





 , Ashlyn P. McClelland 1
, Ashlyn P. McClelland 11 Department of Neuroscience, Rowan-Virtua School of Osteopathic Medicine, Virtua Health College of Medicine and Life Sciences of Rowan University, Stratford, NJ 08084, USA
Abstract
The mammalian tongue is an intricate skeletal muscle organ. From its initial formation to maturation, tongue muscle development involves precisely coordinated processes during embryonic and fetal phases of myogenesis. Extensive research on the regulatory pathways involved in tongue epithelial taste organ development has shown that the Hedgehog (HH) signaling pathway is vital to the formation and epithelial patterning of the tongue and taste organs. Emerging evidence also points to its involvement in the initial formation and spatial patterning of the tongue muscle. HH signaling is a well-established regulator of skeletal muscle development, particularly in limb myogenesis. However, structural and functional differences between limb and tongue muscles, as well as variations in their HH signaling regions, prevent the direct application of findings from limb muscles to the tongue. Consequently, a comprehensive comparative analysis is essential to establish the conserved and divergent mechanisms by which HH signaling operates in these distinct muscle systems. A detailed mechanistic understanding of HH signaling during lingual muscle formation and maturation is vital for fully elucidating its role in tongue function. Further, lingual myogenesis studies pave the way for potential regenerative therapeutic strategies for congenital anomalies and acquired conditions affecting the tongue. Thus, understanding the regulatory mechanisms of tongue muscle development has both biological and clinical importance. This review explores the role of HH signaling throughout the key stages of embryonic tongue muscle development (including myoblast determination, proliferation, differentiation, patterning, and maturation) and compares its role in limb myogenesis.
Keywords
- skeletal muscle
- tongue
- muscle development
- signal transduction
- taste
The mammalian tongue is a highly specialized skeletal muscle [1]. However,
unlike other skeletal muscles, intrinsic tongue muscles are covered by dorsal
epithelium and lamina propria comprising mesenchymal cells. Skeletal muscle
development in the embryo begins with somatic epithelial cells transitioning into
mesenchymal cells, which then differentiate into muscle cells [2, 3]. This
process, known as myogenesis, involves distinct stages regulated by specific
myogenic regulatory factors (MRFs). The MRFs are further modulated by signaling
pathways, including Hedgehog (HH), Wnt, Notch, bone morphogenetic protein (BMP)
and transforming growth factor beta (TGF-
The tightly regulated process of myogenesis involves several distinct stages: myoblast determination, migration, proliferation, differentiation, patterning, and maturation (Fig. 1). It occurs in two distinct phases: embryonic and fetal myogenesis [11]. Embryonic myogenesis initiates the process of muscle development with somites (blocks of mesoderm) differentiating into the dermomyotome (transient epithelium), which delaminates and gives rise to the myotome, the source of skeletal muscle precursors [2, 3]. During the determination stage, mesodermal muscle precursors migrate to their target locations, commit to the myogenic lineage, and develop into myoblasts. This commitment and migration are regulated by transcription factors, primarily Paired Box 3 (Pax3) and its orthologue, Pax7.
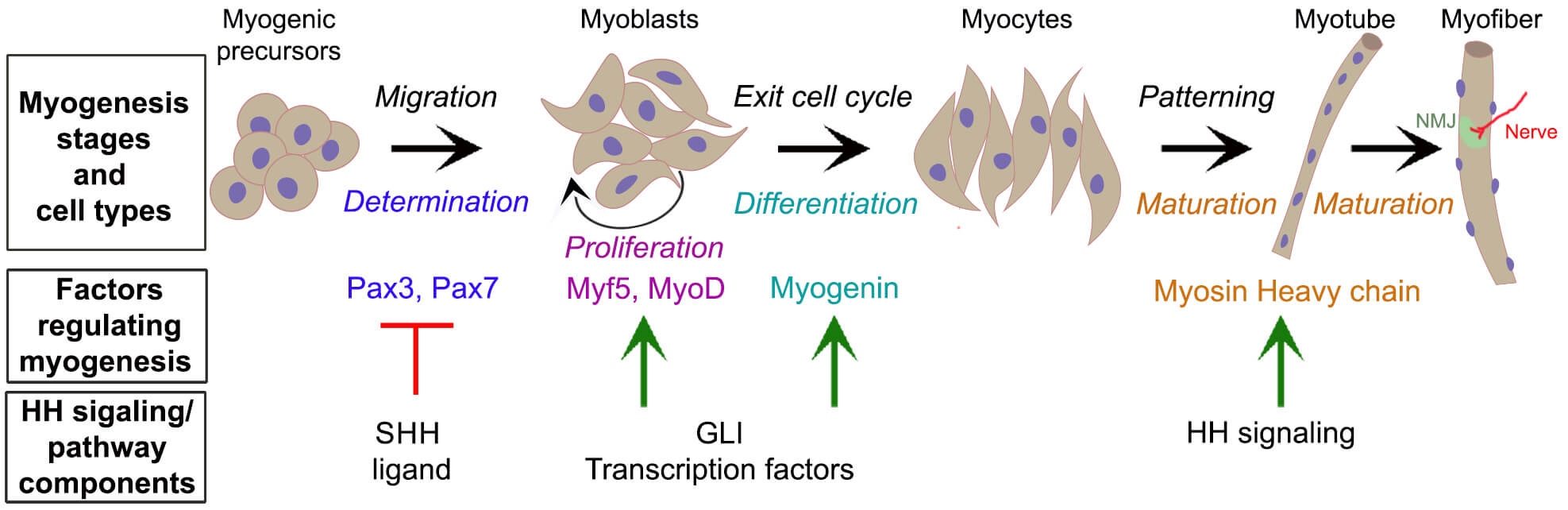 Fig. 1.
Fig. 1.
Hedgehog signaling regulates myogenic factors involved in distinct stages of myogenesis. Schematic diagram illustrating the key myogenic cell types at the various myogenesis stages. Myogenic progenitors migrate to the target site (Migration) and commit to the myogenic lineage (Determination), becoming myoblasts. These myoblasts proliferate (Proliferation) and differentiate (Differentiation) into myocytes, which align and fuse (Patterning) to form multinucleated, immature myotubes (Maturation). These myotubes make synapses with nerves at neuromuscular junctions (NMJ) and become functional myofibers (Maturation). The factors regulating myogenesis are color-matched to their respective stages. These factors are modulated by HH signaling pathway/components, either through inhibition (red line) or activation (green arrows). HH, Hedgehog; SHH, Sonic HH.
As muscle progenitor cells migrate to their destination, they undergo continuous proliferation [12]. Pax3 and Pax7 activate MRFs Myogenic Factor 5 (Myf5) and Myogenic differentiation 1 (MyoD), driving myoblast proliferation [13]. The differentiation stage in the target locations commences with myoblasts exiting the cell cycle and differentiating into myocytes under the antiproliferative effect of myogenin. As myoblasts differentiate, they begin to align, express muscle-specific proteins, and prepare to fuse with other myocytes to form multinucleated myotubes. This aggregation into myotubes marks the transition from progenitor cell to functional muscle cell [14]. Thus, myogenic differentiation is coordinated with an intricate fusion pattern to create their spatial organization [15]. In the final maturation stage, myotubes develop into mature muscle fibers, acquire striations, establish innervation, and develop contractile function. The contractile protein myosin heavy chain (MyHC), predominantly MyHC-embryonic, regulates muscle fiber size, number, and type, and myogenic differentiation genes [16]. Several other MRFs are also required to precisely control myogenesis stages [17].
Embryonic myogenesis, characterized by the formation of primary myofibers, occurs between embryonic day (E) 10 and E12.5 in mouse limb muscle. Fetal myogenesis, which gives rise to secondary myofibers, happens between E14.5 and E17.5 [18]. While embryonic myogenesis is crucial for the initial formation of the organ, fetal myogenesis establishes the muscular foundation necessary for critical postnatal functions [19]. Marked by a shift to fetal myoblasts, which are distinct from their embryonic counterparts, fetal myogenesis promotes muscle proliferation, differentiation, patterning, and functional maturation [11]. This process also establishes the satellite cell population necessary for postnatal muscle growth and regeneration [20]. Thus, the regulation of both embryonic and fetal myogenesis is necessary for proper muscle formation and growth in developing limbs.
Tongue muscle development begins at E10.5 (Fig. 2) with the formation of a central median lingual swelling on the first mandibular arch, followed by a lateral lingual swelling on each side comprising an epithelium and cranial neural crest cell (CNCC)-derived mesenchyme [21]. Muscle progenitors migrate from the somites into the tongue primordium within the lingual swellings starting at E11.5. These lingual swellings subsequently fuse to form the anterior two-thirds of the tongue; the posterior third arises from the third and fourth branchial arches [21]. Following fusion of the lingual swellings, myogenic progenitors develop into myofibers that occupy most of the tongue. HH signaling is known to regulate myoblast determination, tongue formation [22], and initial myofiber patterning [23], but its role in muscle proliferation, differentiation, and maturation in tongue remains poorly understood.
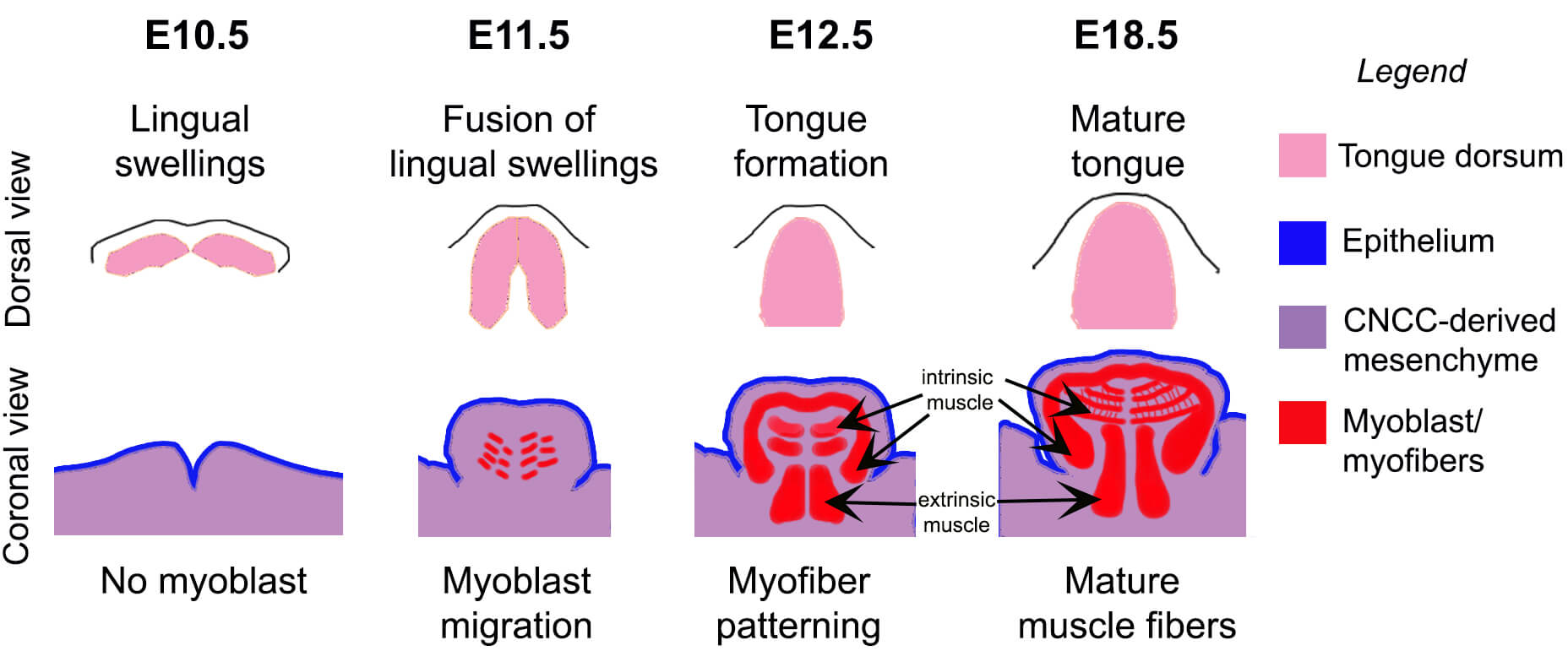 Fig. 2.
Fig. 2.
Stages of mouse embryonic tongue formation. During embryonic day (E)10.5, lingual swellings consisting of both epithelium and mesenchyme become visible. By E11.5, these swellings fuse, initiating the formation of the tongue and allowing the arrival of myoblasts. By E12.5, the anterior tongue body is formed, and myofibers started to acquire patterning. Tongue growth continues and, by E18.5, muscle fibers mature and finalize their patterning. The top panel illustrates the dorsal view (pink), while the bottom panel shows the coronal view. The legend identifies each compartment: epithelium (blue), CNCC-derived mesenchyme (purple), and myoblasts/myofibers (red). Intrinsic and extrinsic muscle types are indicated (arrows). CNCC, cranial neural crest cell.
Skeletal muscles share fundamental characteristics but also exhibit distinct structural and functional differences. All skeletal muscles are composed of striated muscle fibers and require neural input for contraction [24]. However, while both limb and tongue belong to a migratory lineage of somite [21, 25], their origin, function, organization, and innervation vary significantly. All limb muscles originate from the segmented paraxial mesoderm (somites) [26]. In contrast, tongue muscles have a mixed origin, deriving primarily from the occipital somites [27] with contributions from the cranial mesoderm [28] (Table 1).
| Muscle type | Origin | Function | Fiber composition | Tissue organization | Innervation |
| Limb | Somites 9–29 | Movement, joint stability | Mix of slow and fast twitch fibers | Distinct fascicles, defined tendons | Spinal cord motor neurons |
| Tongue | Occipital somites 1–5, cranial mesoderm | Mastication, speech articulation, swallowing | Predominantly fast twitch fibers, some slow twitch | Complex fascicles, extrinsic and intrinsic muscles, lack of defined tendons | Hypoglossal nerve and Vagus nerve (palatoglossus muscle only) |
Limb muscles support voluntary movements, postural stability, and locomotion,
while tongue muscles assist with mastication, speech articulation, and swallowing
(Table 1). Their differing functional requirements for contraction speed, energy
metabolism, and fatigue resistance determine their muscle fiber composition [29, 30]. Broadly, skeletal muscles contain one slow-twitch and three major
fast-twitch fiber types, with variations in their distribution based on
functional demands [30]. For example, the soleus is composed predominantly of a
slow-twitch fiber (~70%), while other leg muscles contain
significantly less (
Muscle organization also differs dramatically between limb and tongue. Limb muscle masses give rise to four major muscle types (flexors, extensors, adductors, abductors) and over 50 muscle fiber types that remain anatomically and functionally separate [34]. Limb muscles have distinct fascicles and defined tendons that attach to bone (Table 1), allowing for efficient force transmission. The tongue, however, incorporates a unique combination of intrinsic and extrinsic muscle fiber types [35] (Fig. 2). Its four intrinsic muscles (superior longitudinal, inferior longitudinal, vertical, and transverse muscle fibers) are entirely contained within the tongue and lack bony attachments. They interweave in complex, multidirectional layers to enable fine motor control for shaping, elongating, and thickening the tongue [33]. The extrinsic muscles (genioglossus, hyoglossus, styloglossus, and palatoglossus) originate from external bony structures and insert into the tongue to facilitate larger positional movements such as protrusion, retraction, and elevation [36]. Together, these two types of muscle fibers enable both the fine motor control and gross movement necessary for the tongue’s diverse functions [36].
Motor neurons originating from the spinal cord innervate limb muscles [37]. In contrast, tongue muscles are predominantly innervated by cranial motor neurons of the hypoglossal nerve [38] with additional vagus nerve innervation to the extrinsic palatoglossus muscle [36]. Thus, the neural control of limb is distinct from tongue.
The MRFs also regulate myogenesis differently in the tongue and limb. Myf5 and MyoD are activated simultaneously in the limb, whereas the tongue exhibits a sequential activation, with Myf5 preceding MyoD [39]. Further, myoblasts contributing to tongue formation predominantly express Myf5 rather than MyoD [40]. Myf5 loss can be compensated in both limb [41] and tongue [39]. However, while MyoD loss can be compensated in the limb [42], in the tongue it causes reduced muscle formation resulting in microglossia [39]. Additionally, myogenin-null embryos exhibit more severe defects in tongue muscle development compared to the limb [43]. These differences may reflect distinct myogenic timelines, as embryonic and fetal myogenesis are temporally separated in the limb but overlap in the tongue [44].
The distinct myogenic phases, embryonic and fetal, are differentially regulated in limb and tongue. For example, Wnt signaling in limb regulates fetal myogenesis but not embryonic myogenesis [45]. Further, Wnt signaling is required for limb muscle cell precursor number and myofiber quantity. In contrast, Wnt5a ligand is expressed in developing tongue muscle cells between E12.5–E14.5 and not apparent in mature myofibers after E15.5 [46]. Muscle-specific Wnt signaling regulates lingual myoblast fusion and differentiation at E14.5 [47], while epithelial specific Wnt signaling controls the number of muscle progenitor cells and their proliferation in tongue [48]. Further, it has been proposed that the tongue muscle progenitor proliferation regulated by Wnt signaling is through its upstream control of Notch signaling [48]. In limb, Notch signaling has established roles in maintaining muscle stem cells by inhibiting differentiation [6, 49]. Recently, Notch1 signaling has been shown to have roles in limb muscle fiber type composition and myofiber maturation [50].
In limb, BMP signaling plays a dual role: promoting embryonic myogenesis while
inhibiting muscle differentiation during fetal myogenesis [51]. While BMP is also
critical for craniofacial development [52], its specific role in regulating
tongue muscle formation has not yet been investigated. TGF-
The HH signaling pathway is a well-established regulator of embryonic tissue development [63]. It requires a membrane receptor Patched1 (PTCH1) that inhibits another membrane receptor, Smoothened (SMO), in the absence of HH ligand. There are three studied HH ligands: SHH, Indian HH (IHH), and Desert HH (DHH) [64]. Among these, SHH expression is extensively reported in the tongue, while both SHH and IHH have been implicated in limb muscle. Importantly, SHH is dual lipidated which limits its free diffusion [65] and thus require additional co-receptors, Growth Arrest Specific 1 (GAS1), Cell-adhesion molecule-related/downregulated by oncogenes (CDON) and brother of CDON (BOC), for ligand reception [66]. Specifically, the dual-lipidated SHH is recruited at the cell surface by the cell adhesion molecules CDON and BOC, which are members of the immunoglobulin superfamily. GAS1, a glycosylphosphatidylinositol (GPI)-anchored membrane protein, subsequently recognizes the ligand and removes its lipid modifications, facilitating SHH binding to PTCH1. When the ligand binds to PTCH1, the SMO inhibition is relieved to modulate downstream signaling. The final effectors of the HH pathway are the GLI transcription factors (GLI1, GLI2, and GLI3), which activate or inhibit target gene expression [67].
SHH decreases Pax3 and Pax7 expression and promotes the expression of other MRFs (Fig. 1) [68, 69]. Although SHH does not directly bind to the MRF, Myf5, its expression is dependent on HH signaling via GLI transcription factors, for both initial and continuous expression [70, 71, 72]. GLI1 and GLI2 can also interact with various regulatory elements of the MRF, MyoD [70, 73, 74]. Further, MyoD regulation can be independent of SHH [75], possibly due to cross-talk of GLI2 with other pathways [73]. The GLI regulation is also observed for myogenin [73, 76]. HH signaling also reportedly mediates expression of MyHC in mussel larval stages [77], in vitro muscle cells [78], and mouse embryonic cardiac tissue [79]. This highlights the versatility and importance of HH signaling in different myogenesis stages.
The expression patterns of HH signaling and its components differ significantly between the tongue and limbs (Fig. 3). In the limb, SHH ligand is expressed in the posterior limb bud mesenchyme, specifically within the zone of polarizing activity, (Fig. 3A, yellow) creating a gradient of GLI3 repressor [80, 81]. This asymmetric distribution of GLI isoforms ensures HH pathway activation in the posterior limb while maintaining pathway repression in the anterior limb. Expression of HH receptor SMO and target gene GLI1 indicates active HH signaling in muscle lineage cells (Pax3+, Myf5+) as well as in SHH surrounding lateral plate-derived mesenchymal cells [42].
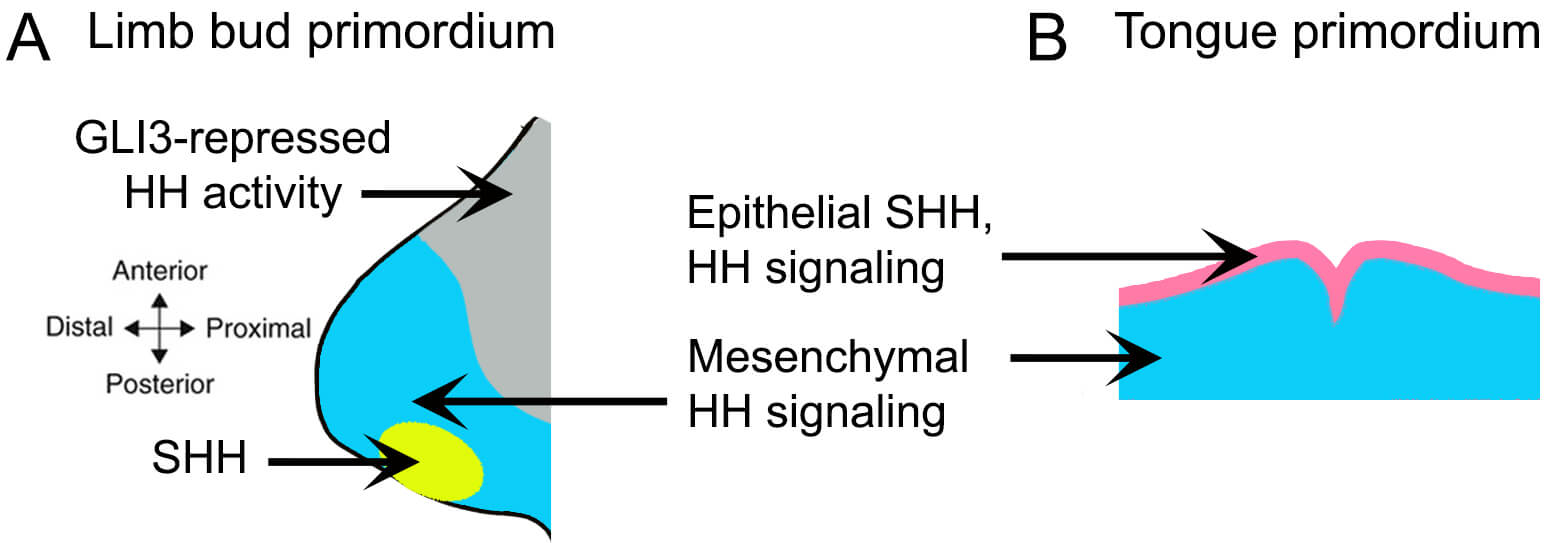 Fig. 3.
Fig. 3.
Distinct expression patterns of the SHH ligand and downstream HH signaling in the limb and tongue primordia. (A) In the limb bud, SHH is expressed in the zone of polarizing activity located in the posterior mesenchyme (yellow), with HH-responsive lateral plate-derived mesenchyme shown in blue. Regions with suppressed HH pathway activity, due to GLI3 repressor function, are indicated in grey. (B) In contrast, SHH expression in the tongue primordium is localized the epithelium (pink), while HH-responsive CNCC-derived mesenchyme is shown in blue.
Conversely, in the embryonic tongue, SHH is present in the entire epithelium of the mandibular arches and the developing tongue [82, 83, 84] (Fig. 3B). Its downstream signaling components, including the SMO receptor and GLI transcriptional activators, are expressed in both the epithelium and the CNCC-derived mesenchyme [83]. However, by E18.5, as the epithelium differentiates into taste and non-taste epithelium, SHH expression becomes restricted to taste cells, and HH signaling occurs only in cells of the taste epithelium [83]. Our recent studies confirm the absence of HH signaling, as indicated by Gli1 expression, in embryonic tongue muscles [85]. Notably, epithelial SHH in the tongue primarily signals to the CNCC-derived mesenchyme, which in turn supports tongue muscle development [23] — a notably distinctive mechanism from the mesoderm-directed HH signaling observed in limb muscle formation. Overall, the HH pathway in tongue development exhibits greater complexity and temporal regulation than in limb development.
Mouse, chick and zebrafish have conserved steps of skeletal muscle development parallel to humans [86, 87]. Studies using vertebrate models or in vitro mouse or chick cells have determined that HH signaling regulates distinct stages of both limb and tongue myogenesis (Table 2, Ref. [22, 23, 42, 73, 83, 85, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102]). Specifically, in limb, Shh ligand is responsible for muscle migration, proliferation, fusion, differentiation of slow fiber types, muscle size and muscle mass. In the tongue, limited studies indicate its critical role in tongue formation and initial myofiber arrangement. Similarly, the HH receptor Smo affects all stages of limb myogenesis; in the tongue, its primary function lies in myoblast migration-mediated tongue formation. Additionally, Smo overexpression can disorganize lingual muscle structure.
| Model system | Functions in limb muscle | Functions in tongue muscle |
| Shh/Ihh Ligand | ||
| Shh Deletion (Shh-/- mouse) | ||
| Shh overexpression | Zebrafish embryos [91, 92] | Mouse epithelium (K14+) [23] |
| Shh/Ihh reduction | Chick embryos (ED3) [93] | ShhMFCS4/− mouse [23] |
| In vitro supplementation | Chicken HH22 limb bud culture and mouse myoblast culture [90] | Research not available |
| Mouse E10.5 limb bud organ culture [89] | ||
| Mouse E13.5 limb myogenic cell culture [42] | ||
| E11 chick embryo myogenic cell culture [94] | ||
| Chicken HH17 limb bud culture [95] | ||
| Zebrafish embryos [91, 92] | ||
| Ihh ligand deletion (Ihh−/− mouse) | Research not available | |
| Smo receptor | ||
| Somite-specific deletion | Pax3𝐶𝑟𝑒; Smofl/fl mouse [42, 88] | Research not available |
| Mesenchyme-specific deletion | Prx1𝐶𝑟𝑒; Smofl/fl mouse (lateral plate-derived) [42] | Wnt1𝐶𝑟𝑒; Smofl/fl mouse (CNCC-derived) [23] |
| Muscle connective tissue-specific deletion | Tcf4GFPCre+neo; Smofl/fl mouse [42] | Research not available |
| Myoblast-specific deletion | Myf5𝐶𝑟𝑒; Smofl/fl and MyoD𝐶𝑟𝑒; Smofl/fl mice [42] | Hand2𝐶𝑟𝑒; Smofl/fl mouse [96] |
| Smo overexpression | Research not available | Hand2𝐶𝑟𝑒; R26SmoM2 mouse [22] |
| SAG treatment (E10.5 mouse) [97] | ||
| In vitro Smo inhibitor | Chick myogenic cells [42] | Tongue formation (microglossia) [83, 99] |
| Mouse embryonic P19 cells overexpressing Gli2 and fibroblast [73] | ||
| Zebrafish Smu (Smo) mutant [98] | ||
| Gas1/Cdon co-receptor | ||
| Gas1 modulation in vivo and in vitro | Gas1 transfection in mouse myoblast C2C12 cells and primary myoblasts derived from Cdon+/+ and Cdon–/– mice [100] | Gas1–/– mouse [85] |
| Gli transcription factors | ||
| Gli Deletion (Gli2–/–; Gli3+/– mouse, Gli2–/–; Gli3–/– mouse, Gli3–/–; Shh–/– mouse) | ||
| Mesenchyme-specific deletion | Research not available | Wnt1𝐶𝑟𝑒; Gli2fl/fl; Gli3fl/fl mice [87] |
| In vitro Gli1 supplementation | Research not available | |
*in the last 25 years; MPC, Myogenic progenitor cells; MyHC, Myosin Heavy Chain.
Another HH receptor, Gas1, also plays a vital role in skeletal myogenesis. Although in limb in vitro, it enhances myotube formation and myoblast differentiation, our recent studies in tongue with in vivo Gas1 deletion suggest it also has roles in muscle cell proliferation, differentiation and maturation during fetal myogenesis [85]. Intriguingly, these effects are not due to muscle-cell specific deletion. Global deletion, including additional epithelium and stromal Gas1 deletion, results in altered lingual intrinsic muscles that consequently affect tongue shape and size [85]. Further, Gas1 functions via alternate pathways to HH signaling. On the other hand, Gli transcription factors initiate MRF activation in limb (Fig. 1), while in tongue its deletion from mesenchyme results in aglossia [87]. Overall, numerous studies indicate essential roles of HH signaling in limb myogenesis. However, no similarly comprehensive body of research exists for tongue myogenesis (Table 2). Available studies indicate vital roles of HH signaling in tongue formation and initial myofiber patterning, but the regulation once the muscles are arranged remains understudied.
In limbs, SHH organizes myoblast distribution along multiple axes and is crucial for distal migration [88]. It directs the anteroposterior axis non-cell-autonomously through lateral plate-derived limb mesenchyme [42], while also acting cell-autonomously on a subset of limb myoblasts to regulate autopod muscle formation [88]. Loss of SHH delays MRF activation, disrupting muscle formation, but does not prevent the proximal migration of myoblasts into the limb bud [88].
Tongue myoblast precursors migrate as a cohesive strand along a distinct and complex pathway [103]. Myoblast determination in the tongue occurs in a posterior-to-anterior direction [23]. Interactions between CNCC- and mesoderm-derived cells contribute to this process through HH signaling [104]. However, reduced SHH expression or inhibition of SHH signaling in CNCC-derivatives does not appear to directly impact myoblast determination (Table 2). In contrast, epithelial SHH acts non-cell-autonomously through mandibular CNCC-derived mesenchyme to guide the unidirectional myoblast migration to the tongue primordium [21]. Deletion of Shh, membrane receptor Smo, or transcription factors Gli2 and Gli3 in CNCC-derivatives leads to aglossia due to impaired myoblast migration [23, 82, 84, 87, 96, 101, 102] (Fig. 4A). This demonstrates the necessity of non-cell-autonomous HH signaling for the migration of tongue myoblast precursors and subsequent muscle formation.
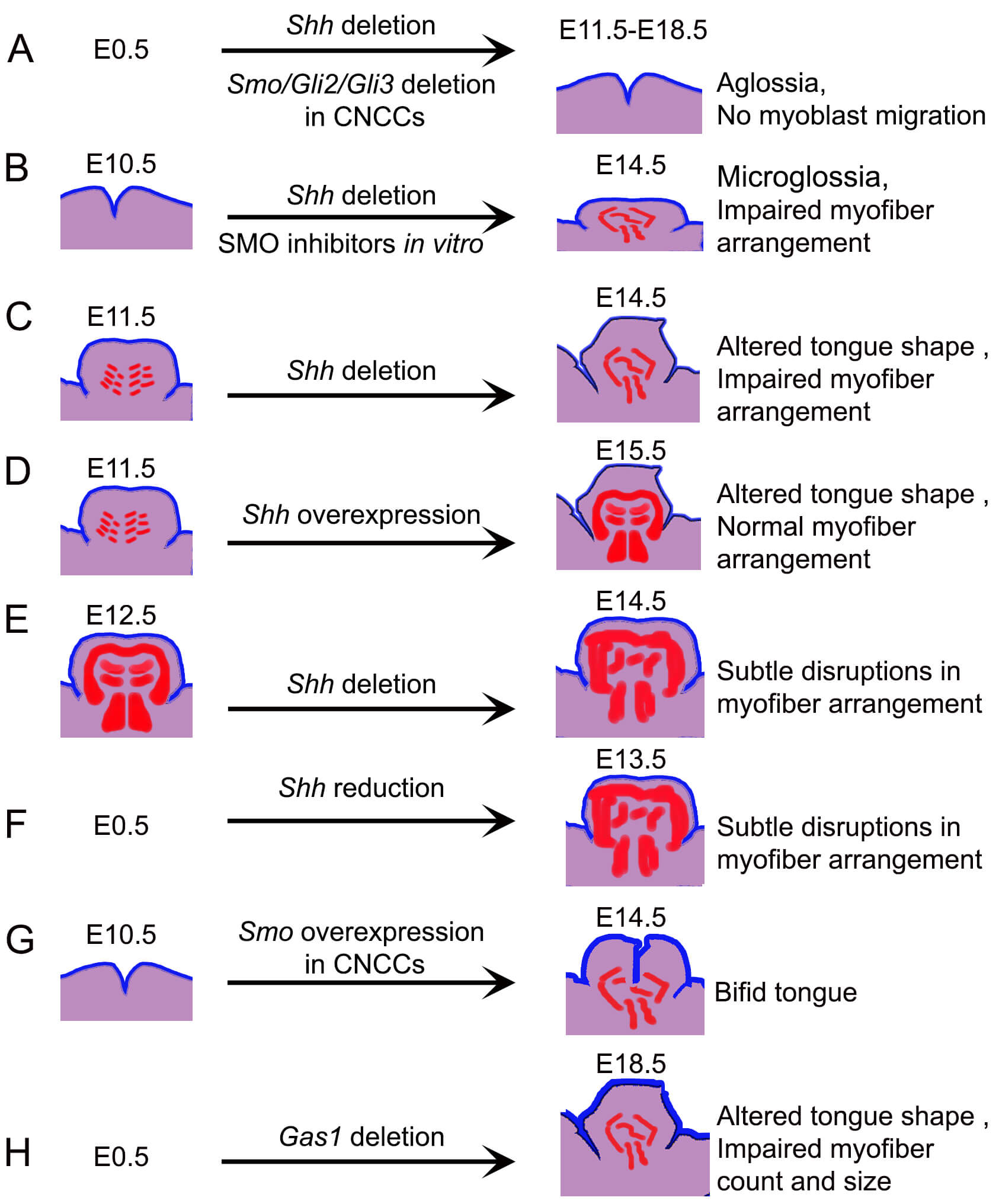 Fig. 4.
Fig. 4.
Graphic illustration of the impact of altering the HH pathway components on tongue morphology and muscle fiber organization. (A) Constitutive deletion of Shh, or CNCC-specific deletion of Smo, Gli2 or Gli3 results in aglossia, attributed to disrupted myoblast migration. (B) Conditional Shh deletion mouse model or in vitro SMO inhibition beginning at E10.5 results in reduced tongue size and loss of the characteristic striated architecture of intrinsic muscles by E14.5. (C) Conditional Shh deletion at E11.5 results in altered tongue shape and disorganized myotube patterning. (D) Ectopic overexpression of Shh in the K14+ epithelial domain perturbs overall tongue morphology but preserves intrinsic muscle organization. (E) Deletion of Shh after the initial establishment of muscle fibers causes only mild disorganization, indicating a temporally restricted requirement for SHH in muscle patterning. (F) Constitutive reduced expression of Shh does not disrupt muscle patterning. (G) Constitutive activation of SMO in CNCC derivatives induces bifid tongue formation. (H) Constitutive deletion of Gas1 reduces tongue size and alters shape due to impaired myofiber count and size. The initiation points of modulations are indicated on the left, whether constitutive (E0.5) or induced at specific developmental stages. The timing of tissue analysis is shown on the right. The colors represent epithelium (blue), CNCC-derived mesenchyme (purple), and myoblasts/myofibers (red).
Together, in the limb, SHH functions both cell-autonomously within myoblasts and non-cell-autonomously through limb mesenchyme to control myoblast distribution and distal migration. In contrast, in the tongue, SHH signaling operates solely in a non-cell-autonomous manner from the epithelium to CNCC-derived mesenchyme, orchestrating myoblast migration but not determination.
Determined myoblasts undergo extensive proliferation while maintaining an undifferentiated state, a process essential for increasing muscle mass [105]. HH-responding Gli1+ cells are present in limb muscle during and after myoblast migration, suggesting ongoing HH pathway activity in muscle progenitors [80, 94]. In the limb, SHH deletion or loss of cell-autonomous HH signaling does not impair the proliferation of embryonic myoblasts in mice [88]. Instead, non-myogenic tissues appear to influence myogenic proliferation and differentiation [106]. In the posterior hindlimb of late chick embryos, SHH indirectly supports myoblast proliferation by maintaining cells in an undifferentiated state, a stage equivalent to E14 in mice [95]. Additionally, ectopic SHH expression in chick muscle cells has been shown to induce myoblast proliferation in vitro [94], reinforcing a potential cell-intrinsic role for HH signaling in promoting limb myoblast proliferation under specific conditions.
Tongue mandibular explant cultures treated with SMO inhibitors exhibit
microglossia [83, 99] (Fig. 4B), primarily due to decreased proliferation of
CNCC-derived mesenchymal cells [99]. Interestingly, despite the reduction in
CNCC-derivatives proliferation upon HH pathway inhibition, the proliferation of
myoblasts and expression of key MRFs such as MyoD and Myf5
remain unaffected [99]. This might be due to that fact that CNCCs support the
proliferation of myogenic precursors through paracrine signaling, particularly
via TGF-
Some in vivo studies of the tongue show that reduction of Shh expression in the epithelium or decreased HH pathway activity in CNCCs, as indicated by reduced expression of target genes Gli1 and Ptch1, does not disrupt either CNCC-derived mesenchyme or myogenic cells proliferation [23, 101, 104]. These results contrast with studies in which CNCC-specific Smo haploinsufficiency resulted in diminished mesenchymal proliferation [84] and that reduction of SHH in distal ectoderm reduces myoblast proliferation at E11.5 [82]. Such discrepancies may reflect potential context-dependent effects or differences in experimental design, timing, or degree of pathway suppression. Our recent research indicates HH-independent roles of receptor Gas1 in regulating Pax7+ myoblast numbers and muscle cell proliferation at E18.5, potentially as a compensatory response to reduced numbers of mature fibers and an increased demand for new fiber formation [85].
Overall, studies on limb and tongue myogenesis suggest that HH signaling may not be not essential for embryonic myoblast proliferation. However, the findings in limb muscle showing that ectopic SHH can induce proliferation underscore its context-dependent role. In contrast, during tongue development, alternative pathways or compensatory mechanisms may regulate myogenic proliferation independently of HH input.
Limb muscles contain variable distributions of both slow and fast fibers (Table 1). HH signaling promotes slow muscle fiber differentiation by inducing slow MyHC expression in a larger precursor pool of myoblast to suppress fast myofiber formation [90, 109, 110]. Overexpression of SHH leads to an increased proportion of slow fibers, whereas continuous deletion disrupts overall muscle formation by E16.5 [89]. Fast fiber formation in the absence of HH signaling in zebrafish studies further support this finding [91, 92]. IHH deletion also results in defective muscle differentiation [93]. Notably, SHH expression ceases in the limb at E16.5 in mice but Ptch1 remains in fetal myoblasts, indicating a possible transition to the IHH signaling seen in the developing bone anlagen [93]. This shift from SHH to IHH highlights the dynamic regulation of HH signaling in limb muscle development and the necessity of maintaining proper signaling levels for muscle fiber differentiation.
Fast-twitch fibers predominate in tongue muscles, with a smaller population of
slow-twitch fibers also present (Table 1). Notably, slow fibers are especially
enriched in the posterior region of the extrinsic genioglossus muscle [32, 111].
If Shh is deleted after myoblast migration, muscle fiber differentiation
into intrinsic or extrinsic types are unimpaired [41] (Fig. 4C). This indicates less dependence on HH signaling in tongue muscle fiber differentiation
than for limb muscle differentiation. On the other hand, alternative pathways,
including the TGF
The slow and fast fiber specification is responsive to innervation [114, 115], which itself can be shaped by CNCC-derived mesenchyme [116, 117]. We hypothesize that HH signaling might regulate fiber specification indirectly through its role in CNCC rather than directly on myogenic cells [118]. In summary, no studies to date have definitively shown that HH signaling directly determines slow vs. fast fiber fate in lingual myogenesis.
Given the necessity of HH signaling in slow fiber-type specification in limb muscles and the relative scarcity of slow fibers in the tongue, the direct impact of HH signaling on fast fiber differentiation in tongue remains unclear and warrants further investigation.
HH signaling plays a multifaceted role in dictating the spatiotemporal organization of limb musculature. It has been shown that non-myogenic tissues, particularly muscle mesenchyme, contribute significantly to muscle patterning [119, 120]. Conditional Smo deletion in such non-muscle limb mesenchyme resulting in a significant loss of antero-posterior muscle patterning while the failure of cell-autonomous removal of Smo activity in the somites themselves to affect this patterning reinforces its non-cell-autonomous role [42]. Myoblasts have receptors for mesenchymal extracellular matrix (ECM) components comprising heparan sulphate proteoglycans [121, 122], which can bind with SHH and mediate distinct patterning [123]. Separately, ectopic expression of Shh in the anterior compartment of the chick wing leads to dramatic muscle re-patterning, with anterior muscles adopting posterior-like characteristics in a mirror-image arrangement [124].
In the mouse tongue, SHH is not present in muscle cells or ECM of embryonic tongue [83, 85]. However, it can still control intrinsic muscle patterning via its interaction with ptch1+ or Smo+ CNCC-derivatives [23]. SHH mediation in the development of lingual tendons also aids indirectly by providing a scaffold for the normal patterning of intrinsic muscle fibers. In mice with Shh deletion from E10.5, the normal striated architecture of the intrinsic muscles was lost and tongue size reduced at E14.5 (Fig. 4B). Once lingual muscles are migrated, Ptch1/HH signaling inhibition in CNCCs specifically leads to hypoglossia, tongue clefting, and disorganized myotube arrangement at E14.5 (Fig. 4C). However, overexpression of Shh in K14+ epithelium at this stage distorts tongue architecture but retains normal arrangement of the intrinsic muscles (Fig. 4D). Further, deletion of Shh after the initial arrangement of muscle fibers results in subtle organizational disruptions (Fig. 4E), suggesting temporal SHH loss at later stages produces milder effects. Notably, a reduction, but not complete elimination, of SHH does not alter lingual muscle patterning [23, 125] (Fig. 4F). The presence of multiple SHH sources in the tongue [9, 126, 127] may account for the graded severity of tongue defects. In contrast, Smo overexpression in CNCC-derivatives resulted in a disorganized muscular structure [22] (Fig. 4F) while pharmacological SMO overactivation caused the formation of bifid tongue [97] (Fig. 4G).
Additionally, deletion of HH co-receptors Gas1 and Boc affects pharyngeal tongue formation at E15.5 [128]. Anterior tongue formation occurred but muscle patterning was not investigated. Expression of HH signaling target gene and activator Gli1 and HH co-receptors Gas1, Cdon, and Boc has previously been observed in embryonic tongue at E11.5 [129]. We recently identified Gas1, both gene and protein, as well as CDON and BOC in the embryonic lingual muscles at E18.5 [85]. Intriguingly, constitutive deletion of Gas1 alters tongue morphology and size as well as the number and size of myofibers at E18.5 (Fig. 4H). HH-responding Gli1+ cells are notably absent in lingual muscles at this stage [83, 85], and their expression in the epithelium and mesenchyme remains unchanged following Gas1 deletion. These findings suggest that the role of HH signaling in muscle organization during tongue development may involve non-canonical or indirect mechanisms.
Overall, an intricate interplay between SHH and the ECM is fundamental to limb muscle patterning. In contrast, in the tongue, SHH acts through CNCC-derived mesenchyme and tendon scaffolding, rather than the ECM, in a temporally regulated manner.
A hallmark of muscle cell maturational progression is the sequential expression of distinct MyHC isoforms. The MyHC isoform expression, including embryonic and neonatal, is tightly regulated during muscle embryonic maturation [130, 131]. Studies on myogenesis regulation have primarily focused on the differentiation of MyHC into slow and fast fibers. However, no literature is currently available for studies specifically focusing on the regulation of neonatal and embryonic MyHC isoforms by HH signaling in either tongue or limb muscles. Our recent studies show that Gas1 deletion significantly reduced MyHC-embryonic expression, but not MyHC-neonatal, without disrupting HH signaling [85].
The development of the NMJ is also a critical step in muscle maturation, enabling effective communication between motor neurons and muscle fibers [132]. SHH is identified as a key regulator in motor neuron development [133, 134]. HH signaling is also essential for the development and patterning of the central nervous system and neural tube, influencing the fate and proliferation of neural progenitors [135, 136]. However, its direct involvement in NMJ formation or maturation in limb or tongue muscles during embryonic development has not been established. We observed a significant reduction in NMJ area following embryonic deletion of Gas1, as well as a percentage overlap between nerve terminals and muscle endplates [85]. Whether these NMJ structural changes in Gas1 mutants translate into muscle functional deficits remains unknown.
Taken together, the role of HH signaling in regulating muscle maturation in both the tongue and limb warrants further investigation. Our recent findings suggest that HH co-receptor Gas1 supports muscle maturation through non-canonical HH signaling.
Overall, HH signaling is essential for embryonic myogenesis in both limb and lingual skeletal muscles. Numerous studies have demonstrated that the loss of key HH pathway components, such as Shh or Smo, leads to significant muscle dysmorphogenesis, though the nature and severity of these defects differ between the limb and tongue (Table 2, Fig. 4). These findings underscore the distinct and context-dependent roles of HH signaling in limb versus tongue muscle development. However, important gaps remain in our understanding of HH signaling in the lingual muscle, particularly during the stages of myoblast proliferation, differentiation, and fiber maturation. Further studies are needed to elucidate HH-mediated fetal myogenesis during tongue embryonic development. A detailed understanding of the pathway regulation would advance our fundamental knowledge of tongue muscle growth and maturation as well as potentially offer new therapeutic avenues for muscle regeneration and repair in congenital or acquired disorders.
AK designed the review. AK and APM performed the literature survey. AK prepared the visualizations. APM prepared the tables. AK drafted the review. Both authors reviewed and edited the final version and contributed to editorial changes in the manuscript. Both authors read and approved the final manuscript. Both authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
We gratefully acknowledge Ms. Lisa M. Price, MSLIS, Research Support Librarian, Rowan-Virtua School of Osteopathic Medicine for English editing of the manuscript.
This work was supported by the National Institutes of Health National Institute on Deafness and Other Communication Disorders grant ECR R21 1R21DC017799 (AK). Further support was provided by the Rowan-Virtua School of Osteopathic Medicine Seed Fund (AK) and Summer Medical Research Fellowship award (APM).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.