







 , Lucia Di Pietro 1, Konstantinos Partsinevelos 1, Saviana Antonella Barbati 2,3, Vincenzo Cardaci 4, Andrea Graziani 2, Renata Mangione 2,3, Giuseppe Lazzarino 5, Barbara Tavazzi 2,3, Valentina Di Pietro 6, Emiliano Maiani 2,3, Francesco Bellia 1, Angela Maria Amorini 1, Giacomo Lazzarino 2,3, Giuseppe Caruso 2,3,*
, Lucia Di Pietro 1, Konstantinos Partsinevelos 1, Saviana Antonella Barbati 2,3, Vincenzo Cardaci 4, Andrea Graziani 2, Renata Mangione 2,3, Giuseppe Lazzarino 5, Barbara Tavazzi 2,3, Valentina Di Pietro 6, Emiliano Maiani 2,3, Francesco Bellia 1, Angela Maria Amorini 1, Giacomo Lazzarino 2,3, Giuseppe Caruso 2,3,*1 Department of Biomedical and Biotechnological Sciences, University of Catania, 95123 Catania, Italy
2 Departmental Faculty of Medicine, UniCamillus—Saint Camillus International University of Health Sciences, 00131 Rome, Italy
3 Unit of Biology and Preclinical Research, IRCCS San Camillo Hospital, 30126 Venice, Italy
4 Vita-Salute San Raffaele University, 20132 Milano, Italy
5 LTA-Biotech srl, 95047 Paternò (CT), Italy
6 School of Infection, Inflammation & Immunology, Department of Inflammation and Ageing, College of Medicine and Health, University of Birmingham, B15 2TT Birmingham, UK
Abstract
Endometriosis is a chronic pathological condition characterized by the growth of endometrial-like tissue outside the uterine cavity and is frequently associated with severe pain, persistent inflammation, and fibrosis within the pelvic region and other parts of the body. The exact causes of endometriosis are not clear, but an innate or adaptive immune response defect has recently been suggested as a factor in the disease’s development. Carnosine is a natural dipeptide formed by the ligation of β-alanine and L-histidine and characterized by a multimodal mechanism of action that includes antioxidant and anti-inflammatory activities. Carnosine has also been shown to modulate glucose, nucleotide, and lipid metabolism as well as the response of immune cells, all processes that play a key role in the context of endometriosis. Despite numerous reviews published on the structure, role, function, and biological activities of carnosine in preclinical and clinical settings, none have focused on its therapeutic potential for the prevention or treatment of reproductive disorders, including endometriosis. In this review, after a brief introduction to the pathogenesis and pathophysiology of endometriosis, we focus on the use of carnosine for the management of reproductive disorders, concluding with its ability to modulate specific cellular and molecular mechanisms closely related to endometriosis. Given the central role of oxidative stress and inflammation across several reproductive disorders, carnosine may represent a promising therapeutic candidate not only in endometriosis, but also in broader reproductive health contexts.
Keywords
- reproductive health
- endometriosis
- carnosine
- inflammation
- oxidative stress
- iron overload
- immune system phenomena
- macrophage polarization
- microglia
- neuroprotection
Endometriosis is a chronic, estrogen-dependent gynecological disease affecting approximately 10–15% of women of reproductive age. It is characterized by the presence of ectopic endometrial-like tissue (glands and stroma) outside the uterine cavity, most commonly within the pelvic region, and is associated with symptoms such as dysmenorrhea, chronic pelvic pain, dyspareunia, and infertility [1]. The pathological lesions are associated with significant morbidity, a detrimental impact on quality of life, and considerable socioeconomic burden [2]. An increasing body of evidence has highlighted the pivotal role of oxidative stress and inflammation in the pathogenesis of endometriosis (Fig. 1) [1, 3].
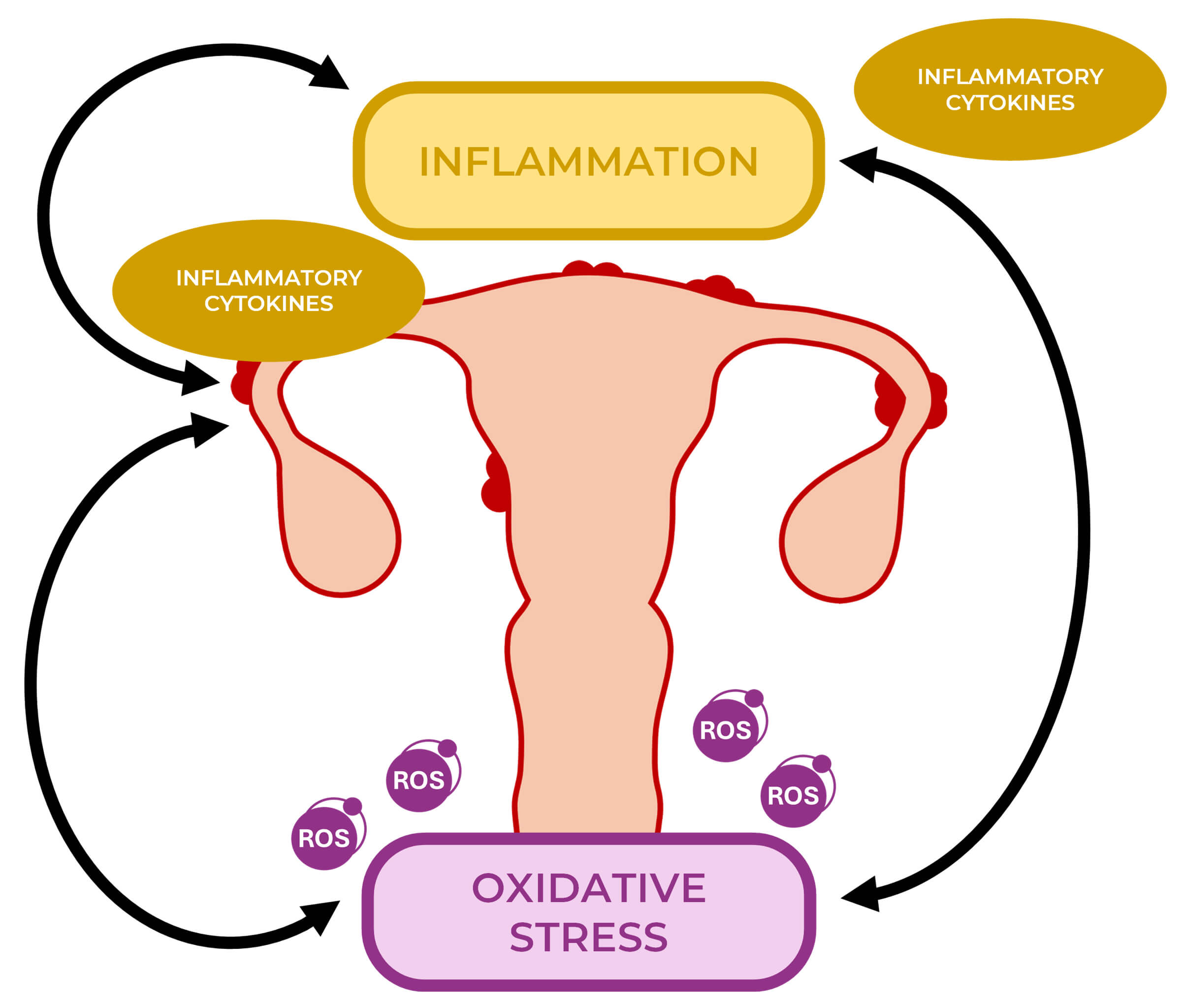 Fig. 1.
Fig. 1.
Oxidative stress and inflammation in the pathogenesis of endometriosis. Oxidative stress and inflammation are key contributors to the pathogenesis of endometriosis. The excessive production of reactive oxygen species (ROS) and the release of inflammatory cytokines create a self-perpetuating cycle that exacerbates disease progression.
Oxidative stress occurs in the presence of an imbalance between reactive oxygen
species (ROS) and antioxidant defenses, leading to oxidative damage and promoting
a pro-inflammatory microenvironment. In particular, retrograde menstruation
introduces erythrocytes and heme into the peritoneal cavity; their breakdown
generates free iron, which catalyzes the formation of ROS via Fenton reactions,
thereby damaging peritoneal tissues and contributing to lesion establishment and
progression [1, 4]. In addition, immunologic dysfunctions, such as impaired
natural killer (NK) cells clearance and the accumulation of activated
macrophages, sustain both oxidative stress and inflammation. Activated
macrophages phagocytize erythrocyte debris and release pro-oxidant and
pro-inflammatory mediators, which further exacerbate this pathologic cycle (Fig. 1) [5, 6]. Given the complex interplay between oxidative stress, inflammation,
and immune dysregulation, targeting oxidative pathways represents an important
strategy for reducing both symptomatic burden and lesion progression. In this
context, the naturally occurring dipeptide carnosine
(
Understanding the multifactorial etiology of endometriosis is essential to identify therapeutic targets. Among the different theories, the major hypotheses explaining lesion formation are represented by retrograde menstruation, iron overload, and immune dysfunction linked to oxidative stress and inflammation.
The most widely accepted theory of retrograde menstruation and implantation, originally postulated by Sampson in the 1920s, suggests that menstrual debris flows backward through the fallopian tubes into the peritoneal cavity, allowing viable endometrial cells to implant and proliferate on peritoneal surfaces [9, 10]. Although retrograde menstruation occurs in many women, the differential progression to endometriosis is thought to depend on additional factors such as iron overload and immune alterations [11]. Retrograde bleeding delivers erythrocytes and heme into the peritoneal cavity; their breakdown leads to the accumulation of free iron, which is responsible for the generation of harmful oxidative species through Fenton reactions [4]. Different studies demonstrate significantly elevated levels of free iron and ferritin in the peritoneal fluid of women with endometriosis compared to controls, with levels correlating with disease severity [12, 13, 14]. In addition, ferritin-loaded macrophages in lesions represent a further marker of iron deposition level. The resulting iron overload and ROS production cause oxidative damage, induce lipid peroxidation, and may trigger ferroptosis in peritoneal and follicular cells, potentially contributing to infertility [15, 16, 17, 18]. ROS accumulation damages lipids, proteins, and DNA within peritoneal and ectopic tissues [19]. Studies have demonstrated increased oxidative biomarker levels (including malondialdehyde (MDA) and 8-hydroxydeoxyguanosine (8-OHdG)) in peritoneal and follicular fluids of affected women [3, 20, 21, 22]. Further, oxidative damage to mitochondria triggers mtDNA leakage, activating innate immune sensors like the cyclic GMP-AMP synthase-stimulator of interferon genes (cGAS–STING) pathway, which promotes autophagy and enhances the invasive and migratory capacity of ectopic stromal cells [23, 24].
Regarding the immunologic aspect of the disease, in women with endometriosis,
immune surveillance mechanisms appear deficient [25, 26]. Key findings include
reduced NK-cell cytotoxicity, altered macrophage activation, and an inflammatory
peritoneal microenvironment [11, 27]. This dysfunction impairs clearance of
ectopic endometrial cells, promotes pro-oxidant and pro-inflammatory cytokine
release (including tumor necrosis factor-
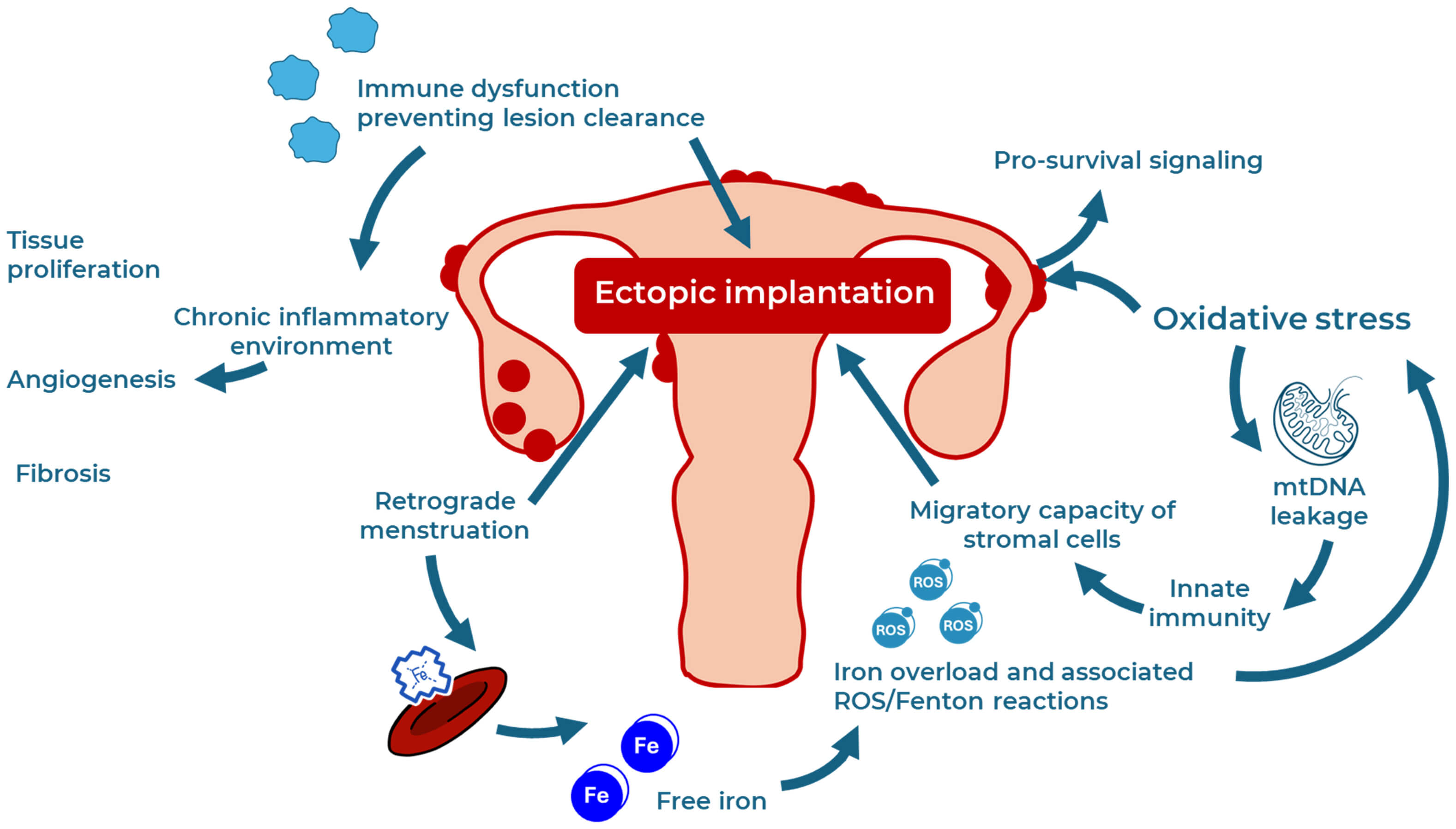 Fig. 2.
Fig. 2.
Multifactorial etiology of endometriosis. Multiple mechanisms contribute to the etiology of endometriosis. These include ectopic implantation through retrograde menstruation, iron overload with consequent ROS generation and Fenton reactions, and immune dysfunctions that hinder lesion clearance. Oxidative stress further amplifies cellular damage, leading to mitochondrial DNA leakage that sustains stromal cell migration via innate immune activation. Additionally, oxidative stress promotes pro-survival signaling in ectopic cells. The resulting chronic inflammation drives tissue proliferation, angiogenesis, and fibrosis, all hallmarks of endometriosis pathogenesis.
Macrophages play a complex and multifaceted role in the development and
progression of endometriosis [34]. In endometriotic lesions, macrophages exhibit
altered phenotypes and functions, contributing not only to impaired clearance of
menstrual debris but also to the establishment of a chronic inflammatory
microenvironment that supports ectopic tissue survival and immune evasion [34].
In this context, macrophages are not able to manage damaged erythrocytes, but
contribute to ROS and cytokine production [35]. These factors activate signaling
pathways, including nuclear factor kappa-light-chain-enhancer of activated B
cells (NF-
| Pathway | Component | Effect |
| ROS and lipid peroxidation | MDA, 8-iso-PGF2 |
Endothelial dysfunction, pain, infertility |
| Iron overload | Free iron, ferritin, Fenton reactions | ROS production, ferroptosis |
| Antioxidant depletion | Thiols, GPX, SOD, catalase, total antioxidant capacity | Sustained oxidative damage |
| Signaling | NF- |
Lesion growth, senescence, inflammation |
| Immune dysfunction | Inadequate lesion clearance |
MDA, malondialdehyde; GPX, glutathione peroxidase; SOD, superoxide dismutase;
NF-
This complex interplay underlines why oxidative stress and iron chelation are considered promising therapeutic targets, and why agents like carnosine, with both antioxidant and metal-chelating functions, deserved detailed exploration in this disease context.
Carnosine is a naturally occurring dipeptide composed of
Although direct studies in endometriosis models are not yet published, research
in non-reproductive oxidative stress models offers a very solid mechanistic
relevance. Carnosine has been reported to directly scavenge a variety of ROS,
such as superoxide anion and hydroxyl radicals, and react with
While the above-mentioned models and carnosine modulatory activity are not directly related to the reproductive system, they illustrate carnosine’s capabilities in protecting against oxidative damage in diverse cell types. Given the established role of oxidative stress, mitochondrial damage, and iron-driven ROS in endometriosis, these mechanisms offer a strong rationale for exploring carnosine in both in vitro and in vivo endometriosis models.
Despite this, some preclinical studies evaluating carnosine relevance in different female reproductive contexts are available and provide encouraging insights into its potential for endometriosis management through reduction of oxidative stress and inflammation. In an in vivo study conducted on female rats exposed to electromagnetic field, closely related to oxidative stress development, DNA damage, and deterioration of the structure and function of the cells, carnosine demonstrated the ability to prevent the loss of primordial and primary follicles, also maintaining the follicle diameter [63]. Additionally, carnosine supplementation during pregnancy in mice enhanced maternal and fetal antioxidant status, with increased SOD and GPX activity and reduced MDA in offsprings, indicating improved redox balance in reproductive tissues [64]. This evidence provides proof of concept about carnosine potential in preserving female fertility by protecting ovarian reserve and enhancing antioxidant defenses under oxidative challenge (Fig. 3).
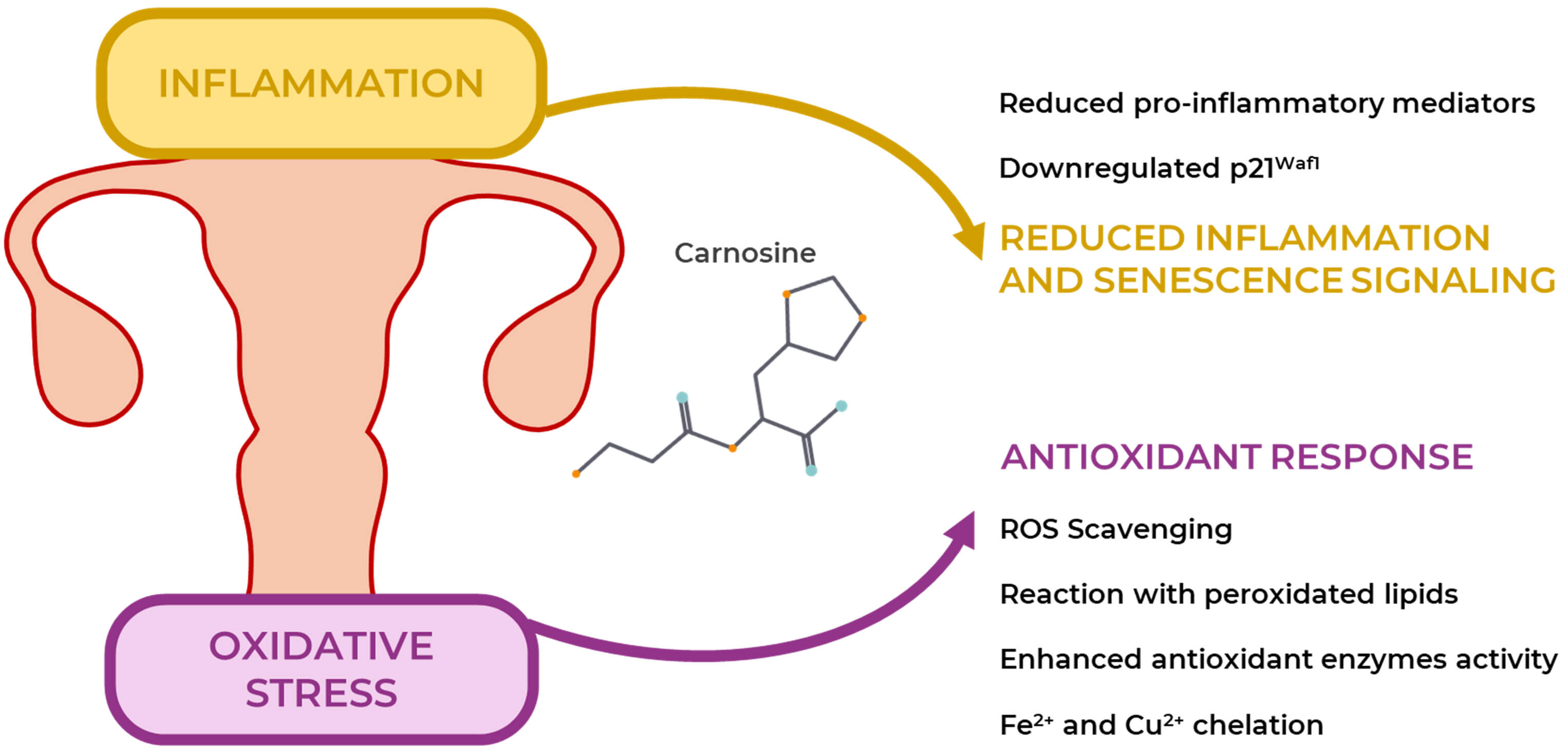 Fig. 3.
Fig. 3.
Carnosine properties in reproductive disorders. Carnosine has shown promising results in counteracting two key features of reproductive disorders: inflammation and oxidative stress. It scavenges ROS, prevents the oxidation of lipids, proteins, and DNA, activates endogenous antioxidant responses via different enzymes and chelates metal ions such as Fe2+ and Cu2+. It reduces inflammation by suppressing proinflammatory mediators and senescence-associated signaling through the downregulation of p21Waf1.
As previously discussed, iron overload plays a pivotal role in promoting oxidative stress through Fenton chemistry, a process relevant to endometriosis and other chronic inflammatory conditions. Carnosine, due to its imidazole group, exhibits significant iron-chelating properties, which contribute to its antioxidant and cytoprotective effects. Mozdzan et al. [65] demonstrated that carnosine effectively chelates Fe2+ and Cu2+ ions and reduces hydroxyl radical generation in vitro, suggesting that its metal-binding capacity could attenuate iron-driven oxidative damage. Similarly, results provided by Kang showed that carnosine and its analogues (e.g., homocarnosine) prevent DNA damage induced by ferritin and H2O2, further underlining a protective role against iron-mediated ROS generation [66]. These findings are further supported by recent in vivo studies. In a mouse model of chronic kidney disease with iron overload, carnosine administration reduced non-heme iron accumulation in tissues and lipid peroxidation levels, while improving redox balance and hemoglobin content [67]. The authors proposed the formation of Fe2+-GSH-carnosine ternary complexes as a mechanism of detoxification. A different study reported that oral carnosine administration was able to mitigate the adverse cardiac remodeling associated with diet-induced obesity in a mouse model of enhanced lipid peroxidation (GPX4 deficient mice). In this context, carnosine significantly reduced iron levels and suppressed collagen-cross-linking in myocardial tissue, strengthening its well-known antifibrotic activity [68]. Collectively, these studies suggest that carnosine may offer therapeutic benefits in disorders involving iron overload by chelating labile iron, preventing hydroxyl radical formation, and activating endogenous antioxidant pathways. This mechanism may be particularly relevant in endometriosis, where excess of iron and the related oxidative stress sustain lesion persistence and infertility [69].
Carnosine exerts significant immunomodulatory effects on macrophages,
influencing both oxidative stress and inflammatory signaling [70, 71, 72]. In
lipopolysaccharide (LPS) + interferon-
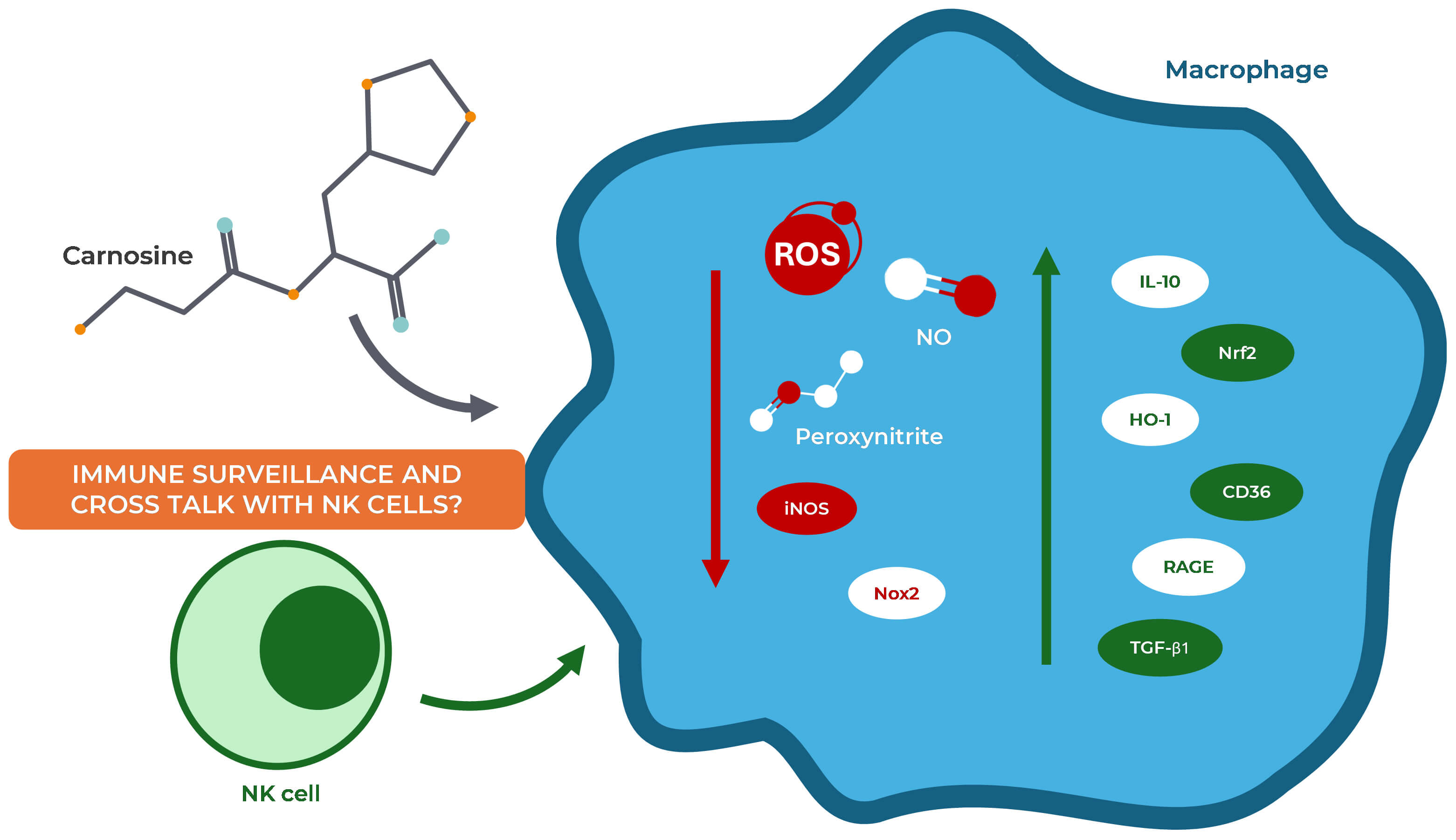 Fig. 4.
Fig. 4.
The role of carnosine in immune response. Carnosine modulates
macrophage activity by reducing the production of ROS, NO, and peroxynitrite. It
attenuates inflammation by downregulating iNOS and Nox2 expression, while
enhancing the anti-inflammatory response through the upregulation of IL-10, Nrf2,
HO-1, CD36, RAGE, and TGF-
Given that endometriosis is characterized by a dysregulated immune response, including defective clearance of ectopic endometrial cells, aberrant macrophage polarization, and impaired T cell and NK cell activity, carnosine’s immunomodulatory effects may offer therapeutic benefit by rebalancing both innate and adaptive immune components within the peritoneal microenvironment.
Beyond the well-established peripheral mechanisms, recent evidence suggests that
neuroinflammation and central nervous system regulation may also represent
underappreciated contributors to reproductive disorders. In the context of
endometriosis, which is frequently associated with central sensitization and
chronic pelvic pain, exploring the role of glial cells provides a novel point of
view to understand how neuroimmune interactions may influence fertility. In
particular, an alternative approach to addressing fertility challenges in women
has been recently proposed by Desroziers [77], who highlighted an interesting and
unconventional link between glial cells and PCOS. In her review, Desroziers [77]
underscores how glial cells, including astrocytes and microglia, can structurally
and functionally modulate neurons related to the gonadotropin-releasing hormone
(GnRH), allowing increased pulsatile or release of GnRH via morphological
remodeling of glial processes. In PCOS-like animal models, abnormal neuronal
wiring, related to increased GABAergic synaptic inputs to GnRH neurons,
correlates with impaired synaptic pruning and suggests a potential, although not
yet fully elucidated, role for glial-mediated shaping of neural circuits. This
concept leads to a captivating hypothesis that glial dysfunction may contribute
to neuroendocrine dysregulation in PCOS by allowing enhanced excitatory input
persistence to GnRH neurons, driving LH hypersecretion and the resultant hormonal
and ovarian symptoms [77, 78]. Interestingly, carnosine exerts multiple
modulatory effects on glial cells that could be linked to these mechanisms. In a
study on human HMC3 microglial cells, carnosine significantly reduced NO
production and improved mitochondrial ATP/ADP ratio [79]. When the same human
cells were challenged with a pro-oxidative and pro-inflammatory stimulus
represented by the combination of LPS and ATP, results obtained by HPLC analysis
reported the ability of carnosine to modulate ROS production and restore the
basal energy metabolism of the glial cells [80]. Moreover, in BV-2 murine
microglial cultures challenged with A
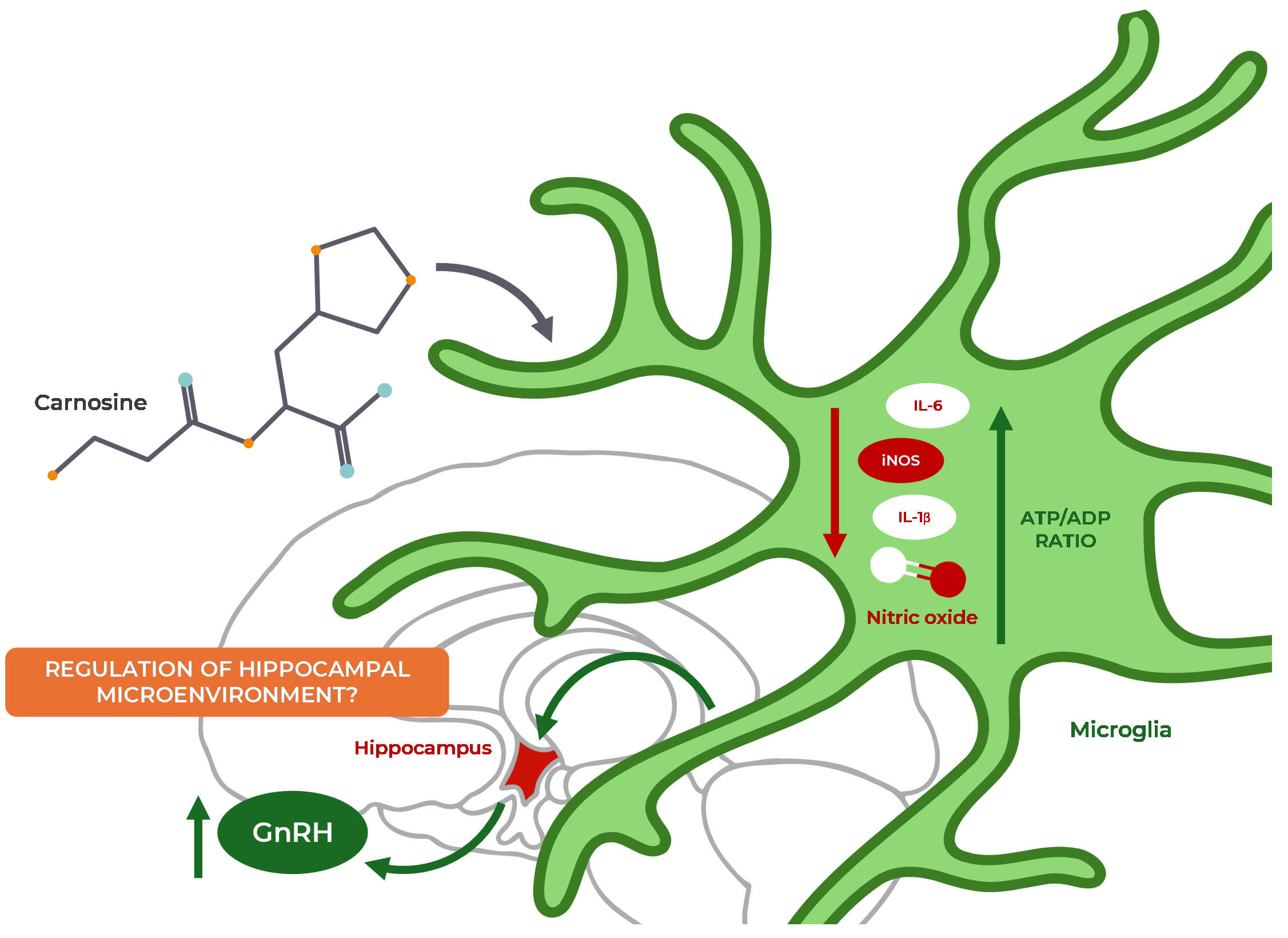 Fig. 5.
Fig. 5.
Proposed mechanisms linking carnosine, microglial regulation,
and fertility. Carnosine is a promising regulator of fertility due to its
antioxidant and anti-inflammatory effects on microglia. It reduces the production
of pro-inflammatory cytokines (IL-6, IL-1
The same model was also employed to assess the transcriptional regulatory
activity of carnosine on glial cells in A
Endometriosis exhibits several disease-specific features that differentiate it
from other chronic inflammatory and oxidative disorders. Lesions are strongly
estrogen-dependent, with aberrant hormone signaling leading to proliferation and
survival of ectopic endometrial cells [85]. Moreover, the progressive fibrotic
remodeling of peritoneal lesions, mediated by excessive extracellular matrix
deposition and myofibroblast activation, represents a distinctive hallmark of
endometriosis [86, 87, 88]. In parallel, the immune microenvironment is characterized
by impaired NK cell cytotoxicity, altered macrophage polarization, and sustained
release of pro-inflammatory cytokines such as TNF-
These features provide a rationale to hypothesize specific mechanisms through which carnosine might exert beneficial effects in endometriosis. Beyond its antioxidant and metal-chelating activities, carnosine shows antiglycating properties that could attenuate fibrotic progression by limiting advanced glycation end-products and tissue stiffening [68, 95]. Its immunomodulatory action on macrophages and cytokine release may help restoring immune surveillance within endometriotic lesions [73, 96]. Furthermore, evidence of carnosine’s ability to regulate glial activation and neuroinflammatory signaling strengthen its potential role in alleviating pain and neuroendocrine alterations disease-associated [50, 97]. Although direct studies in endometriosis are lacking, these unique disease-specific aspects point to potential multimodal mechanisms through which carnosine may act, encouraging further research.
While the focus has been on female reproductive disorders so far, it is important to underline that oxidative stress, immune dysregulation, and metabolic imbalance are also major features of male infertility. These shared pathogenic pathways suggest that carnosine’s antioxidant and cytoprotective effects may extend beyond female contexts, providing benefits in male reproductive health as well. In particular, male infertility is classically related to oxidative insults to spermatozoa, leading to decreased motility, DNA fragmentation, and mitochondrial dysfunction [98]. In different preclinical models of reproductive toxicity in male animals, carnosine demonstrated protective effects via antioxidant and anti-glycating pathways. For instance, in male rats treated with cyclophosphamide hydroxydaunomycin, oncovin, and prednisone (CHOP), a combination of chemotherapeutics commonly used to induce gonadotoxicity in experimental models, carnosine supplementation preserved testicular function, reduced lipid peroxidation, and decreased oxidative DNA damage [99]. Carnosine was also tested in a different model of testicular toxicity induced by sodium valproate, in which the dipeptide, along with Coenzyme Q10 co-administration, was able to increase the levels of reproductive hormones such as testosterone, FSH, and LH in serum, thereby increasing the levels of biochemical parameters such as SOD, GPX, and catalase [100]. Additionally, carnosine was shown to mitigate testicular aging induced by galactose exposure through its anti-glycating and ROS-scavenging properties [101]. Further support for carnosine’s cytoprotective role in the male reproductive system derive from a model of malnutrition-induced hypogonadism, where rats fed with a protein-deficient diet exhibited severe reductions in testicular weight, sperm count and viability, along with hormonal imbalances and increased pro-inflammatory and apoptotic markers in testicular tissue [102]. Carnosine administration reversed these alterations by restoring antioxidant defenses and anti-inflammatory activity. Similarly, in a model of lead (Pb)-induced reproductive toxicity, characterized by increased oxidative stress, mitochondrial dysfunction, and poor sperm parameters, carnosine supplementation alleviated these alterations, confirming its protective role for mitochondria and redox homeostasis [103]. Beyond animal models, carnosine has also demonstrated promising results in human sperm manipulation contexts. When added during semen processing, carnosine improved mitochondrial activity and beat-cross frequency (BCF), supporting its potential in assisted reproduction technologies [104]. These beneficial mitochondrial effects were reproposed and assessed in studies on quail sperms, where carnosine, present in seminal plasma, improved different motility parameters after in vitro storage, suggesting an innovative and critical function of imidazole dipeptides in sperm preservation [105]. This function appears significantly relevant in semen cryopreservation, where oxidative stress is a critical factor. In stallion semen, higher carnosine levels were associated with better tolerance to cooling and freezing, and with reduced MDA levels, proving that carnosine was effective in removing lipid peroxidation products. These findings suggest that carnosine may act as a natural buffer against cryo-induced oxidative insults, potentially enhancing sperm resistance during biotechnological processes [106]. This evidence confirms that carnosine supports male reproductive health by attenuating oxidative damage, preserving mitochondrial integrity, and sustaining hormonal and spermatogenic homeostasis under stress conditions (Fig. 6).
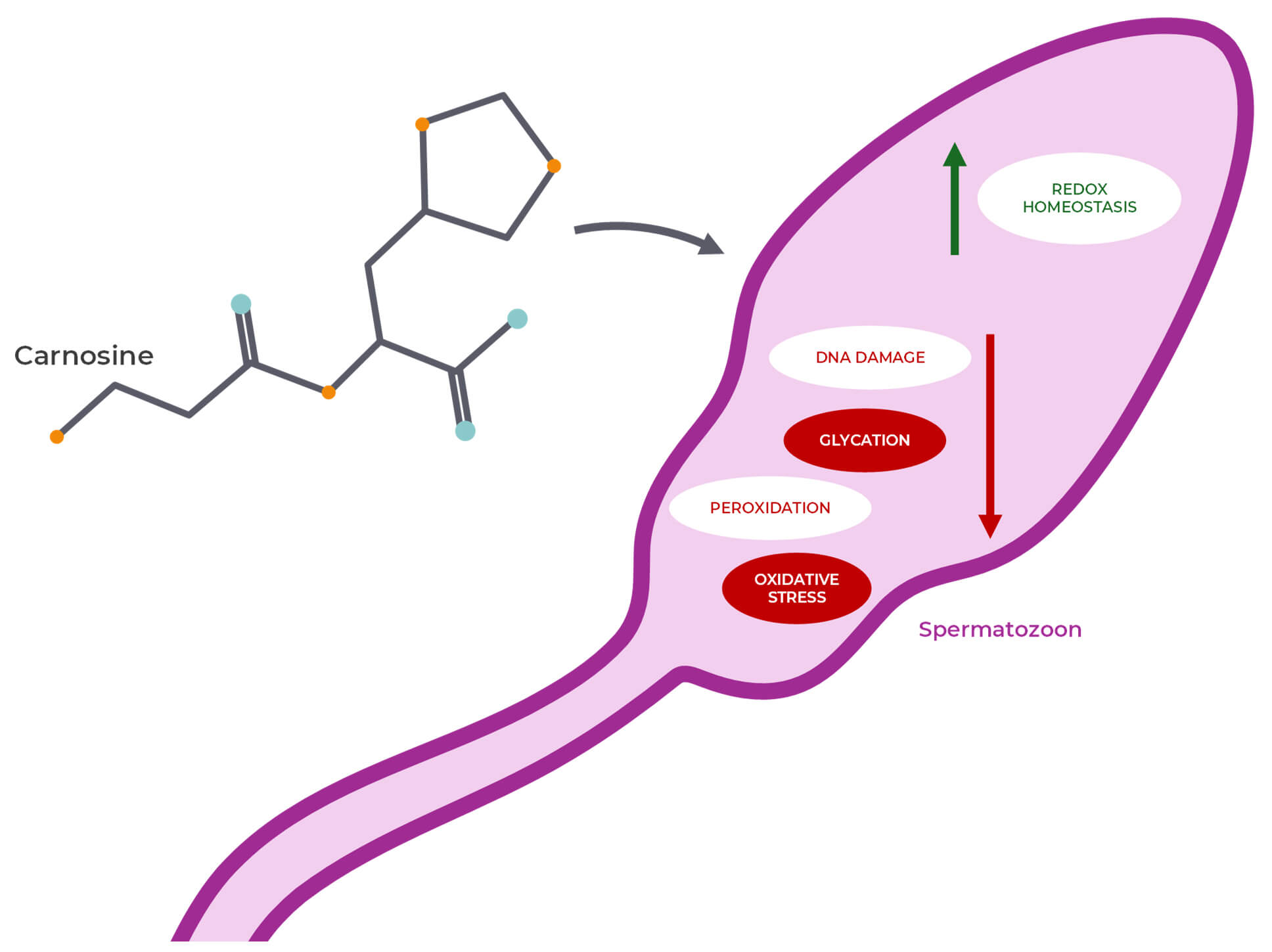 Fig. 6.
Fig. 6.
Role of carnosine in male reproductive health. Carnosine has shown protective activity towards male reproductive health, decreasing DNA damage and glycation, along with oxidative stress and the related peroxidation of lipids. Overall, it also allows an improvement of redox homeostasis.
Although endometriosis represents a different disease related to women, the oxidative and immune pathways implicated in its pathogenesis show a notable overlap with those involved in male infertility. This overlap suggests that carnosine, due to its broad-spectrum cytoprotective actions, could be considered as a supportive intervention in both contexts.
To date, no clinical trial has specifically evaluated carnosine supplementation
in women with endometriosis. However, human studies in related inflammatory and
oxidative contexts provide translational insights. A meta-analysis of randomized
controlled trials involving histidine-containing dipeptides (including carnosine)
reported significant reductions in systemic oxidative markers (e.g., MDA and
8-OHdG) and inflammatory markers (e.g., C-reactive protein (CRP) and
TNF-
There are numerous evidence showing that the natural dipeptide carnosine possesses a therapeutic potential in the context of human reproduction. It has shown to exert its antioxidant and potential protective effects on sperm and reproductive tissues. Additionally, carnosine has shown to play a role in several aspects of female reproduction, including ovarian health, fetal development, and potentially influencing pregnancy outcomes. In particular, studies have shown the ability of carnosine to protect ovarian follicles from damage caused by electromagnetic fields, potentially improving fertility. Furthermore, maternal supplementation with carnosine has shown promise in enhancing fetal growth and development in animal models. In addition to the above direct positive modulatory effects, carnosine has shown to be able to modulate endometriosis-related markers as well as macrophages and microglia, the latter emerging as an innovative regulator of female fertility, and in particular in the context of endometriosis.
In summary, carnosine has shown very promising results in supporting reproductive health, but further research is needed to fully understand its therapeutic potential on reproductive disorders, also strengthening the possible benefits of carnosine administration for prevention and/or treatment of endometriosis. In this context, a critical limitation of the current literature is represented by the absence of direct in vitro and in vivo studies using established endometriosis models (e.g., rodent models of endometriosis, human endometriotic stromal cell cultures), along with the lack of clinical studies focused on the role of carnosine in the management of endometriosis markers and symptoms. Carnosine’s multimodal potential including antioxidant, chelating, and immunomodulatory properties is deeply reported in the literature, and systematic reviews of clinical studies on the role of oxidative stress and the potential of antioxidant therapy in endometriosis identify several candidates that have been tested, but reports on carnosine are still missing [109, 110, 111]. Given the absence of direct carnosine-endometriosis studies, in the present review we chose to focus on mechanistic and preclinical evidence regarding the multimodal potential of this dipeptide, and on well-established pathogenic pathways in endometriosis (iron overload, ROS, immune dysfunction) to justify the consideration of carnosine as a possible therapeutic candidate. This approach is intentionally hypothesis-generating: the conclusions drawn here are provisional and aim to motivate dedicated new in vitro, in vivo, and early clinical studies. Future research must prioritize investigating the effects of carnosine in this specific disease to validate the promising mechanisms proposed herein as well as to determine optimal dosing and delivery strategies.
8-isoPGF2
Project administration and conceptualization of the manuscript: GCaro and GCaru; literature search: GCaro, LDP, KP, SAB, VC, AG, RM, GiuL, BT, VDP, EM, FB, AMA, GiaL, and GCaru; writing—original draft: GCaro and GCaru; preparation of the figures: GCaro, LDP, and GCaru; writing—review & editing: GCaro, LDP, KP, SAB, VC, AG, RM, GiuL, BT, VDP, EM, FB, AMA, GiaL, GCaru. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This work was supported by the Italian Ministry of Health (Ricerca Corrente) and partially supported by PRIN 2022 grant “Counteracting Human Infertility Pathophysiology (CHIP)”, number 2022KREEEF- University of Catania.
The authors declare no conflict of interest. In particular, the judgments in data interpretation and writing were not influenced by the relationship between Prof. Giuseppe Lazzarino and LTA-Biotech srl.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.