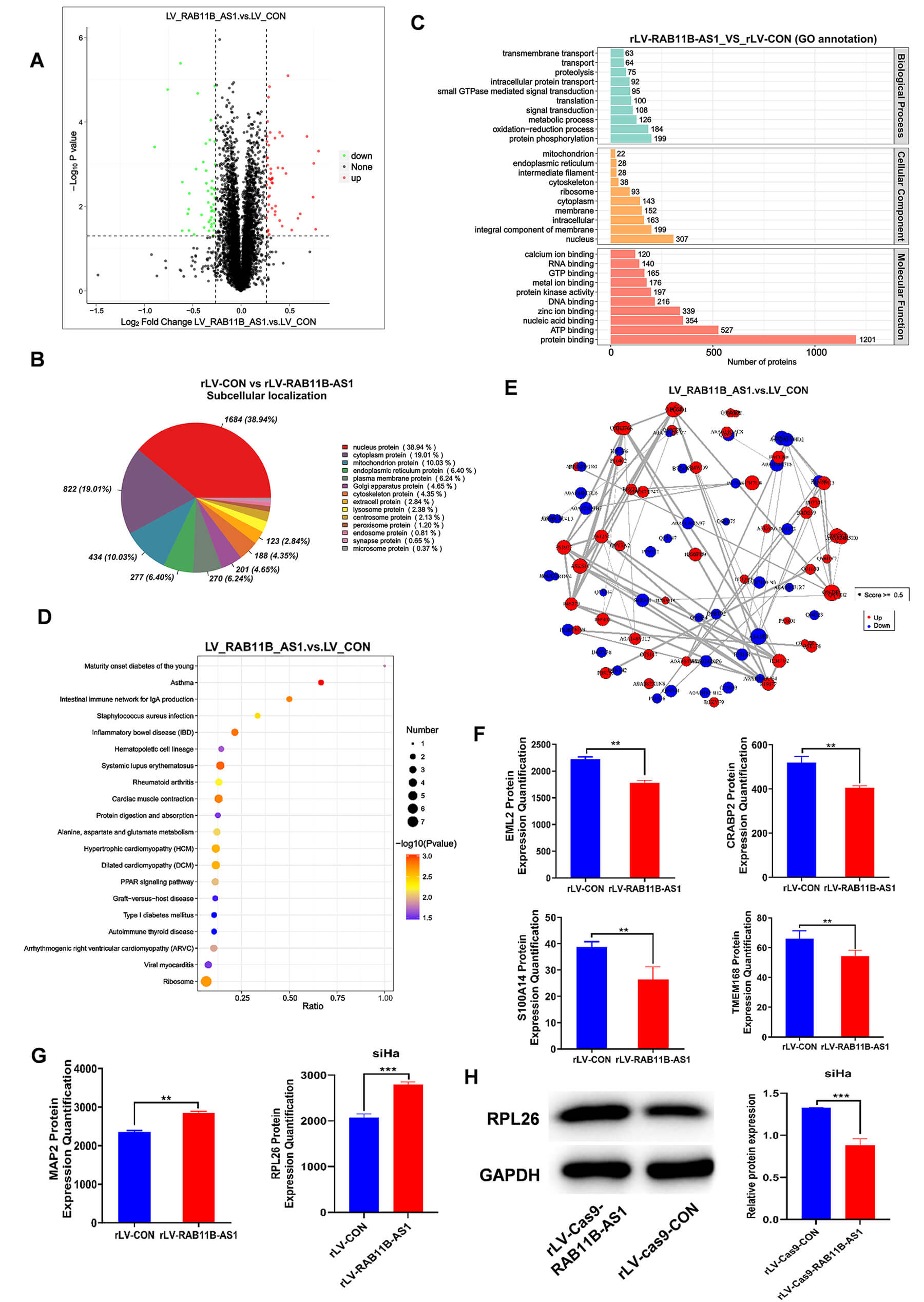 Fig. 4.
Fig. 4.
RAB11B-AS1 up-regulates the protein expression
of RPL26. (A) Volcano plots of DEPs in the rLV-RAB11B-AS1 and rLV-CON
groups. (B) Subcellular localization of DEPs. (C) Analysis of proteomics GO
enrichment. Abscissa presents the number of enriched proteins and the ordinate
presents the GO enrichment annotation. (D) Scatter diagram of proteomics KEGG
enrichment. Abscissa presents the Ratio and the ordinate presents KEGG enrichment
items. (E) Protein-protein interaction networks. Red presents upregulated
proteins and blue present downregulated proteins. (F,G) Tandem mass tag labeled
protein quantification analysis was performed in SiHa infected with
rLV-RAB11B-AS1 and rLV-CON. (H) RPL26 was detected by Western
blot analysis using antibodies against RPL26 with actin as a loading
control. All measurement data are expressed as mean
To further elucidate the downstream mechanism by which RAB11B-AS1 modulates CC progression via RPL26, we transduced RAB11B-AS1-overexpressing cells with RPL26-targeting shRNA; western-blot analysis confirmed efficient knockdown (Fig. 5A). Volcano-plot comparison of rLV-CON versus rLV-RAB11B-AS1+rLV-shRNA-RPL26 revealed 320 up-regulated genes (including S100A14, EML2, CRABP2 and TMEM168) and 725 down-regulated genes (including MAP2) (Fig. 5B). Gene Ontology enrichment analyses identified receptor-regulator activity, peptide binding and transmembrane-signaling-receptor activity as significantly enriched among the up-regulated genes in the rLV-RAB11B-AS1 group, yet these same categories were conversely enriched among the down-regulated genes in the rLV-RAB11B-AS1+rLV-shRNA-RPL26 group (Fig. 5C,D). KEGG pathway mapping further demonstrated that Pentose and glucuronate interconversions, cell adhesion molecules (CAMs), JAK-STAT signaling and cytokine–cytokine receptor interaction were up-regulated in rLV-RAB11B-AS1 cells but significantly down-regulated upon RPL26 silencing (Fig. 5E,F).
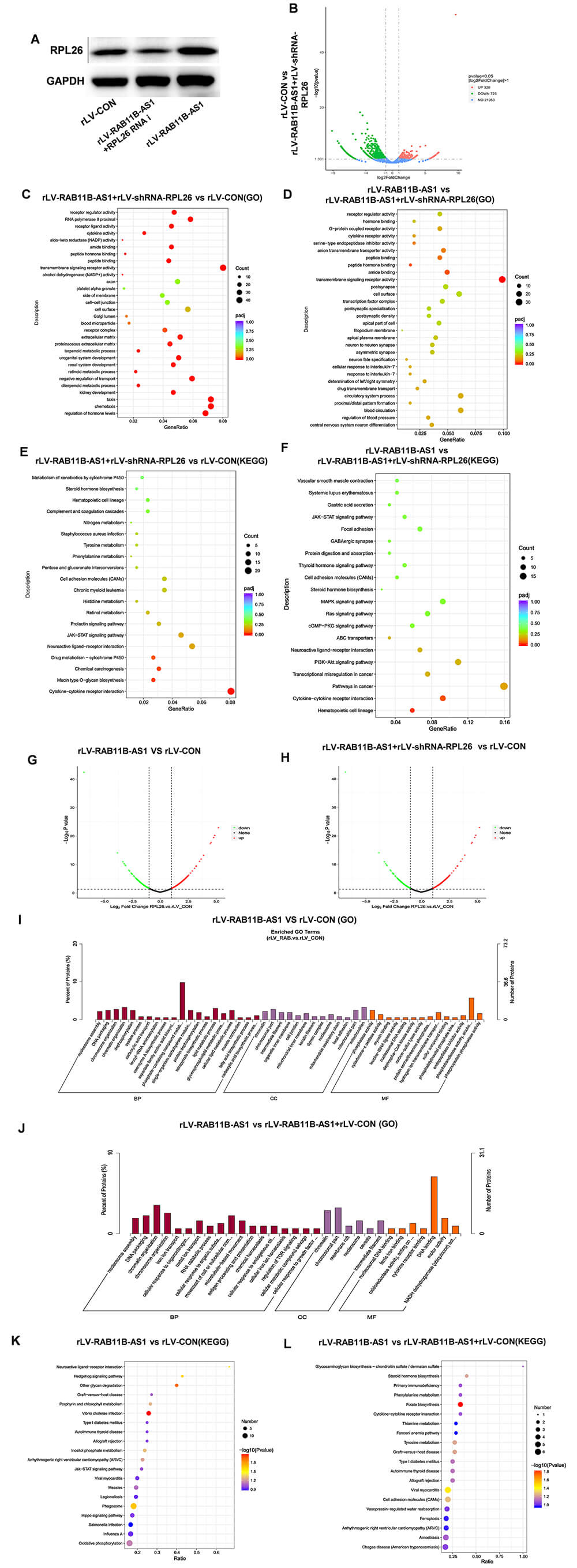 Fig. 5.
Fig. 5.
RAB11B-AS1 regulates the expression of cancer related-genes by enhancing RPL26. (A) Western blot analysis was used to detect the efficiency of RPL26 knockdown based on RAB11B-AS1 overexpression. (B) Volcano plot of differentially expressed transcripts between the rLV-CON group and the rLV-RAB11B-AS1 + rLV-shRNA-RPL26 group. (C) Scatter plot of transcriptomic GO enrichment analysis comparing the rLV-CON group and the rLV-RAB11B-AS1 + rLV-shRNA-RPL26 group group. (D) Scatter plot of transcriptomic GO enrichment analysis comparing the rLV-RAB11B-AS1 group and the rLV-RAB11B-AS1 + rLV-shRNA-RPL26 group group. (E) KEGG pathway enrichment analysis of differentially expressed genes between the rLV-CON group and the rLV-RAB11B-AS1 + rLV-shRNA-RPL26 group group. (F) KEGG pathway enrichment analysis of differentially expressed genes between the rLV-RAB11B-AS1 group and the rLV-RAB11B-AS1 + rLV-shRNA-RPL26 group. (G) Volcano plot displaying differentially expressed transcripts between the rLV-CON group and the rLV‑RAB11B‑AS1 group. The x‑axis represents log₂(fold change), and the y‑axis represents –log10 (p value). (H) Volcano plot of differentially expressed transcripts between the rLV-CON group and the rLV-RAB11B-AS1 + rLV-shRNA-RPL26 group. (I) Scatter plot of proteomic GO enrichment analysis comparing the rLV-CON group and the rLV-RAB11B-AS1 group. (J) Scatter plot of proteomic GO enrichment analysis comparing the rLV-CON group and the rLV-RAB11B-AS1 + rLV-shRNA-RPL26 group. The x axis shows the GO term annotations, and the y axis indicates the number of enriched proteins. (K) Scatter plot of proteomic KEGG pathway enrichment analysis between the rLV-CON group and the rLV-RAB11B-AS1 group. (L) Scatter plot of proteomic KEGG pathway enrichment analysis between the rLV-CON group and the rLV-RAB11B-AS1 + rLV-shRNA-RPL26 group.
To further validate the above findings, we performed a proteomic analysis. Volcano plot analysis identified a total of 505 DEPs, including 209 upregulated proteins and 397 downregulated proteins in the rLV-RAB11B-AS1 group compared with the rLV-CON group. A total of 415 DEPs were identified, including 214 upregulated proteins and 201 downregulated proteins in the rLV-RAB11B-AS1+ rLV-shRNA-RPL26 group compared with the rLV-CON group (Fig. 5G,H). Proteomics GO analysis showed that the DEPs in the rLV-RAB11B-AS1 group exhibited significant enrichment in several terms-including dephosphorylation compared to the rLV-CON group. Conversely, the GO terms, cell junction, dephosphorylation were not enriched in the rLV-RAB11B-AS1+rLV-shRNA-RPL26 group compared with the rLV-CON group (Fig. 5I,J). Proteomics KEGG analysis showed that the DEPs were highly enriched in rLV-RAB11B-AS1 group compared to the rLV-CON group, including phagosome, Hedgehog signaling pathway, and inositol phosphate metabolism. These KEGG terms, were not enriched in the rLV-RAB11B-AS1+rLV-shRNA-RPL26 group compared with the rLV-CON group (Fig. 5K,L). Collectively, these observations suggest that RAB11B-AS1 regulates the expression of cancer related-genes by enhancing RPL26.
To explore the effect of RPL26 on RAB11B-AS1 overexpression inhibiting CC, we performed in vitro and in vivo experiments. CCK-8 assays showed that compared with the rLV-CON group, the proliferation ability was significantly decreased in the rLV-RAB11B-AS1 group; however, the proliferation ability was significantly increased in the rLV-RAB11B-AS1+rLV-shRNA-RPL26 group (Fig. 6A). Furthermore, compared with the rLV-CON group, the colony formation rate was notably decreased in the rLV-RAB11B-AS1 group, but was notably increased in the rLV-RAB11B-AS1+ rLV-shRNA-RPL26 group (Fig. 6B). Next, tumorigenesis test in vivo was performed in nude mice. Compared with the rLV-CON group, the average weight of xenograft tumors was significantly decreased in the rLV-RAB11B-AS1 group. However, compared with the rLV-RAB11B-AS1 group, the average weight of xenograft tumors was remarkably increased in the rLV-RAB11B-AS1+rLV-shRNA-RPL26 group (Fig. 6C,D). Tumor onset time was significantly increased in the rLV-RAB11B-AS1 group compared with the rLV-CON group. However, compared with the rLV-RAB11B-AS1 group, the appearance time of xenograft tumors was significantly shortened in the rLV-RAB11B-AS1+ rLV-shRNA-RPL26 group (Fig. 6E). H&E staining showed that poorly differentiated cells were significantly decreased in the rLV-RAB11B-AS1 group compared with the rLV-CON group but markedly increased in the rLV-RAB11B-AS1+ rLV-shRNA-RPL26 group compared with the rLV-CON group (Fig. 6F,G). Compared with the rLV-CON group, PCNA positivity rate was significantly decreased in the rLV-RAB11B-AS1 group. However, compared with therLV-RAB11B-AS1 group, the PCNA positive rate was significantly increased in the rLV-RAB11B-AS1+rLV-shRNA-RPL26 group (Fig. 6F,H).
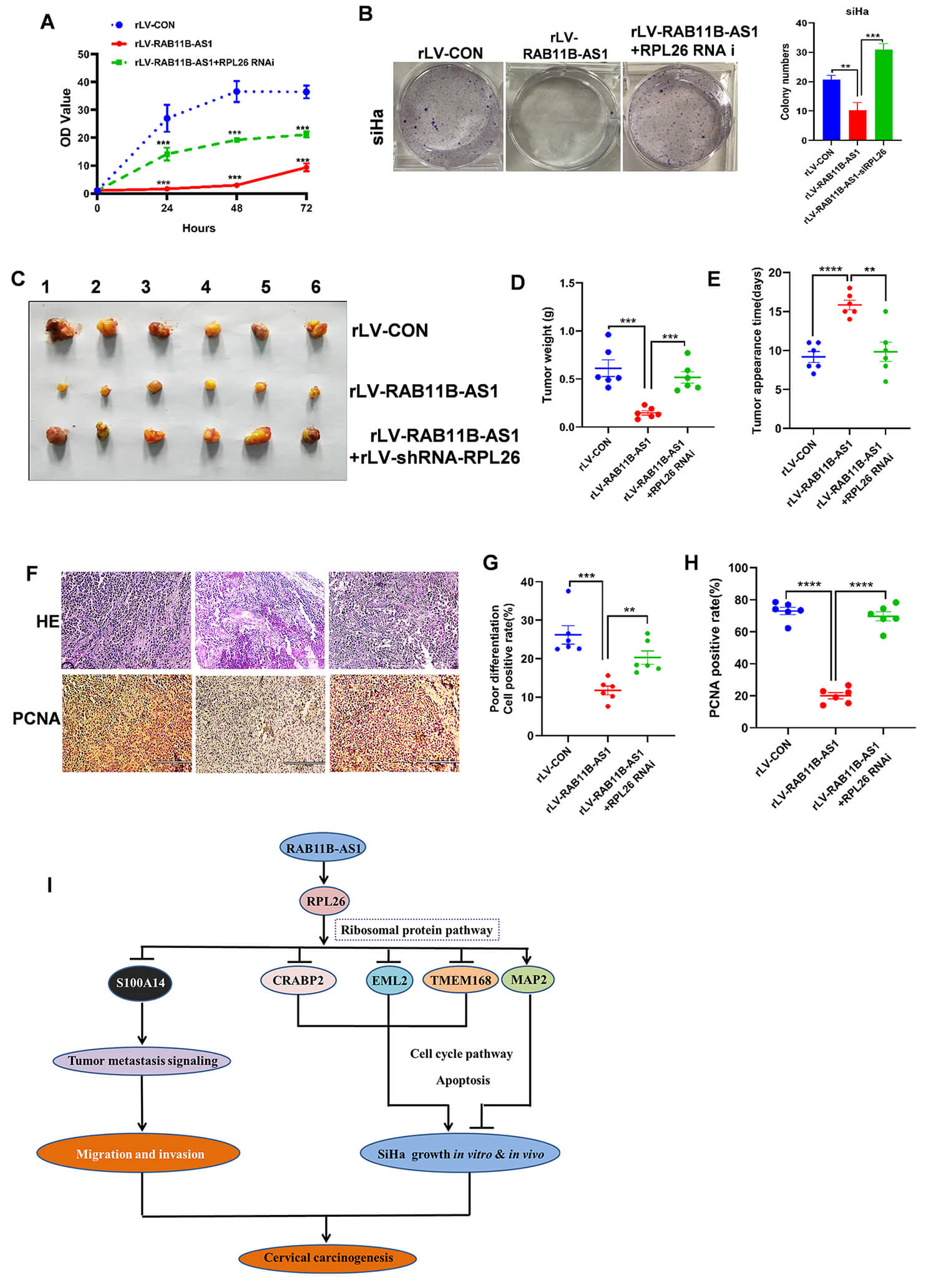 Fig. 6.
Fig. 6.
RPL26 knockdown abrogates the suppressor function of
RAB11B-AS1 in vivo. (A) CCK-8 assay was used to detect the
proliferation trend of rLV-CON, rLV-RAB11B-AS1 and
rLV-RAB11B-AS1+rLV-shRNA-RPL26 groups. (B) Cell colony
formation ability in the rLV-CON, rLV-RAB11B-AS1, and
rLV-RAB11B-AS1-rLV-shRNA-RPL26 groups. Representative images
from three independent biological replicates are shown. (C) The nude mice were
euthanized at 30 days and the transplanted tumors were imaged, including the
rLV-CON, rLV-RAB11B-AS1 and
rLV-RAB11B-AS1-rLV-shRNA-RPL26 group. (D) Comparison of average
xenograft tumor weight among the rLV-CON, rLV-RAB11B-AS1 and
rLV-RAB11B-AS1-rLV-shRNA-RPL26 group. (E) Comparison of
appearance time (days) of xenograft tumor among the rLV-CON,
rLV-RAB11B-AS1 and rLV-RAB11B-AS1-rLV-shRNA-RPL26
groups. (F) H&E staining and immunohistochemistry with anti-PCNA from xenograft
tumor among the rLV-CON, rLV-RAB11B-AS1 and
rLV-RAB11B-AS1-rLV-shRNA-RPL26 groups. The picture of H&E
staining (upper) and immunohistochemistry with anti-PCNA (lower); scale bar, 200
µm. (G) Analysis of H&E staining cell positive rate (%) from xenograft
tumor among the rLV-CON, rLV-RAB11B-AS1 and
rLV-RAB11B-AS1-rLV-shRNA-RPL26 groups. (H) Analysis of PCNA
positive rate (%) from xenograft tumor among the rLV-CON,
rLV-RAB11B-AS1 and rLV-RAB11B-AS1-rLV-shRNA-RPL26
groups. (I) Schematic diagram illustrating the mechanism by which RAB11B-AS1
inhibits cervical cancer progression through the regulation of ribosomal
proteins, cell cycle, and adhesion molecule-related pathways. All quantitative
data are presented as mean
Globally, CC is the second most frequent cause of death among women aged between the ages of 20 and 39 years [12]. 5-year overall survival rate of patients with locally advanced CC is less than 70% [4]. Recently, although great progress has been made in the diagnosis and treatment of human CC, the precise mechanisms of cervical carcinogenesis remain largely unknown and require further study. This study clearly showed that RAB11B-AS1 can inhibit the progression of CC through the regulation of cancer-related pathways based on ribosomal proteins, cell cycle and CAMs among others (Fig. 6I). These findings provide basic data for the study of human CC.
A growing body of research indicates that various oncogenic or tumor-suppressive lncRNAs in cervical cancer function as competitive endogenous RNAs (ceRNAs), sequestering specific miRNAs and thereby relieving repression on downstream target genes. This regulatory mode ultimately alters key malignant phenotypes, including cell-cycle progression, EMT, and metastatic potential [13]. For example, HIF1A-AS2 has been shown to act as a molecular sponge for miR-34b-5p, enhances cervical cancer cell proliferation and invasion while suppressing apoptosis [14]. Therefore, further elucidation of lncRNA-mediated regulatory mechanisms in cervical cancer is of substantial clinical significance and may provide new avenues for biomarker development and targeted therapy. The present study demonstrated a decreased expression of RAB11B-AS1 in cervical cancer specimens versus matched para-cancerous tissues, which was inversely correlated with pathological grade of human CC. Notably, our results suggest that RAB11B-AS1 reduced the growth ability, increases apoptosis, and inhibits cell migration in human cervical cancer cells. Disrupting calcium-dependent energy metabolism leads to apoptosis in Triple-negative breast cancer (TNBC) cells [15] . Especially, inducing apoptosis is the main goal of cancer therapy [16]. In this study, excessive RAB11B-AS1 promotes both early and late apoptosis. These findings align with existing reports, suggesting that RAB11B-AS1 likely suppresses CC by inducing apoptosis.
In particular, our results demonstrate that RAB11B-AS1 was negatively correlated with CC metastasis. Cell-cell adhesion and the extracellular matrix are well-known determinants of cancer invasion and metastasis [17]. Moreover, EMT-mediated migration promotes cancer progression [18]. Thus, we postulate that RAB11B-AS1 inhibit the CC migration and invasion by changing cell-cell junctions or the EMT. Intriguingly, our results suggest that RAB11B-AS1 regulates several cancer related-genes in CC. Studies indicate that RPL26 is the main target of ubiquitin fold modifier 1 conjugation, which is a ribosomal modification [19]. UFMylation of RPL26 is associated with endoplasmic reticulum protein homeostasis [20]. Reports suggest that RPL26 is a p53 regulator and increases p53 expression [21, 22, 23] by inhibiting the activity of HDM2 [24] and Mdm2 [25]. Studies have demonstrated that MAP2 plays a pivotal role in the neurite outgrowth of neuronal cells [26] and negatively regulates tumorigenesis [27]. The antitumor functions of RAB11B-AS1, such as inhibiting cell proliferation, migration, and invasion of CC cells [28], may be linked to its ability to increase the expression of MAP2, which is itself a known inhibitor of these cellular processes [27].
Studies have shown that the proteins echinoderm microtubule-associated protein-like-2, endothelial monocyte-activating protein 2 (EMAPII) regulate tumorigenesis [29]. In our study, we found that RAB11B-AS1 inhibited the expression of EMAPII leading us to postulate that RAB11B-AS1 inhibits CC growth through a separate function of EMAPII, which requires further investigation. Study indicates that CRABP2 is a cellular binding-protein and, an intracellular retinoic acid transporter [30]. Reports suggest that altered CRABP2 expression promotes more severe outcomes in several cancers [31] and regulates breast cancer invasion dependent on its ER status [32]. These data suggest that RAB11B-AS1 may block the progression of CC by inhibiting CRABP2. S100A14 is involved in cell proliferation, cell differentiation and tumor metastasis [33]. In particular, S100A14 promotes cell proliferation by altering the cellular G2/M phase, migration and invasion in CC [34], suggesting that RAB11B-AS1 may affect these signaling pathways by reducing S100A14 expression in CC. Consequently, RAB11B-AS1 regulation of key proteins, including RPL26, EML2, CRABP2, TMEM168, MAP2, and S100A14 is critical for inhibiting cervical carcinogenesis.
In HeLa cells, the G1/S checkpoint has been shown to be regulated by miRNAs [35]. Strikingly, our results showed that RAB11B-AS1 affects cancer-related pathways related to ribosomal proteins, cell cycle and CAMs. RAB11B-AS1 also regulates cancer metastasis-related adhesion junction signaling pathways by altering actin. Our study indicates that RAB11B-AS1 inhibits CC metastasis potentially through its involvement in cell-cell adhesion. This aligns with research indicating that the ability of cancer to invade and metastasize is tied to cell adhesion properties [36]. In particular, our results showed that RAB11B-AS1 also affects important cancer pathways by upregulating HO-1, plakoglobin, and GSTs, and downregulating TRK. The study reveals that HO-1 exerts anti-inflammatory and cytoprotective effects by regulating p53/Mdm2 signaling axis [37]. Plakoglobin is highly differentially expressed in breast cancer and contributes to breast cancer metastasis [38]. GSTs are a superfamily of enzymes that can conjugate glutathione [39] and are involved in cancer cell growth and differentiation [40]. TRK pathway aberrations are involved in the pathogenesis of many cancer types [41, 42, 43]. Taken together, our study found that RAB11B-AS1 promoted apoptosis and inhibited proliferation and metastasis through RPL26-mediated p53 pathway.
While this study provides initial insights into the oncogenic role of the long non-coding RNA RAB11B-AS1 in cervical cancer, several limitations merit consideration. First, functional assays were restricted to a single cervical cancer cell line (SiHa); thus, the generalizability of our findings remains to be validated in additional models. Second, potential off-target effects cannot be excluded, necessitating further investigation to strengthen the clinical relevance of our observations. Third, downstream mechanisms of RAB11B-AS1 were inferred solely from transcriptomic profiling, without experimental validation.
To summarize, our findings demonstrate that RAB11B-AS1 suppresses the progression of human CC by regulating key the cancer-related pathways involving ribosomal proteins, cell-cycle regulation, and CAMs. Specifically, RPL26 plays a key role for RAB11B-AS1 exerts its effects in human CC. These results offer crucial foundational data that could inform future strategies for CC treatment.
The datasets used and analyzed during the current study are available from the corresponding author upon reasonable request.
XT and YOY conceived the study and participated in the study design, performance, coordination, and manuscript writing. XG, YY, ZZ performed the research. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All procedures related to patients were carried out in accordance with International Ethical Guidelines for Biomedical Research Involving Human Subjects (CIOMS). The studies were approved by the Ethics Committee of Shanghai Tongji Hospital (Approval Number: K-W-2025-011). This study was conducted in accordance with the guiding principles of the Declaration of Helsinki. All the mouse experiments were approved by the Animal Ethics Committee of Tongji University (Approval Number: TJAB04225102). The housing of the mice was conducted in accordance with the guidelines stipulated by the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals (NIH Publication No. 8023, revised 1978). The patients/participants provided their written informed consent to participate in this study.
Not applicable.
This study was supported by a grant from the National Natural Science Foundation of China (NCSF no.81974404) and (NCSF no.81702745).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.