








 , Yan Jiang 1,†, Jiangbo Wan 1, Difan Zhang 1, Fang Huang 1,*
, Yan Jiang 1,†, Jiangbo Wan 1, Difan Zhang 1, Fang Huang 1,* , Siguo Hao 1,*
, Siguo Hao 1,*1 Department of Hematology, Xinhua Hospital Affiliated to Shanghai Jiao Tong University School of Medicine, 200090 Shanghai, China
†These authors contributed equally.
Abstract
Mesenchymal stem cells (MSCs) are one of the effective treatments for acute graft-versus-host disease (aGVHD) following allogeneic hematopoietic stem cell transplantation (allo-HSCT) due to their potent immunoregulatory function. Given the limited quantity and high heterogeneity of freshly isolated MSCs, extensive in vitro expansion is essential for clinical application. Prolonged passaging of MSCs leads to a decline in therapeutic efficacy, with the underlying mechanisms remaining unclear. This study aimed to explore the mechanism and intervention strategies of immunoregulatory dysfunction of MSCs during passaging.
We compared the therapeutic effects of MSCs at early passages (passage 6, P6-MSCs) and later passages (passage 12, P12-MSCs) in a mouse GVHD model. We also analyzed the expression of Gal-3 in MSCs at different passages and its role in macrophage polarization. Additionally, the selective Gal-3 inhibitor TD139 was evaluated for its effects on Gal-3 expression and the immunoregulatory function of MSCs.
Our data showed an inverse correlation between passage number and therapeutic efficacy in MSCs. Late-passage MSCs (P12) exhibited significantly reduced efficacy in alleviating GVHD compared to early-passage MSCs (P6). The expression of Gal-3 was markedly upregulated in late-passage MSCs (P12), and it was found to directly inhibit anti-inflammatory M2-like macrophage polarization. Our research demonstrated that TD139 dose-dependently suppresses Gal-3 expression in MSCs and restores their immunoregulatory function.
Gal-3 contributes to the decline in the immunomodulatory capabilities of MSCs. TD139, a Gal-3 inhibitor, has a potentially positive effect on rescuing the immunoregulation dysfunction of MSCs in late-passage and may represent a novel strategy to enhance the therapeutic potential of late-passage MSCs for GVHD treatment.
Keywords
- mesenchymal stem cells
- galectin-3
- GB-0139 (TD139)
- graft vs host disease
Allogeneic hematopoietic stem cell transplantation (allo-HSCT) is widely utilized for the treatment of various hematologic malignancies, demonstrating significant immunotherapeutic potential [1]. However, it is constrained by severe graft-versus-host-disease (GVHD), which is a life-threatening complication [2]. The conditioning regimen in allo-HSCT activates antigen-presenting cells (APCs)—predominantly macrophages and dendritic cells—that present host antigens to donor T cells and initiate a robust cytokine response. Effector T cells, macrophages and pro-inflammatory cytokines target skin, lungs, gut and liver, leading to organ damage, which is a clinical manifestation of aGVHD [3].
Mesenchymal stem cells (MSCs) represent a unique population of multipotent precursor cells, defined by their capacities for self-renewal and differentiation into a range of cell lineages, including chondrocytes, osteoblasts, adipocytes, cardiomyocytes, and neurons [4, 5]. Furthermore, MSCs exhibit potent immunoregulatory properties, modulating both innate and adaptive immune responses. This immunoregulatory effect is primarily induced by inflammatory cytokines within the inflammatory microenvironment [6, 7]. MSCs have been demonstrated to facilitate the shift of macrophages from the pro-inflammatory M1 phenotype to the anti-inflammatory M2 phenotype, while simultaneously inhibiting macrophage infiltration at inflammatory sites [8]. Additionally, MSCs directly inhibit the activation of cytotoxic CD8+ T cells and the differentiation of T helper 1 (Th1) and Th17 cells [9]. Simultaneously, they facilitate the differentiation and activity of CD4+CD25+FOXP3+ Treg cells [10, 11]. This combined capacity for self-renewal and multilineage differentiation, coupled with their immunoregulatory function, underscores the potential of MSCs as a viable therapeutic modality.
MSCs are isolatable from a range of tissue sources, including bone marrow, adipose tissue, umbilical cord, and peripheral blood [12, 13]. Given the limited abundance of MSCs in these tissues, extensive ex vivo expansion is required to achieve the sufficient cell numbers necessary for in vivo application [14]. However, multiple studies have shown that MSCs may undergo a decline in their immunosuppressive and regenerative capacities after extended passages [15, 16, 17, 18].
Galectin-3 (Gal-3, gene name: Lgals3), a
Selective galectin inhibitors (TAZTDG, TD139, GB0139) modulate multiple
pathophysiological processes, demonstrating significant potential as therapeutic
candidates [26]. Among them, TD139 is a small-molecule inhibitor that selectively
targets Gal3 by disrupting its
Macrophages constitute a vital element of the innate immune system and mediate critical functions in both inflammatory responses and host defense [31]. In response to differing microenvironments, macrophages adopt distinct functional phenotypes: classically activated macrophages (M1-like) and alternatively activated macrophages (M2-like) [32]. M1-like macrophages are activated by lipopolysaccharide (LPS), resulting in the release of pro-inflammatory cytokines. These macrophages primarily mediate inflammatory responses and contribute to tissue damage. In contrast, M2-like macrophages are activated by interleukin-4 (IL-4), triggering the secretion of anti-inflammatory cytokines. These macrophages predominantly participate in anti-inflammatory responses and tissue repair [33].
The precise mechanism underlying the decline in the immunoregulatory effects of MSCs after multiple passages remains to be elucidated and constituted the primary focus of this study. Our results demonstrated that the effect of late-passage MSCs on M2-like macrophage polarization was diminished both in vivo and in vitro. Additionally, Galectin-3 (Gal-3) was significantly upregulated in late-passage MSCs compared to early-passage MSCs. Furthermore, Gal-3 directly inhibited M2-like macrophage polarization. Therefore, we hypothesized that the attenuation of the immunoregulatory function of MSCs was attributed to the overexpression of Gal-3 in late-passage MSCs. Notably, TD139, a selective Gal-3 inhibitor, dose-dependently suppressed Gal-3 expression in MSCs, particularly in late-passage MSCs. More importantly, TD139 enhances the immunoregulatory potential of late-passage MSCs by promoting M2 macrophage polarization. Based on these findings, the combination of TD139 and MSCs may represent a novel therapeutic strategy for GVHD and other diseases.
Fetal bovine serum (FBS), penicillin-streptomycin (P/S), trypsin-ethylenediaminetetraacetic acid (EDTA) were purchased from Gibco (Grand Island, NY, USA). CD4 and CD8 MicroBeads and the AutoMACS system were purchased from Miltenyi Biotec. (Shanghai, China). Biolegend (San Diego, CA, USA) -provided antibodies included: APC-conjugated CD29 (303008), CD19 (115512), and F4/80 (123115); PE-conjugated CD11c (117307), Sca1 (108107), CD206 (141705), and inducible nitric oxide synthase (iNOS) (696805); with PE-Cy7-CD44 (103029) and FITC-CD11b (101205). Hieff qPCR SYBR Green Master Mix was acquired from Yeasen (Shanghai, China). FOXP3/Transcription Factor Staining Buffer set was purchased from eBioscience (San Diego, CA, USA). All primers were obtained from Sangon Biotech (Shanghai, China). TD139, Galectin-3 were purchased from Selleck&Bimake (Selleck&Bimake, Shanghai, China).
Male BALB/c (H-2d) and C57BL/6 (H-2b) mice (10–12 weeks old) were obtained from Shanghai Laboratory Animal Center (SLAC, Shanghai, China) and maintained in specific pathogen-free (SPF) conditions with autoclaved water and standard chow. All animal tests were conducted in compliance with the guidelines established by the Ethics Committee of Xinhua Hospital Affiliated with the Shanghai Jiao Tong University School of Medicine (Ethics Approval No. XHEC-F-2022-076). All mice in this study were euthanized by cervical dislocation, in strict accordance with the guidelines outlined by the Ethics Committee of Xinhua Hospital Affiliated with the Shanghai Jiao Tong University School of Medicine (Shanghai, China).
MSCs isolated from bone marrow of C57BL/6 mice were cultured in Dulbecco modified Eagle medium-low glucose (DMEM-L; Hyclone, Shanghai, China) containing 10% heat-inactivated fetal bovine serum (FBS; Gibco, USA) and 1% penicillin-streptomycin (10,000 U/mL penicillin, 10,000 µg/mL streptomycin) (1% P/S; Gibco, USA) at 37 °C in a humidified atmosphere of 5% CO2. MSCs at passage 2 (P2- MSCs) were passaged by 0.25% trypsin-ethylenediaminetetraacetic acid (EDTA; Gibco, Burlington, Canada) at a 1:3 ratio when they were 80%–90% confluent, up to P12. Here, the MSCs were classified as early passages of MSCs (passage 4 to 6-MSCs) and late passages (passage 7 to 12-MSCs). MSCs exhibited typical fibroblast-like morphology, characterized by long, polygonal, plastic-adherent cells, and display the typical immunophenotypes, including Sca1 (+), CD29 (+), CD44 (+), CD19 (–), and CD11b (–) (Supplementary Fig. 1A,B).
The Raw264.7 cells, a macrophage cell line, were purchased from the American Type Culture Collection (Manassas, VA, USA). Cells were cultured in Dulbecco modified Eagle medium glucose (DMEM; Hyclone, China) containing 10% FBS and 1% P/S at 37 °C in a humidified atmosphere of 5% CO2.
All cell lines (including Raw264.7) were validated by STR profiling and tested negative for mycoplasma contamination.
Firstly, total RNA was extracted from cells using EZ-press RNA Purification Kit
(EZBioscience, CA, USA) according to manufacturer’s protocol. Total RNA meeting
purity criteria (A260/A280 ratio 1.8–2.0) was converted to complementary
deoxyribonucleic acid (cDNA) with PrimerScript™ RT Master Mix
(Takara, Shiga, Japan) per manufacturer’s instructions. qRT-PCR was performed
using relevant gene-specific primers and Hieff qPCR SYBR Green Master Mix
(Yeasen, China) on Step One Plus Real-time PCR System (Life Technologies, CA,
USA). The messenger ribonucleic acid (mRNA) expression levels of target genes
were normalized to
| Gene | Forward Primers (5′-3′) | Reverse Primers (5′-3′) |
| CATTGCTGACAGGATGCAGAAGG | TGCTGGAAGGTGGACAGTGAGG | |
| Galectin-3 (LGALS3) | AGACAGCTTTTCGCTTAACGA | GGGTAGGCACTAGGAGGAGC |
| TGF- |
CTCCCGTGGCTTCTAGTGC | GCCTTAGTTTGGACAGGATCTG |
| IL-10 | GCTCTTACTGACTGGCATGAG | CGCAGCTCTAGGAGCATGTG |
| Arg | AGCACTGAGGAAAGCTGGTC | TACGTCTCGCAAGCCAATGT |
| Ym1 | CAGGTCTGGCAATTCTTCTGAA | GTCTTGCTCATGTGTGTAAGTGA |
| CD206 (MR) | AAACACAGACTGACCCTTCCC | GTTAGTGTACCGCACCCTCC |
| IL-1 |
GTCACAAGAAACCATGGCACAT | GCCCATCAGAGGCAAGGA |
| IL-6 | GAACAACGATGATGCACTTGC | CTTCATGTACTCCAGGTAGCTATGGT |
| TNF- |
CTTCTGTCTACTGAACTTCGGG | CACTTGGTGGTTTGCTACGAC |
| COX2 | GCCTACTACAAGTGTTTCTTTTTGCA | CATTTTGTTTGATTGTTCACACCAT |
| CCL2 | AGGTGTCCCAAAGAAGCTGTA | ATGTCTGGACCCATTCCTTCT |
| CCL3 | TGACACTCTGCAACCAAGTCTTC | AACGATGAATTGGCGTGGAA |
| CCL4 | TTCTCTTACACCTCCCGGCAG | GTACTCAGTGACCCAGGGCTCA |
| iNOS | CAGCTGGGCTGTACAAACCTT | CATTGGAAGTGAAGCGTTTCG |
| Galectin-1 (Lgals1) | AACCTGGGGAATGTCTCAAAGT | GGTGATGCACACCTCTGTGA |
qRT-PCR, Quantitative Real-time PCR; MSCs, Mesenchymal stem cells;
TGF-
Fluorescein conjugated monoclonal antibodies (mAbs) used for experiments include phycoerythrin (PE)-conjugated mAbs to mouse, Fluorescein isothiocyanate (FITC)-conjugated mAbs to mouse, allophycocyanin (APC)-conjugated mAbs to mouse and R-Phycoerythrin-Cyanine 7 (PE-Cy7)-conjugated mAbs to mouse. Antibody information are provided in the Reagents and Animals section. In brief, for cell surface antigen staining, cells were collected, washed with phosphate buffered solution (PBS) and incubated with relevant fluorescein-conjugated mAb in the dark for 30 minutes at 4 °C. For cell nuclear/intracellular antigen staining, after stained with cell surface antigen, cells were fixed and permeabilized with eBioscience™ FOXP3/Transcription Factor Staining Buffer set (Invitrogen, CA, USA) according to the manufacture’s protocol followed by intracellular staining with relevant fluorescein-conjugated mAb in the dark for 30 minutes at room temperature. Finally, cells were washed with PBS and performed flow cytometric analysis. Flow cytometry (FCM) was performed by using BD FACS CantoII (Becton-Dickinson Bioscience, Franklin Lakes, NJ, USA) and analysis was performed by using FlowJo software (Version 10.8.1; BD Biosciences, Ashland, OR, USA).
Mice underwent bone marrow (BM) transplantation (BMT) according to the protocol
described previously [34]. Briefly, recipient mice (BALB/c, male, 10–12 weeks)
were randomly divided into three groups as follows: the T cell depleted-bone
marrow cells (TCD-BMC) + T cells + vehicle group, the TCD-BMC + T cells + P6-MSCs
group and the TCD-BMC + T cells + P12-MSCs group. At day 0, after BALB/c
recipients received 7.0 Gy (137Cesium source at 100 cGy/min) total body
irradiation (TBI), TCD-BMC (5
Spleens harvested from C57BL/6 mice were grinded through 40-µm nylon cell
strainers and obtained splenic cell suspension. CD4+/CD8+ T cell
isolation or depletion was performed using CD4 and CD8a MicroBeads and the
AutoMACS system (Miltenyi Biotec, Germany) according to the manufacturer’s
protocol. Briefly, 10 µL of CD4 MicroBeads and 10 µL of CD8a
MicroBeads were added per 1
Bone marrow cells (BMC) were isolated from the femurs and tibias of C57BL/6 mice
following humane euthanasia. The marrow cavity was flushed with ice-cold
DMEM-complete medium using a 25-gauge needle to obtain a single-cell suspension,
which was sequentially filtered through a 40-µm nylon cell strainer to
remove debris. Red blood cells in the suspension were lysed with ACK lysis
buffer, and the resulting nucleated cell pellet was resuspended in MACS Buffer at
a concentration of 1
Macrophages were harvested from the peritoneal lavage of recipient mice at day 21 after BMT. After intraperitoneal injection of pre-cooled PBS, gentle abdominal massage was performed prior to peritoneal lavage fluid collection. Finally, the harvested cells were stained for flow cytometric analysis using anti-mouse CD11b-FITC, anti-mouse F4/80-APC and anti-mouse CD206-PE and anti-mouse iNOS-PE antibodies. CD11b+F4/80+ double-positive cells were identified as peritoneal macrophages (Supplementary Fig. 3).
To investigate the effects of MSCs on macrophages at different passages, P6-MSCs
and P12-MSCs were seeded into the lower chamber of transwell (Corning, USA,
catalog number 3412, pore diameter 0.4 µm, polycarbonate membrane) at a
density of 2
Raw264.7 cells were seeded into 24-well plates at a density of 3
MSCs at different passages were seeded in 96-well cell culture plates at
3
Raw 264.7 macrophages were seeded in 24-well cell culture plates at 1.5
All experiments were independently repeated a minimum of two to three times. The
figure legends provide detailed information on reproducibility and sample sizes.
Data are presented as mean
To determine whether passage number of MSCs would affects the therapeutic
efficacy of MSCs in GVHD, recipient mice were received MSCs at different passages
after BMT (Fig. 1A). Although there were no significant differences in survival
of mice among three groups (Fig. 1B), the clinical GVHD scores were significantly higher in
recipients treated with P12-MSCs compared to those treated with P6-MSCs (Fig. 1C,
p
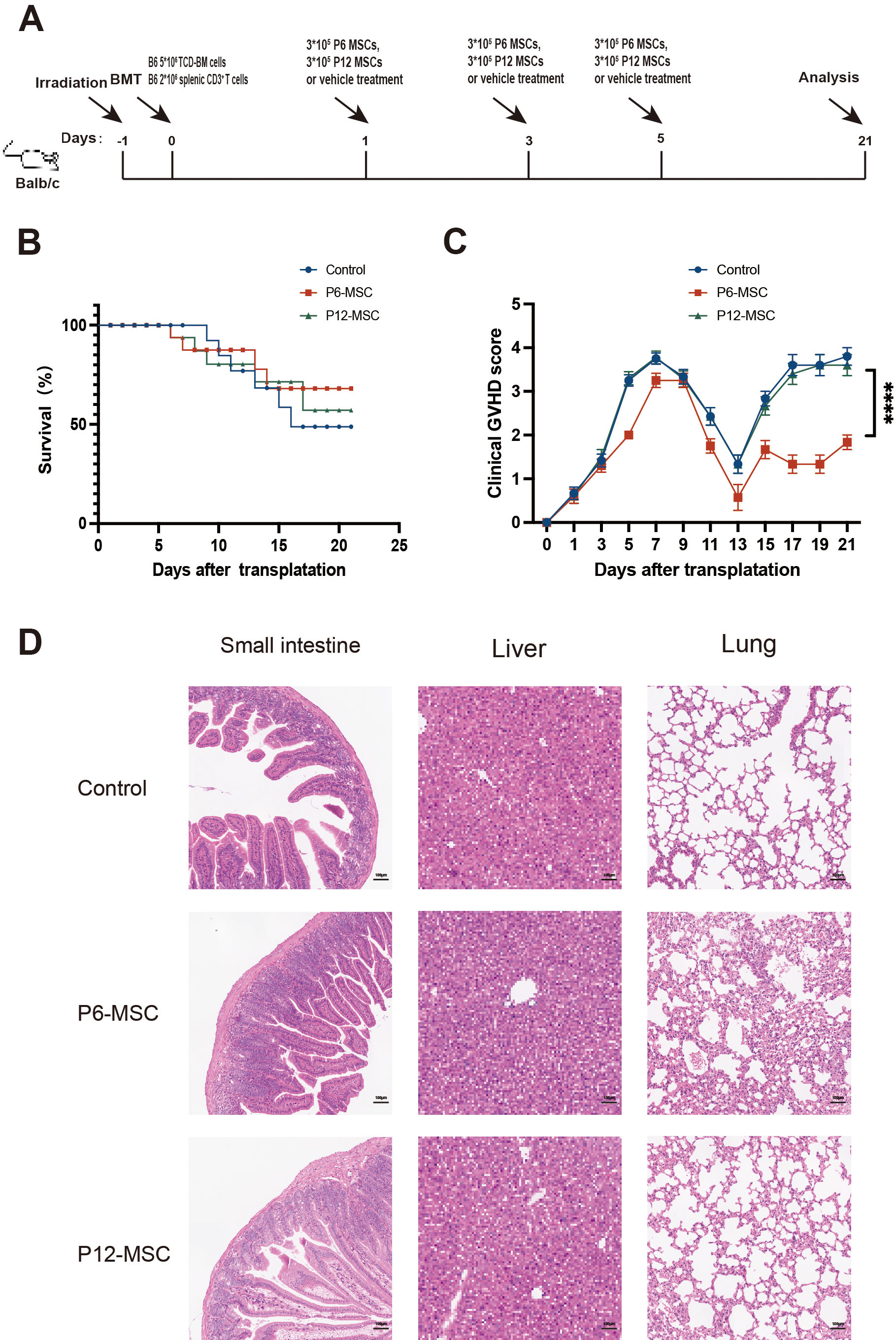 Fig. 1.
Fig. 1.
Effects of MSCs at different passages on the GVHD mice. (A)
Experimental scheme for GVHD model establishment and treatment. (B) Kaplan-Meier
survival curve of GVHD mice treated with PBS, P6-MSCs, P12-MSCs. (C) Clinical
GVHD score of GVHD mice treated with PBS, P6-MSCs, P12-MSCs. (D) Representative
histopathology images of liver, lung and small intestine from GVHD mice treated
with PBS, P6-MSC and P12-MSC on day 21 after transplantation. Scale bar, 100
µm. Data were pooled from 3 independent experiments. N = 3–5 in each
group. ****p
Next, we sought to explore the potential mechanisms underlying the variation in
MSCs at different passages in the context of GVHD. Given the powerful
immunoregulatory effects of MSCs on macrophages, we next assessed the
polarization of macrophages isolated from peritoneal lavage liquid of GVHD mice
model treated with MSCs at different passages. Compared to the P6-MSCs-treated
mice, the proportion of F4/80+ CD11b+ CD206+ M2-like macrophages
was lower in the P12-MSCs-treated mice (Fig. 2A, p
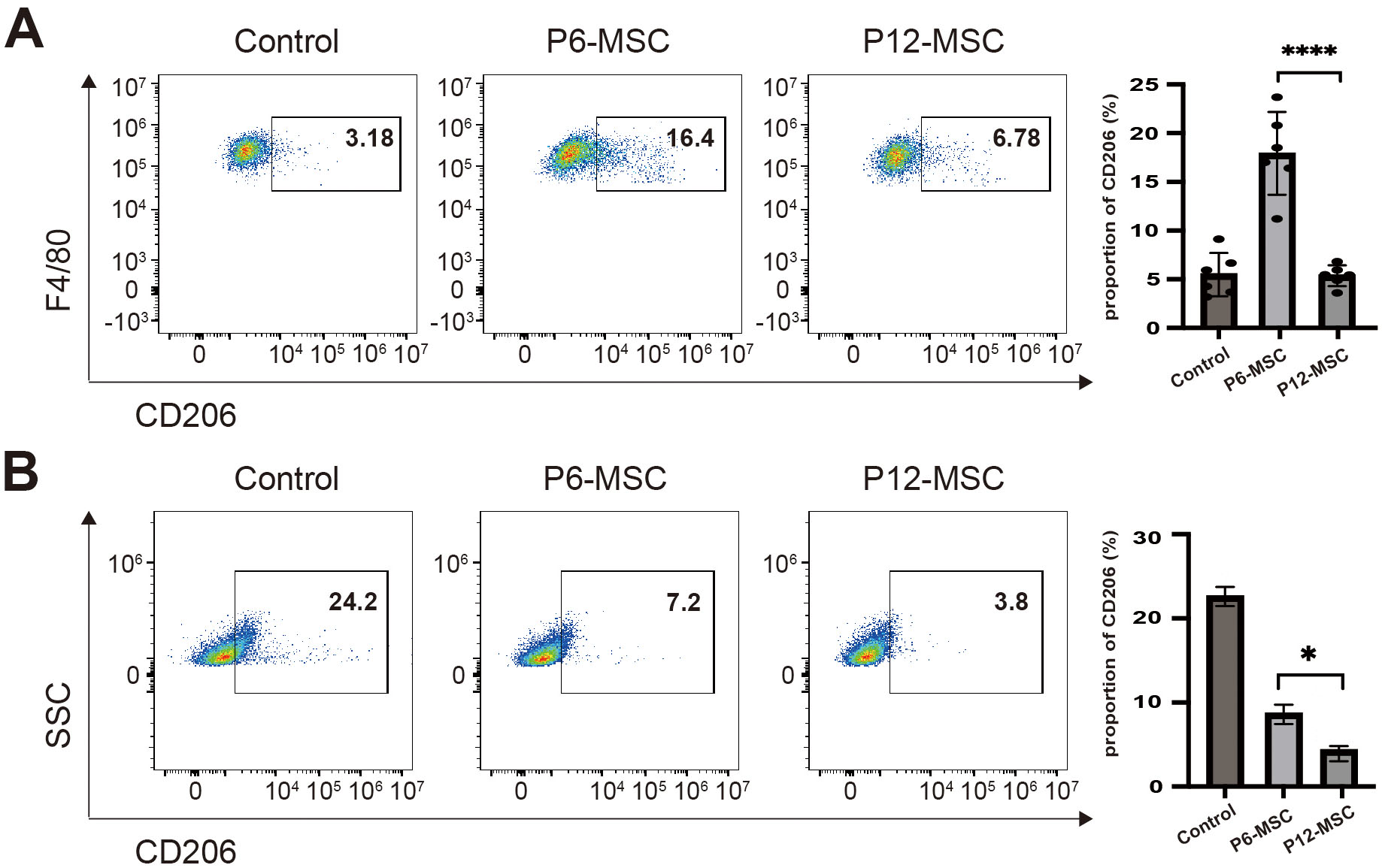 Fig. 2.
Fig. 2.
The effect of MSCs on macrophage polarization both in
vivo and in vitro. (A) Flow cytometric analysis of CD206+ M2-like
macrophage proportion within F4/80+CD11b+ peritoneal macrophages from
GVHD mice treated with PBS, P6-MSCs or P12-MSCs on day 21 post-transplantation.
(B) Flow cytometric analysis of CD206+ M2-like macrophages percentages in
Raw264.7 cells co-cultured with vehicle control (culture medium alone), P6-MSCs,
or P12-MSCs without LPS stimulation for 48 hours. Data were pooled from 3
independent experiments. *p
To simulate the in vivo inflammatory environment, we supplemented the MSCs-Raw264.7 macrophage co-culture system with LPS. Although the proportion of CD206+ M2-like macrophages and iNOS+ M1-like macrophages did not differ significantly between P6-MSCs-treated and P12-MSCs-treated groups, mRNA expression of M2-associated anti-inflammatory mediators, such as IL-10 was reduced, while the mRNA expression of M1-associated pro-inflammatory mediators, including L-6, CCL4, and COX2, was elevated in the P12-MSCs-treated group compared to the P6-MSCs-treated group (Fig. 3A,B). Our results indicated that the therapeutic efficacy of MSCs at different passages for acute GVHD could be related to macrophage polarization and the effect of MSCs at late passage on M2-like macrophages polarization was notably weakened.
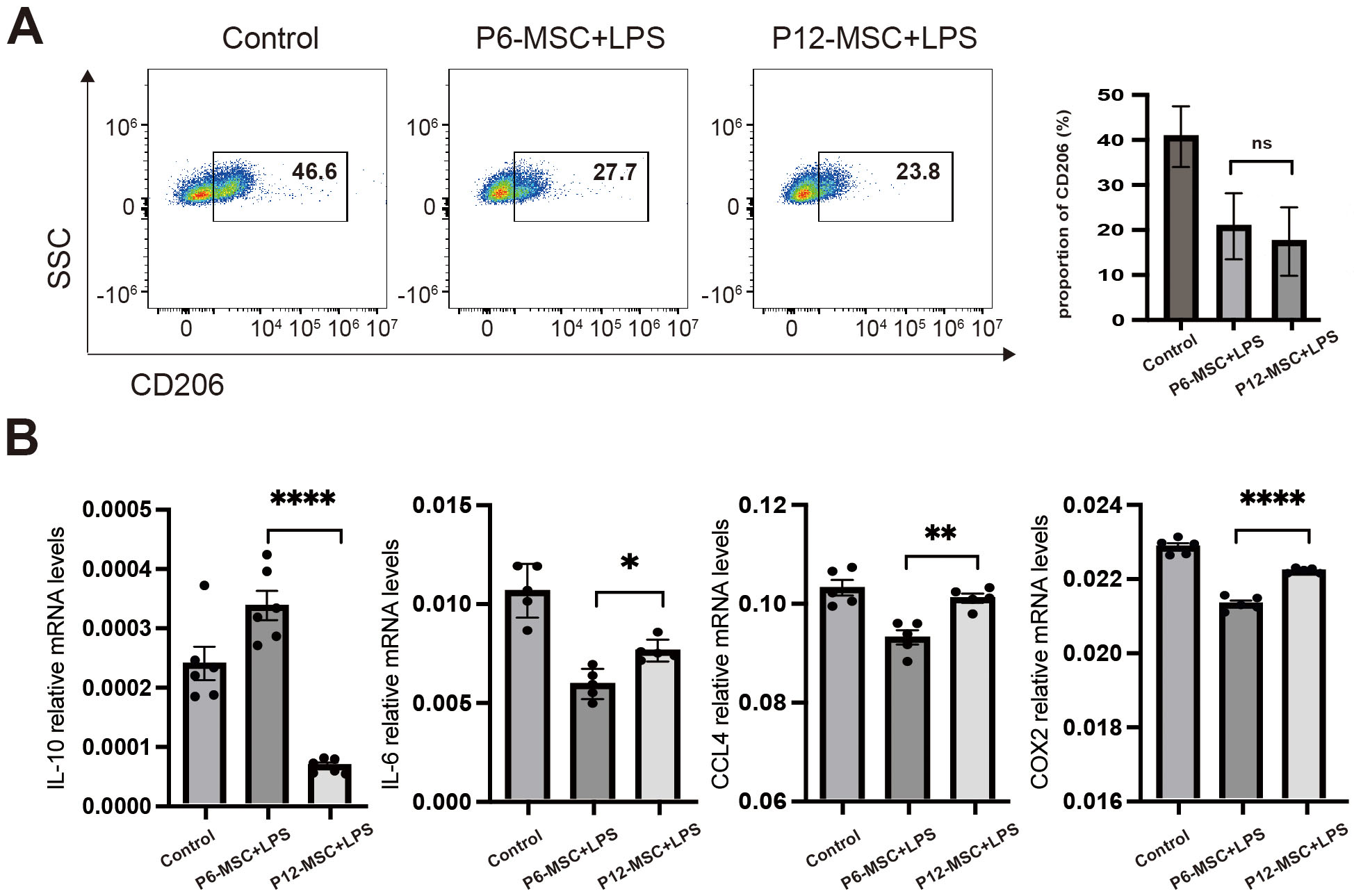 Fig. 3.
Fig. 3.
The effect of MSCs on macrophage polarization in the present of
LPS. Raw264.7 cells were cocultured with vehicle control (culture medium alone),
P6-MSCs or P12-MSCs in the present of 100 ng/mL LPS for 48 h, distinct from Fig. 2 (no LPS). (A) Flow cytometric analysis of CD206⁺ cell percentages in Raw264.7
macrophages. (B) qPCR analysis of macrophage polarization-related mediator
expression (IL-10, IL-6, CCL4, and COX2) in Raw264.7 macrophages. Data were
pooled from 3 independent experiments. Data are presented as the mean
Previous studies have demonstrated that the activation of Gal-3 enhances
inflammation [23], whereas the downregulation of Gal-3 can suppress inflammatory
responses [24]. Several studies have indicated that Gal-3 promotes a
pro-inflammatory response by inhibiting M2 macrophage polarization and inducing
apoptosis [21, 22]. MSCs constitutively express Gal-3 at both the mRNA and
protein levels, thereby exerting immunomodulatory effect via galectins [36].
Moreover, Gal-3 is a new informative model to predict aGVHD development [37].
Based on this, we inferred that Gal-3 expression may be linked to the effects of
MSCs on macrophages, to verify this point, we first analyzed the expression of
Gal-3 in MSCs at different passages. As shown in Fig. 4, Gal-3 expression
escalated starting at passage10 and markedly higher in MSCs at later passages
compared to those at earlier passages. Then, we assessed the characteristics of
Raw 264.7 macrophages in the presence of Gal-3. As shown in Fig. 5A, the
proportion of iNOS+ M1-like macrophages notably increased in Gal-3-treated
Raw 264.7 cells in a dose-dependent manner after 48 h. The M1-like macrophage
phenotype is characterized by the expression of chemokines such as iNOS, CCL2,
CCL3, CCL4 and the induction of pro-inflammatory mediators including
IL-1
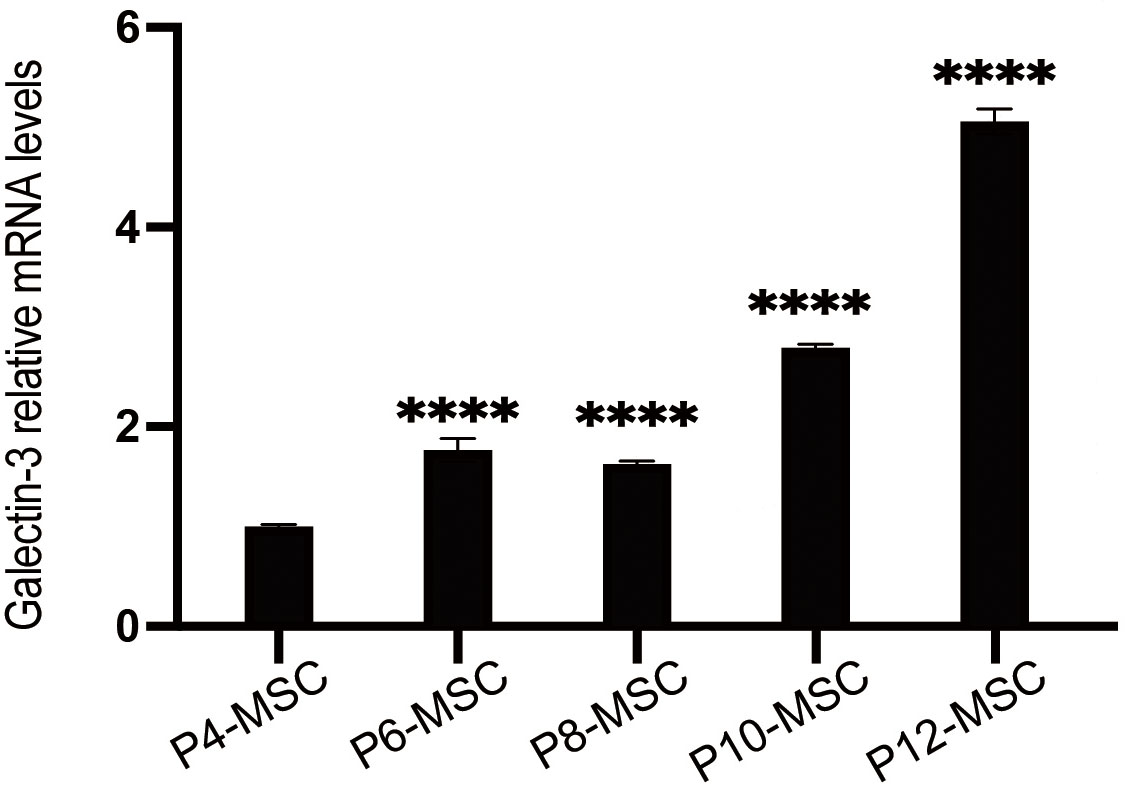 Fig. 4.
Fig. 4.
The mRNA expression of Galectin-3 in MSCs at different passages
by qPCR. Data were pooled from 3 independent experiments. Data are presented as
the mean
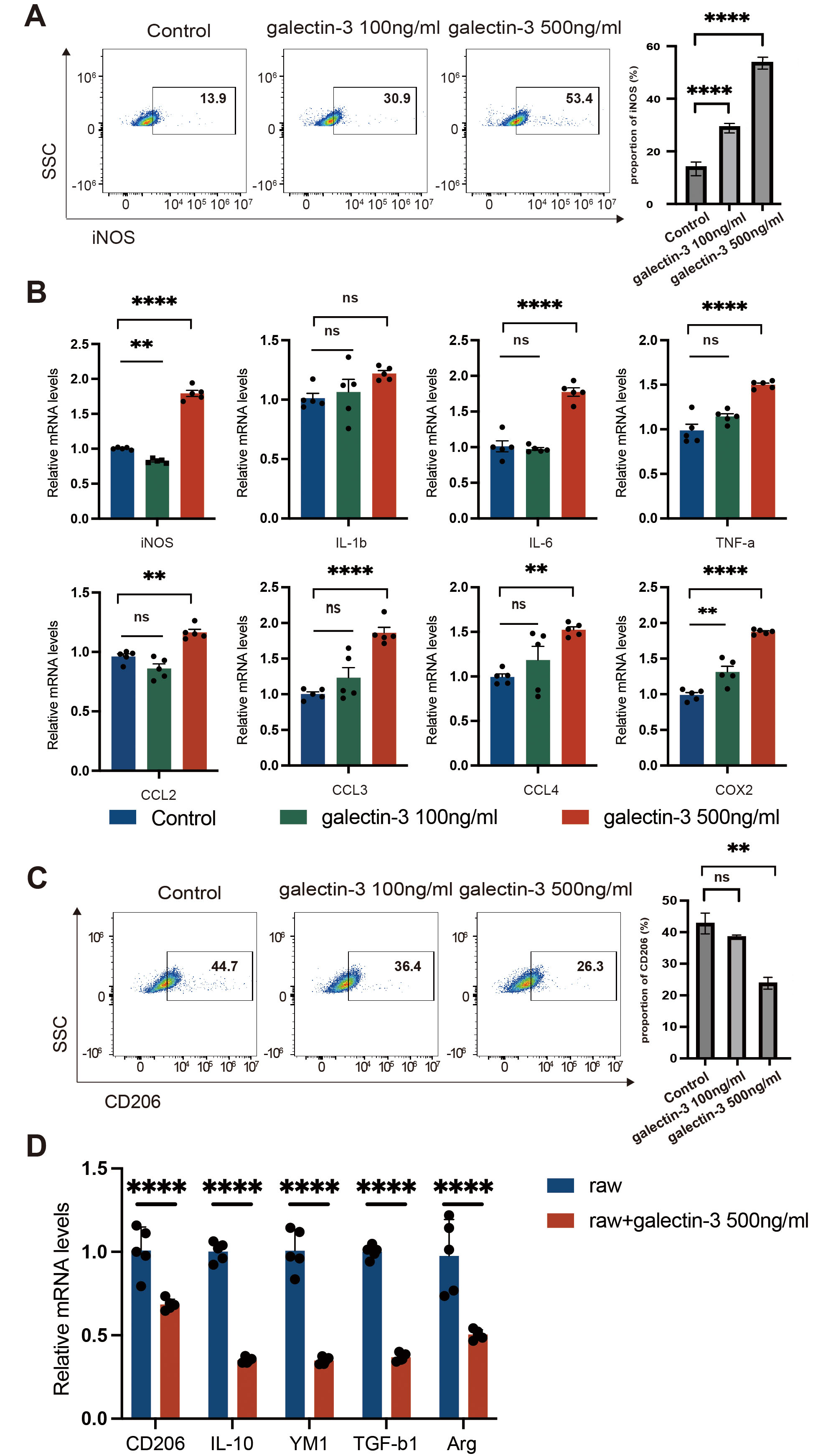 Fig. 5.
Fig. 5.
The effect of Gal-3 on macrophage polarization. Raw264.7 cells
were treated with vehicle, Gal-3 (100 ng/mL) or Gal-3 (500 ng/mL) for 48 h. (A)
Flow cytometry analysis of iNOS+ M1 macrophage proportion in Raw264.7
macrophages. (B) qPCR analysis of iNOS+ M1 macrophage polarization-related
mediator expression (iNOS, IL-1
Based on the above data, it is evident that elevated expression of Gal-3 in MSCs
at later passages results in a diminished effect of MSCs on M2-like macrophage
polarization. In this study, we sought to assess the potential
applications of Gal-3 inhibitors. While prior studies have focused on Gal-3, our
data demonstrated that Gal-1 is also expressed in MSC (Supplementary Fig.
4A). TD139, a selective small-molecule inhibitor of Gal-3, with no impact on
Gal-1 (Supplementary Fig. 4B), and is currently undergoing Phase II
clinical trials for idiopathic pulmonary fibrosis [28, 38]. Treatment with TD139
at concentrations ranging from 10–100 µM for 48 hours led to a significant
dose-dependent reduction of Gal-3 expression in P12-MSCs (Fig. 6A). Based on this
observation, a concentration of 100 µM TD139 was selected for subsequent
experiments. Similarly, Gal-3 expression was also inhibited in P6-MSCs following
high-dose TD139 treatment (Fig. 6B, p
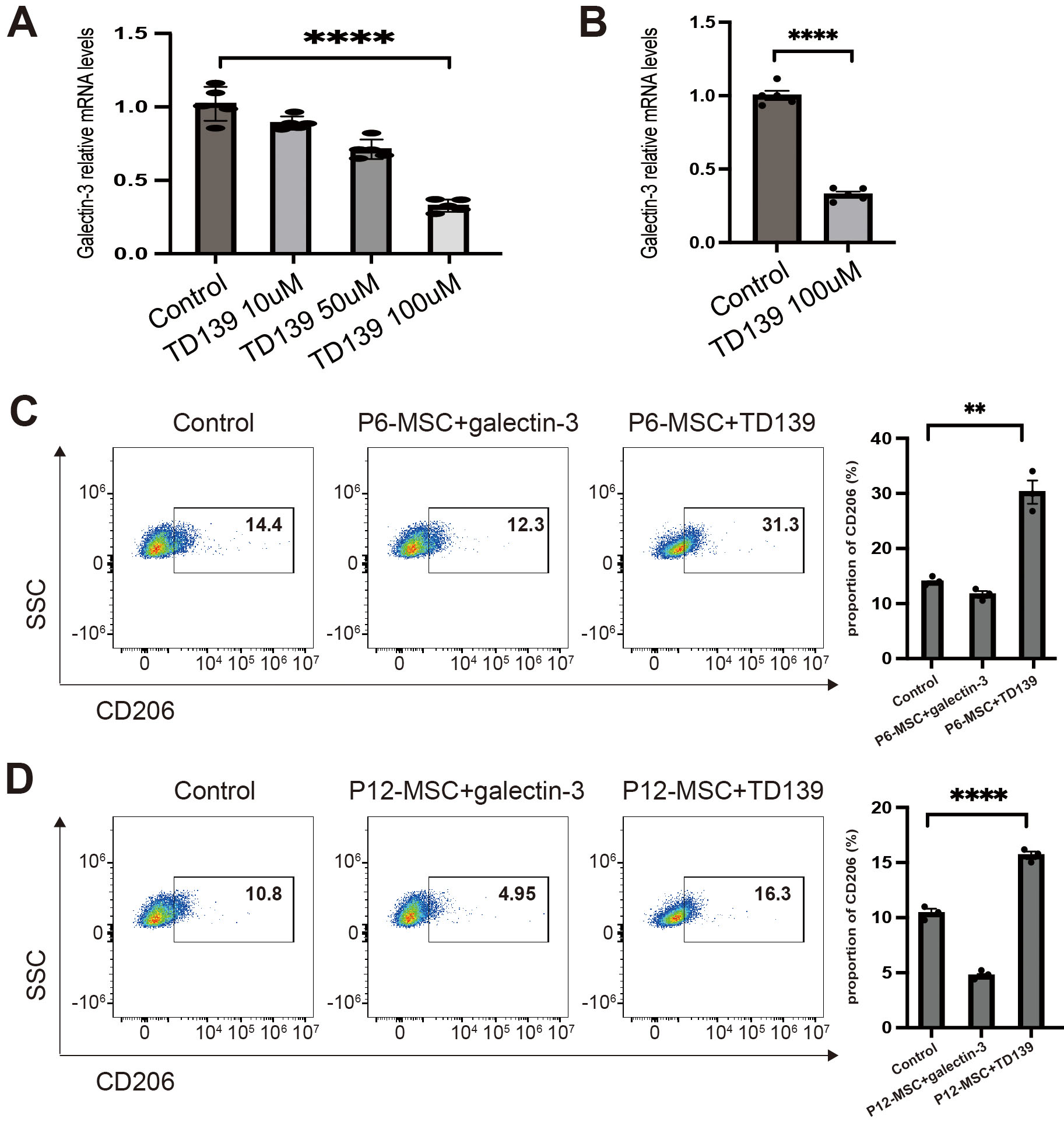 Fig. 6.
Fig. 6.
The effect of TD139 on Gal-3 expression of MSCs and macrophage
polarization induced by MSCs. MSCs at different passages were treated with DMSO,
TD139 (10 µM), TD139 (50 µM) or TD139 (100 µM) for 48 h (A,B).
MSCs at different passages cocultured with Raw264.7 macrophages were treated with
vehicle, Gal-3 (500 ng/mL) or TD139 (100 µM) for 48 h (C,D). (A) qPCR
analysis of Gal-3 expression in P12-MSCs. (B) qPCR analysis of Gal-3 expression
in P6-MSCs. (C) Flow cytometry analysis of CD206+ M2 macrophage proportion
in Raw264.7 macrophages cocultured with P6-MSCs. (D) Flow cytometry analysis of
CD206+ M2 macrophage proportion in Raw264.7 macrophages cocultured with
P12-MSCs. Data were pooled from 3 independent experiments. Data are presented as
the mean
To determine whether TD139 affect other profiles of MSCs, we also examined the immunophenotype of MSCs in presence of TD139. The results revealed that MSCs cultured with or without TD139 exhibited typical fibroblast-like morphology, characterized by long, polygonal, plastic-adherent cells, and displayed the expected immunophenotypes of MSCs, including Sca1 (+), CD29 (+), CD44 (–), CD19 (–), and CD11b (–) (data not shown). These findings indicated that TD139 effectively inhibits Gal-3 expression in MSCs without altering their fundamental biological properties, including morphology and immunophenotype.
Since TD139 inhibits Gal-3 expression in MSCs, we inferred that TD139 could
improve the immunoregulatory function of MSCs. In this study, we investigated the
impact of TD139 on macrophage polarization induced by MSCs. As shown in Fig. 6,
after incubation with TD139 at a concentration of 100 µM, the proportion of
CD206+ M2-like macrophages relative to Raw264.7 macrophages was
significantly increased in the TD139-treated P6-MSCs group compared to the
vehicle-treated P6-MSCs group (Fig. 6C, p
In the past few years, due to its immunosuppressive ability, MSCs have been applied for the treatment of GVHD after allo-HSCT, including gut GVHD, as well as for the management of refractory Crohn’s disease [4, 39, 40, 41]. MSCs derived from various biological sources usually require ex vivo expansion to achieve sufficient cell numbers for in vivo applications [14]. However, repeated in vitro passaging negatively affects the biological characteristics of MSCs, leading to a decline in their immunosuppressive and regenerative potential with each passage [16, 17, 18]. Therefore, it is necessary to develop strategies to save the immunosuppressive capability of MSCs during passaging, particularly for MSCs at later passage.
Several preclinical and clinical studies have documented the advantageous impacts of MSCs on GVHD [42, 43, 44]. Nevertheless, other studies have shown that in vitro aging adversely affects the immunosuppressive function of MSCs. Gao et al.’s study [45] elucidated that MSCs undergo a progressive aging process in vitro, and the cellular senescence influences their immunosuppressive activity. Von Bahr et al. [46] reported that in GVHD patients, 1-year survival was 75% in patients who received early-passage MSCs (from passages 1 to 2) in contrast to 21% in those who using later passage MSCs (from passages 3 to 4). In this study, we compared the therapeutic efficacy of MSCs at different passages in aGVHD mice model. Our data showed that compared to the control group, the severity of GVHD and tissue damage were not significantly different in the late-passage MSC group, whereas they were notably decreased in the early-passage MSC group, indicating that the therapeutic efficacy of MSCs diminishes with increasing passage.
In the pathogenesis of aGVHD, conditioning regimens in allo-HSCT initially activate host antigen-presenting cells, mainly macrophages and dendritic cells, thereby positioning macrophages as a central focus of research [34]. Previous studies have shown that with the progressive expansion in vitro, MSCs have undergone a phenotypic transformation from anti-inflammatory to pro-inflammatory, which leads to the deterioration of their therapeutic effects [47, 48, 49]. Thus, in this study, we examined peritoneal macrophages derived from aGVHD mice under various treatment. Consistent with previous studies, M2-like macrophages were increased in aGVHD mice treated with early-passage MSCs [50, 51, 52]. However, in contrast to the early-passage MSC treatment group, the number of M2-like macrophages was significantly reduced in the late-passage MSC group, suggesting that the variability in therapeutic efficacy of MSCs at different passages is associated with macrophage polarization. Subsequently, we conducted experiments with the Raw264.7 macrophage cell line, and the results showed that the effect of late-passage MSCs on M2-like macrophages polarization was notably weakened compared to early-passage MSCs. Taken together, these results underscored the relationship between the immunomodulatory efficient of MSCs at different passages and macrophages polarization.
Gal-3 belongs to the beta-galactoside-binding animal lectin family, manifesting
pleiotropic functions such as mediating cell–cell interactions, cell-matrix
adhesion, and transmembrane signaling. It is produced by MSCs and exerts
immunomodulatory effects [53]. Accumulating evidences have revealed that Gal-3
plays a pivotal role in macrophage polarization and the pathogenesis of aGVHD
[37, 54]. Li H et al. [54] provided evidence indicating that Gal-3
enhances inflammatory response by directly binding to NLRP3 in RAW264.7
macrophages. Therefore, we inferred that there may be an association between
Gal-3, MSCs and macrophages. In the present study, we observed that Gal-3
expression was significantly increased in late-passage MSCs. Moreover, Gal-3
promoted an inflammatory response in Raw264.7 macrophages, notably by increasing
M1-like macrophage polarization and expression of iNOS, IL-6, TNF-
TD139 has emerged as the foremost small molecule inhibitor targeting Gal-3 and has been extensively investigated in various experimental studies for its potential applications [60]. Currently, it represents a promising therapeutic agent for idiopathic pulmonary fibrosis in the phase II clinical trial [28]. Previous studies have underscored the therapeutic potential in lung fibrosis, cancer metastasis and hepatitis [61, 62, 63]. Here, we investigated the therapeutic role of TD139 in GVHD mice and its effects on MSCs for the first time. As shown in Supplementary Fig. 6, TD139 treatment in Raw 264.7 cells reduced the iNOS+ M1 macrophage population while increasing the CD206+ M2 macrophage subset. This data indicated that TD139 directly inhibits M1-like macrophage polarization while promoting M2-like polarization. Nevertheless, as shown in Fig. 6, our results strongly indicated that TD139 dose-dependently suppressed the expression of Gal-3 in MSCs without affecting their biological properties. Inhibition of Gal-3 by TD139 significantly promoted M2-like macrophage polarization in both the P6-MSCs-Raw264.7 and P12-MSCs-Raw264.7 coculture systems, suggesting that TD139 enhanced the effect of MSCs on macrophages by inducing M2-like polarization. More importantly, in the presence of TD139, the proportion of M2-like macrophages was higher in Raw264.7 cells cocultured with P12-MSCs than in those cocultured with P6-MSCs. Although TD139 exhibited direct effects on macrophage polarization, its efficacy was substantially potentiated with the present of MSCs, suggesting that TD139 partially restored the immunomodulatory function of MSCs. Taken together, these findings suggested that TD139 can save the diminished immunoregulatory function of MSCs, particularly late-passage MSCs, enabling more MSCs to be obtained through passaging while preserving their immunoregulatory capabilities.
Although TD139-pretreated MSCs is a promising strategy for GVHD therapy, several
challenges remain in their clinical application. We attempted to apply TD139 for
pretreating MSCs and subsequently inject TD139-pretreated MSCs into GVHD mice.
However, the results were not satisfactory (data not shown). In our opinion, the
following factors may contribute to this outcome: First, TD139 may not fully
inhibit the expression of Gal-3 in MSCs. Second, after TD139 pretreatment of MSCs
in vitro, the expression of Gal-3 in MSCs may recover in vivo.
To address these issues, we plan to use small interfering RNA (siRNA) to inhibit
the expression of Gal-3 in MSCs for a prolonged period in future studies.
Additionally, we aim to apply TD139 and MSCs simultaneously, rather than
pretreating MSCs with TD139. In addition, our data demonstrated that Gal-3
inhibited M2-like macrophage polarization, mechanistic details require further
study. Prior evidence implicated Gal-3 in regulating macrophage function through
NF-
In conclusion, as shown in Fig. 7, the reduced therapeutic effect of late passage MSCs in treating GVHD is related to their impaired capacity to induce M2 macrophage polarization. Our study showed that the expression of Gal-3 increased in late passage MSCs, and that Gal-3 can suppress M2 macrophage polarization. Therefore, it is likely that Gal-3 plays a pivotal role in mediating the ability of MSCs to regulate macrophage polarization. Notably, treatment with TD139, a selective Gal-3 inhibitor, can restore the immunoregulatory function of late passage MSCs in vitro.
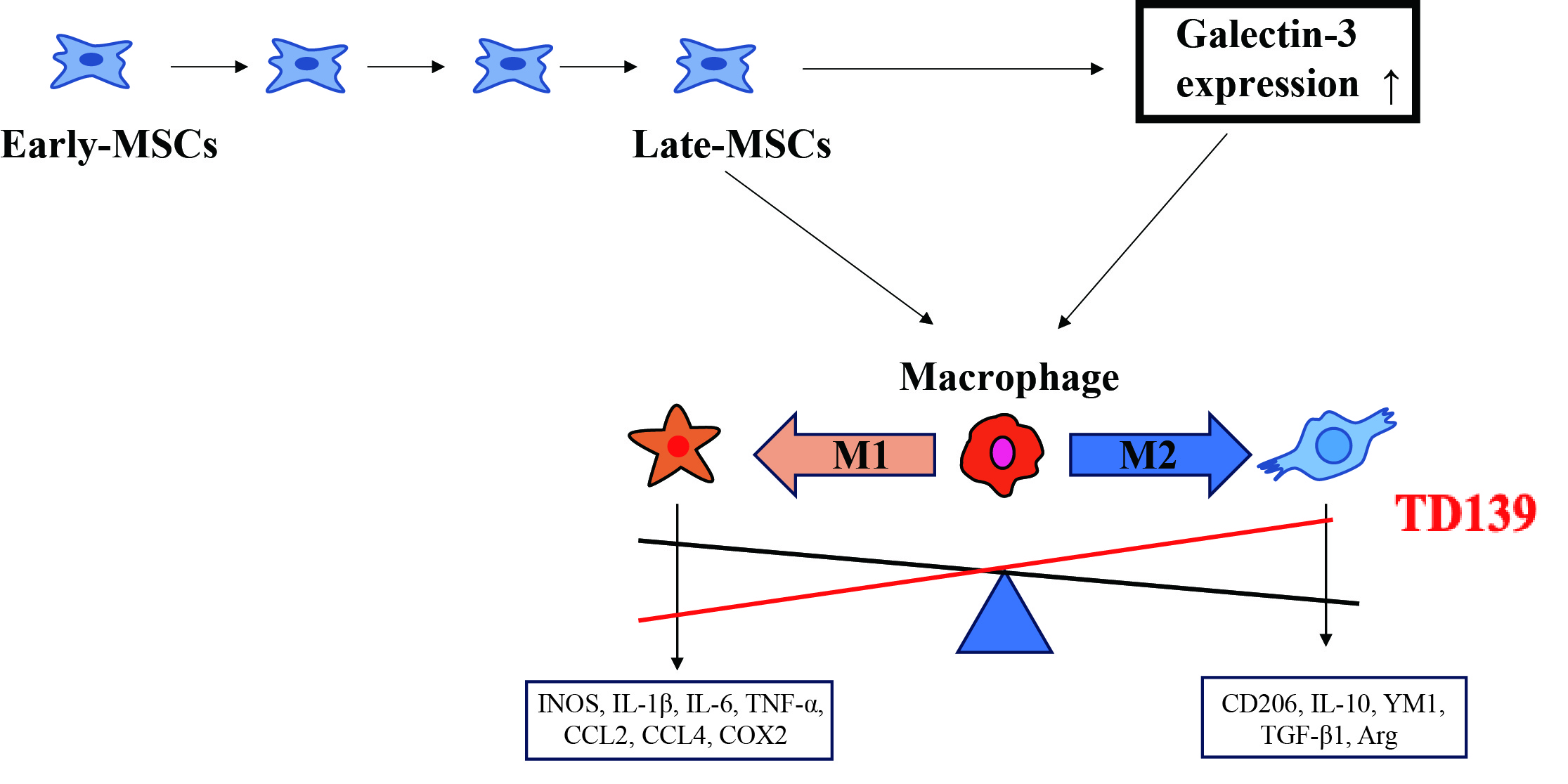 Fig. 7.
Fig. 7.
The diagram of main results. MSCs, mesenchymal stem
cells; IL-1
In summary, our study demonstrated that passaging in vitro impairs the therapeutic efficacy of MSCs in GVHD due to elevated Gal-3 expression in late-passage MSCs. Gal-3 directly suppressed M2-like macrophage polarization, thereby attenuating the immunoregulatory function of late-passage MSCs. TD139, a selective Gal-3 inhibitor, could restores the immunoregulatory function of MSCs, facilitating the clinical application of expanded MSCs for GVHD treatment.
MSCs, mesenchymal stem cells; allo-HSCT, Allogeneic hematopoietic stem cell transplantation; GVHD, graft versus host diseases; APCs, antigen-presenting cells; Gal-3, Galectin-3; LPS, lipopolysaccharide; IL-10, interleukin 10; IL-6, interleukin 6; COX2, cyclooxygenase 2; CCL2, CC-chemokine ligand 2; CCL3, CC-chemokine ligand 3; CCL4, CC-chemokine ligand 4; Arg, Arginase; IL-1
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The concept was conceived by SGH. The overall study design was developed by MHW, JBW and SGH. The experiments were performed by MHW, FH, YJ, DFZ, and JBW. The data analysis was performed by MHW and YJ. The manuscript was written by MHW, JBW and SGH. The research was supervised by JBW and SGH. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
The animal study was approved by the Ethics Committee of Xinhua Hospital Affiliated to the Shanghai Jiao Tong University School of Medicine (Ethics Approval No. XHEC-F-2022-076). All experimental procedures were performed in accordance with the 3R principle (Replacement, Reduction, and Refinement) for animal research, aiming to minimize animal suffering and optimize experimental design.
Not applicable.
This research received no external funding.
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBL39918.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.