





 , Md Wasim Bari 2, Satoshi Kishigami 1,2,3,4,*
, Md Wasim Bari 2, Satoshi Kishigami 1,2,3,4,*
1 Department of Integrated Applied Life Science, Integrated Graduate School of Medicine, Engineering, and Agricultural Sciences, University of Yamanashi, 400-8510 Yamanashi, Japan
2 Faculty of Life and Environmental Sciences, University of Yamanashi, 400-8510 Yamanashi, Japan
3 Center for Advanced Assisted Reproductive Technologies, University of Yamanashi, 400-8510 Yamanashi, Japan
4 Advanced Biotechnology Center, University of Yamanashi, 400-8510 Yamanashi, Japan
Abstract
Lysosomes serve not only in the degradation of cellular components but also as calcium (Ca2+) stores. In this study, we investigated the effects of trans-Ned19, an inhibitor of lysosomal calcium channels known to block two-pore channels (TPCs), on fertilization and oocyte activation in mice.
Pronuclear formation was assessed via Hoechst 33342 staining, cortical granule release was evaluated using Lens culinaris agglutinin-fluorescein isothiocyanate (LCA-FITC) staining, intracellular Ca2+ levels were monitored with Cal-520 AM, and sperm motility was analyzed using a sperm motility analysis system (SMAS).
In strontium (Sr2+)-induced oocyte activation, trans-Ned19 significantly reduced pronuclear formation at 8 h post-activation. Cortical granule release and Ca2+ oscillations were also markedly suppressed. In contrast, during in vitro fertilization (IVF), trans-Ned19 treatment significantly decreased the fertilization rate; however, pronuclear formation and cortical granule release remained comparable to controls in fertilized embryos. Notably, when IVF was performed using zona pellucida-free oocytes, the fertilization rate in the trans-Ned19 group was similar to that of the controls. However, a significant increase in polyspermy was observed. Furthermore, trans-Ned19 significantly impaired sperm motility parameters, including straight-line velocity, curvilinear velocity, and average path velocity.
These findings suggest that lysosomal TPCs are essential for both normal fertilization and artificial oocyte activation in mice.
Keywords
- calcium channels
- in vitro fertilization
- oocytes
In mammals, mature oocytes are transformed into zygotes through fertilization
with sperm. This process, known as oocyte activation, begins with a series of
intracellular calcium (Ca2+) oscillations, followed by events such as
cortical granule release and pronuclear formation, ultimately leading to
embryonic development [1, 2]. During physiological oocyte activation,
sperm-derived phospholipase C zeta (PLC
Lysosomes not only function as degradative organelles for intra- and extracellular components but also serve as intracellular Ca2+ stores [9, 10]. They interact with other calcium-regulating organelles, such as the endoplasmic reticulum and mitochondria, thereby modulating cytoplasmic Ca2+ signaling [11, 12]. Lysosomal membranes harbor calcium -permeable two-pore channels (TPCs), which play a key role in regulating intracellular Ca2+ dynamics [13]. The animal TPC family comprises three subtypes: TPC1, TPC2, and TPC3 [14]. In humans and mice, only TPC1 and TPC2 are expressed, with distinct subcellular localizations-TPC1 is distributed across various endolysosomal compartments, whereas TPC2 is predominantly localized to late endosomes and lysosomes [15]. TPCs are activated by nicotinic acid adenine dinucleotide phosphate (NAADP) [16]. And the NAADP-TPC pathway has been implicated in fertilization. For example, TPC knockdown in sea star oocytes disrupts embryonic development due to impaired Ca2+ signaling [17]. In mice, NAADP-mediated Ca2+ signaling via TPC1 regulates the acrosome reaction in sperm, and the presence of NAADP synthase has also been reported in human sperm [18, 19, 20]. Nonetheless, the role of TPCs in mammalian fertilization and oocyte activation remains largely unexplored.
Trans-Ned19 is a selective antagonist of NAADP and effectively inhibits NAADP-induced Ca2+ release [21]. Its inhibitory activity has been characterized across a broad concentration range, from 1 µM to 300 µM [21, 22].
In this study, we investigated the potential involvement of TPC-mediated Ca2+ signaling in fertilization and oocyte activation using the NAADP antagonist trans-Ned19.
Institute of Cancer Research (ICR) female mice aged 8 weeks or older and ICR male mice aged 10 weeks or older were used. Mice were purchased from SLC (Japan SLC, Inc., Shizuoka, Japan) and bred in-house. They were maintained in a specific pathogen-free facility under controlled conditions (25 °C, 50% humidity, 14 h light/10 h dark cycle). Standard pelleted food and water were provided ad libitum. All animal procedures were approved by the Animal Experimentation Committee (protocol number A4-10) at the University of Yamanashi, Japan, and were conducted in accordance with ethical guidelines. Animal handling and experimentation adhered to the 3Rs principle and followed the Guidelines for Proper Conduct of Animal Experiments (Science Council of Japan).
Female mice aged 8 weeks or older were administered 7.5 IU of pregnant mare serum gonadotropin (Aska Pharmaceutical, Tokyo, Japan) via intraperitoneal injection, followed 48 h later by 7.5 IU of human chorionic gonadotropin (hCG; Aska Pharmaceutical). Cumulus-oocyte complexes (COCs) were collected from the oviducts approximately 16 h after the hCG injection. All mice used in this study were euthanized by cervical dislocation.
For IVF, sperm collected from the caudal epididymis of male mice aged 10 weeks
or older were pre-incubated in a 100 µL droplet of human tubal fluid
(HTF) medium at 37 °C and 5% (v/v) CO2 for 1 h to induce
capacitation (Supplementary Table 1). Subsequently, COCs and capacitated
sperm were co-incubated in a 100 µL droplet of HTF medium at a final
sperm concentration of 1.0
For Sr2+-induced oocyte activation, COCs were transferred to a 100 µL droplet of HEPES-buffered Chatot-Ziomek-Bavister (CZB) medium containing 0.1% (w/v) bovine testicular hyaluronidase (Sigma-Aldrich, St. Louis, MO, USA) and incubated for 20 min (Supplementary Table 2). They were then moved to a 15 µL droplet of CZB medium and cultured for 1 h. Cumulus-free oocytes were further incubated in a 15 µL droplet of Ca2+-free CZB medium containing 5 mM SrCl2 (Fujifilm Wako Pure Chemical Corporation, Osaka, Japan) at 37 °C under 5% CO2 for 8 h (Supplementary Table 3). When trans-Ned19 (Sigma-Aldrich) was used, it was added to both fertilization and activation media at a final concentration of 300 µM.
To observe pronuclear formation, activated oocytes or oocytes after IVF were incubated in CZB medium containing 1 µg/mL Hoechst 33342 (Sigma-Aldrich) for more than 10 min and observed using a fluorescence microscope (BZ-X710 or BZ-X800, Keyence, Osaka, Japan).
Oocytes were cultured in CZB medium containing 5 µg/mL Lens culinaris agglutinin-fluorescein isothiocyanate (LCA-FITC; Vector Laboratories, Newark, CA, USA) for at least 10 min following 2 h of Sr2+-induced activation or insemination. They were observed at room temperature using a fluorescence microscope. Cortical granule release was evaluated visually based on fluorescent images.
Before Sr2+-induced activation, oocytes were pre-incubated in CZB medium containing 5 µM Cal-520 AM (AAT Bioquest, Pleasanton, CA, USA) for 1 h. Time-lapse imaging was then performed using a fluorescence microscope. When trans-Ned19 was used, it was added to the activation medium at a final concentration of 300 µM. During imaging, oocytes were maintained in an incubation chamber with 5% CO2, a top heater set to 45 °C and a bath heater set to 41 °C. Imaging was conducted at 30-second intervals for 3 h. Calcium signals were analyzed by measuring the average pixel value in the cytoplasmic region using Python 3.11.2 (Python Software Foundation, https://www.python.org/) and the OpenCV image processing library (https://opencv.org/).
After 1 h of capacitation in a 100 µL droplet of HTF medium, sperm were further incubated in HTF medium containing 300 µM trans-Ned19 for 2 h. Sperm motility was assessed at room temperature using a sperm motility analysis system (SMAS; DITECT, Tokyo, Japan) with its built-in analysis software.
Statistical analyses were performed using the chi-square test or Fisher’s exact
test, depending on the expected frequencies in 2
To examine the effect of trans-Ned19 on pronuclear formation, 300 µM trans-Ned19 was added to the insemination medium for IVF and the activation medium for Sr2+-induced oocyte activation. Pronuclei were assessed by Hoechst 33342 staining 8 h after the onset of insemination or Sr2+-induced activation. As a result, trans-Ned19 significantly decreased the fertilization rate during IVF, although pronuclear formation was observed in all fertilized embryos except one (Fig. 1A and Table 1). In contrast, during Sr2+-induced oocyte activation, trans-Ned19 treatment significantly reduced the pronuclear formation rate. However, when trans-Ned19 was added 2 hours after the onset of Sr2+-induced activation, pronuclei were observed in nearly all embryos (Fig. 1B and Table 1). Interestingly, the incidence of small and incompletely expanded pronuclei—classified as “1PN like”—was significantly increased (Fig. 1C and Table 1). These findings suggest that trans-Ned19 does not inhibit sperm-induced oocyte activation but rather interferes with fertilization. In contrast, trans-Ned19 strongly impairs Sr2+-induced oocyte activation.
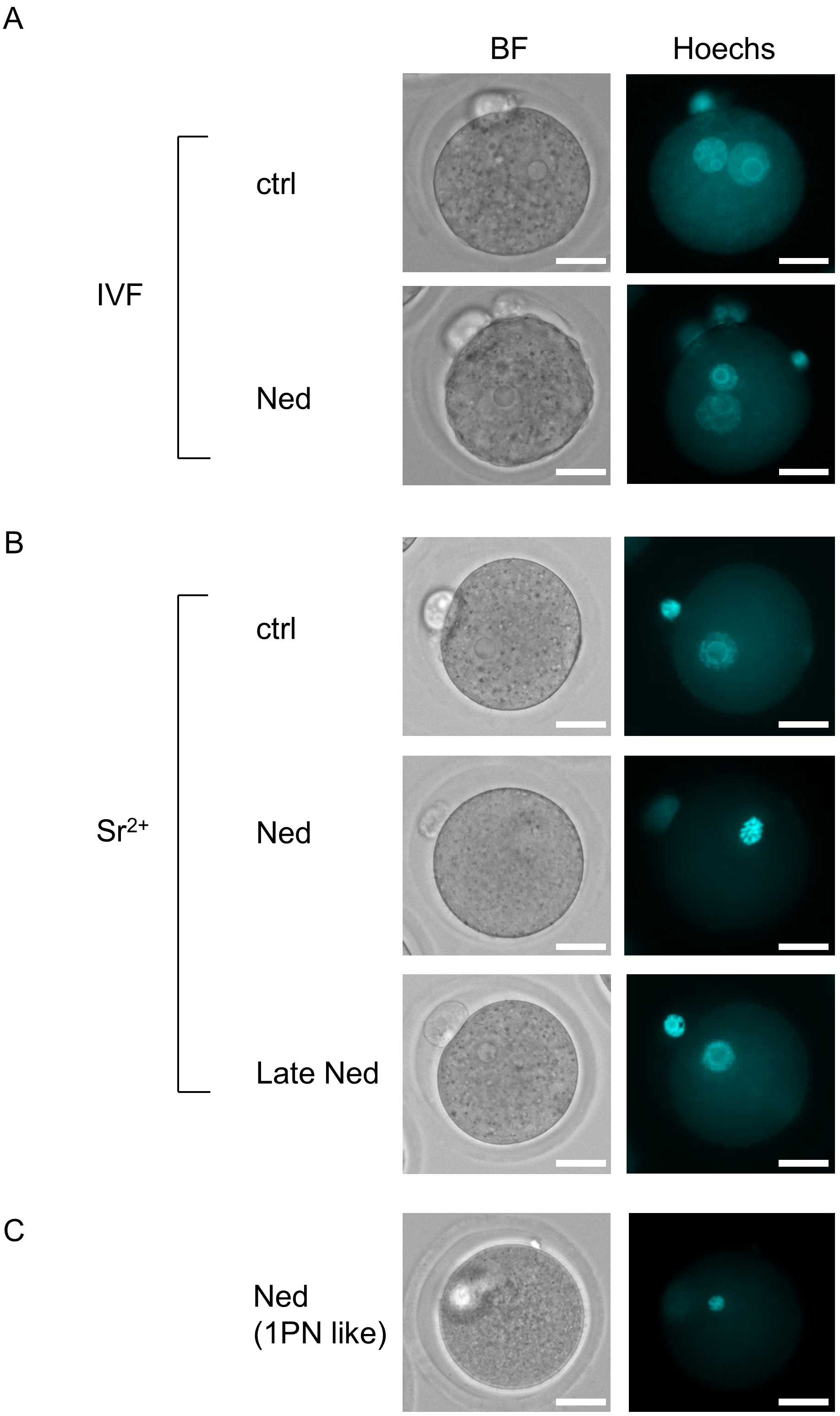 Fig. 1.
Fig. 1.
Effects of trans-Ned19 on pronuclear formation after in vitro fertilization (IVF) and strontium (Sr2+)-induced activation. (A) Representative images showing pronuclear formation in trans-Ned19-untreated (control) and trans-Ned19-treated groups 8 h after IVF. Ctrl, trans-Ned19-untreated group; Ned, group treated with trans-Ned19 from the start of insemination. (B) Representative images of oocytes 8 h after Sr2+-induced activation. Ctrl, trans-Ned19-untreated group; Ned, group treated with trans-Ned19 from the start of activation; Late Ned, group treated with trans-Ned19 starting 2 h after the onset of Sr2+-induced activation. (C) “1PN like” embryo in the trans-Ned19-treated group after Sr2+- induced activation. BF, bright-field image; Hoechst, fluorescence image stained with Hoechst 33342. Scale bar: 25 µm.
| No. of oocyte | MII | 0PN with a spem | PN formed | |||||||
| Total (%) | 1PN (%) | 1PN like (%) | 2PN (%) | 3PN |
Others | |||||
| IVF | ctrl | 51 | 6 (12)a | 0 (0) | 45 (88) | 3 (7)a | 0 (0) | 41 (91) | 1 (2) | 0 (0) |
| Ned | 90 | 24 (27)b | 1 (1) | 65 (72) | 0 (0)b | 0 (0) | 62 (95) | 3 (5) | 0 (0) | |
| Sr2+ | ctrl | 49 | 0 (0)a | - | 49 (100) | 47 (96)a | 0 (0)a | 2 (4) | 0 (0) | 0 (0) |
| Ned | 49 | 17 (35)b | - | 32 (65) | 22 (69)b | 7 (22)b | 1 (3) | 0 (0) | 2 (6) | |
| Late Ned | 50 | 0 (0)a | - | 50 (100) | 49 (98)a | 0 (0)a | 0 (0) | 0 (0) | 1 (2) | |
MⅡ, metaphase II oocyte; 0PN, embryo with sperm but without expanded pronucleus;
1PN, embryo with one pronucleus; 1PN like, embryo showing fluorescence resembling
a single unexpanded pronucleus; 2PN, embryo with two pronuclei; 3PN
Next, we examined cortical granule signals 2 h after the onset of insemination or activation. As a result, cortical granule signals were observed in all fertilized embryos during IVF, regardless of trans-Ned19 treatment (Fig. 2A and Table 2). In contrast, in Sr2+-induced oocyte activation, embryos lacking cortical granule signals were observed in the trans-Ned19-treated group (Fig. 2B and Table 2). Additionally, there was a significant increase in embryos displaying small granule signals, which were distinct from the prominent cortical granule signals observed in the control group (Fig. 2B and Table 2). These small granule signals were rarely detected in the control group. These results indicate that trans-Ned19 does not affect cortical granule release during sperm-induced oocyte activation but suppresses it during Sr2+-induced oocyte activation.
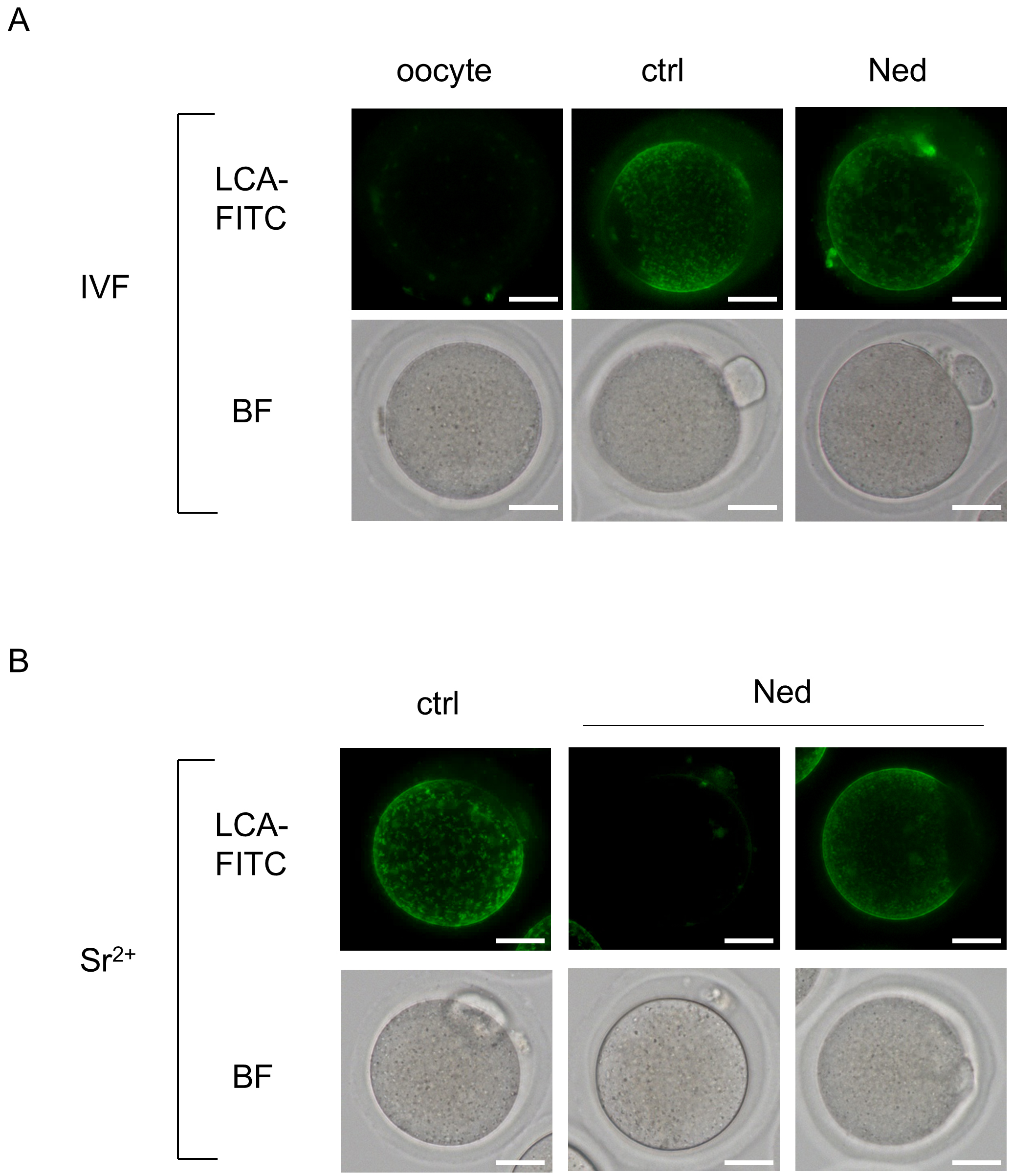 Fig. 2.
Fig. 2.
Lens culinaris agglutinin-fluorescein isothiocyanate (LCA-FITC) staining results 2 h after IVF and Sr2+-induced activation. (A) Results of IVF. Oocyte, metaphase II oocyte; Ctrl, trans-Ned19-untreated group; Ned, group treated with trans-Ned19 from the start of insemination. (B) Results of Sr2+-induced activation. Oocyte, metaphase II oocyte; ctrl, trans-Ned19-untreated group; Ned, group treated with trans-Ned19 from the start of activation. For trans-Ned19-treated embryos during Sr2+-induced activation, the left image shows an embryo without cortical granule release, and the right image shows an embryo with small granule signals. BF, bright field image. Scale bar: 25 µm.
| No. of oocyte | Fertilized (%) | LCA-FITC positive | ||||
| Total (%) | Normal particles (%) | Small particles (%) | ||||
| IVF | ctrl | 46 | 38 (83) | 38 (100) | 38 (100) | 0 (0) |
| Ned | 67 | 52 (78) | 52 (100) | 52 (100) | 0 (0) | |
| Sr2+ | ctrl | 48 | - | 48 (100)a | 47 (98)a | 1 (2)a |
| Ned | 49 | - | 42 (86)b | 26 (62)b | 16 (38)b | |
Normal particles: normal lens culinaris agglutinin-fluorescein
isothiocyanate (LCA-FITC) signal pattern; Small particles: small LCA-FITC signal
pattern. In IVF, the percentage of fertilization was calculated based on the
total number of oocytes tested (No. of oocyte), and the percentages of Normal
particles and Small particles were calculated based on the number of fertilized
embryos (Fertilized). In the case of Sr2+-induced activation, the
percentages of Normal particles and Small particles were calculated based on the
total number of oocytes tested (No. of oocyte). For both IVF and
Sr2+-induced activation, the control data (ctrl) partially overlap with
those reported in our previous study. a, b Significant differences
(Chi-square test or Fisher’s exact test, p
Trans-Ned19 inhibited cortical granule release, an event that occurs at an early stage of Sr2+-induced oocyte activation. To further investigate whether Ca2+ oscillations—which occur earlier than cortical granule release and initiate oocyte activation—were induced, we monitored cytoplasmic Ca2+ levels using the fluorescent Ca2+ indicator Cal-520 AM. As a result, the number of embryos exhibiting an increase in cytoplasmic Ca2+ signals following Sr2+-induced activation was significantly reduced in the trans-Ned19-treated group compared to the control group (Fig. 3). These findings indicate that trans-Ned19 inhibits the initiation step of Sr2+-induced oocyte activation by suppressing Ca2+ oscillations.
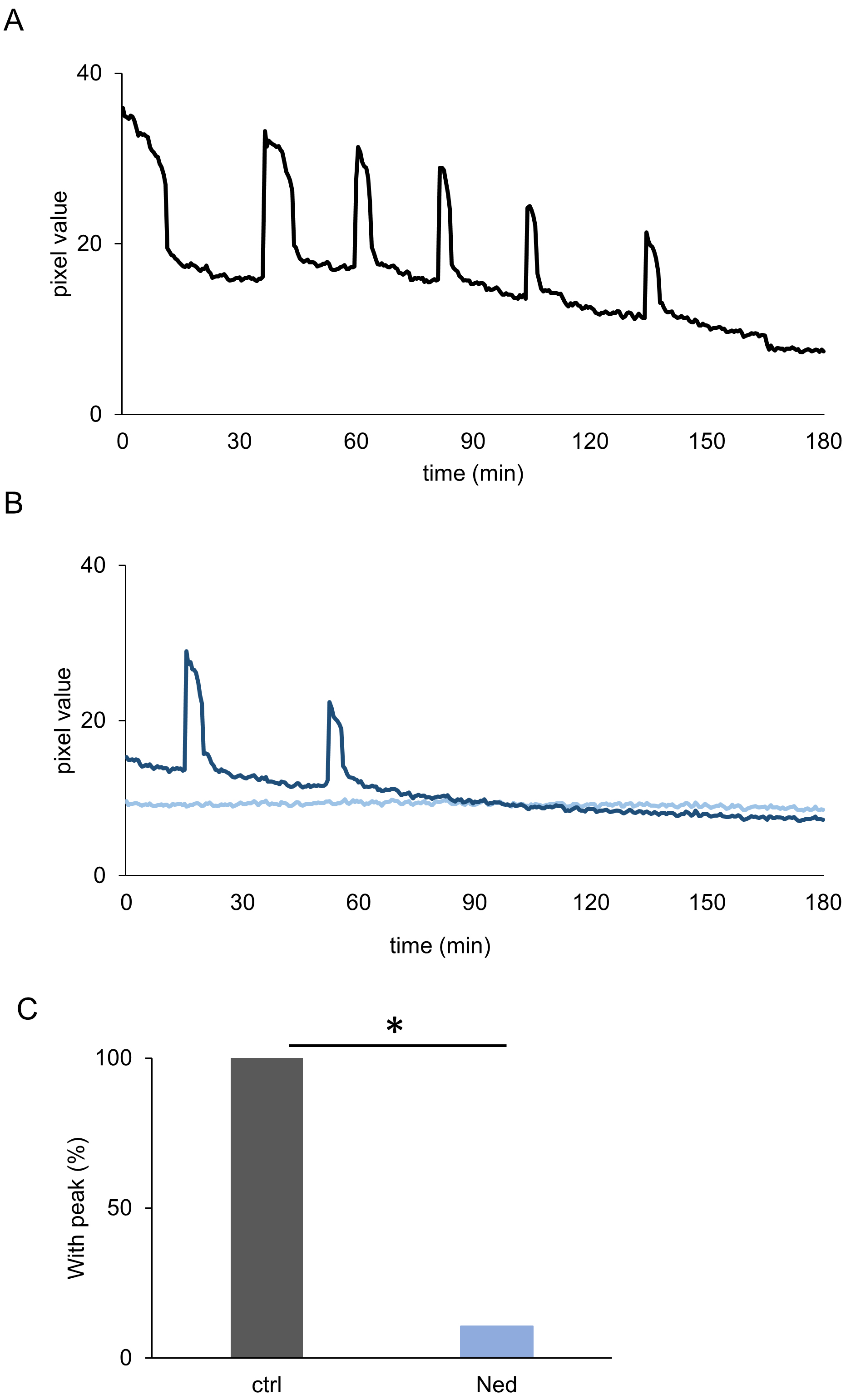 Fig. 3.
Fig. 3.
Cytoplasmic calcium (Ca2+) signal imaging results up to 3 h
after the start of Sr2+-induced activation. (A) Changes in pixel values
within the embryonic cytoplasmic region in the trans-Ned19-untreated group. (B)
Changes in pixel values within the embryonic cytoplasmic region in the
trans-Ned19-treated group. In both (A,B), the horizontal axis represents time
(min) from the start of imaging, and the vertical axis represents pixel value.
The graphs show results from representative embryos. (C) Percentage of embryos
showing an increase in pixel values in the trans-Ned19-untreated group (ctrl) and
the trans-Ned19-treated group (Ned). The control data (ctrl) overlap with those
reported in our previous study. *Significant differences (Chi-square test,
p
Previously, we demonstrated that CQ significantly reduced the fertilization rate
in IVF; however, when using oocytes with the zona pellucida removed, the
fertilization rate was restored to the level of the control group [5]. Similarly,
in this study, we found that while trans-Ned19 treatment did not significantly
affect the pronuclear formation rate, it tended to reduce the fertilization rate
in IVF. To investigate this further, IVF was performed using zona-free oocytes,
and pronuclear formation was assessed 8 h later. In the trans-Ned19-treated
group, the fertilization rate was comparable to that of the control group,
indicating the removal of the zona pellucida restored fertilization competence.
Expanded pronuclei were also observed, similar to those in the control group
(Fig. 4 and Table 3). Notably, trans-Ned19 treatment significantly increased the
number of embryos with more than three pronuclei (3PN
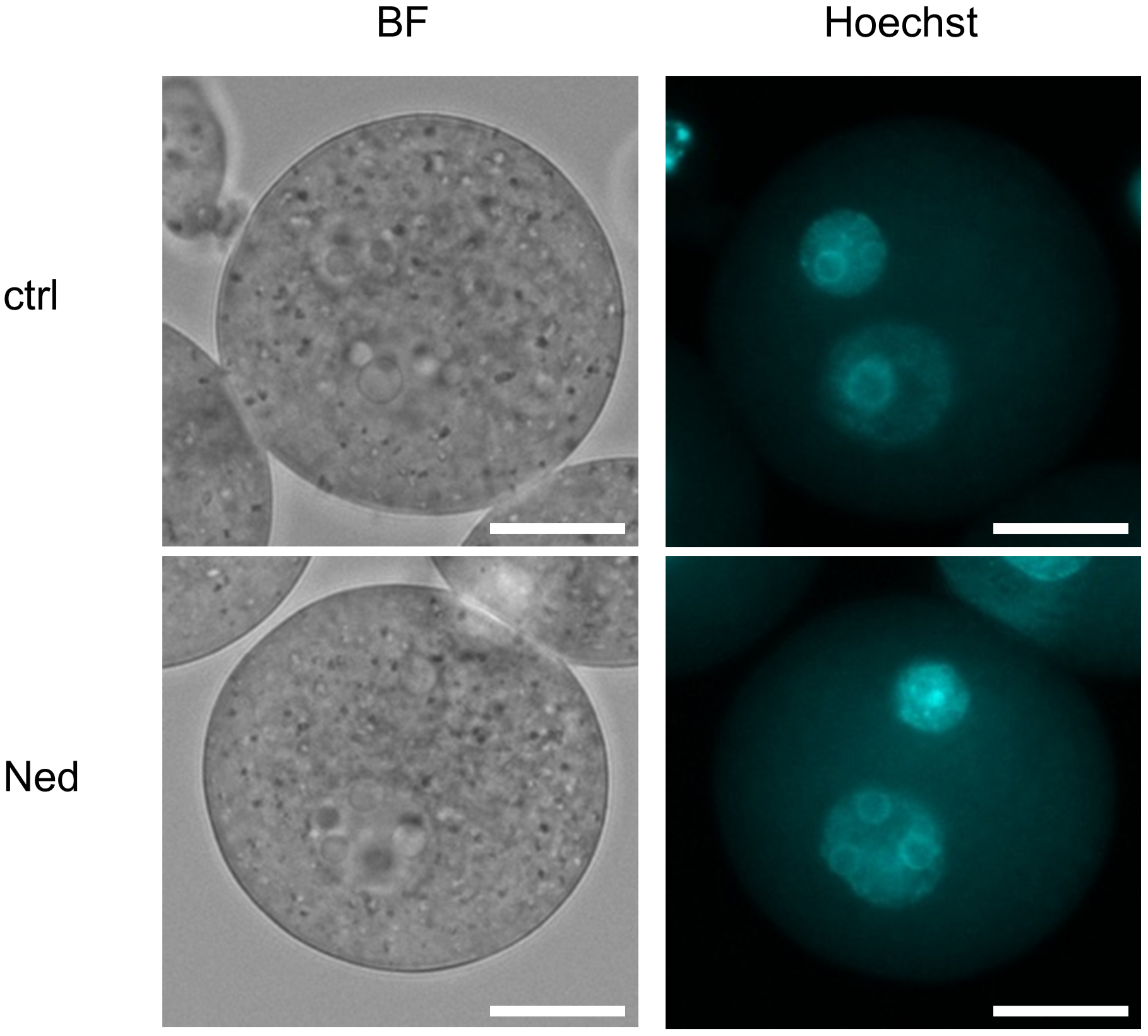 Fig. 4.
Fig. 4.
Hoechst staining after 8 h after insemination using zona-free oocytes. Representative images showing pronuclear formation in the trans-Ned19-untreated and trans-Ned19-treated groups 8 h after zona-free IVF. Ctrl, trans-Ned19-untreated group; Ned, group treated with trans-Ned19 from the start of insemination; BF, bright-field image; Hoechst, fluorescent image stained with Hoechst 33342. Scale bar: 25 µm.
| No. of oocyte | MII | 1PN | 2PN | 3PN | |
| ctrl | 47 | 0 (0) | 3 (6) | 37 (79)a | 7 (15)a |
| Ned | 56 | 1 (2) | 3 (5) | 25 (45)b | 27 (48)b |
MII, metaphase II oocyte; 1PN, embryo with one pronucleus; 2PN, embryo with two
pronuclei; 3PN
Since trans-Ned19 reduced the fertilization rate by affecting sperm penetration through the zona pellucida, we examined whether trans-Ned19 treatment impairs sperm motility. Sperm were treated with trans-Ned19 for 2 h after capacitation, and their motility was subsequently assessed, as fertilization is considered to occur within 1–2 h after insemination under IVF conditions. As a result, trans-Ned19 treatment significantly reduced the proportion of sperm exhibiting high motility, as indicated by decreased straight-line velocity, curvilinear velocity, and average path velocity (Fig. 5). In addition, the proportion of immotile sperm was significantly increased, with a similar trend observed as early as 1 hour after trans-Ned19 treatment (data not shown). These findings suggest that the impaired passage of sperm through the zona pellucida is partly due to reduced motility caused by trans-Ned19 treatment.
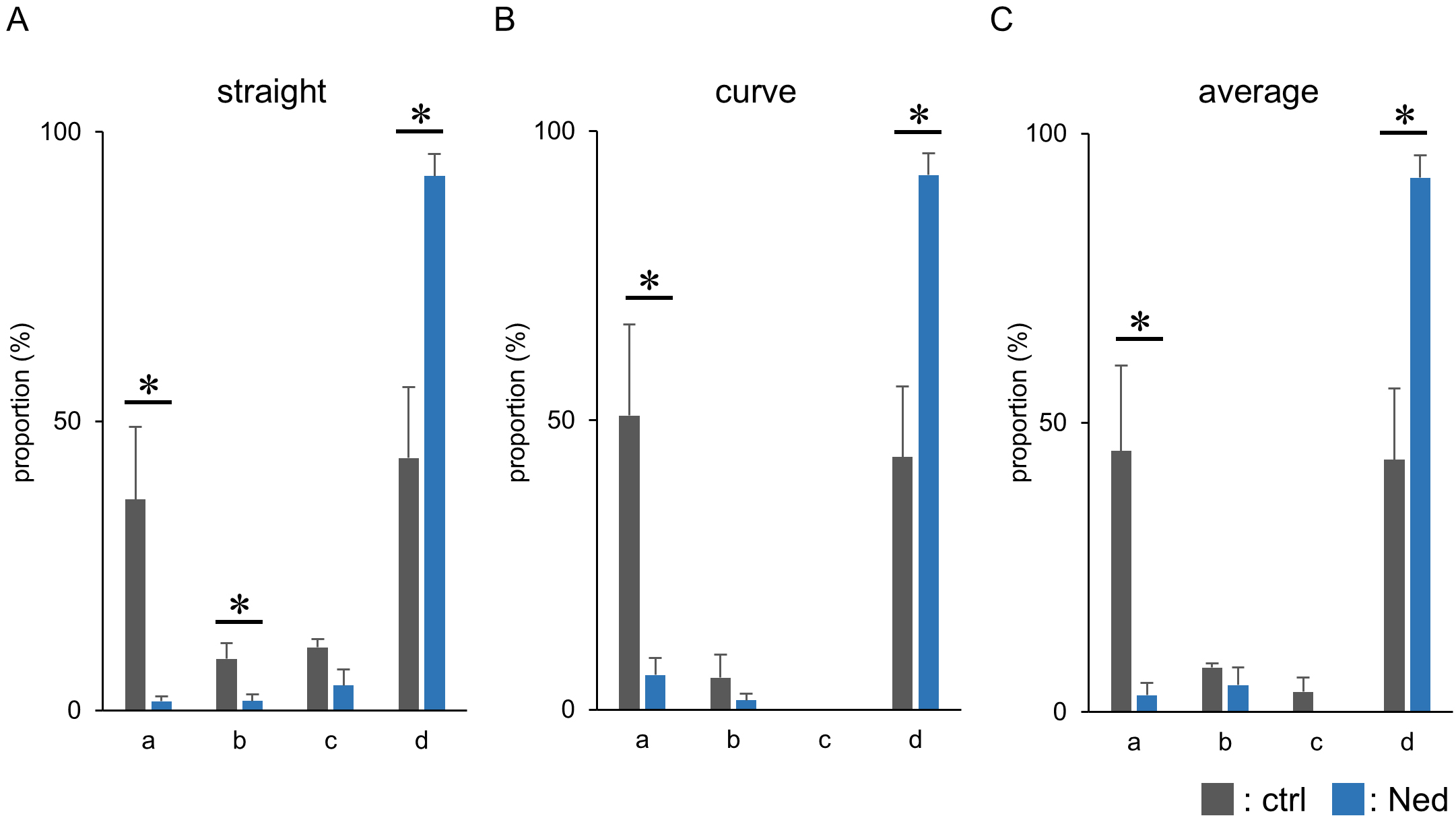 Fig. 5.
Fig. 5.
Sperm motility distribution 2 h after trans-Ned19 treatment.
(A–C) Graphs showing sperm motility characteristics measured 2 h after
trans-Ned19 treatment. (A) Straight-line velocity; (B) curvilinear velocity; (C)
average path velocity. In each graph, the horizontal axis (a–d) indicates
velocity ranges: a:
For oocyte activation, calcium is released from intracellular Ca2+ stores, primarily the endoplasmic reticulum (ER), through the activation of IP3-gated Ca2+ release channels (IP3 receptors) [1]. In animal cells, NAADP, another Ca2+-mobilizing messenger, initiates Ca2+ release from the endolysosomal system via activation of TPCs [13]. However, the involvement of TPCs in oocyte activation remains unknown in mammals. In this study, we demonstrated that inhibition of TPCs by trans-Ned19 had little effect on sperm-induced oocyte activation, whereas it significantly suppressed Sr2+-induced oocyte activation by inhibiting Ca2+ oscillations. These results suggest that the cascade of sperm-induced oocyte activation is distinct from that of Sr2+-induced activation, consistent with our previous findings [6].
Although both TPC1 and TPC2 are present in mice [15], at least TPC1, which is localized to lysosomes and endosome, is expressed in mouse oocytes [23]. In sperm, TPC1 has been reported to localize to the acrosome and nuclear redundant vesicles [18]. Trans-Ned19, a selective and cell-permeant NAADP antagonist, is widely used to study the functional roles of TPCs [24]. In smooth muscle cells, 100 µM trans-NED 19 reduces TPC1-mediated Ca2+ elevation by nearly 60% [25]. In the present study, 300 µM trans-NED19 significantly suppressed Sr2+-induced Ca2+ oscillations, leading to a failure of normal pronuclear formation in approximately 50% of embryos. In mice, Sr2+ influx—which induces Ca2+ oscillations and triggers oocyte activation—occurs via transient receptor potential vanilloid 3 (TRPV3) channels [26]. Therefore, it is reasonable to propose that increased intracellular Sr2+ may induce oocyte activation via Ca2+ release from lysosomes, likely through TPCs, either directly or indirectly. However, even under trans-Ned19 treatment, expanded pronuclei were observed in approximately 50% of embryos, suggesting that pronuclear formation during Sr2+-induced activation is not solely dependent on TPC function. Consistently, it has been suggested that Sr2+ can sensitize IP3 receptors and promote Ca2+ oscillations [5]. Furthermore, when trans-Ned19 was added 2 h after the start of Sr2+-induced activation, it did not affect pronuclear formation. In mice, pronuclei are typically formed approximately 3 h after fertilization, suggesting that TPCs are not involved in the later stages of pronuclear formation [27].
In contrast, in IVF embryos, expanded pronuclei were observed in nearly all fertilized embryos, even under trans-Ned19 treatment. This suggests that lysosomal TPCs are not essential for sperm-induced oocyte activation. These findings support the notion that TPC1-mediated Ca2+ release contributes to Sr2+-induced oocyte activation, whereas sperm-induced oocyte activation likely proceeds through a distinct Ca2+ signaling pathway independent of TPC1. Together, our results highlight mechanistic differences between artificial and physiological modes of oocyte activation and underscore the selective involvement of TPCs in non-physiological activation pathways.
Although trans-Ned19 treatment significantly reduced the fertilization rate in IVF, the rate became comparable to that in the control group after removal of the zona pellucida. Furthermore, trans-Ned19 treatment significantly decreased sperm motility, suggesting that TPCs in sperm may be required for maintaining normal motility. While the role of TPCs in sperm motility remains to be fully elucidated, Ca2+ signaling is known to regulate sperm motility, and inhibition of TPC-derived Ca2+ signaling by trans-Ned19 may contribute to the observed reduction [28]. In addition, TPCs have been reported to be involved in the regulating the acrosome reaction. Inhibition of TPCs by trans-Ned19 has been reported to impair the acrosome reaction in sperm, and a recent study suggests that this may occur via TPC-mediated Ca2+ signaling, potentially contributing to reduced fertilization rates [18, 19].
Although trans-Ned19 has been widely used as a selective TPC inhibitor, its effects may not be entirely specific and can vary depending on the experimental context. While trans-Ned19 has been employed in numerous studies investigating NAADP-sensitive Ca2+ signaling, its specificity and potential off-target effects may differ depending on cell type, dosage, and exposure conditions. In the present study, we focused on identifying the potential involvement of TPCs in fertilization and oocyte activation using trans-Ned19 as a pharmacological tool. However, we acknowledge the limitation of relying on a single inhibitor at a high concentration and recognize the need for further validation. Future studies incorporating additional pharmacological agents—such as SG-094, cis-Ned19, and TPC agonists (e.g., TPC2-A1-N and TPC2-A1-P)—will be essential to strengthen causal interpretation and expand upon the current findings.
On the other hand, the significantly increased incidence of polyspermy observed in zona-free IVF under trans-Ned19 treatment suggests that TPCs are involved in the mechanisms that prevent polyspermy in oocytes.
Rejection of polyspermy is known to occur at both the zona pellucida and the oocyte membrane, referred to as the “zona block” and “membrane block”, respectively. In IVF using zona-free oocytes, the membrane block is considered the primary barrier to polyspermy [29]. Although this block is thought to be associated with the loss of the oocyte membrane protein IZUMO1 receptor, also known as folate receptor 4 (JUNO) following fertilization [30], the underlying mechanisms remain largely unclear. Since cortical granule release was observed even under trans-Ned19 treatment, it is suggested that TPC involvement in polyspermy prevention occurs mainly at the level of the oocyte membrane. Interestingly, an increase in cytosolic Ca2+ alone is not sufficient to establish the membrane block [18]. In normal fertilization, TPC-mediated activity may be required for the establishment of this membrane block. Future studies should aim to elucidate how TPCs contribute to this process.
Cortical granules undergo Ca2+-dependent exocytosis, releasing their contents into the perivitelline space and modifying the zona pellucida to prevent polyspermy—a process known as the cortical reaction. This secretion mediated by the soluble NSF-attachment protein receptor (SNARE) protein pathway [31]. Trans-Ned19 treatment did not suppress cortical granule release in fertilized embryos, suggesting that TPC activity is not required for the cortical reaction during sperm-induced oocyte activation, consistent with its dispensability in pronuclear formation. In contrast, during Sr2+-induced oocyte activation, trans-Ned19 treatment significantly decreased the number of embryos exhibiting normal granule signals and significantly increased those with small granule signals. This suggests that TPC function is required for normal cortical reactions in Sr2+-induced activation. Even under trans-Ned19 treatment, 53% of embryos exhibited cortical granule release patterns similar to the control group, and the pronuclear formation rate was 47%, suggesting that some embryos successfully completed cortical granule release but failed to form expanded pronuclei. Conversely, embryos with small granule signals may have failed to form pronuclei, indicating a correlation between the granule signal pattern and pronuclear formation. These observations suggest that incomplete activation signals under trans-Ned19 treatment result in abnormal cortical reactions and defective pronuclear formation.
Inhibition of TPCs by trans-Ned19 had minimal impact on sperm-induced oocyte activation during IVF, whereas Sr2+-induced oocyte activation was inhibited from the initiation stage—specifically, at the onset of Ca2+ oscillations—leading to significant reductions in cortical granule release and pronuclear formation. Additionally, trans-Ned19 treatment was associated with a trend toward decreased sperm motility and fertilization rates, which were restored upon removal of the zona pellucida. These effects were similar to those observed with CQ treatment [6], suggesting that lysosome-derived Ca2+ signaling, including TPC-mediated pathways, may be a common target of both agents. More importantly, TPCs may contribute to the prevention of polyspermy, highlighting their role in post-fertilization events. Together, these findings provide new insights into the mechanisms of oocyte activation and hold potential implications for the advancement of assisted reproductive technologies.
CQ, chloroquine; TPC, two-pore channel; NAADP, nicotinic acid adenine dinucleotide phosphate; IVF, in vitro fertilization.
All raw data for this study are available from the corresponding author.
TY and SK designed the research study. TY and MWB performed the experiments. SK provided guidance and support throughout the research. TY analyzed the data. TY and SK wrote the manuscript. All authors contributed to revisions of the manuscript, reviewed and approved the final version, and agreed to be accountable for all aspects of the work.
All the animal experiments were approved by the Animal Experimentation Committee at the University of Yamanashi, Japan, and conducted in accordance with the ethical guidelines (protocol number A4-10). Animal handling and experimentation adhered to the 3Rs principle and followed the Guidelines for Proper Conduct of Animal Experiments issued by the Science Council of Japan.
We gratefully acknowledge discussions and technical support with Dr. H. Harayama in Kobe University, Drs. T. Wakayama, S. Wakayama, D. Ito, and Ms. Y. Kanda at the Advanced Biotechnology Center, and also Ms. S. Furusato at the Center for advanced Assisted Reproductive Technologies, University of Yamanashi as well as all the lab members.
This work was supported by JSPS KAKENHI Grant (Numbers 20K06443 and 24K01937 to SK).
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBL42710.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.