




 , Aleksey Tyulenev 1, Tatyana Kalashnikova 1, Lyubov Sutormina 1, Vadim Ushakov 1, Oleg Oktyabrsky 1
, Aleksey Tyulenev 1, Tatyana Kalashnikova 1, Lyubov Sutormina 1, Vadim Ushakov 1, Oleg Oktyabrsky 11 Institute of Ecology and Genetics of Microorganisms, Perm Federal Research Center, Russian Academy of Sciences, 614081 Perm, Russia
Abstract
Endogenous and exogenous H2S can influence the virulence of bacteria and their susceptibility to antibiotics and oxidative stress. Escherichia coli and Bacillus subtilis, when grown in minimal medium with sulfate as the sole sulfur source, produce H2S when treated with cystine or under stress conditions, including exposure to chloramphenicol and ciprofloxacin. However, it is unknown whether Mycobacterium smegmatis is capable of producing sulfide under these conditions and how this production affects cell physiology.
Real-time monitoring of dissolved oxygen (dO2), pH, extracellular K+, and sulfide was performed directly in culture flasks using selective electrodes. Changes in the level of low molecular weight (LMW) thiols were recorded using spectrophotometric methods and high performance liquid chromatography (HPLC).
Sudden addition of cystine or chloramphenicol to growing M. smegmatis cultures increased the intracellular level of cysteine and induced its homeostasis mechanisms, which include the export of excess cysteine from cells and its incorporation into mycothiol (MSH), along with desulfurization with H2S formation. Ciprofloxacin also increased intracellular cysteine concentration and sulfide production but did not induce cysteine release. Both antibiotics inhibited growth and respiration, whereas cystine transiently increased respiration and glucose uptake in M. smegmatis, in contrast to E. coli, which showed a transient inhibition of these processes.
The mechanisms of cysteine homeostasis under the action of antibiotics in M. smegmatis are similar to those in E. coli and B. subtilis, indicating the universal nature of stress response. The opposing effects of cystine-derived H2S on physiological parameters in M. smegmatis and E. coli may be important factors contributing to their susceptibility to antibiotics.
Keywords
- chloramphenicol
- ciprofloxacin
- H2S
- cysteine
- mycothiol
- Mycobacterium smegmatis
The rapid development of antibiotic resistance of pathogenic bacteria requires searching for adjuvants of existing drugs to improve their therapeutic efficacy and targets to create novel antibiotics. Research into the influence of cysteine and its derivatives on the virulence of bacteria and their susceptibility to antibiotics and oxidative stress is one of the actively developing areas [1]. Endogenous H2S, formed during L-cysteine degradation, reduced the sensitivity of bacteria to antibiotics and oxidative stress, while mutations and inhibitors of H2S-producing enzymes increased the efficacy of antimicrobial drugs [2, 3, 4]. High intracellular cysteine levels are potentially harmful, since cysteine is capable of autoxidation yielding ROS and can potentiate the Fenton reaction giving rise to toxic hydroxyl radicals by maintaining the Fe2+ pool [5, 6]. To explain the interplay between L-cysteine metabolism, H2S production and oxidative stress, a model has been proposed where 3-mercaptopyruvate sulfotransferase (3MST) protects Escherichia coli against oxidative stress by catalyzing cysteine degradation to form H2S, which binds free iron required for the Fenton reaction [3]. However, instead of enhancing antibiotic tolerance, exogenous and endogenous H2S in Mycobacterium tuberculosis and exogenous H2S in Acinetobacter baumannii sensitized them to antibiotics by triggering a pro-oxidative redox imbalance [7, 8, 9]. Endogenous H2S in mycobacteria has been shown to be an effector molecule that maintains bioenergetic homeostasis by stimulating respiration, plays a key role in central metabolism, regulates redox homeostasis, and increases the susceptibility of M. tuberculosis to antituberculosis drugs clofazimine and rifampicin through its pro-oxidant function [9]. Host-derived H2S has also been shown to stimulate M. tuberculosis respiration, primarily through cytochrome bd oxidase, and regulate genes involved in sulfur and copper metabolism, as well as the Dos regulon [8, 10]. The effect of H2S on antibiotic sensitivity apparently depends on the bacterial species and can be determined by the features of endogenous H2S production or the nature of its effect on metabolic pathways. We have previously shown that antibiotic exposure of E. coli and Bacillus subtilis is accompanied by changes in low molecular weight thiol levels and H2S production [11, 12, 13, 14]. It still remains unknown whether mycobacteria are capable of producing H2S in response to antibiotics.
Addition of cystine to E. coli growing in minimal medium with sulfate has been shown to cause cytoplasmic cysteine overload and stimulate cysteine export, its incorporation into glutathione (GSH) and degradation to form H2S [6, 13, 15, 16, 17]. Multiple bacteria possess cysteine-inducible cysteine-specific efflux pumps for its export [18, 19, 20]. However, no such transporters have been detected in M. tuberculosis and the ability of mycobacteria to export cysteine into the medium remains unknown, although recently the involvement of the Rv0191 efflux pump in the export of low mlecular weight (LMW) thiols has been suggested [21]. L-cysteine supplementation has been shown to stimulate H2S production in Bacillus anthracis, Pseudomonas aeruginosa, and Staphylococcus aureus [2, 3]. Various E. coli enzymes are known to possess in vitro cysteine desulfhydrase activity (TnaA, CysK, CysM, MalY, MetC, 3MST, IscS, CyuA); some of them may specifically cope with cysteine toxicity [2, 22, 23, 24, 25]. Addition of cysteine to M. tuberculosis was also found to increase H2S production in a cysteine desulfhydrase (Cds1)-dependent manner, suggesting that this pathway could be used to get rid of excess cysteine [9].
Recently, in a series of studies, we have shown that a transient increase in intracellular cysteine concentration and activation of cysteine homeostasis mechanisms occur not only when cysteine/cystine is added to the growth medium, but also as a consequence of abrupt protein synthesis inhibition during various stresses in E. coli and B. subtilis [11, 12, 26, 27]. These stresses involve nutrient starvation (depletion of glucose, phosphate, or nitrogen) and exposure to antibiotics such as chloramphenicol, tetracycline, and ciprofloxacin. Excess cysteine formed in E. coli under these conditions was mainly incorporated into glutathione, and was also exported into the medium and partially degraded to form H2S. Sulfide release from cells into the medium under stress can be recorded using selective sulfide or platinum electrodes as abrupt changes in the redox potential (Eh jumps). In our earlier studies, Eh jumps suppressed by the thiol reagent N-ethylmaleimide (NEM) occurred upon exposure to various stressors (starvation, elevated temperature, ultraviolet irradiation, treatment with acetate and antibiotics, etc.) on Gram-negative and Gram-positive bacteria, indicating the universal nature of the observed phenomenon [28].
Like other mycobacteria, Mycobacterium smegmatis contains no glutathione and uses mycothiol as its major LMW thiol, whose concentration in cells is comparable to that of glutathione in E. coli [29, 30, 31]. Mycothiol in mycobacteria performs many of the functions of glutathione in Gram-negative bacteria, including the maintenance of the intracellular redox balance, as well as protection against reactive oxygen and nitrogen species, alkylating agents, and antibiotics. Mycothiol (MSH) levels may be influenced by environmental conditions, such as oxidative or osmotic stress or nutrient starvation [30]. In response to the increased need for antioxidant molecules under hostile conditions such as oxidative stress, M. tuberculosis actively upregulates CysM-dependent L-cysteine biosynthesis through the transcription factors AosR and SigH, turn enhancing mycothiol and ergothioneine production [21]. The expression of MSH biosynthetic genes was also elevated by acidic conditions [32]. It was shown that MSH turnover occurs in M. smegmatis and that MSH can be a cysteine source [30]. Since MSH is autoxidized much more slowly than cysteine, it has been suggested that cysteine incorporation into MSH is a way to conserve cysteine and limit its intracellular concentration. However, no literature data on how the emergence of excess cysteine affects MSH levels are available.
In this study, we examined how cystine supplementation and treatment with chloramphenicol or ciprofloxacin affect changes in cysteine, mycothiol, and H2S levels in M. smegmatis. We also monitored the effects of these treatments on bacterial growth and respiration, which may be important for explaining the effects of H2S on antibiotic tolerance. M. smegmatis is frequently used as a model organism to study M. tuberculosis due to its genetic similarity, conserved metabolic pathways (including biosynthesis and turnover of LMW thiols), and lower pathogenicity. However, despite the similarity and widespread use of M. smegmatis to investigate the mechanisms of drug resistance in tuberculosis, these bacteria have some differences in gene expression, metabolic pathways, and cell structure. In particular, M. tuberculosis was shown to produce significantly more H2S compared to M. smegmatis [9]. Moreover, these bacteria differ significantly in growth rate, which may play an important role in altering thiol levels during antibiotic-mediated growth inhibition. This suggests that some caution is needed when generalizing the results obtained in M. smegmatis to M. tuberculosis.
The American Type Culture Collection (ATCC) strain M. smegmatis 700084/mc2155 was used in this study. Bacterial cultures were grown in 250 mL flasks at 37 °C with shaking at 150 rpm in 100 mL of minimal M9 medium with glucose (4 g/L) [33]. This medium additionally contained glycerol (4 g/L), Tween 80 (0.15%), FeCl3 (10 µM), CaCl2 (0.2 mM), MgSO4 (2 mM) and 1 mL of trace elements prepared as described previously [34]. Bacteria were pre-grown for 24 hours in the presence of ampicillin (25 µg/mL); part of the culture was then transferred to fresh medium of the same composition. The overnight culture was centrifuged, diluted in 100 mL of fresh medium without antibiotic to OD600 0.25 and cultured as described above. Some experiments on studying H2S production were carried out in the glycerol-free medium. In this case, the overnight culture after centrifugation was transferred to a medium of the above composition, excluding glycerol. Cystine (Cys, 30 µM), chloramphenicol (Cam, 25 µg/mL) and ciprofloxacin (CF, 10 and 30 µg/mL) were added when OD600 had reached 0.4. The antibiotic concentrations used were selected based on preliminary experiments on their ability to rapidly inhibit bacterial growth and respiration when added to growing cultures. Ciprofloxacin concentrations (10 and 30 µg/mL) were 40–120 times higher than the MIC values (0.25 µg/mL) for M. smegmatis mc2155 reported in the literature [35].
The following equation was used to calculate the specific growth rate
(µ): µ =
A Clark InPro 6800 oxygen electrode (Mettler Toledo, Greifensee, Switzerland) and an ESC-10601/7 pH electrode (IT Company, Moscow, Russia) placed in flasks with M. smegmatis cultures were used for continuous measurement of dissolved oxygen and pH in the medium, respectively. Data recorded using the dO2/pH controller of a BioFlo 110 fermenter (New Brunswick Scientific Co., Edison, NJ, USA).
The sulfide content in the culture medium was monitored using an XC-S2--001 sulfide-specific chalcogenide electrode (Sensor Systems Company, St. Petersburg, Russia) and a cpX-2 computer pH/ion meter (IBI, Pushchino, Russia). The advantages of this electrode are the ability to measure at physiological pH (the electrode operates in the pH range from 6 to 12), no reaction to changes in oxygen content and high sensitivity (sensitivity threshold 5 nM). A standard curve for determining sulfide concentration was generated using known amounts of Na2S.
Changes in K+ concentration in the medium were recorded using an ELIS-121K K+-selective electrode (IT Company, Moscow, Russia) and a cpX-2 computer pH/ion meter (IBI, Pushchino, Russia). When measuring potassium, its content in the growth medium was reduced to 0.2 mM. Synchronous processing of all data from the sensor system was carried out using the RS-232 and Modbus protocols and the Advantech OPC Server v3.0 software package (Advantech Co., Shing-Tien, New Taipei City, Taiwan, China).
Gaseous H2S was determined based on its specific reaction with lead acetate [Pb(Ac)2]. The lead sulfide formed during the reaction produces a brown spot on Pb(Ac)2-soaked paper strips fixed above the surface of the liquid culture [2]. Sequential replacement of paper strips every 30 min allowed monitoring of the H2S production kinetics. The color intensity was quantified using ImageJ1.54g software (NIH, Bethesda, MD, USA) after scanning the spots. The detection threshold of this method was 0.1 µM.
L-cysteine was determined using our modification of the Gaitonde method [26]. Briefly, culture samples collected at different time points were concentrated ten-fold and sonicated in 0.1 M Tris-HCl (pH 8.6). After protein removal with 0.5 M perchloric acid followed by treatment with potassium hydroxide to pH 8.6, the resulting supernatant was evaporated on an RV10 rotary evaporator (IKA, Staufen, Germany) and treated with dithiothreitol (10 mM) to reduce cystine to cysteine. To determine extracellular cysteine, culture samples were passed through a membrane filter to remove cells; the resulting filtrates were concentrated by rotary evaporation, the protein was then removed and cystine was reduced to cysteine as described above. Cysteine assay in reduced samples was performed as described previously [26]. Standard curves were prepared using known amounts of cysteine treated in the same way as the culture samples.
The intracellular LMW thiols, including mycothiol, were determined using the
high performance liquid chromatography (HPLC) method with fluorometric detection
following the reaction of thiols with the fluorescent dye monobromobimane (mBBr)
[36]. M. smegmatis culture samples (15 mL) were centrifuged (5 min at
8000
All the experiments were performed at least in triplicate, with results
presented as means
Compared to M. tuberculosis, a slow-growing virulent strain (doubling
time, 24 h), M. smegmatis is a fast-growing non-virulent strain
(doubling time 90 min) [37]. Under our experimental conditions, M.
smegmatis grew at an average rate of 0.25
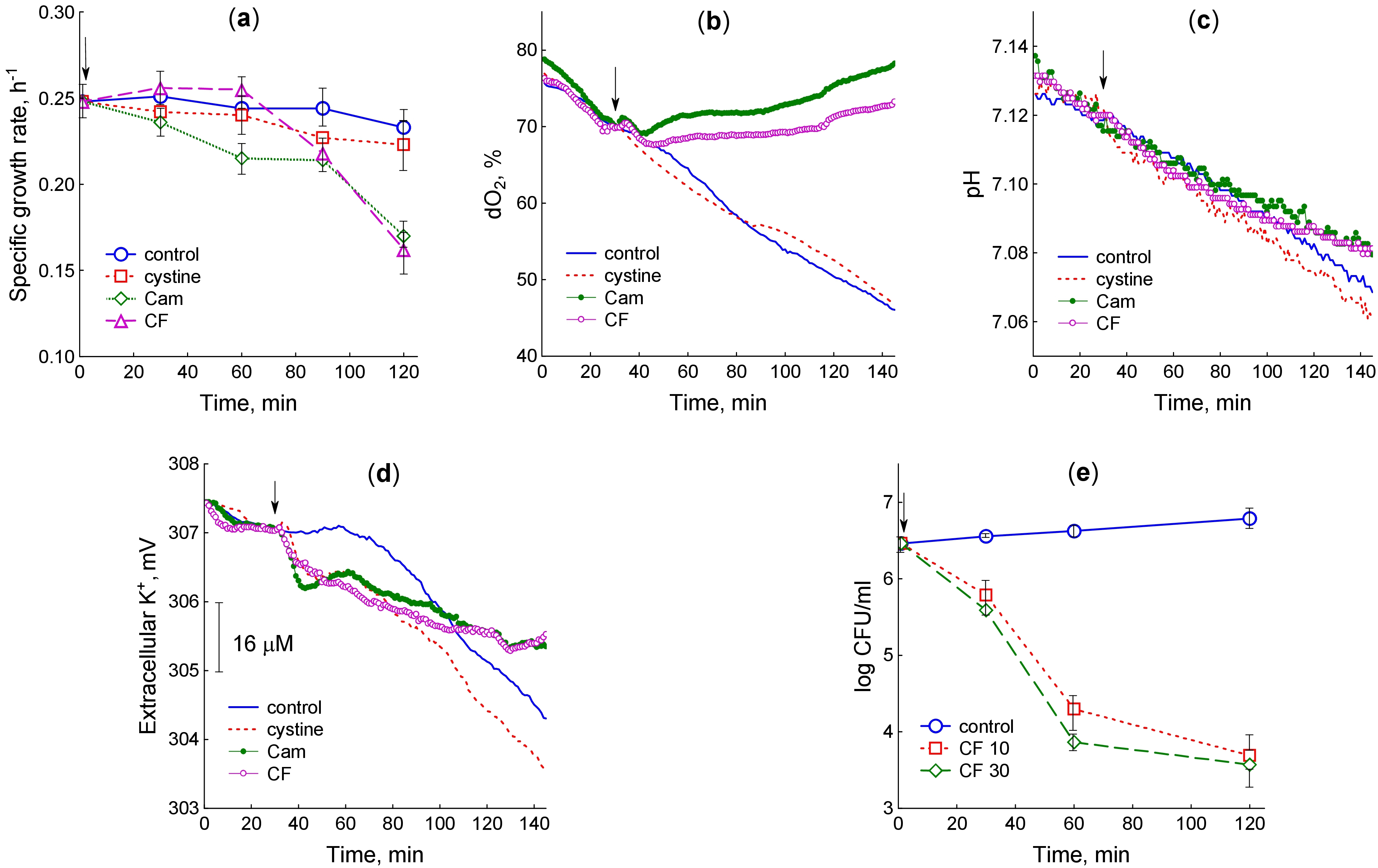 Fig. 1.
Fig. 1.
Changes in physiological parameters of M. smegmatis upon treatment with cysteine, chloramphenicol and ciprofloxacin. (a) Specific growth rate. (b) Concentration of dissolved oxygen. (c) pH of the medium. (d) Extracellular K+ concentration. (e) Ciprofloxacin-induced killing curves. The arrow shows the moment of adding the test compounds. (a,e) The average of the data from several experiments is plotted with the standard error indicated (bars). (b–d) The graphs show typical curves obtained from three independent experiments. Cys, Cystine; Cam, chloramphenicol; CF, ciprofloxacin; dO2, dissolved oxygen.
Due to the rapid oxygen consumption by cells, the content of dissolved oxygen in
control cultures gradually decreased with increasing culture density, despite
constant rotation of the flasks (Fig. 1b). The rate of oxygen consumption
expressed as a percentage of dO2 per OD600 per minute was 0.36
Glucose metabolism during M. smegmatis growth was accompanied by
accumulation of acidic by-products, which reduces pH (Fig. 1c). In this regard,
sensitive pH recording can be used as a real-time indicator of glucose
consumption. Two hours after the test compounds had been added, the change in pH
relative to the baseline (
During bacterial growth, the K+ content in the medium decreased due to its
uptake by the cells (Fig. 1d). Treatment of M. smegmatis with cystine
and antibiotics caused an abrupt decline in K+ concentration in the medium,
indicating an accelerated influx of K+ ions into the cells. The change in
the K+ sensor potential relative to the initial value 10 min after treatment
with cystine and antibiotics (
We also tested the bactericidal effect of ciprofloxacin on M. smegmatis (Fig. 1e). The maximum decrease in CFU was observed during the first hour of exposure to the antibiotic, when the bacteria maintained a high growth rate. It indicates that, like in the case of E. coli, growing bacteria with high metabolic activity suffer more damage, preventing colony formation during subsequent cultivation on antibiotic-free plates [38]. There was no significant difference in the effects of 10 and 30 µg/mL ciprofloxacin.
Cysteine or cystine can be transported into M. smegmatis and M. tuberculosis [39], which may lead to intracellular cysteine overload and activate desulfurization of excess cysteine to form H2S, as observed in E. coli [13, 16]. Indeed, genetic and biochemical evidence has been provided that conversion of cysteine to H2S mediated by Cds1 desulfhydrase functions as a sink for excess cysteine in M. tuberculosis, but H2S production in M. smegmatis was barely detectable [9]. Using a highly sensitive method for recording sulfide with a selective electrode, we studied the changes in H2S production upon exposure of M. smegmatis to cystine and antibiotics.
Under our experimental conditions, the addition of 30 µM cystine caused an
abrupt drop of 52.6
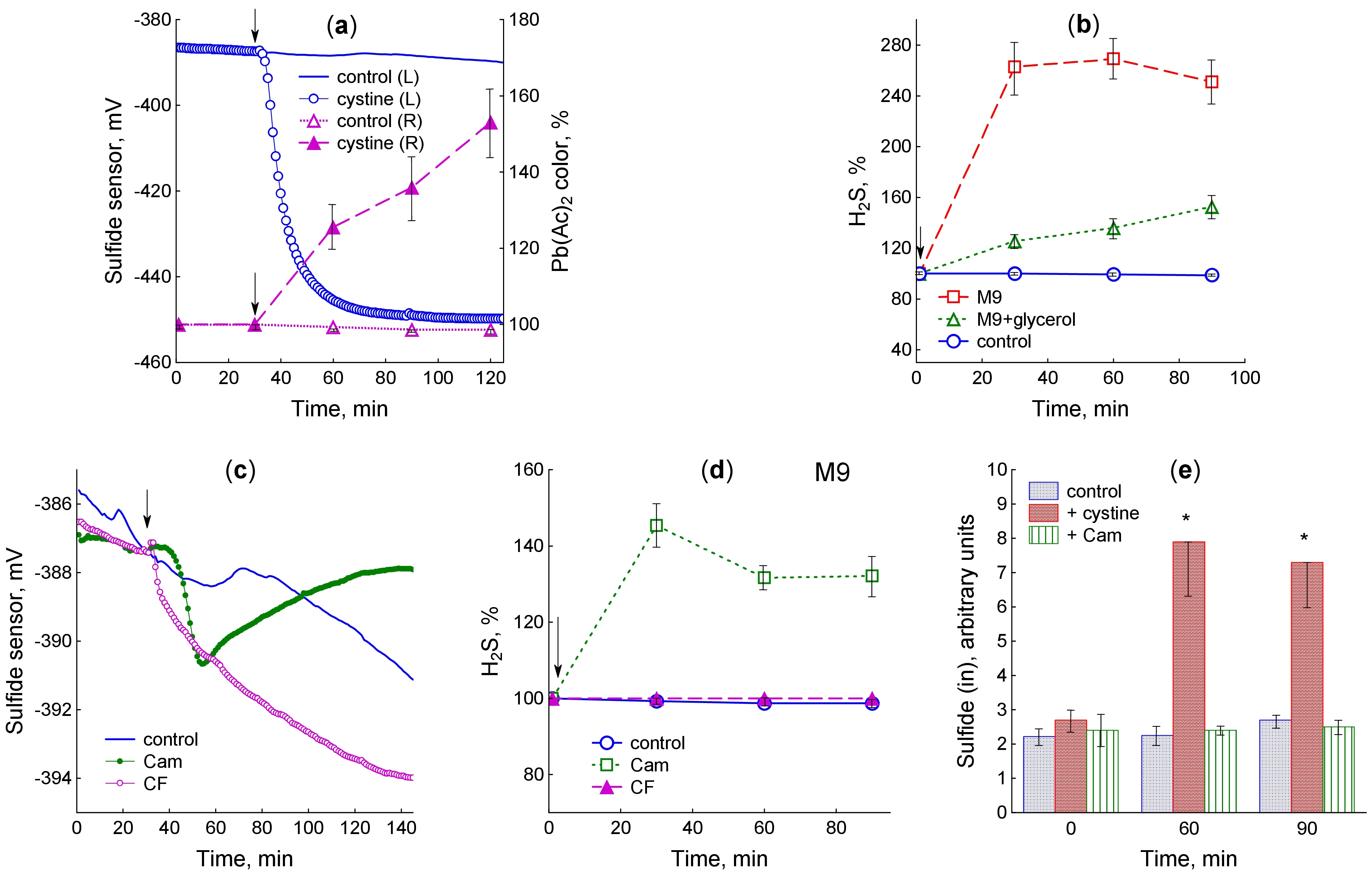 Fig. 2.
Fig. 2.
M. smegmatis releases H2S upon addition of cystine
and antibiotics. (a) H2S production after cystine addition to
glycerol-containing medium: left panel (L) – sulfide sensor readings; right
panel (R) – color change of Pb(Ac)2-soaked paper strips upon their
successive replacement every 30 min. (b) H2S accumulation kinetics
determined using the Pb(Ac)2-soaked paper strips after cystine addition to
the medium with and without glycerol. (c) Sulfide sensor readings after addition
of chloramphenicol and ciprofloxacin to the glycerol-containing medium. (d) The
kinetics of H2S accumulation determined using the Pb(Ac)2-soaked paper
strips after addition of chloramphenicol and ciprofloxacin to the glycerol-free
medium. (e) The H2S levels in extracts of cells grown in glycerol-containing
medium determined by high performance liquid chromatography (HPLC). Results of HPLC analysis are expressed as arbitrary
units (peak area divided by OD600). The arrow shows the moment of adding the
test compounds at OD600 0.4. Mean values and standard errors (vertical
bars) are shown. Significant differences compared to control are marked with an
asterisk (*) (p
Chloramphenicol caused the sulfide electrode potential to drop by 4.2
HPLC analysis of monobromobimane derivatives revealed the presence of sulfide in the cell extract (Supplementary Fig. 2). After 60-min exposure, cells treated with cystine contained 4 times more sulfide than controls (Fig. 2e). Treatment with chloramphenicol did not affect the level of intracellular sulfide (Fig. 2e).
The methods used allow the assessment of H2S accumulation in the liquid medium (sulfide electrode), in the gas phase (lead acetate-soaked paper strips), and in cell extracts (HPLC analysis of mBBr derivatives) and differ in sample preparation and detection threshold (5 nM, 0.1 µM and 2 nM, respectively), which may explain some of the discrepancies in the data obtained by these methods, especially at low levels of H2S production. In general, the use of three different detection methods revealed H2S production by M. smegmatis cells upon addition of cystine to the medium. H2S release from cells upon exposure to chloramphenicol and ciprofloxacin was only confidently recorded using a selective sulfide electrode, indicating that the level of H2S production is weak under these conditions. Nevertheless, in all the cases, sulfide release could be caused by an excess cysteine in the cytoplasm, prompting us to measure its levels in the studied situations.
In the absence of exposures, intracellular cysteine in M. smegmatis was
maintained at a nearly constant level (0.115
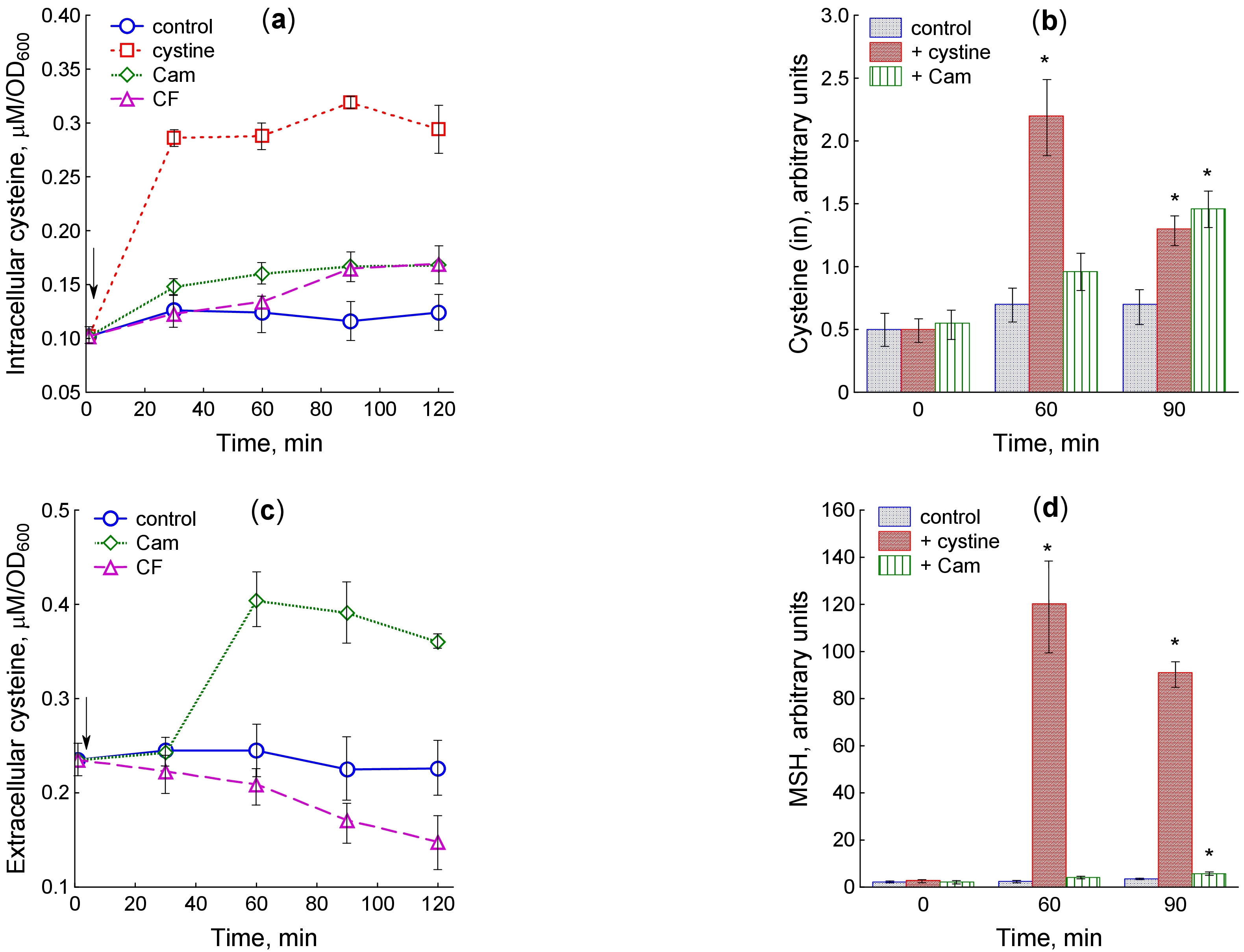 Fig. 3.
Fig. 3.
Changes in cysteine and mycothiol levels upon treatment of
M. smegmatis with cystine and antibiotics. (a) Intracellular cysteine
determined using the Gaitonde’s method, (b) intracellular cysteine determined by
HPLC, (c) extracellular cysteine determined using the Gaitonde’s method, (d)
mycothiol (MSH) levels determined by HPLC. The arrow shows the moment of adding the
test compounds at OD600 0.4. Results of HPLC analysis are presented in
arbitrary units (the peak area divided by OD600). Significant differences
compared to control are marked with an asterisk (*) (p
Since excessive cysteine is cytotoxic, multiple bacteria utilize cysteine-specific efflux pumps for its export [18, 19, 20]. We measured the concentration of extracellular cysteine during exposure to antibiotics in order to elucidate the involvement of cysteine export in regulation of its intracellular homeostasis under stress. Extracellular cysteine concentration per OD600 unit was 2-fold higher than its intracellular level, remaining approximately constant during growth in the untreated culture. Chloramphenicol accelerated cysteine efflux and increased its extracellular concentration by 1.8 times 60 min after the start of exposure (Fig. 3c). In contrast to chloramphenicol, ciprofloxacin gradually reduced the extracellular cysteine levels.
In addition to cysteine and sulfide, HPLC analysis of mBBr derivatives revealed
the presence of mycothiol (Supplementary Fig. 2). Intracellular
mycothiol levels unchanged during growth of M. smegmatis in the control
culture, but increased significantly after cystine supplementation (Fig. 3d;
Supplementary Fig. 2). When exposed to chloramphenicol, the mycothiol
level statistically significantly increased after 90-min incubation with the
antibiotic, coinciding in time with the maximum increase in cysteine content
(Fig. 3d; Supplementary Fig. 2). Statistical analysis of the data
presented in Fig. 2b,d revealed a high correlation between the cysteine and
mycothiol levels when M. smegmatis was treated with cystine (r = 0.95,
p
In this work, we have shown that, just like in E. coli and B. subtilis, the main cause of endogenous H2S production in M. smegmatis is an increase in intracellular cysteine concentration, which may result from over import of exogenous cysteine/cystine or from dramatic inhibition of protein synthesis by chloramphenicol treatment. In both situations, the increase in intracellular cysteine levels induced mechanisms of its homeostasis. In E. coli, when protein synthesis is inhibited by antibiotics or as a result of nutrient starvation, most of the excess cysteine is incorporated into glutathione, while the remaining portion is exported into the medium and degraded to form H2S [11, 12, 26]. All these mechanisms of cysteine homeostasis are maximally activated during intracellular cysteine overload as a result of excess cystine import when it is suddenly added to a medium with sulfate as the only source of sulfur [6, 13, 16, 17]. During E. coli growth in cysteine-containing Luria-Bertani (LB) medium (Miller), H2S production by cells is carried out without any external influences. Under these conditions, the onset of H2S release coincides with the slowdown of growth and respiration, which may also indicate the occurrence of excess cysteine at this stage of culture growth [27, 38].
In B. subtilis, which does not contain GSH, as well as in the glutathione-deprived gshA mutant of E. coli, the main pathways to restore cysteine homeostasis are its export and degradation to form H2S, which is more intense and prolonged than in the wt strain of E. coli [12, 13, 14, 26]. It appears that bacillithiol, the major LMW thiol in B. subtilis as well as other Firmicutes [42], does not function as a cysteine buffer, unlike GSH in E. coli.
Mycothiol is the unique protective thiol of Actinobacteria [30] and the most abundant LMW thiol in M. smegmatis [29]. In this work, we have shown that an increase in intracellular cysteine is accompanied by an increase in mycothiol levels, indicating its role as a cysteine buffer in M. smegmatis. We also observed cysteine release from cells and H2S production upon exposure to chloramphenicol, indicating that M. smegmatis possesses all the mechanisms of cysteine homeostasis characteristic of E. coli: incorporation of excess cysteine into buffer molecules, release into the medium, and desulfurization.
Measurements showed that M. smegmatis growing under normal conditions
contained a low cysteine level (0.115
E. coli can utilize cysteine-specific efflux pumps (EamA, EamB, Bcr) to
export cysteine into the medium [18, 19, 20]. Thus, constitutive expression of the
cysteine exporter EamA in E. coli has been shown to prevent H2S
formation by reducing the intracellular concentration of cysteine [26, 44].
However, no such transporters have been identified in M. tuberculosis.
Nevertheless, we observed cysteine release from M. smegmatis cells
and its accumulation in the medium upon exposure to chloramphenicol, which may
indicate that an unidentified cysteine export system is present in this
bacterium. In E. coli, AlaE was found to be the primary exporter of
excessive intracellular cysteine, although this protein was previously identified
as an alanine exporter [17]. In contrast to E. coli, where ciprofloxacin
(10 µg/mL) caused a 3-fold accumulation of cysteine in the medium
compared to the control [45], M. smegmatis did not increase
extracellular cysteine levels under these conditions. Chloramphenicol-induced
cysteine release in M. smegmatis was also 2.6 times lower than that in
E. coli. This may be explained by the slower inhibition of growth and
hence protein synthesis in M. smegmatis compared to E. coli,
where the higher initial growth rate (0.68
M. tuberculosis encodes multiple enzymes that may produce H2S. Cysteine desulfurization involving Cds1 has been shown to be the major, although not the only, source of endogenous H2S in M. tuberculosis [9]. Cds1 is also present in M. smegmatis [9], but the involvement of this or other enzymes in H2S generation upon addition of cystine and chloramphenicol needs to be further studied. M. smegmatis has been reported to produce little H2S [9]. However, our experiments showed that sudden addition of cystine to M. smegmatis growing in sulfate medium causes the release of 0.3 µM sulfide, which is 4.5 times lower than that of E. coli (1.4 µM) [13], but is well detectable by the methods we used. The difference in H2S production may be due to variations in the culture medium composition. We found that excluding glycerol from the medium intensified H2S generation when cells were treated with cystine and chloramphenicol, which may be due to changes in the cell wall composition and permeability [40, 41]. The amount of sulfide released by M. smegmatis under the action of chloramphenicol is significantly lower than that of E. coli (10–15 nM and 180 nM, respectively). The reason, as in the case of cysteine export, may be a smaller excess of intracellular cysteine that occurs in M. smegmatis during rapid inhibition of protein synthesis.
Antibiotic-induced H2S formation appears to result from disruption of cysteine homeostasis due to protein synthesis inhibition. Under these conditions, growth inhibition in M. smegmatis was accompanied by a decrease in the rate of respiration and glucose consumption, as observed previously in E. coli [11, 12, 13]. However, although both bacteria released H2S when cystine was added, the physiological response was opposite. Reversible inhibition of growth and respiration was observed in the case of E. coli [13], whereas temporary stimulation of respiration and activation of glucose consumption occurred in M. smegmatis. These findings can be attributed to the peculiarities of the effect of H2S on cytochrome oxidases: in E. coli, low micromolar concentrations of H2S inhibit cytochrome bo oxidase [16, 47], while in M. tuberculosis, H2S stimulates respiration via cytochrome bc1/aa3 and, primarily, via cytochrome bd [9]. Another evidence of the participation of H2S in regulation of energy processes, including the membrane potential, in M. smegmatis may be the abrupt acceleration of K+ entry into cells, whose kinetics coincide with sulfide production upon addition of cystine and antibiotics. The ability of H2S to interact with K+ ion channels has been previously shown [48]. The special role of exogenous and endogenous H2S in regulating central metabolism and accelerating respiration and growth in mycobacteria may be among the reasons for the opposite effect of H2S on antibiotic susceptibility in M. tuberculosis and E. coli [2, 8, 9].
We have previously shown that the absence of one of the mechanisms of cysteine homeostasis leads to increased activation of the remaining mechanisms when excess cysteine appears in the cytoplasm. In particular, the lack of glutathione in the gshA mutant of E. coli causes more intense production of H2S and cysteine export when cells are treated with ciprofloxacin, whereas the mstA mutant, which does not produce H2S under these conditions, synthesizes more glutathione than the wt strain does [13]. The gshA mutant exhibited an increased sensitivity to ciprofloxacin in minimal M9 medium, especially with fractional addition of cystine during cultivation. In the present work, we showed that with increasing concentration of intracellular cysteine in M. smegmatis, the level of mycothiol rises, similar to that of GSH in E. coli. We speculate that, similar to E. coli, a decrease in MSH levels due to mutations or inhibitors of the enzymes involved in its synthesis will intensify H2S production under stress conditions. Since exogenous and endogenous H2S have been shown to enhance the killing of M. tuberculosis by the anti-TB antibiotics clofazimine and rifampin [8, 9], it is conceivable that inhibition of MSH synthesis may increase the susceptibility of mycobacteria to antibiotics via the same mechanism. Mycothiol is essential for M. tuberculosis survival and intracellular levels of this thiol are associated with changes in resistance to antibiotics and oxidative stress [39]. M. smegmatis devoid of MSH exhibited an increased susceptibility to several antibiotics, such as erythromycin, azithromycin, rifamycin S, penicillin G, and vancomycin [30, 49], suggesting that MSH is involved in multiple detoxification mechanisms, one of which may be the maintenance of cysteine homeostasis. Although caution is needed when extrapolating the results obtained with M. smegmatis to M. tuberculosis and further more in-depth studies are needed, our findings may be promising in searching for ways to improve the efficacy of anti-TB drugs.
In this study, we showed that, similar to E. coli and B. subtilis, addition of cystine and chloramphenicol to growing M. smegmatis induces intracellular cysteine accumulation and activates homeostatic mechanisms such as cysteine export, its degradation to H2S, and incorporation into mycothiol as a cysteine buffer. Ciprofloxacin also increased intracellular cysteine concentration and sulfide production but did not induce cysteine release. Our findings indicate that activation of cysteine homeostasis mechanisms may be part of a universal stress response across bacterial species. The molecular mechanisms underlying these processes in mycobacteria require further investigation, including key gene knockout experiments.
The datasets used or analyzed during the current study are available from the corresponding author on reasonable request.
Conceptualization, GS and OO; Investigation, AT, LS, TK, and VU; Formal analysis, OO, GS, AT, LS, TK, and VU; Methodology, OO, GS, AT, LS, TK, and VU; Data curation, GS and OO; Writing–original draft preparation, GS; Writing–review and editing, OO and GS. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
This study does not involve human or animal subjects; therefore, according to Russian regulations and policies, ethical committee approval is not necessary.
In this study, we used the equipment of Centre of collective usage of scientific instruments of Perm Federal Research Centre.
This work was supported by assignments # 124020500028-4.
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBL44441.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.