










 , Angela Trovato Salinaro 1
, Angela Trovato Salinaro 11 Department of Biomedical and Biotechnological Sciences, School of Medicine, University of Catania, 95123 Catania, Italy
2 Occupational Medicine, Department of Clinical and Experimental Medicine, University of Catania, 95123 Catania, Italy
3 Department of Agriculture, Food and Environment (Di3A), University of Catania, 95123 Catania, Italy
4 Department of Medicine and Surgery, Kore University of Enna, 94100 Enna, Italy
†These authors contributed equally.
Abstract
The global increase in diabetes mellitus has been accompanied by a significant rise in related complications. Diabetic patients frequently experience ocular surface disorders and multiple studies have demonstrated that the diabetic corneal epithelium is characterized by increased cellular fragility and compromised barrier integrity. It has been demonstrated that the processes of oxidative stress and inflammation are pivotal in causing ocular tissue damage in diabetic patients. Numerous studies have explored the protective effects of various antioxidants, especially those sourced from plants. Cynara cardunculus L. var. altilis (DC.), a species widely integrated into the Mediterranean diet and commonly known as cultivated cardoon (CC), is particularly rich in bioactive phenolic compounds, recognized for their antioxidant effects.
The current work focuses on assessing the effect of CC leaf extracts on high glucose-treated human corneal epithelial cells (HCEpiCs). HCEpiCs were cultured for 24 h in a medium supplemented with glucose up to a concentration of 25 mM. Mannitol treatment was included to distinguish whether the observed effects were due to glucose metabolism or solely osmotic stress. To evaluate the effect of CC extracts, corneal cells were pre-incubated with the CC extract 10–20 μg/mL for 24 h before high glucose (HG) treatment. Cell viability, transepithelial electrical resistance, wound healing assay and reactive oxygen species (ROS) measurements were performed after HG treatment. To evaluate the levels of oxidative stress, the antioxidant enzymes superoxide dismutase (SOD) and catalase (CAT), along with glutathione (GSH) levels were assayed. The mitogen-activated protein kinase ERK1/2/cytosolic phospholipases A2 (cPLA2)/cyclooxygenase-2 (COX-2) signaling pathway, triggering the inflammatory process, was evaluated by western blot analyses.
Our findings indicate that CC extract (i) improved viability, reducing oxidative stress by lowering ROS production and enhancing SOD, CAT activities and GSH content in human corneal epithelial cells exposed to high glucose concentrations; (ii) maintained a high TEER and promoted corneal epithelial wound healing; and (iii) induced down-regulation of the ERK 1/2/cPLA2/COX-2 signaling pathway involved in the inflammatory process and ROS production.
CC leaf extract could improve human corneal epithelial cell function suggesting its potential as a preventive agent against the development of chronic diabetic complications.
Graphical Abstract
Keywords
- catalase
- cyclooxygenase 2
- Mediterranean diet
- oxidative stress
- signal transduction
- corneal epithelial cells
- diabetes complications
- inflammation
- Cynara extracts
- cytosolic phospholipases A2
Diabetes mellitus (DM) is one of the major global health challenges today, due to the rapid increase in its incidence. This disease affects individuals of all ages, social conditions and demographic groups, and is spreading virtually everywhere in the world. DM is currently considered a disease with a large number of hidden cases, as those diagnosed represent only a small part of the real prevalence [1]. This chronic metabolic disorder develops when the pancreas fails to produce adequate amounts of insulin or when body tissues cannot effectively respond to the hormone’s action, leading to disturbances in glucose regulation. When diabetes is not kept under control, excessive glucose in the blood (hyperglycemia) occurs, which, over time, can seriously affect several body systems, especially blood vessels and the nervous system [2].
Approximately 50% of people with diabetes develop a form of corneal degeneration known as diabetic keratopathy [3]. The epithelial surface of the cornea is organized into 5–7 strata of non-keratinized squamous cells arranged in a stratified pattern which are essential for maintaining the transparency and structural integrity of the cornea. Due to the avascular nature of the cornea, the supply of nutrients and glucose to the corneal epithelial cells occurs primarily by diffusion from the aqueous humor rather than from the tear film [4]. Because the aqueous humor contains high glucose (HG) concentrations under hyperglycemic conditions, corneal epithelial cells in patients with diabetes mellitus are subject to prolonged contact with glucose, which contributes to a spectrum of pathological epithelial changes [5]. Disturbance of the corneal homeostatic balance has been widely observed in patients with diabetes. This condition is associated with a malfunction of the epithelial barrier, changes in the structure of epithelial cells and reduced corneal sensitivity [6]. Even small changes in the diabetic cornea can impair healing processes, leading to prolonged recovery times or non-healing epithelial lesions. The delayed healing process exposes patients to serious visual complications, including gradual epithelial thinning, the onset of secondary damage, and the triggering of cytokines that facilitate fibrotic scar development [5].
Corneal epithelial cells form the outermost layer of the ocular surface, protecting the eye from microbes, allergens, and chemicals [7]. The prompt renewal of the epithelial layer, along with the restoration of its protective barrier, is crucial for maintaining corneal architecture and vision-related function [8]. It has been demonstrated that exposure to HG levels leads to dysfunction of the corneal epithelial barrier, leading to a notable reduction in transepithelial electrical resistance (TEER). Therefore, gaining insight into the pathophysiological processes affecting the cornea in patients with diabetes is crucial for maintaining its structural and functional integrity [9, 10].
Moreover, increased oxidative stress resulting from elevated reactive oxygen
species (ROS) production has been identified as a key factor in the development
of diabetic complications [10]. The cornea, due to its location on the external
part of the eye, is constantly exposed to high concentrations of oxygen and solar
radiation, both sources of oxidative stress. To counteract this problem, the
cornea contains various enzymatic antioxidant defenses, including superoxide
dismutase (SOD) and catalase (CAT), and non-enzymatic ones, including glutathione
(GSH). SOD catalyzes the conversion (dismutation) of the superoxide anion
(O2
Studies have linked changes in ROS production to diabetic corneal problems, suggesting that HG and ROS production leads to systemic corneal disease, characterized by peripheral neuropathy, delayed corneal wound healing, and impaired cell migration [12, 13]. Although oxidative stress has been widely studied, the specific mechanisms by which hyperglycemia modulates reactive oxygen species generation and interferes with the repair of corneal epithelial barrier function are not yet fully elucidated.
In previous studies we demonstrated, in HG-treated human retinal microvascular endothelial cells, an activation of cytosolic phospholipases A2 (cPLA2) and of mitogen-activated protein kinase (MAPK) ERK1/2 [14] and an increase of cyclooxygenase-2 (COX-2) protein levels and release of inflammatory cytokines [15], highlighting that the activation of the ERK/cPLA2/COX-2 axis represents an early event after exposure to HG, mediating retinal damage. Cytosolic PLA2 catalyzes the hydrolysis of membrane glycerophospholipids, primarily phosphatidylcholine, at the sn-2 position, releasing arachidonic acid (AA), which is a precursor for eicosanoids such as prostaglandins E2 (PGE2) and leukotrienes (LTs). Cytosolic PLA2 activation is triggered by various pro-inflammatory signals, initiating a Ca2+-dependent translocation from the cytosolic space toward the perinuclear membranes. This process involves phosphorylation of Ser-505 in the catalytic domain by MAPK ERK1/2 and phosphorylation of Ser-727 by other MAPKs [16]. To date, few studies have elucidated the role of the signaling pathway in diabetic corneal diseases.
Cynara cardunculus L. var. altilis (DC.) is a perennial plant native to the Mediterranean region, popularly known as cultivated cardoon and closely related to the more widespread globe artichoke [C. cardunculus L. var. scolymus (L.) Fiori] and the wild cardoon [C. cardunculus L. var. sylvestris (Lamk) Fiori], the progenitor of both cultivated species. Traditionally grown for its edible, fleshy leaf midribs and for the use of its flowers as a source of vegetable rennet [17], the cultivated cardoon has recently attracted interest for its potential in the development of natural extracts for pharmacological applications [18]. The primary reason for this interest is its considerable abundance of bioactive molecules, including phenolic acids, flavonoids, and sesquiterpene lactones [19, 20], which have been shown to possess antioxidant, anti-inflammatory, and hepatoprotective properties [21]. Among the three C. cardunculus taxa, cultivated cardoon is considered the most promising for the extraction of natural bioactive compounds, as it may produce more biomass than wild cardoon and globe artichoke [22].
It has been demonstrated that globe artichoke leaf extract can lower sorbitol levels in rat eye lenses exposed to high glucose conditions. Sorbitol buildup is a hallmark of diabetic retinopathy, and reducing its accumulation could offer protective benefits against this eye condition. Experimental evidence from both ex vivo and in vivo models demonstrates that artichoke leaf extract plays a protective role by decreasing oxidative stress and suppressing inflammatory processes, while simultaneously enhancing the efficiency of the endogenous antioxidant defense system [23, 24, 25]. The cultivated cardoon, a versatile crop in the Mediterranean diet, is particularly rich in bioactive phenolic compounds, recognized for their antioxidant effects, as well as dietary fiber and essential minerals [26]. Polyphenols possess strong antioxidant properties that enable them to neutralize reactive oxygen species, thereby limiting oxidative stress. This protective effect can reduce the risk of oxidative damage to ocular tissues, which is a key factor in the development of age-related eye diseases such as cataracts and macular degeneration [23]. Clearly, these multifaceted actions cannot be performed by a single compound, but rather by a complex mixture of active ingredients, mainly mono- and dicaffeoylquinic acids and flavonoids such as luteolin, which exert additive or synergistic pharmacological effects.
In the present study, we investigated the early cellular and molecular responses of HG-treated human corneal epithelial cells to cultivated cardoon (CC) leaf extract. Our findings indicate that CC extract improved viability, reduced ROS production, increased antioxidant enzyme activities and migration and reduced the inflammatory process after glucose treatment, suggesting its potential as a preventive agent against the development of chronic diabetic complications. The results, obtained after 24- and 48-hour treatments, are preliminary to understanding CC’s potential in diabetic keratopathy.
All reagents were from Sigma (St. Louis, MO, USA) or Merck (Darmstadt, Germany) or VWR (Leighton Buzzard, UK). Standards were purchased from Extrasynthese (Lyon, France) or Roth (Karlsruhe, Germany).
The cultivated cardoon ‘Altilis 41’, developed by researchers at Catania
University as part of a breeding program on C. cardunculus L., was
selected as the target genotype due to its high production of biomass and
bioactive compounds [26, 27]. Seeds were placed in plastic pots (diameter: 10 cm;
height: 8 cm) filled with a 1:1 (v/v) mixture of peat-based substrate
(Profi-Substrate, Gramoflor, Germany) and sand. The pots were placed in an
unheated greenhouse and irrigated, when necessary, with deionized water (EC
Per each replicate, a lyophilized sample (100
Caffeoylquinic acids and flavone profiles were analyzed as already described
[29] with some modifications, using a Zorbax Eclipse XDB-C18 column (4.6
Human corneal epithelial cells (HCEpiCs, Innoprot, P10871, Derio, Spain) were grown in Corneal Epithelial Cell Complete Medium (CECM, Innoprot, P60131). Cells were mycoplasma-free (Innoprot, REF: P10871-IM). To validate the authenticity of the cell lines, Innoprot employs Genetic Identity Verification through Short Tandem Repeat (STR) profiling, which is a standard method for human cells.
For high glucose treatments, cells were cultured for 24 h in their respective growth media supplemented with glucose to a concentration of 25 mM. Mannitol treatment was included to distinguish whether the observed effects were due to glucose metabolism or solely osmotic stress. Cells were incubated, for the same time period of 24 h, to the respective media supplemented with 25 mM mannitol.
CC leaf powders, obtained by freeze-drying the plant material, were dissolved in dimethyl sulfoxide (DMSO). The stock solution 0.1 mM, contained a mixture of chemicals extracted from the CC leaves. To evaluate the effect of CC leaf extracts, corneal cells were pre-incubated with the CC extract for 24 h before HG treatment. During the incubation of these cell cultures for the following 24 h, in the presence or absence of 25 mM glucose, the cultures were replaced with artichoke extract in fresh medium. Schematically, experiments were performed on five groups of corneal cultures, incubated with CECM containing 5.5 mM glucose (NG), as follows:
(1) normal glucose group (NG): 24 h incubation with CECM + 24 h incubation with fresh CECM;
(2) high glucose group (HG): 24 h incubation with CECM + 24 h incubation with fresh CECM containing glucose to a concentration of 25 mM;
(3) mannitol group (M): 24 h incubation with CECM + 24 h incubation with fresh CECM containing mannitol to a concentration of 25 mM;
(4) cultivated cardoon group (CC): 24 h incubation with CECM containing CC leaf extract + 24 h incubation with fresh CECM containing CC extract;
(5) cultivated cardoon + high glucose group (CC + HG): 24 h incubation with CECM containing CC extract + 24 h incubation with fresh CECM containing CC leaf extract and HG.
Cell viability was determined by MTT assay (Chemicon, Temecula, CA, USA) as previously described [30].
HCEpiCs (7
The effect of CC leaf extract on cell migration after treatment with HG was evaluated by wound healing assay, as previously described [13].
Reactive oxygen species (ROS) were detected using Cellular Reactive Oxygen Species Detection Assay Kit (ab113851, Abcam, Cambridge, UK) as previously described [32].
After different treatments, the activities of antioxidant enzymes Superoxide dismutase (SOD), catalase (CAT) and the levels of glutathione (GSH) were assayed using commercial kits (SOD Activity CS0009, CAT Activity MAK531 and GSH Assay MAK517, Sigma-Aldrich, Burlington, MA, USA), according to the manufacturer’s instructions.
Following treatments, HCEpiCs were lysed and Bradford assay was employed to
quantify total protein content. A total of 40 µg of protein from
each sample was separated by SDS-PAGE and transferred to membranes according to
established protocols [15]. Primary antibodies against cPLA2 (1:1000,
sc-376636, Santa Cruz, CA, USA) and phospho-cPLA2 (1:1000, 2831, Cell
Signaling Technology, Danvers, MA, USA), as well as ERK1/2 (1:1000, 4696, Cell
Signaling Technology, Danvers, MA, USA) and phospho-ERK1/2 (1:1000, 4370, Cell
Signaling Technology, Danvers, MA, USA),
Following treatments, supernatants of cell cultures were used to determine PGE2 (Cayman Chemical, Ann Arbor, MI, USA) and interleukin-6 (IL-6) levels (R&D Systems Inc., Minneapolis, MN, USA), according to the manufacturer’s instructions. Detection range was 7.8–1000 pg/mL for PGE2 and 3.1–300 pg/mL for IL-6.
Experiments were conducted in triplicate, each comprising four samples per group
(n = 4). Values are from mean
HPLC-MS was employed to both identify and quantify ten distinct bioactive compounds present in the CC leaf extract, providing a detailed phytochemical profile, belonging to the groups of caffeoylquinic acids and flavones. The latter represented approximately 87% of the total quantified polyphenols, consistent with previous findings in similar species [33]. Among the flavones, luteolin and its derivatives were the most abundant, with luteolin and luteolin-7-O-glucoside reaching values of 3.97 and 2.68 g kg-1 of dry matter, respectively (Fig. 1), in agreement with the findings of Sałata et al. [19]. Apigenin-7-O-glucoside and 1,5-O-dicaffeoylquinic acid were the predominant compounds within the apigenin and caffeoylquinic acid groups, with amounts of 1.39 and 0.94 g kg-1 of dry matter, respectively, as also observed in other phenolic-rich leaf extracts [26, 34].
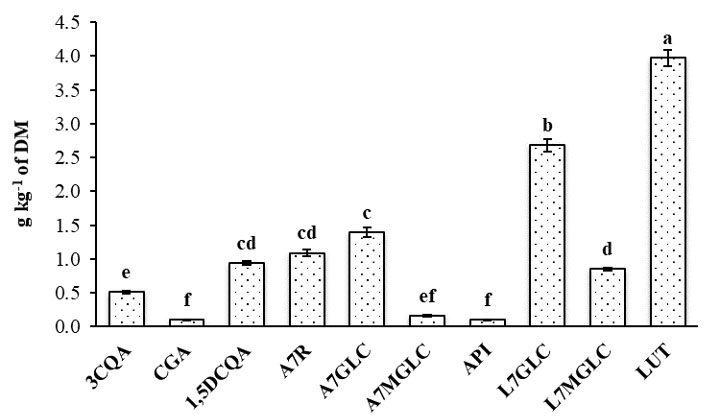 Fig. 1.
Fig. 1.
Content (
MTT-based cytotoxicity tests were performed to determine the effects of CC leaf extract on cell viability following treatment of HCEpiCs with increasing CC leaf extract concentrations (2.5–5.0–10–15–20 µg/mL). As shown in Fig. 2A, all concentrations caused no effect on cell viability after incubation for 24 and 48 h. In another set of experiments, HCEpiCs were cultured in a medium supplemented with 25 mM glucose (HG) for 24 h and 48 h in the absence and the presence of increasing concentrations of CC leaf extract (2.5–5.0–10–15–20 µg/mL), as schematically described in the Materials and Methods section. Mannitol treatment was included to distinguish whether the observed effects were due to glucose metabolism or solely osmotic stress. The treatment with HG for 24 and 48 h induced a decrease in cell viability by about 31% and 81%, respectively, in comparison with cells incubated in NG medium. Preincubation of the cells for 24 h with increasing concentrations of CC leaf extract 10–15–20 µg/mL and incubation for another 24 h with fresh medium containing CC leaf extracts in the presence of HG significantly increased cell viability by about 16%, 49% and 50%, respectively, when evaluated against the corresponding values observed in HG-treated cells (Fig. 2B). After 48 h in the presence of HG, 10–15–20 µg/mL of CC leaf extract induced an increase of cell viability by about 2.4, 2.7 and 2.9 fold, respectively, in comparison with cells treated with HG. The presence of lower concentrations of CC leaf extract (2.5 and 5 µg/mL) did not significantly alter the viability compared with cells treated with HG. Treatment with mannitol 25 mM did not alter cell viability.
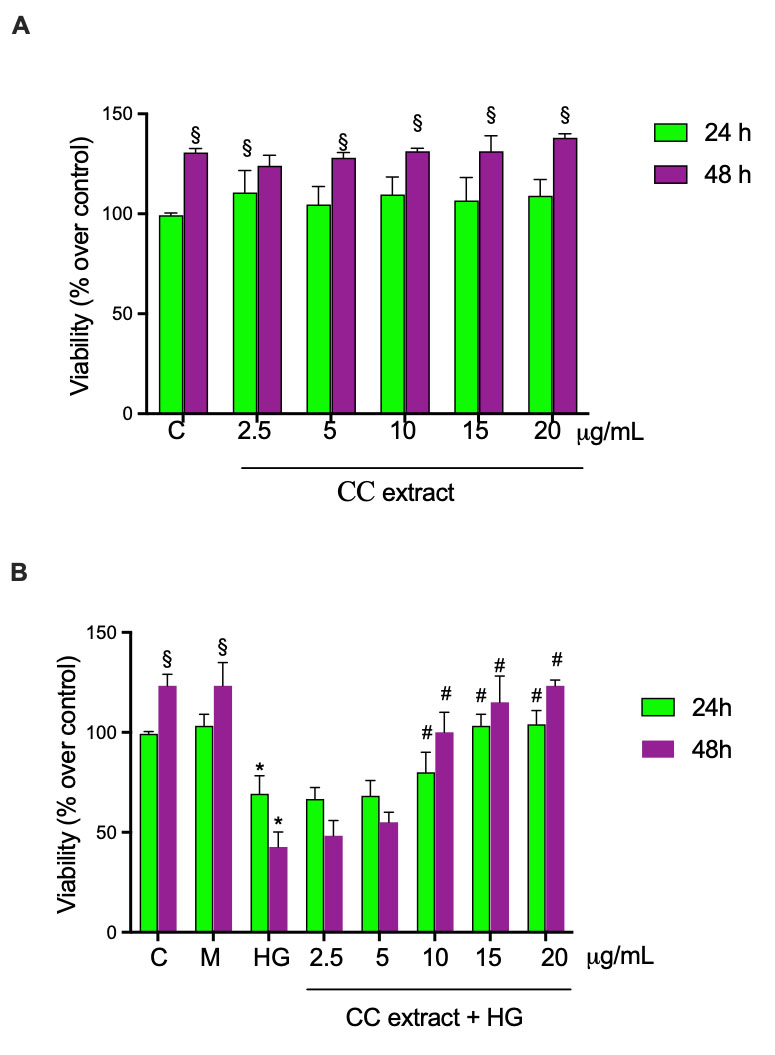 Fig. 2.
Fig. 2.
Cell viability was evaluated by MTT assays in human corneal
epithelial cells (HCEpiCs). Cells were treated for 24 and 48 h with cultivated
cardoon (CC) leaf extract 2.5–5–10–15 and 20 µg/mL in the absence
(A) and in the presence (B) of 25 mM glucose (HG). Mannitol 25 mM (M) was used as
osmotic control. Results are presented as the mean
Following these findings, further treatments were performed using HG for 24 h, excluding the incubation for 48 h that was excessively harmful for HCEpiCs. Moreover, the CC leaf extract was used at concentrations of 10 and 20 µg/mL, which gave the best protection results after treatment with HG.
The effect of CC leaf extract on epithelial barrier integrity was evaluated by TEER assays. HCEpiCs were plated on Transwell membranes and incubated for 6 h, 24 h and 48 h with 25 mM glucose (HG) in the absence or in the presence of 10 µg/mL and 20 µg/mL of CC leaf extract. As shown in Fig. 3, the treatment with HG significantly reduced the values by approximately 28%, 42% and 54% after 6 h, 24 h and 48 h, respectively, compared with untreated cells. Preincubation of the cells for 24 h with 10 µg/mL of CC leaf extract and incubation with fresh medium containing 10 µg/mL of the CC leaf extract in presence of HG for 6 h, 24 h and 48 h, significantly increased TEER by about 10%, 16% and 30%, respectively, when evaluated in comparison with HG-treated cells. Even more, preincubation of the cells for 24 h with 20 µg/mL of CC leaf extract and incubation with fresh medium containing 20 µg/mL of the CC leaf extract in presence of HG for 6 h, 24 h and 48 h, significantly increased TEER by about 30%, 42% and 67%, respectively, in comparison with HG-treated cells. Treatment with mannitol (M) 25 mM did not alter cell viability. The presence of 10 µg/mL or 20 µg/mL of CC leaf extract on control cells incubated for the same time in absence of HG did not modify TEER values compared with cells incubated without CC leaf extract (data not shown).
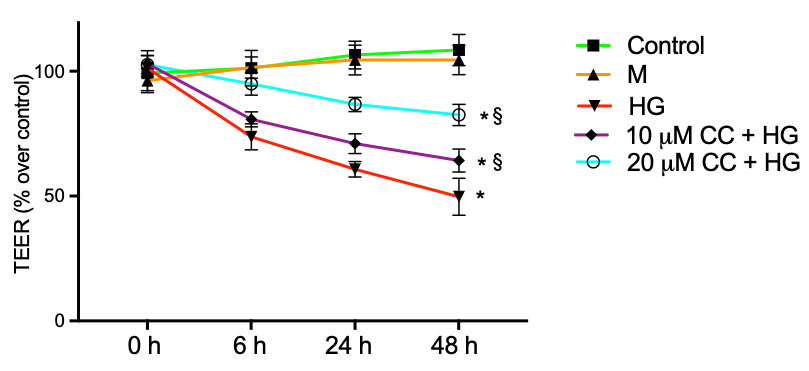 Fig. 3.
Fig. 3.
Transepithelial electrical resistance (TEER). Barrier integrity
was evaluated in HCEpiCs by TEER measurements after incubation for 24 h with 5.5
mM glucose (NG, Control) or 25 mM high glucose (HG) in absence or in presence of
10 µg/mL (10 µg/mL + HG) or 20 µg/mL (20
µg/mL + HG) of cultivated cardoon (CC) leaf extract. Mannitol 25 mM
(M) was used as osmotic control. Results are presented as the mean
The wound healing assay was employed to evaluate the effect of CC leaf extract on HCEpiCs under high glucose environment. As shown in Fig. 4A,B, the treatment with HG reduced the wound closure by about 50% after 6 and 24 h and by 63% after 48 h, in comparison to untreated cells. Preincubation of the cells for 24 h with 10 µg/mL or 20 µg/mL of CC extract and incubation with fresh medium containing 10 µg/mL or 20 µg/mL of the CC leaf extract in presence of HG for 24 h induced an increase of 1.5 and 1.8 fold, respectively, and after 48 h, an increase of 2.0 and 2.4 fold, respectively, in comparison to HG-treated cells.
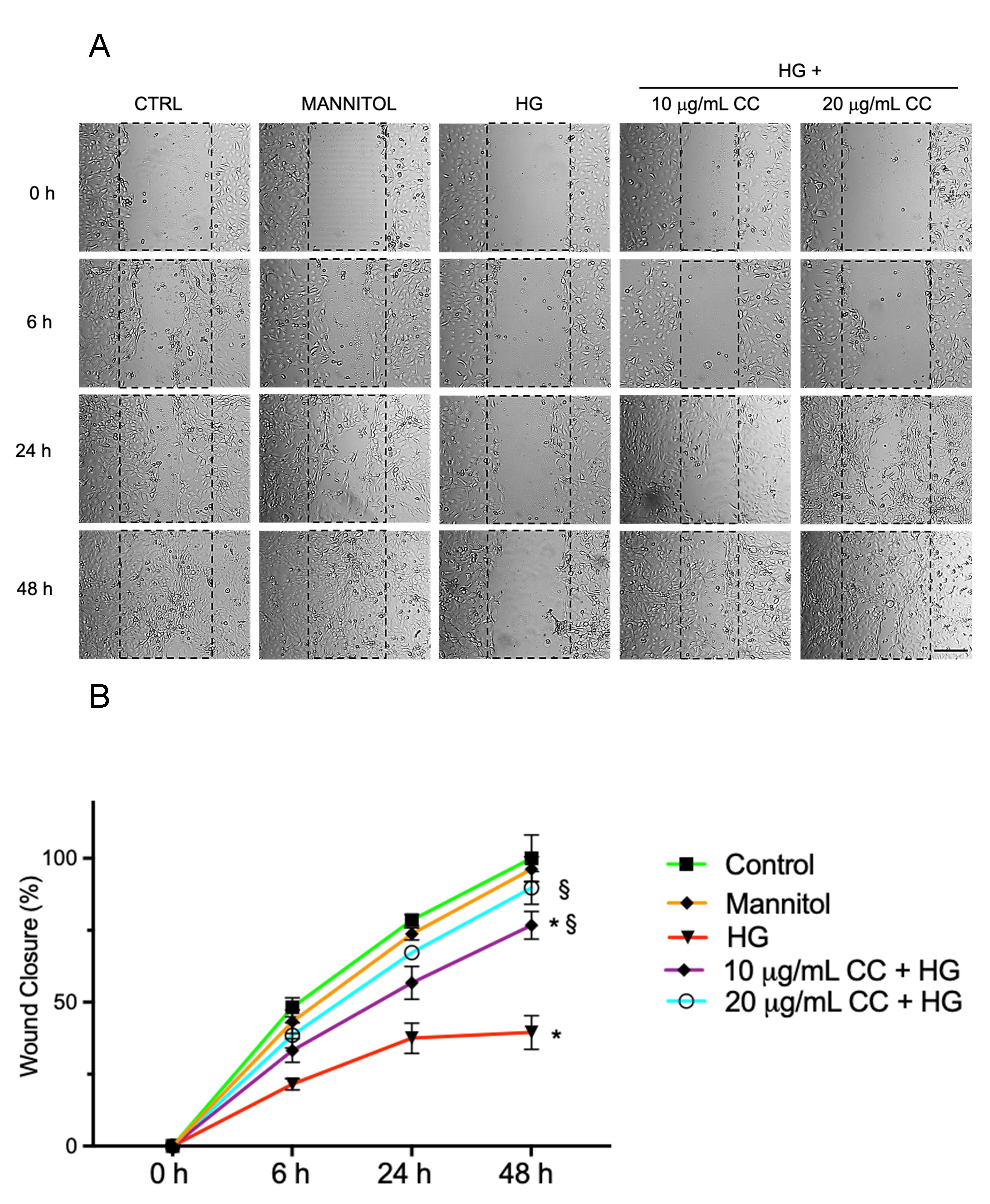 Fig. 4.
Fig. 4.
Migration of HCEpiCs after wound healing. (A) selected images
illustrating wound healing assays carried out in HCEpiCs incubated with normal
glucose (NG, 5.5 mM glucose, Control), Mannitol 25 mM, high glucose (HG, 25 mM
glucose), or with HG in presence of 10 µg/mL (10 µg/mL
+ HG) or 20 µg/mL (20 µg/mL + HG) of cultivated cardoon
(CC) leaf extract, immediately after the wound at 0 h and after 6, 24 and 48 h.
Images were captured using a Leica microscope at 20
High concentrations of glucose have been shown to cause the production of intracellular ROS, which leads to oxidative stress in rabbit corneal epithelial cells [35]. Our results also demonstrated a significant increase in ROS production after incubation of HCEpiCs with HG for 24 h compared with control cells incubated in a medium containing 5.5 mM glucose (NG). In Fig. 5, ROS production was significantly increased by 5.4 fold in comparison with control cells incubated in NG medium. Preincubation with 10 µg/mL and 20 µg/mL of CC leaf extract reduced ROS production by 45% and 74%, respectively, in comparison with HG-treated cells. Treatment with mannitol (M) 25 mM did not induce ROS production.
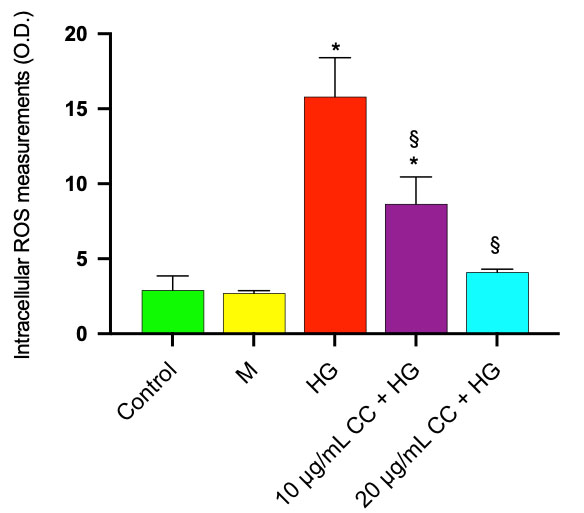 Fig. 5.
Fig. 5.
ROS production was evaluated in HCEpiCs after incubation with
DCFDA as described in Materials and Methods. HCEpiCs were incubated for 24 h in
the absence (Control) or in the presence of 25 mM glucose (HG) or in the presence
of 10 µg/mL (10 µg/mL CC + HG) or in the presence of 20
µg/mL (20 µg/mL CC + HG) of cultivated cardoon (CC)
leaf extract. Mannitol 25 mM (M) was used as osmotic control. Results are
presented as the mean
To determine whether CC leaf extract modulates the antioxidant defenses of HCEpiCs, we quantified the activities of superoxide dismutase (SOD) and catalase (CAT), as well as the intracellular concentration of glutathione (GSH). These biomarkers represent the primary enzymatic and non-enzymatic mechanisms responsible for scavenging reactive oxygen species within cells. The treatment with HG induced a significant decrease of SOD and CAT activities and GSH level. In Fig. 6A,B, the levels of SOD and CAT activity were reduced by 4.4 and 2.3 fold, respectively, after incubation with HG in comparison with control cells in normal glucose. Incubation of the cells with HG in association with 10 µg/mL or 20 µg/mL of CC leaf extract, induced an increase of SOD activity by 3.3 and 4.5 fold, respectively, and CAT activity by 1.8 and 2.2 fold, respectively, in comparison to HG-treated cells without CC leaf extract. GSH content was also significantly reduced by 2.3 fold after HG treatment compared with untreated cells (Fig. 6C). When the HG-treatment was conducted in association with 10 µg/mL or 20 µg/mL of CC leaf extract, the GSH amount increased by 1.8 and 2.2 fold, respectively, in comparison to HG-treated cells without CC leaf extract, highlighting a protective effect.
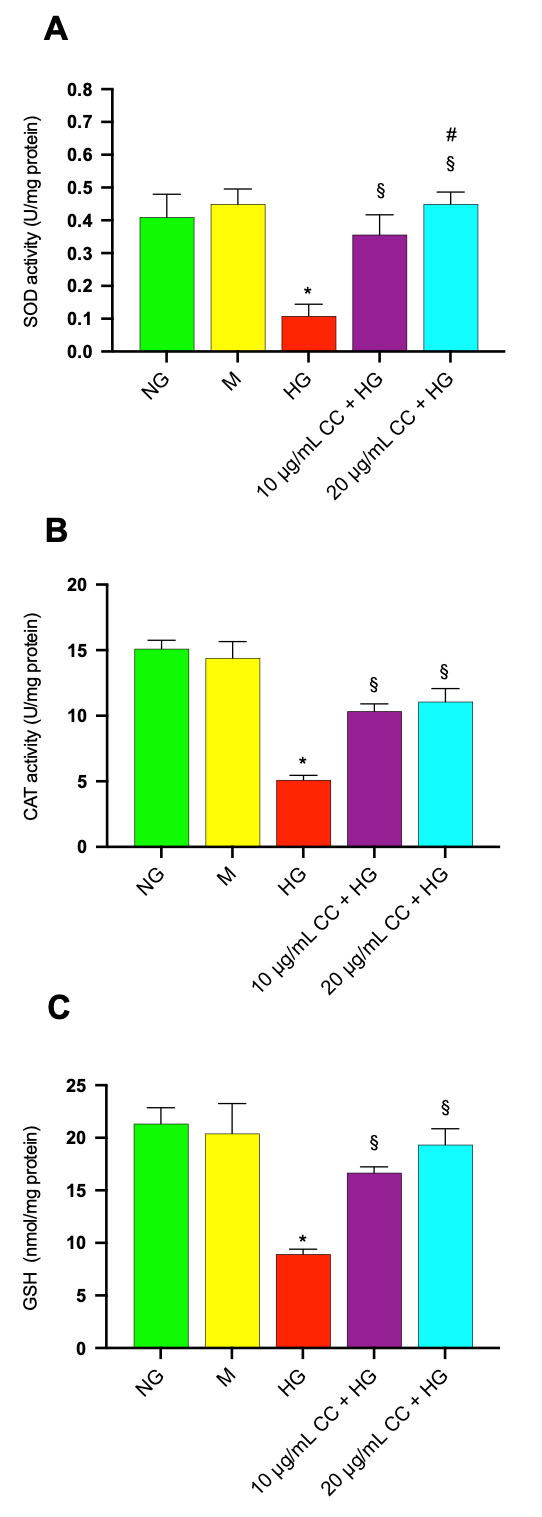 Fig. 6.
Fig. 6.
Effect of CC leaf extract on antioxidant defenses in
HCEpiCs exposed to high glucose: modulation of SOD and CAT activities (A, B) and
intracellular GSH levels (C). (A) Superoxide dismutase (SOD), (B) catalase (CAT)
antioxidant activity and (C) glutathione (GSH) content were evaluated in HCEpiCs
incubated for 24 h in the absence (Control, NG) or in the presence of 25 mM
glucose (HG) or in the presence of 10 µg/mL (10 µg/mL
CC + HG) or in the presence of 20 µg/mL (20 µg/mL CC +
HG) of cultivated cardoon (CC) leaf extract. Mannitol 25 mM (M) was used as
osmotic control. Results are presented as the mean
Since our in vitro study demonstrated that the activation of the ERK/cPLA2/COX-2 axis represents an early event after exposure of microvascular endothelial cells to HG [15], we investigated the potential involvement of ERK 1/2 and cPLA2 in our experimental model. It is well established that the phosphorylation of cPLA2 is essential for its activation, enabling it to release arachidonic acid from membrane phospholipids, a process that is regulated by ERK 1/2.
Immunoblots for p-ERK 1/2 and total ERK 1/2 are shown in Fig. 7A. Treatment of HCEpiCs with HG led to a significant 2.0 fold increase in phospho-ERK 1/2 levels compared with control cells, as shown in Fig. 7D (p-ERK 1/2/ERK 1/2 ratio). Incubation of the cells with HG in association with 10 µg/mL or 20 µg/mL of CC leaf extract reduced the phosphorylation of ERK 1/2 by about 2.2 fold and 2.6 fold, respectively, compared with HG-treated cells.
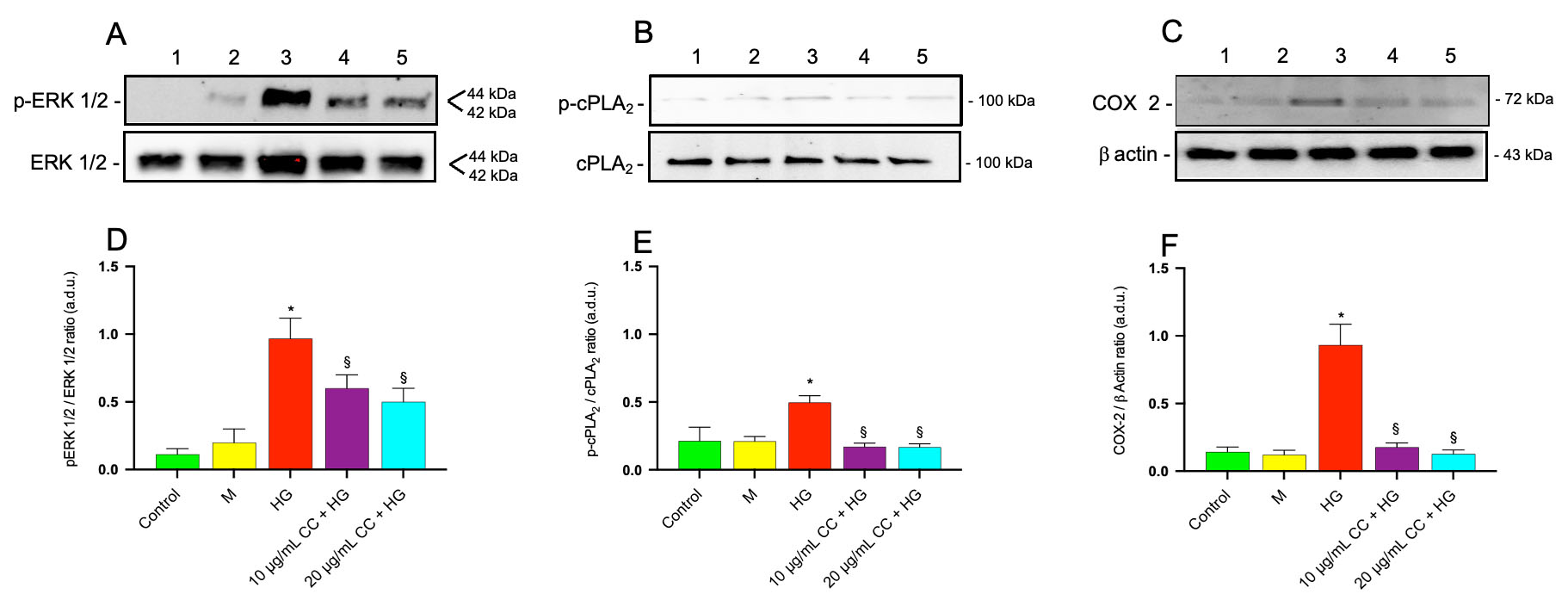 Fig. 7.
Fig. 7.
Effect of CC leaf extract on ERK 1/2, cPLA2 phosphorylation and
COX-2 expression in HCEpiCs exposed to high glucose. Immunoblot analysis was
conducted using specific antibodies targeting p-ERK 1/2 and total ERK 1/2 (A),
activated (phosphorylated, p-cPLA2) and total cPLA2 (B), as well as
COX-2 (C) proteins. The blots were also probed with an anti
Immunoblots for p-cPLA2 and total cPLA2 are shown in Fig. 7B and quantitative analysis of western blots (ratio of p-cPLA2/cPLA2) reported in Fig. 7E demonstrated a 3.2 fold increase in the phosphorylated (active) form of cPLA2 in HCEpiCs treated with HG compared with control cells. Incubation of the cells with HG in the presence of 10 µg/mL or 20 µg/mL of CC leaf extract reduced the phosphorylation of cPLA2 by 2.8 fold and 3.0 fold, respectively, compared with HG-treated cells. This HG-induced increase in cPLA2 phosphorylation supports enhanced enzyme activity, leading to more arachidonic acid being available for cyclooxygenase activity. Based on this, we assessed COX-2 protein levels after treating HCEpiCs with HG, both in the presence and the absence of CC leaf extract.
Immunoblots for COX-2 are shown in Fig. 7C. The quantitative analysis of
immunoblots for COX-2, represented by the COX-2/
No significant change in p-ERK, p-cPLA2 and COX-2 expression was observed in Mannitol treated cells compared with controls.
Since COX-2 levels were significantly increased after HG treatment, PGE2 release
was quantified in HCEpiC treated with HG in the absence/presence of 10
µg/mL or 20 µg/mL of CC leaf extract. Treatment of
HCEpiCs with HG increased PGE2 release by 2.5 fold in comparison to untreated
cells (Fig. 8A). The treatment with HG in association with 10 µg/mL
or 20 µg/mL of CC leaf extract reduced the release by 1.6 and 1.9
fold, respectively. PGE2 levels in the medium of untreated HCEpiC were 380
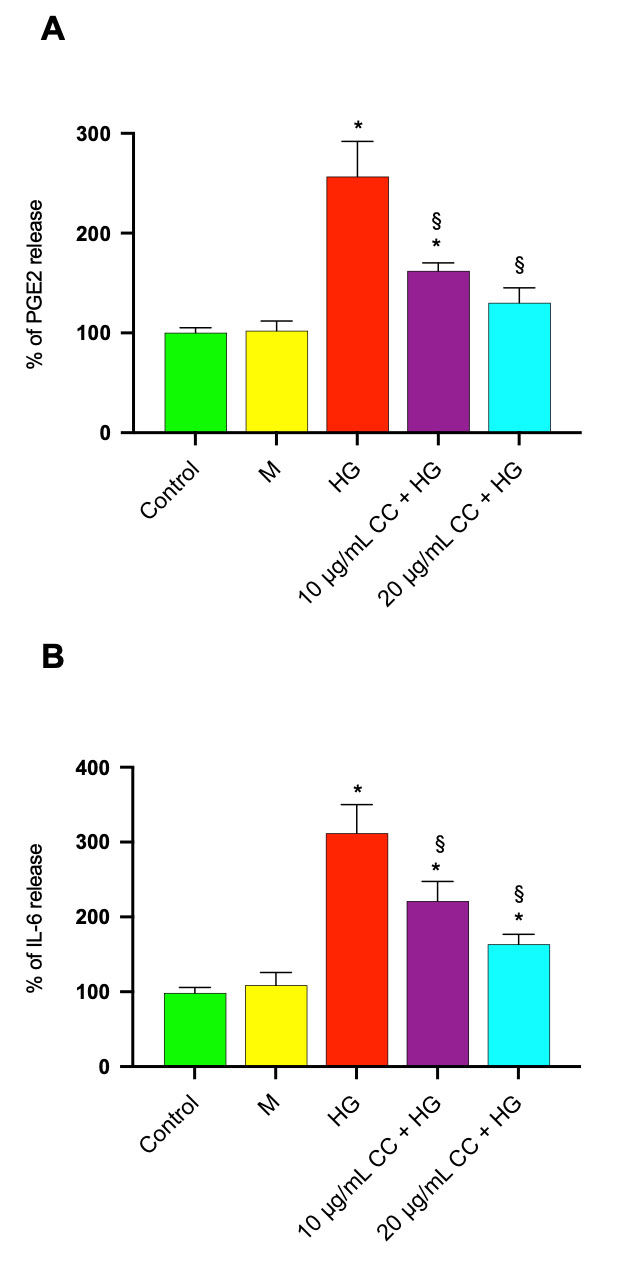 Fig. 8.
Fig. 8.
Effect of CC leaf extract on PGE2 and IL-6 release in HCEpiCs
exposed to high glucose. Prostaglandins E2 (PGE2) and interleukin-6 (IL-6)
release were evaluated in medium of HCEpiCs incubated for 24 h in the absence
(Control) or in the presence of 25 mM glucose (HG) or in the presence of 10
µg/mL (10 µg/mL CC + HG) or in the presence of 20
µg/mL (20 µg/mL CC + HG) of cultivated cardoon (CC)
leaf extract. Mannitol 25 mM (M) was used as osmotic control. After incubation,
aliquots of supernatants were used for PGE2 (A) and IL-6 (B) determination by
ELISA Assay Kits. Results are presented as the mean
To confirm that treatment with CC leaf extract was able to reduce the
inflammatory process, IL-6 levels were measured in the medium of HCEpiC treated
with HG in the absence/presence of 10 µg/mL or 20 µg/mL
of CC leaf extract. As shown in Fig. 8B, HG treatment increased IL-6 release by
about 3.0 fold compared to untreated control cells. The treatment with HG in
association with 10 µg/mL or 20 µg/mL of CC leaf
extract reduced the release by 1.4 and 1.8 fold, respectively. IL-6 levels in the
medium of untreated HCEpiC were 127
No significant change in PGE2 and IL-6 release was observed in Mannitol treated cells compared with controls.
The global rise in diabetes mellitus has led to a worrying increase in complications associated with the disease. Chronic hyperglycemia causes significant biochemical and metabolic changes that result in both macrovascular and microvascular damage. Macrovascular complications primarily affect the heart and brain, while microvascular damage contributes to neuropathy, nephropathy, and ocular tissue injuries [2]. Patients with diabetes often experience a range of ocular surface issues, including epithelial defects, recurrent corneal erosion, ulcers, corneal edema, superficial punctate keratitis, and delayed corneal epithelial wound healing. In particular, diabetic keratopathy is an ocular complication of diabetic peripheral neuropathy, characterized by impaired corneal epithelial wound healing and reduced subbasal corneal nerve plexus density and hypoesthesia [6]. Clinically, this manifests as ocular surface discomfort, often described as burning or dryness, due to impaired corneal innervation and epithelial integrity [36]. Multiple studies have demonstrated that the diabetic corneal epithelium is characterized by increased cellular fragility, decreased epithelial cell density, reduced epithelial thickness, and compromised barrier integrity. These structural and functional abnormalities are indicative of disrupted cellular homeostasis, involving dysregulation of critical processes such as proliferation, differentiation and migration [37].
It has been shown that both oxidative stress and the inflammatory process are
responsible for the damage to the ocular tissues of diabetic patients.
Recognizing the significant role of oxidative stress in the progression of
diabetic complications, extensive research has investigated the protective
potential of various antioxidants, particularly those derived from plants.
Natural compounds such as
Polyphenols, a diverse class of secondary metabolites found in plant-based foods such as fruit, vegetables, nuts and tea, contribute to the maintenance of ocular physiological homeostasis and promote tissue repair under pathological conditions [39, 40].
Cynara cardunculus L. botanical complex is recognized as a rich source of health-promoting compounds, including polyphenols, inulin, vitamins, and minerals [26, 27]. Leaf extracts of both globe artichoke and cardoons have traditionally been used extensively in folk medicine, particularly for the treatment of liver disorders. The therapeutic effects of these extracts have often been attributed to key phenolic constituents particularly present in the heads and leaves, such as 1caffeoylquinic acids and flavones [41].
In our study, CC leaf extract alone did not alter cell viability, as reported in Fig. 2A, nor did it affect any of the measured parameters (data not shown). The treatment of HCEpiCs with 25 mM glucose significantly reduced corneal cell viability, confirming results by others that demonstrated a decreased epithelial cell density in the cornea in the presence of elevated glucose levels, both in animal models and in human research studies [42, 43]. To select the CC leaf extract concentration with the best protective effect on HG-treated HCEpiCs, cell viability was determined using MTT assay, which provides information on cellular metabolic activity, thereby discriminating between living and dead cells. The results demonstrated that the pre-treatment of corneal cells with cultivated cardoon extract preserved their viability by protecting them from HG damage.
Physiologically, to fulfill its protective barrier function, the cornea maintains a high TEER, guaranteed by the presence of tight junctions between epithelial cells, as well as a marked regenerative capacity which, in response to an insult, activates mechanisms of cell proliferation and migration [4]. Ultrastructural analysis via scanning electron microscopy revealed disrupted intercellular architecture in diabetic rats, associated with compromised integrity among corneal cells, responsible of the epithelial barrier [44] and in vitro studies also demonstrated significantly lower TEER values in corneal epithelial cell monolayers under diabetic conditions [12]. Our findings confirmed an HG-induced significative reduction in TEER, indicating a compromise in the integrity of the corneal epithelial barrier, and demonstrated that CC leaf extract maintained high TEER values in the presence of 25 mM glucose. Furthermore, we confirmed that HG treatment slows down the wound healing of HCEpiC [13]. In the present study, the treatment with HG in the presence of CC leaf extract effectively accelerated migration and wound closure to normal levels. Rapid re-epithelialization is essential to ensure the physiological function of the cornea; CC leaf extract, by ensuring a re-epithelialization process, potentially prevents corneal damage. In diabetic patients, delayed epithelial healing increases the risk of ocular complication and microbial infections, particularly in the context of an already compromised immune system [45, 46]. Consequently, considerable efforts have been directed toward identifying agents capable of enhancing epithelial repair in the diabetic cornea such as cytokines [47], silk-derived proteins [48] and natural substances [49]. Our findings showed the effect of CC extract in promoting and accelerating cell migration, counteracting the damage of HG, and represent a promising outcome in the context of therapies based on the use of natural substances.
A close correlation has been also shown between diabetic corneal dysfunction and increased ROS production. Prolonged hyperglycemia can trigger glucose-induced protein auto-oxidation and promote ROS generation [35].
Oxidative stress is the consequence of a disrupted balance in which ROS are generated at a rate that surpasses the cell’s antioxidant capacity to neutralize or repair the resulting oxidative damage, particularly present in the conjunctiva and cornea, highly exposed to UV radiation and particularly vulnerable to oxidative stress [50]. In diabetic mice, corneal injury leads to heightened oxidative stress, which significantly hinder corneal epithelial healing and elevate the risk of persistent epithelial defects [51].
The endogenous antioxidant defense system includes both enzymatic and nonenzymatic antioxidants. Enzymatic antioxidants consist of SOD, CAT, glutathione peroxidase and glutathione oxidase, and, among the non-enzymatic antioxidants, a significant role is played by GSH, synthesized by metabolic processes [52]. In patients with type 2 diabetes mellitus, insulin treatment has been shown to only partially reduce oxidative stress, as evidenced by persistently low GSH levels [10].
We therefore examined the effect of CC leaf extract on ROS concentrations and on several antioxidant enzyme activities and we found that CC leaf extract significantly decreased the production of ROS, increased SOD and CAT antioxidant activities as well as GSH content in HCEpiCs in a high glucose environment.
In eukaryotic cells, the bulk of oxidative metabolism is carried out in the mitochondria and endoplasmic reticulum. Within these organelles, standard redox reactions inevitably lead to the generation of free radicals, which, if not efficiently neutralized, can contribute to oxidative stress. The mitochondrial electron transport chain is responsible for producing approximately 90% of cellular ROS [53]. Moreover, metabolic pathways involving enzymes such as COX-2 and cPLA2 play a key role in ROS generation. A study on an in vivo/ex vivo hypoxia model showed that the activation of cPLA2 is involved in ventilator-induced diaphragm dysfunction by promoting mitochondrial ROS production, highlighting a positive interaction between cPLA2 activity and oxidative stress at the mitochondrial level [54]. It also demonstrated in microglial cells, stimulated with lipopolysaccharide (LPS), an increase in the cPLA2 activation alongside ERK1/2 activation and the ROS production induced by LPS was significantly reduced by cPLA2 inhibitors such as arachidonyl trifluoromethyl ketone (AACOCF3) but not by the iPLA2 inhibitor BEL, indicating that cPLA2 is specifically involved in mediating LPS-induced oxidative stress in microglial cells [55]. Moreover, in lens epithelial cells, cPLA2 plays a crucial role in PDGF-stimulated cell growth, with its activation tightly regulated by calcium-dependent membrane translocation. This membrane targeting is essential for cPLA2 to activate NADPH oxidase, leading to increased ROS production. These findings highlight that cPLA2 plays a key role in linking calcium signaling to oxidative stress in corneal epithelial cells [56].
Therefore, elevated intracellular ROS not only damage macromolecules but also function as signaling molecules, triggering a cascade of biochemical and molecular responses that disrupt normal cellular signaling pathways [57, 58]. Our results showed the significant inhibitory effect of CC extract on HG-induced cPLA2 and ERK1/2 phosphorylated forms and COX-2 and on PGE2 and IL-6 production. These findings suggest that CC extract may exert wide protective effects on glucose stressed HCEpiCs. One could hypothesize that, during ocular pathologies due to diabetes complications, free radicals could activate the ERK1/2/cPLA2/COX-2 signaling, triggering the inflammatory process and, at the same time, the activation of this pathway could lead to the production of other ROS, creating a positive feedback loop (Fig. 9). We have not directly tested whether ROS act exclusively as upstream activators or are generated as a downstream product of this cascade. Further investigation will be necessary to validate the regulatory role of ROS in this context.
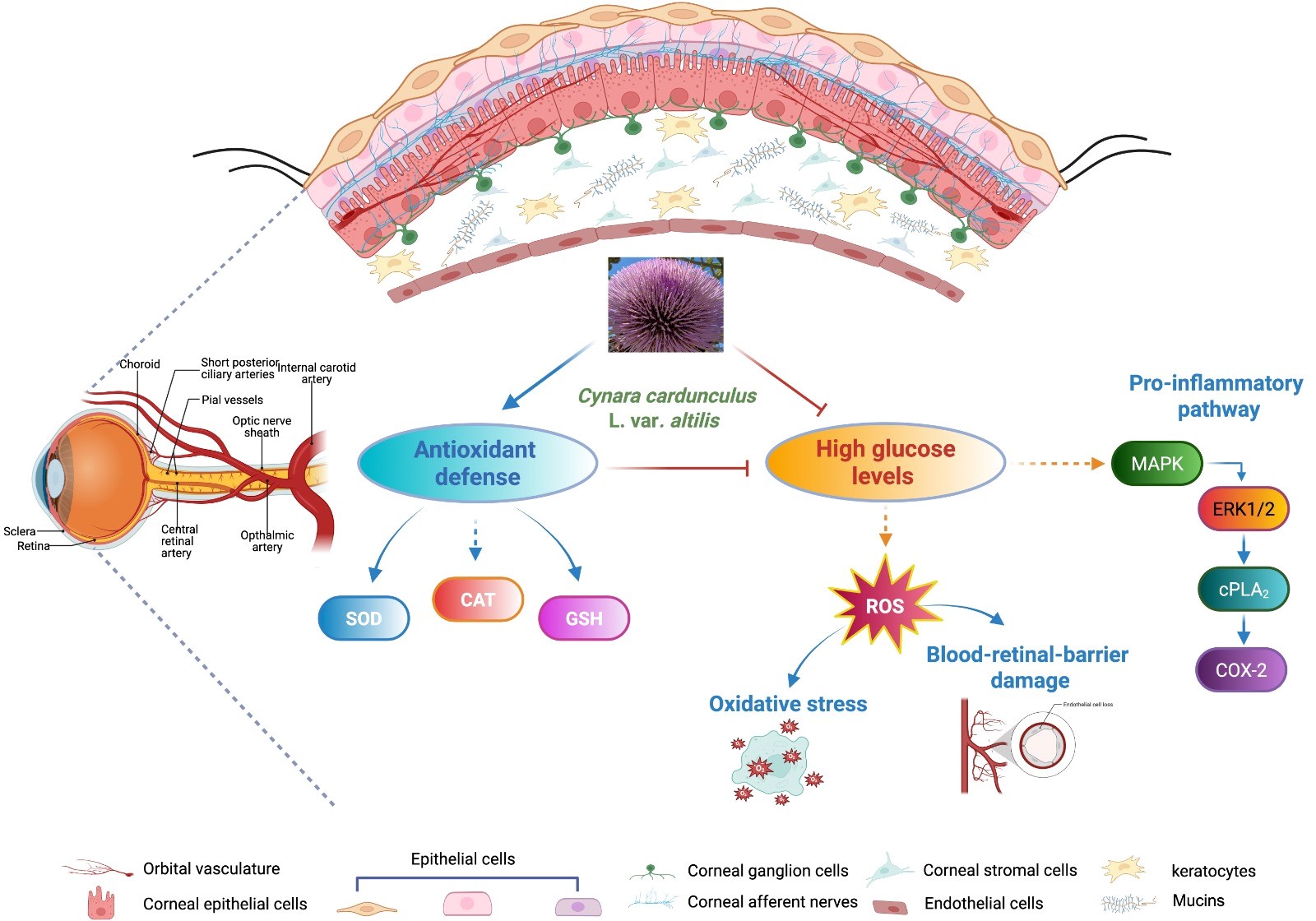 Fig. 9.
Fig. 9.
Potential signaling pathways modulated by Cynara cardunculus L. cultivated cardoon (CC) leaf extract in high glucose exposed human corneal cells. Cynara cardunculus L. extract enhances the antioxidant defense enzymes including SOD, CAT activities and GSH content and inhibits ROS production and pro-inflammatory pathways, especially ERK 1/2/cPLA2/COX-2 signaling in high glucose exposed human corneal cells, highlighting the possibility that it could perform a protective function in the management of diabetic keratopathy. BioRender. Scuto, M. (2025) https://BioRender.com/an2rc7d.
In this scenario, identifying therapeutics that could alleviate these conditions is crucial for advancing new treatment strategies for diabetic keratopathy. Our results indicate that CC leaf extract could have strong potential as a therapeutic agent, controlling both the inflammatory process and the production of ROS, thus maintaining intact the protective and regenerative functions of the cornea.
This study reports preliminary results on the effect of CC extract after in vitro treatment of corneal cells with HG. It is hoped that the results obtained will lay the foundation for future in vivo and clinical studies. Clearly, CC extract alone will not be able to replace the traditional therapy that diabetic patients undergo, but it could be used as a dietary supplement to reduce the damage in diabetic keratopathy.
Despite decades of research into oxidative stress as a key pathogenic factor in the development of diabetes mellitus and its complications, this area remains highly relevant. Contemporary approaches to diabetes management are increasingly centered on personalized antioxidant therapies, incorporating targeted delivery systems and strategies that focus on specific sources of ROS.
Further investigation should be carried out to identify reliable biochemical markers of free radical activity and to explore the interplay between oxidative stress and inflammation, aiming to better understand their combined roles in ocular disease progression.
We investigated the effects of CC extract on cell migration and viability using an in vitro model of hyperglycemia-induced corneal cells. Our results demonstrated that cultivated cardoon leaf extract (i) reduced oxidative stress by lowering ROS production and enhancing SOD, CAT activities and GSH content in high glucose exposed human corneal cells; (ii) maintained a high TEER and promoted corneal epithelial wound healing; (iii) induced down-regulation of the ERK 1/2/cPLA2/COX-2 signaling pathway involved in the inflammatory process and ROS production.
Collectively, these findings indicate that cultivated cardoon extract has the potential to enhance human corneal epithelial cell function. This provides a scientific basis for considering its development as a therapeutic agent in the prevention or treatment of diabetic keratopathy.
All data reported in this paper will also be shared by the lead contact upon request.
GL, ATS, and CDA designed the research study. AC, AA, CP, SL, GP and MS performed the research. GL, ATS, CDA and CL analyzed the data. GL, ATS, and CDA wrote the manuscript. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
The authors thank Antony Bridgewood for language assistance. For some experimental procedures the authors used instruments available in the Service Center B.R.I.T. BIO-Nanotech Research and Innovation Tower. The authors thank AUDI project proposal.
This research was funded by “University of Catania under Grant Scheme PIACERI”, Linea di Intervento 1, University of Catania, Italy.
The authors declare no conflict of interest. Given her role as the Editorial Board member, Gabriella Lupo had no involvement in the peer-review of this article and has no access to information regarding its peer review. Given her role as the Guest Editor, Maria Scuto had no involvement in the peer-review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process for this article was delegated to Adrian Gericke.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.