




 , Bernard Moussian 2,7,*,§
, Bernard Moussian 2,7,*,§
1 Dr. Margarete Fischer-Bosch Institute of Clinical Pharmacology, 70376 Stuttgart, Germany
2 Animal Genetics, University of Tuebingen, 72076 Tuebingen, Germany
3 School of Pharmaceutical Science and Technology, Tianjin University, 300072 Tianjin, China (present address)
4 University of Tuebingen, 72074 Tuebingen, Germany
5 Institute of Clinical Pharmacology, Klinikum Nuremberg, Paracelsus Medical University Nuremberg, 90419 Nuremberg, Germany (present address)
6 Departments of Clinical Pharmacology, Pharmacy and Biochemistry, University of Tuebingen, 72076 Tuebingen, Germany
7 University Côte d’Azur, INRAE, CNRS, Sophia Agrobiotech Institute, 06903 Sophia Antipolis, France
†These authors contributed equally.
§These authors contributed equally.
Abstract
Membrane transport proteins are critical determinants of systemic and intracellular drug levels, thereby contributing substantially to drug response and/or adverse drug reactions. Therefore, the U.S. Food and Drug Administration and the European Medicines Agency, the regulatory authorities for drug approval in the U.S. and Europe, respectively, recommend pre-clinical testing of selected drug transporters during the drug development process to elucidate clinically relevant drug–drug interactions (DDIs). In the current proof-of-principle study, we describe the generation of fruit flies expressing human membrane drug transporters in their salivary glands to enable DDI studies in a time-saving manner and at low costs.
Using the Gal4/upstream activation sequence (UAS) expression system, we established fruit flies expressing human organic cation transporters (hOCTs) 1 and 2 and genetic variants thereof. Both transporters are key drug uptake transporters in humans and are recommended for pre-clinical DDI studies. After injecting fluorescent hOCT substrates, their accumulation in salivary gland cells was observed by confocal laser scanning microscopy.
We demonstrate the feasibility of expressing hOCT1 and hOCT2 in the salivary glands of fruit fly embryos and subsequent alteration by clinically relevant genetic variants, corroborating results from mammalian cell experiments. Moreover, we show an OCT-dependent accumulation of the prototypic fluorescent OCT substrates ethidium (Et+) and 4-Di-1-ASP (4-(4-(dimethylamino)styryl)-N-methylpyridinium, ASP+) in the salivary gland cells and subsequent inhibition by clinically-used OCT drug inhibitors.
Based on the handling procedure and the lack of need for Animal Protection Act approval, we propose that the humanized Drosophila melanogaster fruit fly model opens a new avenue for pre-clinical functional transporter studies.
Keywords
- Drosophila melanogaster
- membrane transport proteins
- organic cation transporter 1
- organic cation transporter 2
- organic anion transporters
- ATP binding cassette transporter subfamily B member 1
- drug development
- drug interactions
- pharmacogenomic variants
Plasma membrane transporters are critical determinants of drug disposition in
humans. Particularly transporters from the ATP-binding cassette (ABC) and the
solute carrier (SLC) superfamily are directly involved in processes affecting
drug pharmacokinetics. This has been discussed in detail in several review
articles [1, 2, 3] and further details can be found in the references cited in these
review articles. Membrane transporters and their genetic variants therefore
contribute to drug failure, drug resistance and adverse drug reactions (ADRs).
These roles have been summarized in several review articles [4, 5, 6]. For further
information, additional studies are cited within these reviews. Of the various
transporter families, human organic cation transporter (hOCT) 1 (encoded by
SLC22A1) and hOCT2 (SLC22A2) and human organic anion
transporting polypeptide human organic anion transporting polypeptide 1B1
(hOATP1B1) (SLCO1B1), hOATP1B3 (SLCO1B3) and hOATP2B1
(SLCO2B1) have emerged as particularly significant due to their roles in
the uptake and distribution of a wide range of clinically relevant drugs. Human
OCT1 and hOCT2 mediate the uptake of positively charged compounds including
metformin, morphine, sumatriptan and tropisetron, particularly in the liver and
kidney. Similarly, hOATP1B1, hOATP1B3 and hOATP2B1 are crucial hepatic
transporters responsible for the uptake of various anionic drugs such as statins,
certain antibiotics and anticancer agents. Functional impairments or inhibition
of these transporters can lead to altered systemic drug levels, reduced efficacy
or increased toxicity. The clinical importance of these transporters is
emphasized by the observed clinical consequences of common and rare genetic
transporter variants. For instance, variants in SLC22A1 have been linked
to altered metformin pharmacokinetics and therapeutic response in patients with
type 2 diabetes mellitus. Moreover, the common genetic variant c.521T
Current research on membrane transporters typically utilizes in vitro systems such as transfected mammalian cell lines, membrane vesicles and hepatocyte cultures to characterize transporter function and drug interactions [11]. These in vitro data are often complemented by in vivo studies in rodents, including knockout mouse models, which require ethical approval with implications for cost and timelines. While rodent models have significantly advanced the understanding of transporter function [12], they present challenges including species-to-species differences in transporter expression and function, long development times and resource-intensive maintenance.
To address these limitations, there is growing interest in identifying simpler models that can provide functional insights at a lower cost and with greater experimental flexibility. Here, Drosophila melanogaster (fruit fly) offers numerous advantages: fruit flies are genetically tractable, inexpensive to maintain and allow for rapid, tissue-specific gene manipulation [13]. Therefore, we sought to develop a new strategy using the fruit fly as a novel model to implement human membrane transporter testing in the pre-clinical drug development process (summarized in Fig. 1A–D) [14]. Using the galactose-responsive transcription factor Gal4/upstream activation sequence (Gal4/UAS) expression system [15], human membrane transporters and their clinically-relevant genetic variants are expressed in the salivary glands of Drosophila embryos (Fig. 1A,B). Drosophila embryos were chosen because they are amenable to micromanipulation and injection procedures. Moreover, they are transparent and can be immobilized so that fluorescent compounds can be easily visualized by confocal fluorescence imaging. Although Drosophila analogs of human transporters have been described [14, 16], they were not utilized due to the absence of direct one-to-one orthologs and the limited functional characterization available. The insect salivary glands instead of the Malpighian tubules (analogous to kidney) or the fat body (analogous to liver) were chosen as the target organ for three reasons: (i) salivary glands are simple epithelial organs consisting of polarized cells with one side facing the hemolymph (blood) system and the other side facing the glandular lumen, thus resembling the physiology of human hepatocytes and proximal tubular epithelial cells of the kidney, which are major sites of drug-relevant transporter-related barrier function [4, 17]; (ii) they are of suitable size amenable for light and confocal fluorescence microscopy; (iii) they are not essential during embryonic and larval stages, thus allowing expression and functional analyses of human proteins without compromising viability. These humanized flies are used in their embryonic stage to study the interaction of drugs with membrane transporter proteins by applying transporter-specific fluorescent tracers together with an investigational drug (Fig. 1C,D). Since fruit fly embryos are transparent, the fluorescent tracers can be assessed directly in the living embryos. For instance, reduced accumulation of the transporter-specific fluorescent tracer and subsequent reduced fluorescence within the salivary gland cells will be determined if the investigational drug competes with the fluorescent tracer for uptake by the membrane transporter (Fig. 1D). The functional consequences of transporter genetic variants can also be assessed.
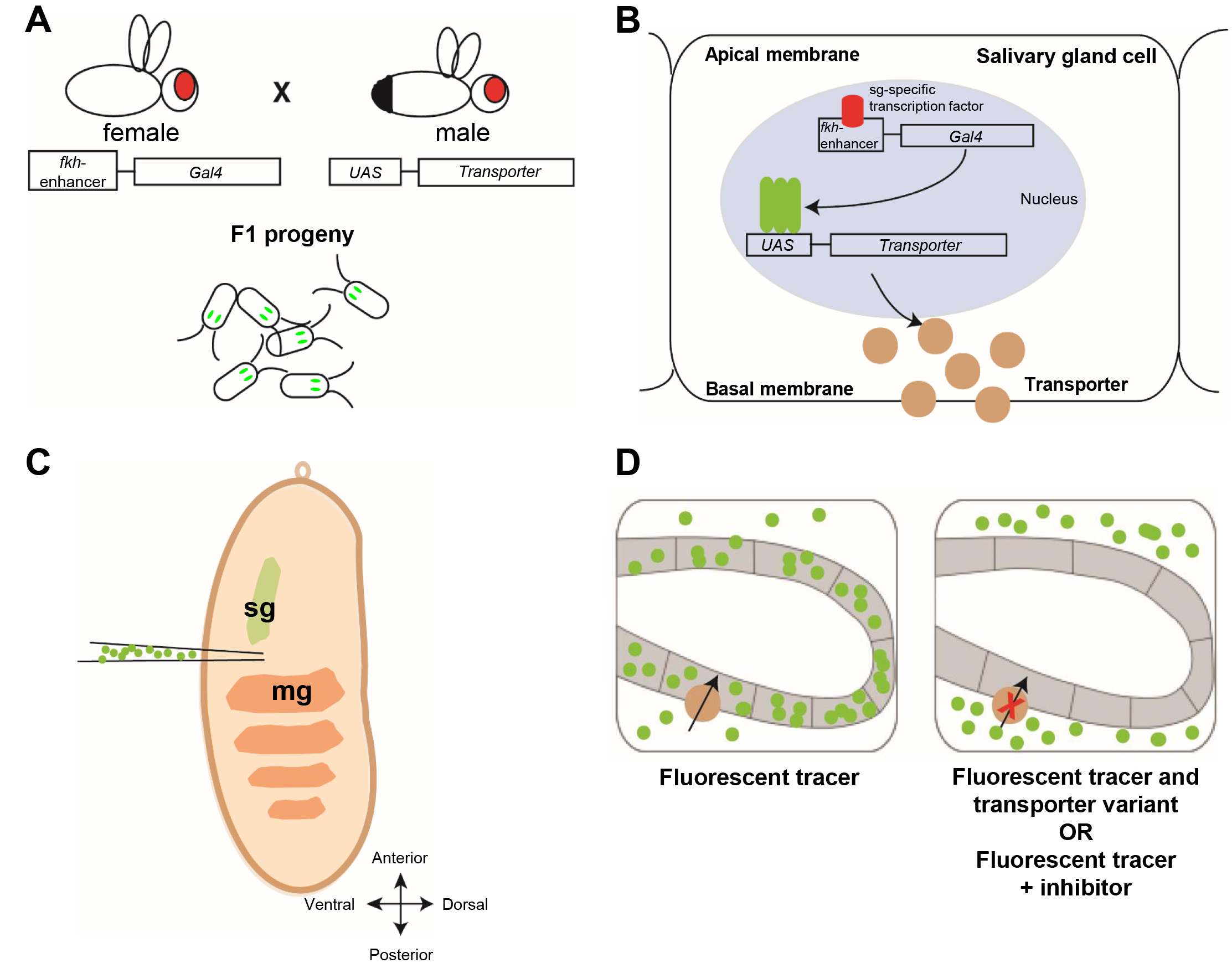 Fig. 1.
Fig. 1.
The general principle of the Drosophila human membrane transporter test system and its establishment for human organic cation transporter (hOCT) 1. (A,B) The Gal4/upstream activation sequence (Gal4/UAS) system mediates human membrane transporter expression in salivary glands. (A) Male flies with a UAS-transporter insertion in their genome are crossed with females harboring a fkh-Gal4 insertion. The eggs with both fkh-Gal4 and UAS-transporter are harvested for injection. Their salivary glands are marked by green fluorescent protein (GFP) expression (green). (B) In the eggs, expressed under the control of the fkh enhancer, the Gal4 transcription factor drives the expression of the transporter coding gene specifically in epithelial cells of the salivary glands during embryonic stage 14–17. Membrane uptake transporters (beige circles) will locate to the basolateral side of the membrane of salivary gland cells. (C) Injection of fluorescent tracers or drugs. For a homogenous distribution of injected molecules, the needle is stung into the embryos at the ventral side between the salivary gland (sg) and midgut (mg). (D) Uptake transporters in the basolateral membrane (beige circles) mediate the accumulation of fluorescent tracers provided by injection (green dots) into the salivary gland cells; simultaneous injection of the fluorescent tracer and an inhibitory drug or expression of reduced function variants prevents accumulation of the fluorescent tracer in the cell. The figure was drawn using Adobe Illustrator CS6 (Adobe Inc., San Jose, CA, USA).
The cDNAs encoding full-length hSLC22A1 (NM_003057.2) and hSLC22A2 (NM_003058.2) were cloned into pcDNA3.1 as described [18] and variants thereof were produced by site-directed mutagenesis using the QuikChange II XL Site-Directed Mutagenesis kit (product number 200521, Agilent Technologies, Waldbronn, Germany). Mutagenesis primer sequences are given in Supplementary Table 1.
Transporter genes were then amplified from the respective pcDNA3.1 constructs using specific primers designed for In-Fusion cloning (Takara Bio Europe SAS, Saint-Germain-en-Laye, France) (Supplementary Table 2A). Previously generated constructs of hSLCO1B1 and hSLCO1B3 (common haplotype 1) [19], hSLCO2B1 [20] and human ATP-binding cassette transporter B1 (hABCB1) [21] served as templates as well. Amplicons were cloned into the pUAST-attB transposon-vector (Drosophila Genomics Resource Center, Bloomington, IN, USA) using the In-Fusion HD cloning kit (product number 639650, Takara Bio Europe SAS, Saint-Germain-en-Laye, France).
The transgenic flies were generated by BestGene Inc. (Chino Hills, CA, USA). The pUAST-attB human transporter gene constructs were injected into y[1] w[1118]; PBac{y[+]-attP-9A}VK00005 flies that harbor the attP-site at the position 75A10 on chromosome 3 and an endogenous PhiC31 source. The pUAST-attB-based transposon was, hence, inserted into this position of the genome. To visualize the salivary glands, a paired organ with a monolayer of cuboidal epithelial cells in the anterior half of the embryo (see Fig. 2), and the expression of the target gene later in Gal4-driven expression experiments, we recombined each UAS-hOCT1/2 insertion to a UAS-green fluorescent protein (GFP) insertion on the same chromosome. Information on the fly lines and their sources are given in Supplementary Table 3 and schemes of the fly crossing experiments are presented in Supplementary Fig. 1. All the recombined lines were validated through Sanger sequencing of the respective transgenes (primers in Supplementary Table 2B). The fly line harboring the salivary gland specific fkh-Gal4 on the 3rd chromosome was obtained from Marta Llimargas (IBMB, Barcelona, Spain). Fly lines were reared on standard soybean-molasses-yeast-agar medium at 18 or 25 °C.
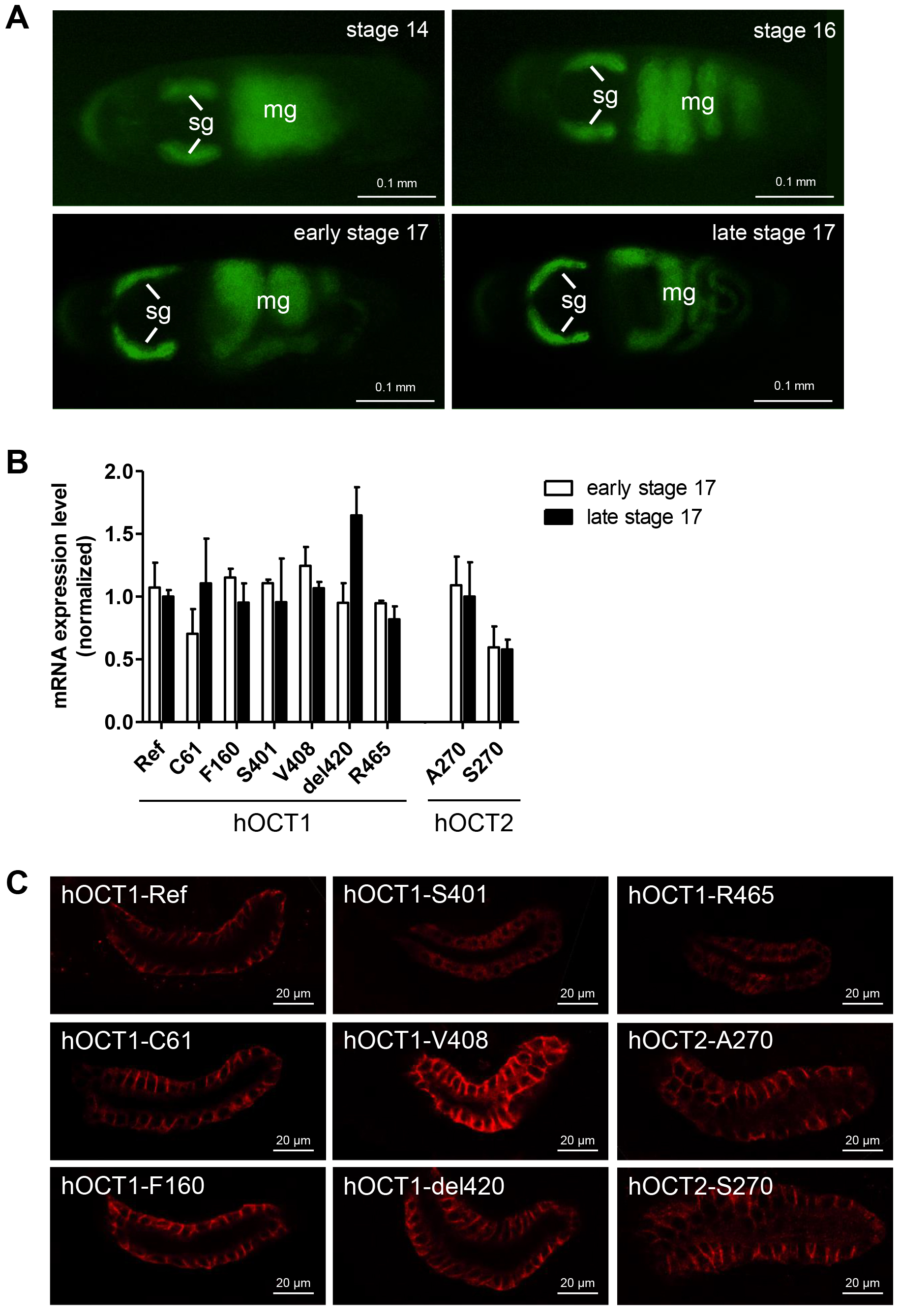 Fig. 2.
Fig. 2.
fkh-Gal4 driven expression during fruit fly embryonic development. (A) GFP expression in the salivary glands (sg) was promoted by fkh-Gal4. The stages were determined by the morphology of the midgut (mg) that auto-fluoresces. Scale bar = 0.1 mm. (B) Transcript levels of fkh-driven expression of hOCT1 and hOCT2 and variants thereof were analyzed by quantitative polymerase chain reaction (PCR) in both early (white) and late (black) stage 17 embryos. The data are normalized against the transcript level of hOCT1-Ref for hOCT1 variants and hOCT2-A270 for hOCT2-S270 in late stage 17 embryos. Bars show standard errors of 3 experiments, in which 70–100 eggs were prepared for RNA extraction. (C) Localization of hOCT1 and hOCT2 and variants thereof (red signal) in salivary gland cells by immunodetection and confocal microscopy. Scale bar = 20 µm.
Fruit fly strains expressing recombinant human membrane transporters and the respective UAS-human transporter expression vectors are available on request by qualified researchers for their own use.
Additional information is given in the Supplementary Methods.
The RNeasy mini kit plus RNase-Free DNase Set (product numbers 74134 and 79254, Qiagen, Hilden, Germany) was used to extract RNA from 70–100 dechorionated eggs [22] for each sample. The GFP-positive eggs were identified based on the observation of the GFP signal in salivary glands under a fluorescence microscope (AZ100, Nikon, Tokyo, Japan). Gut morphology was used for staging as described [23]. Transcripts were quantified using the FastStart Essential DNA Green Master kit (product number 06402712001, Roche, Basel, Switzerland) on a Roche LightCycler Nano instrument (Roche, Basel, Switzerland). The transcript levels of the reference gene RpS20 were used for normalization of gene expression as described [24]. Additional information and polymerase chain reaction (PCR) primers are given in the Supplementary Methods and in Supplementary Table 4.
The embryos were fixed applying the slow formaldehyde fixation method [25]. Detection of hOCT1 or hOCT2 was performed with anti-human OCT1 monoclonal antibody 2C5 (MA5-15730, Thermo Fisher Scientific, Karlsruhe, Germany) at a dilution of 1:500 and anti-human OCT2 monoclonal antibody MAB6547 (R&D systems, Minneapolis, MN, USA) at a dilution of 1:50. The secondary antibody was AlexaFluor 568 goat anti-mouse IgG (Cat.No. A-11004, Thermo Fisher Scientific/Invitrogen, Karlsruhe, Germany, 1:500). Immunolocalization of hABCB1 was performed using antibody HPA002199 (Merck KgaA/Sigma Aldrich, Darmstadt, Germany) at a dilution of 1:100 and AlexaFluor 568 goat anti-rabbit IgG (Cat.No. A-11011, Thermo Fisher Scientific/Invitrogen, Karlsruhe, Germany, 1:500) as secondary antibody. Immunodetection was carried out following standard protocols [22]. Staining of human OCT1 or OCT2 in the salivary gland cells was observed with a fluorescence microscope (AZ100, Nikon, Tokyo, Japan) or an inverted confocal microscope (Zeiss LSM 510, Oberkochen, Germany).
Immunodetection of human OATP1B1, OATP1B3, and OATP2B1 in embryos is described in the Supplementary Methods and in Supplementary Table 5.
For collection of eggs, flies were kept at 25 °C in egg laying cages on
an apple juice agar plate prepared as described [22] garnished with a spot of
fresh baker’s yeast. Stage 16 eggs were dechorionated, and selected based on the
presence of GFP. For each injection, 50 to 60 eggs were collected. The eggs were
lined on a fresh apple juice agar plate, anterior to posterior, and the lateral
side facing up. Homemade heptane glue prepared according to Walker & Dahmann
[22] was brushed to a stripe on a cover slip. The egg line was transferred to the
cover slip by gently touching the glue stripe. The cover slide was put on a
microscope slide, and incubated for 8 minutes in a dry chamber (humidity
Ethidium bromide (EtBr), mercury dibromofluorescein disodium (MDBF) and 4′,5′-dibromofluorescein (DBF) were from Merck KgaA/Sigma Aldrich (Darmstadt, Germany), cholyl-lysyl-fluorescein (CLF) was from Corning Inc. (Tewksbury, MA, USA) and 4-(4-(dimethylamino)styryl)-N-methylpyridinium iodide was from Thermo Fisher Scientific, Karlsruhe, Germany (D288). By confocal microscopy, the fluorescence of ethidium (Et+) was excited with the 543 nm laser and detected through the 560/615 nm emission filter. The other fluorescent substrates were excited with the 488 nm laser and detected using the 505/530 nm emission filter.
The 488 nm excitation laser and 505/530 nm emission filter were used to detect the GFP signal and the 543 nm laser and the 560/615 emission filter to detect the red fluorescent protein (RFP) signal. Only alive embryos identified by active movement were recorded. For imaging, the plane of the specimen with maximal salivary gland lumen area was focused on.
To quantify the uptake of Et+ by salivary gland cells, the ratio of the
Et+ signal between salivary gland and epidermis cells (sg/epi) was
determined. The measurement was performed using ImageJ 1.50i (National Institute
of Health, Bethesda, MD, USA). Microscopy images were first converted to 16-bit
gray scale images. An area with four to six cells with the strongest Et+
signal was chosen by the polygon select function, and the mean gray value of this
area was measured. The mean gray value of the epidermis at the same focal plane
was measured to obtain the sg/epi ratio. To compare the Et+ uptake
efficiency between different hOCT variants or after inhibitor treatment, the
sg/epi ratio from each group was subtracted from the sg/epi ratio of the negative
control group, and normalized to the result of hOCT1-Ref or hOCT2-A270.
Statistical significance was analyzed using Student’s t-test.
Differences were considered to be significant when p
The cimetidine concentration that achieved half-maximum inhibition (IC50) of Et+ accumulation was determined by non-linear regression using Prism 5.04 (GraphPad, Boston, MA, USA) applying the following equation: (Y = bottom + (top–bottom)/(1 + ((X/IC50)slope))).
The generation of human embryonic kidney 293 (HEK293) FlpIn cells stably expressing hOCT1, hOCT2 and variants thereof has been described [27, 28]. The cell lines were received from Dr. Kathy M. Giacomini (Dept. of Bioengineering and Therapeutic Sciences, University of California, San Francisco, USA). The cell lines were demonstrated to stably overexpress the respective transporter by using microarray analyses. As expected, each transporter was highly expressed only in its corresponding overexpressing cell line when compared to the vector-transfected control cell line [29]. Cells were cultured in high glucose Dulbecco’s Modified Eagle’s Medium (D5796, Merck KgaA/Sigma Aldrich, Darmstadt, Germany) supplemented with 10% fetal bovine serum (Merck KgaA/Sigma Aldrich, Darmstadt, Germany), 100 U/mL penicillin and 100 mg/mL streptomycin (Lonza, Basel, Switzerland) and 85 µg/mL hygromycin B (Thermo Fisher Scientific/Invitrogen, Karlsruhe, Germany) at 37 °C and 5% CO2. Cells were regularly tested for mycoplasma and were negative for mycoplasma contamination using a PCR detection kit (Cat. No. 11-1025, Venor®GeM Classic mycoplasma detection kit, Minerva Biolabs GmbH, Berlin, Germany).
Cells were grown to confluence, trypsinized and collected by centrifugation. Cell pellets were resuspended in 10 mL growth medium and 5 mL growth medium were added. 200 µL of the cell suspension was then seeded in each well of 96-well black plates (View plate 96F TC black, #6005182, Revvity/Perkin Elmer, Waltham, MA, USA), which had been pre-treated with poly-L-lysine (Biochrom AG, Berlin, Germany) for better cell attachment. Cells were then incubated for 24 h at 37 °C and 5% CO2. For analysis of cellular Et+ accumulation, the cells were washed once with Hanks’ balanced salt solution (HBSS, Thermo Fisher Scientific/Gibco #14025, Karlsruhe, Germany) warmed to 37 °C and then incubated at 37 °C and 5% CO2 with 100 µL/well HBSS containing 5 µM EtBr (HP47.1, Carl Roth, Karlsruhe, Germany). Cellular accumulation was terminated after different time points by removing the HBSS/EtBr solution and washing the cells twice with ice-cold HBSS. After final removal of the ice-cold HBSS, fluorescence of the cells was measured in a multimode plate reader (EnSpire, Revvity/Perkin Elmer, Waltham, MA, USA) at an excitation wavelength of 535 nm and an emission wavelength of 590 nm. Subsequently, cells were lysed by adding 100 µL lysis buffer (10 mM MeOH/1 mM Tris/EDTA 1:1, pH 8) per well and incubated at room temperature for 10 min. Protein content of the lysed cells was determined by the bicinchoninic acid assay as described [18].
Time-dependent accumulation values of Et+ were fitted by non-linear
regression (least squares fit) with Prism 5.04 (GraphPad, Boston, MA, USA) as
described [30] using the following formula [31]: cin = kin/kout
As a proof of principle, we established fruit flies expressing the human organic cation transporters hOCT1 (SLC22A1) or hOCT2 (SLC22A2). We selected these transporters because they are clinically relevant and therefore recommended for pre-clinical testing during drug development by the regulatory authorities from the US and Europe, FDA and EMA, respectively, as well as the International Transporter Consortium [17]. Of note, hOCT1 mediates the uptake of several frequently used medications (e.g., metformin, morphine, sumatriptan, tropisetron) from the blood into the hepatocytes, while hOCT2 is the major uptake transporter in kidney proximal tubule cells, with consequences for drugs such as cisplatin and metformin [17, 32]. Moreover, genetic variations in both proteins are crucial for drug response and adverse drug reactions (ADRs) [7].
To construct transposons for transgenic flies, the open reading frames coding for hOCT1 reference (hOCT1-Ref) and six common hOCT1 variants (C61, F160, S401, V408, del420, R465) [27] and for hOCT2-S270 and hOCT2-A270 [28] (Supplementary Table 6) were cloned downstream of the UAS regulatory sequence. Transgenic lines were generated with the respective transposons that were integrated into the fly genome at the same position on chromosome 3 to avoid chromosomal position effects on gene expression. To visualize salivary gland-specific expression, these insertions were recombined to a UAS-GFP insertion for GFP co-expression.
The salivary gland-specific expression of hOCT1 and hOCT2 was induced by Gal4 expressed under the control of the fkh promoter [33] (Fig. 2A). The expression levels of different hOCT1 and hOCT2 variants remained stable in stage 17 embryos for several hours (Fig. 2B). The hOCT1-Ref protein and both hOCT2 proteins were localized to the lateral and basal plasma membrane of salivary gland cells (Fig. 2C). This localization corresponds to the localization of hOCT1 in human hepatocytes [34] and hOCT2 in renal proximal tubule epithelial cells [35]. Four hOCT1 variants (C61, F160, V408, del420) were also localized in the basolateral membrane, whereas variants S401 and R465 were predominantly found in the cytoplasm (Fig. 2C). These results are consistent with previous studies using mammalian HEK293 cells (Supplementary Table 6). The only exception is variant hOCT1-C61, which is predominantly localized intracellularly when expressed in mammalian cells [36]. One reason for this discrepancy might be that hOCT1-C61 is temperature-sensitive, i.e., that it is properly trafficked to the plasma membrane at the temperature of the fly experiments, but not at 37 °C in the mammalian cells. An example of temperature-sensitive trafficking to the plasma membrane is given by a common variant in the ABC transporter cystic fibrosis transmembrane conductance regulator (CFTR), deltaF508. This variant is retained in the endoplasmic reticulum (ER) at 37 °C, but traffics out of the ER to the plasma membrane at lower temperatures of 23–30 °C [37]. The mechanistic investigation of temperature sensitivity of hOCT1-C61 is of interest and should be explored in future studies, e.g., by rearing embryos at higher temperatures or lowering the temperature in the mammalian cell system. However, it was not intended to do this in the present study, as we wanted to build a medium to high-throughput system that could be used with a minimum of parameters to be set.
We chose ethidium (Et+) to study function of human organic cation uptake transporters because it is a fluorescent cation and membrane impermeable requiring transporters in the plasma membrane to enter cells [38]. After injection of 0.5 µM EtBr into the hemolymph of stage 17 wild type embryos, the distribution of Et+ in the embryos was observed by confocal microscopy (Fig. 3A). Et+ accumulated in epidermal and tracheal cells, but not in the salivary gland cells. This observation suggests the presence of endogenous Drosophila transporters capable of transporting organic cations in epidermal and tracheal cells. It further confirms that expression of human OCT1 in the salivary gland cells can be used to monitor hOCT1 activity because there was no interference with endogenous transport activity. Indeed, only in embryos expressing hOCT1-Ref in their salivary glands, an Et+ signal was detected in the salivary gland cells as early as 30 minutes after injection (Fig. 3B). The course of Et+ uptake by salivary gland cells was recorded every 30 minutes for five hours. Et+ accumulation increased up to 5 hours, after which the larvae hatched from the embryos. The hatching indicated that viability was not compromised and it is safe to use Et+ in the embryos. This demonstrates that hOCT1-Ref transports Et+ into the salivary gland cells. In contrast, the salivary glands of embryos of the negative control group were free of Et+ (Fig. 3B).
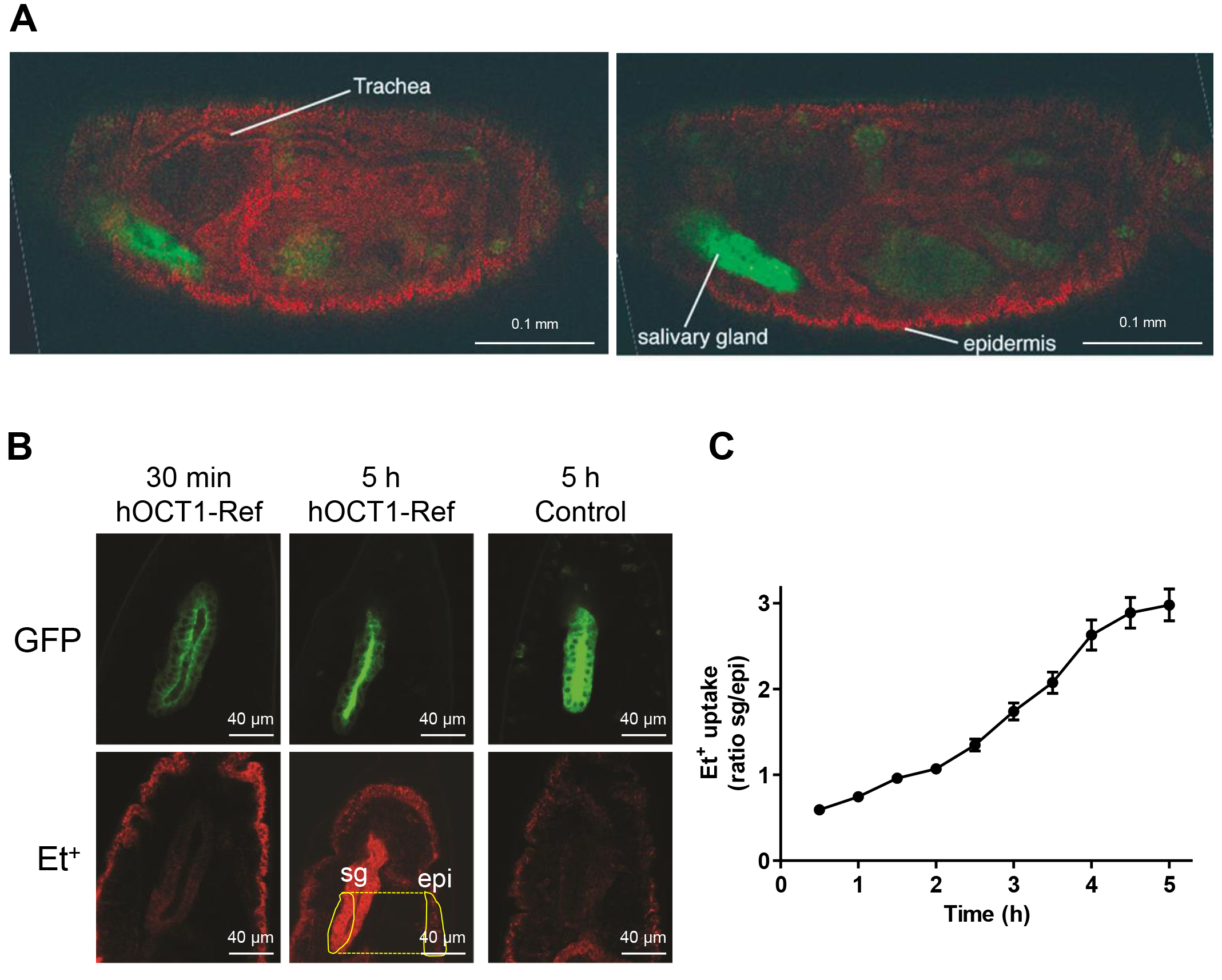 Fig. 3.
Fig. 3.
Establishment of the Drosophila human membrane transporter test system for hOCT1. (A) Tissue distribution of ethidium (Et+) in wild type fruit fly embryos 4 hours after injection (red). Left and right images show the same embryo at different focal planes. The embryo was injected with 0.5 µM ethidium bromide (EtBr) and incubated for four hours at room temperature in darkness. The dye (red) filled the whole body lumen of the embryo, and was strongly concentrated in epidermal and tracheal cells. The position of the salivary gland (labeled with GFP, green) was free of the red Et+-signal. Scale bar = 0.1 mm. (B) Et+ (red, 0.5 µM EtBr injected) accumulates specifically in hOCT1-Ref positive salivary glands, which are visualized through GFP co-expression (green). Et+ accumulation was already detectable 30 min after injection and increased during 5 hours. As exemplified here, the signal intensities in the areas of the salivary glands (sg) and the epidermis (epi) marked with yellow lines were used to calculate the sg/epi ratio. The salivary glands of negative control embryos were free of Et+ staining even five hours after injection. Scale bar = 40 µm. (C) Time course of Et+ uptake. The Et+ uptake into the salivary gland cells increased linearly over time. Bars indicate standard errors of 20–21 injections.
Since the Et+ signal intensity varied slightly between individual embryos
due to variable volumes of injected dye, we used the hOCT-independent uptake of
Et+ by epidermal cells as an internal reference to directly compare the
uptake activity of different hOCT variants. We calculated the ratio of Et+
intensity between the salivary glands and the epidermis (sg/epi) to quantify the
salivary gland Et+ uptake (Fig. 3B). The ratio determined for hOCT1-Ref
increased over time from 0.55
The activity of genetic variants of hOCT1 and hOCT2 was analyzed through injection of 0.5 µM EtBr. Et+ uptake was recorded at 1 hour and 4 hours post-injection. The transport activities of all six hOCT1 variants were significantly lower than the activity of hOCT1-Ref (Fig. 4A,B; Table 1a, 1b). Three classes of hOCT1 variants were identified: (1) variants with moderate activity reduction of 25% (hOCT1-V408) and 30% (hOCT1-C61, hOCT1-F160), (2) variants with strong activity reduction of 60% (hOCT1-del420) and 80% (hOCT1-S401), and (3) the null-function variant hOCT1-R465 (Table 1b). Because in vitro data of Et+ uptake by hOCT1 variants were not available, we measured uptake in mammalian HEK293 cells expressing different hOCT1 variants (Supplementary Fig. 3A,B,D) and could show that Et+ uptake by transgenic hOCT1-expressing fly embryos is similar to data obtained with hOCT1-expressing mammalian cells. For variants V408, F160, S401 and R465 these results are in line with data for various other compounds, which has been reviewed previously [6, 7]. Of note, we observed a strong activity reduction for the del420 variant, both in the transgenic fly embryos and in the mammalian HEK293 cell line. This is in line with the notion that the del420 variant has substrate-specific transport activity. Thereby, Et+ can be added to the substrates, for which a strong reduction of transport activity can be observed (e.g., metformin, tropisetron, tyramine, monocroatline), while various others are known to be transported similar to the reference sequence (e.g., cycloguanil, debrisoquine, sparteine, sumatriptan) [6, 7]. The only exception is C61, which shows a strong activity reduction in the mammalian cell system (Supplementary Fig. 3A,D) as expected for this variant [6, 7], but only a moderate activity reduction in the fly embryos (Fig. 4B). This is probably due to the predominant localization of C61 in the plasma membrane of salivary gland cells (Fig. 2C) as discussed above in subsection 3.1. Both hOCT2 variants showed similar Et+ transport activities (Fig. 4C), again in agreement with the uptake experiments using mammalian HEK293 cells (Supplementary Fig. 3C,D) and results for other compounds [7]. Nevertheless, strictly speaking, we cannot exclude that differences in protein levels of the variants might explain the differences in Et+ uptake, although the transgenes are inserted at the same chromosomal position ensuring identical expression levels.
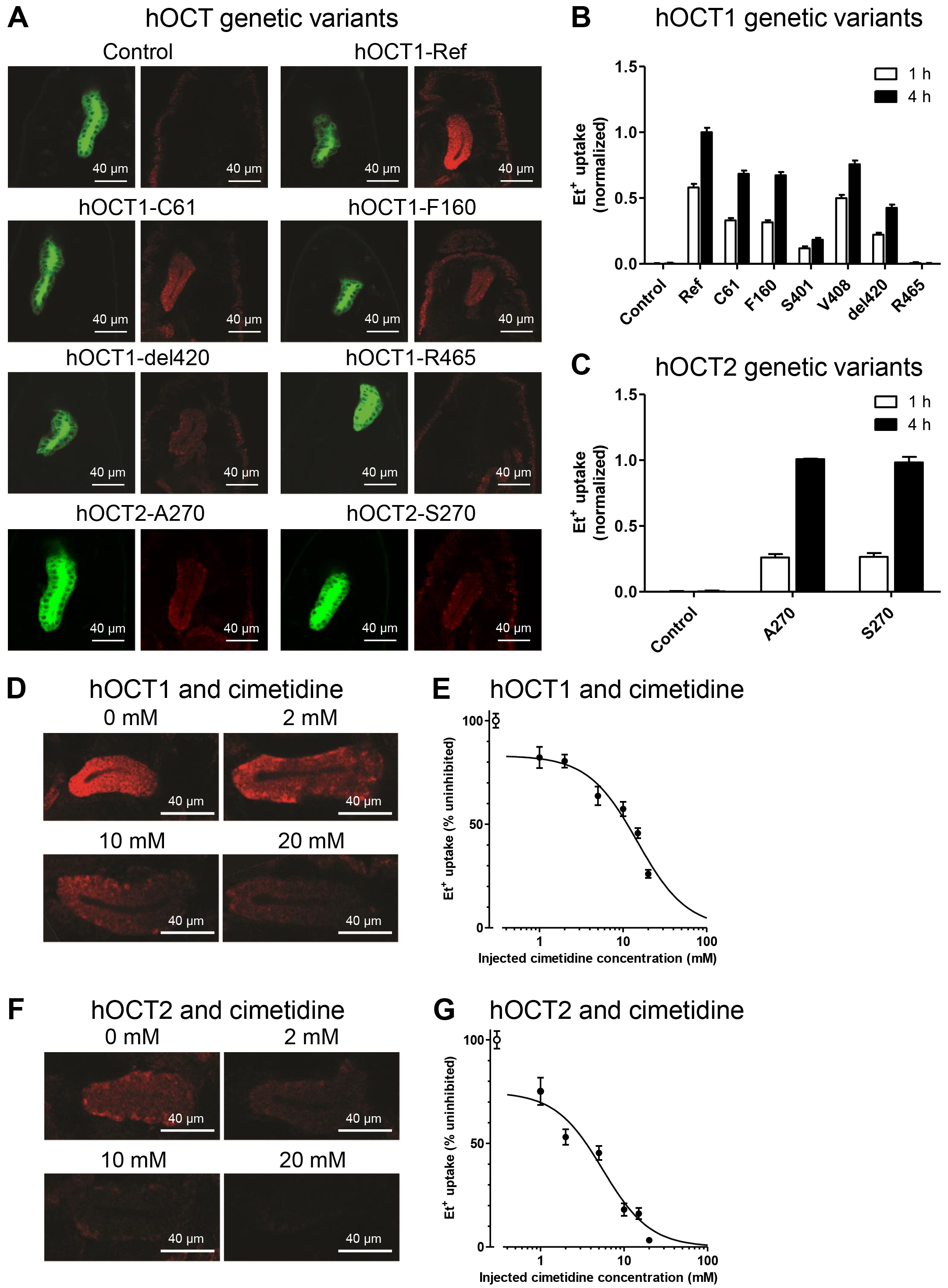 Fig. 4.
Fig. 4.
Application of the Drosophila human membrane
transporter test system for clinically-relevant genetic hOCT variants and
transport inhibitor cimetidine. (A) Representative images of ethidium (Et+)
uptake by hOCT1-Ref, hOCT2 and genetic variants thereof. Embryos of the negative
control did not express any hOCT1 or hOCT2. The uptake of Et+ was
detected by confocal microscopy 4 h after injection. 4 of the 6 investigated
variants are shown. Scale bar = 40 µm. (B) Quantification of
Et+ uptake by hOCT1 genetic variants. Data were normalized to the uptake by
hOCT1-Ref after 4 h. Bars indicate standard errors of 34–90 injections. (C)
Quantification of Et+ uptake by hOCT2-A270 and hOCT2-S270. Data were
normalized to the uptake by hOCT2-A270 after 4 h. Bars indicate standard errors
of 64–90 injections. (D,F) Co-injection of 0.5 µM EtBr with 2 mM,
10 mM and 20 mM cimetidine caused a concentration dependent reduction of Et+
uptake by hOCT1- or hOCT2-expressing salivary gland cells compared to the uptake
by salivary gland cells in absence of cimetidine (0 mM) 4 h post-injection. Scale
bar = 40 µm. (E,G) Inhibitor-response curves for the effects of
cimetidine on Et+ uptake by hOCT1-Ref and hOCT2-A270. Assuming a 17-fold
dilution after injection, the apparent IC50 values are 0.87
| Genetic variant | 0.5 µM EtBr | 6.25 µM EtBr | Activity ratio | ||||
| Et+ signal (sg/epi) | Et+ signal minus control (sg/epi) | Activity (% to hOCT1-Ref) | Et+ signal (sg/epi) | Et+ signal minus control (sg/epi) | Activity (% to hOCT1-Ref) | 0.5 µM:6.25 µM | |
| Neg. control | 0.27 |
– | – | 0.4 |
– | – | – |
| Ref | 1.9 |
1.63 | 100 | 1.4 |
1.0 | 100 | – |
| C61 | 1.2 |
0.93 | 57.1 | 1.1 |
0.7 | 70 | 0.82 |
| F160 | 1.2 |
0.93 | 57.1 | 1.3 |
0.9 | 90 | 0.63 |
| S401 | 0.61 |
0.34 | 20.9 | 0.69 |
0.29 | 29 | 0.72 |
| V408 | 1.7 |
1.43 | 87.7 | 1.4 |
1.0 | 100 | 0.88 |
| del420 | 0.9 |
0.63 | 38.7 | 0.7 |
0.3 | 30 | 1.29 |
| R465 | 0.28 |
0.01 | 0.6 | 0.4 |
0 | 0 | – |
sg/epi, salivary gland/epidermis; Et+, ethidium; EtBr, ethidium bromide; hOCT1, human organic cation transporter 1.
| Genetic variant | 0.5 µM EtBr | 6.25 µM EtBr | Activity ratio | ||||
| Et+ signal (sg/epi) | Et+ signal minus control (sg/epi) | Activity (% to hOCT1-Ref) | Et+ signal (sg/epi) | Et+ signal minus control (sg/epi) | Activity (% to hOCT1-Ref) | 0.5 µM:6.25 µM | |
| Neg. control | 0.37 |
– | – | 0.5 |
– | – | – |
| Ref | 3.2 |
2.83 | 100 | 3.5 |
3 | 100 | – |
| C61 | 2.3 |
1.93 | 68.2 | 2.3 |
1.8 | 60 | 1.13 |
| F160 | 2.3 |
1.93 | 68.2 | 2.7 |
2.2 | 73.3 | 0.93 |
| S401 | 0.89 |
0.52 | 18.4 | 1.3 |
0.8 | 26.7 | 0.69 |
| V408 | 2.5 |
2.13 | 75.3 | 2.8 |
2.3 | 76.7 | 0.98 |
| del420 | 1.5 |
1.13 | 39.9 | 1.4 |
0.9 | 30 | 1.33 |
| R465 | 0.31 |
0 | 0 | 0.4 |
0 | 0 | – |
sg/epi, salivary gland/epidermis; Et+, ethidium; EtBr, ethidium bromide; hOCT1, human organic cation transporter 1.
Values given are means
We next investigated the effect of Et+ concentration on uptake efficiency in the Drosophila test system by injecting two different concentrations, i.e., 0.5 µM (low) and 6.25 µM (high). For each variant, the ratio between the Et+ signal detected in the salivary glands and the epidermis was generally independent of the concentration 4 hours post-injection. Moreover, the transport activity of the different variants gave similar results at high and low Et+ concentrations (Table 1a, 1b). We also monitored the uptake efficiency of high and low concentrations of Et+ in dependence of time (Table 1a, 1b). The transport activity of the different variants was stable regardless of the different incubation times of 1 or 4 hours. These experiments demonstrate that the Drosophila test system is highly flexible, providing stable results independent of substrate concentration and incubation time.
To further elucidate our method as a test system for drug-drug interaction (DDI)
experiments, we analyzed the effects of cimetidine, a known inhibitor of hOCT1
and hOCT2 [40], on hOCT activity in fly embryos. The inhibitory effect of
cimetidine increased with increasing concentrations (Fig. 4D). An IC50 value
of 14.8
In addition, injection of prazosin (4 mM), a potent inhibitor of hOCT1
(IC50 2 µM) but not of hOCT2 (IC50
The transport of a substrate from the basal side of an epithelial cell to its apical side requires the concerted activity of a transporter in the basal plasma membrane for substrate uptake and a transporter in the apical plasma membrane for substrate efflux. The basally localized hOCT1 and the apically localized hABCB1 are one such pair of transporters needed for the transport of a number of cations, including drugs, across epithelia [6]. Thus, we aimed to analyze the functionality of hABCB1 in order to reconstitute the complete hOCT1/hABCB1 transport system in D. melanogaster. First, we verified that hABCB1 is expressed in the salivary gland cells of humanized hABCB1 embryos by immunodetection and quantitative polymerase chain reaction (qPCR) (Supplementary Fig. 4). The function of hOCT1 and hABCB1 was evaluated by injecting EtBr or rhodamine 123, which are fluorescent substrates not only of hOCT1 but also of hABCB1 [38, 42, 43], into the embryos expressing both genes in the salivary glands. Et+ or rhodamine 123 accumulated within the cells of the salivary glands, but not in the lumen. Similarly, when both transporters were expressed in the tracheal system, Et+ was only taken up into the cells of the tracheae but not excreted into the lumen (Supplementary Fig. 4). Based on this lack of excretion into the lumen of the salivary glands or tracheae we conclude that the hOCT1/hABCB1 humanized flies cannot be used for our goal to investigate the vectorial transport of fluorescent organic cations and their interference by drugs. It was beyond the scope of this study to elucidate the underlying mechanisms of this apparent non-functionality, but it will be of interest to examine this issue in future studies. One possible reason for the apparent non-functionality may include predominant intracellular localization of hABCB1, which could be investigated, e.g., by high-resolution confocal imaging. Another reason maybe an altered glycosylation in Drosophila as compared to humans. Although some types of N-glycan structures are conserved between Drosophila and humans and even though they share some similar substrate specificities, there are only half the number of glycosyltransferases in Drosophila compared to humans [44]. Therefore, when hABCB1 is translated in Drosophila embryos, the N-glycosylation process may show differences from that in humans. There is evidence from rat hepatocytes, that only a fully N-glycosylated Abcb1 is associated with functional activity [45]. The apparent non-functionality of hABCB1 in Drosophila appears to be a limitation specific to this transporter, because other human ABC transporters such as human ATP-binding cassette transporter B9 (ABCB9) [46] and ATP-binding cassette transporter C1 (ABCC1) [47] were functionally expressed in the Drosophila cell line S2 and in Drosophila neurons, respectively.
We have shown that organic cation uptake transporters are functional in fruit fly embryos. We next sought to analyze the function of human organic anion transporters (OATPs) in flies. We chose hOATP1B1, hOATP1B3, and hOATP2B1 for analysis because they are clinically relevant transport proteins that mediate the uptake of various drugs (e.g., statins, antibiotics) from blood into hepatocytes. Their importance has been highlighted in several reviews [6, 7], which reference also additional studies. These transporters are therefore also recommended by the FDA and EMA for pre-clinical testing. We generated flies harboring the cDNA of the respective transporters downstream of the UAS promoter and verified their expression in embryos by qPCR (Supplementary Fig. 5). However, specific antibodies against hOATP1B1 (Supplementary Fig. 6) and hOATP1B3 (Supplementary Figs. 7,8) failed to recognize the respective proteins in the salivary glands or any other tissue tested. In contrast, localization of hOATP2B1 was successful. The expression of hOATP2B1 in different tissues was detected using a specific antibody (Supplementary Fig. 9). However, the hOATP2B1 signal was not detected at the plasma membrane but inside the cells.
Although the human OATP proteins could not be immunolocalized in the plasma membrane of salivary gland cells, tracheal cells or oenocytes, the transporters may nonetheless be functionally expressed below the limit of immunodetection. Therefore, the function of hOATP1B1, hOATP1B3, and hOATP2B1 was assessed by injection of the fluorescent OATP substrates MDBF, DBF, and CLF [48] into embryos expressing hOATP1B1 [48], hOATP1B3, and hOATP2B1. For visualization of the target tissue, each insertion was recombined to UAS-RFP. Four hours after injection, salivary glands and tracheae were observed by confocal microscopy. The dye signals were observed in the salivary gland (Supplementary Figs. 10–12) and tracheal lumina (Supplementary Figs. 13–15) in control and humanized embryos. No signal was seen in oenocytes (Supplementary Figs. 16–18). Based on these results, we conclude that fluorescent hOATP substrates were taken up into the lumen of the salivary glands and tracheae of embryos independently of hOATP1B1, hOATP1B3, and hOATP2B1 activity. All three hOATPs were apparently also not active in oenocytes.
Taken together, in contrast to organic cation transporters and contrary to
expectations, the three hOATPs studied lack a detectable functional expression in
D. melanogaster under our current conditions. One may speculate that
transporter functionality might be improved by several strategies in future
studies, although this might be challenging because knowledge on mechanisms of
hOATPs trafficking is incomplete. So far, several mechanisms have been identified
as being necessary for membrane trafficking, e.g., of hOATP1B1, hOATP1B3 and
hOATP2B1: interaction of hOATP1B1, but not of hOATP1B3, with PDZ domain
containing 1 (PDZK1) [49], amino acid residues in transmembrane domain 11 of
hOATP1B1 [50], N-glycosylation of hOATP1B1 [51], the N-terminal region of
hOATP1B3 [52] and protein kinase C mediated, clathrin-dependent internalization
[53]. Potential strategies for improving transporter functionality may
consequently include (i) co-expression of human PDZK1 to provide the proper
interacting protein, (ii) generation of fused lobes (fdl)
knockout flies, because the
In summary, we have developed a novel system that utilizes Drosophila embryos for the expression of functional human membrane transporters. We successfully demonstrated that the data on inhibitors and clinically-relevant genetic variants of hOCT1 and hOCT2 correlate with those obtained in mammalian expression systems. However, there is the limitation that human membrane transporters do not appear to be functionally expressed in the fruit fly in general, which needs to be studied for each transporter individually. Nevertheless, the fruit fly system can be extended for those transporters and their clinically-relevant genetic variants that can be functionally expressed. It can be speculated that differences between Drosophila and humans in drug distribution may exist and may arise from the fruit fly’s embryos open circulatory system and significantly smaller body size, which likely result in different drug distribution patterns and faster drug distribution compared to humans. Therefore, Drosophila is best suited as a preclinical screening tool for primary small-molecule screening or post-screening validation to narrow down a large set of drug candidate drugs to a much smaller pool of lead compounds as suggested by Pandey & Nichols [13]. The incorporation of Drosophila in high-throughput screening is expected to accelerate the identification of effective lead compounds because it enables screening within a whole-animal, systems level context. The system can be significantly enhanced and accelerated through the integration of microfluidic technologies that can support medium- or high-throughput analyses. Specifically, the use of microfluidic arrays will allow for automated, parallelized injection protocols and high-resolution, in vivo fluorescence microscopy [56, 57, 58].
As additional innovation and advantage compared to currently used mammalian cell systems, we hypothesize that pharmacodynamic consequences of transporter-mediated DDIs can be assessed. Our work constitutes the basis for future studies, in which human transporters—such as hOCT1 and hOCT2—can be expressed specifically, e.g., in the brain and flies are fed with toxic agents (e.g., cisplatin) and simultaneously with drugs of interest that may counteract toxicity because they inhibit hOCT function. Due to low costs to generate and maintain humanized Drosophila stocks and the lack of ethical constraints, this system opens a new avenue of pre-clinical evaluation of human membrane transporters to replace currently existing models.
The datasets used or analyzed during the current study are available from the corresponding author on reasonable request.
MS, ATN, BM, YW and ES designed research. YW, JY and SUJ performed research. YW, JY, SUJ, ES, BM, MS, and ATN analyzed data. YW, JY, BM, MS and ATN wrote the paper. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
The authors thank K.M. Giacomini for providing the HEK cell lines and for critical reading of the manuscript. In addition, we thank Silvia Hübner and Nicole Gehring for excellent technical assistance.
This work was supported by the Robert-Bosch Stiftung, Stuttgart, Germany, and the Interfaculty Centre for Pharmacogenomics and Pharma Research (ICEPHA) Grant Tuebingen–Stuttgart, Germany.
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBL42817.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.