

 , Oliver Stefani 4, Germaine Cornelissen 5
, Oliver Stefani 4, Germaine Cornelissen 51 Department of Biology, Tyumen Medical University, 625023 Tyumen, Russia
2 Laboratory for Chronobiology and Chronomedicine, Research Institute of Biomedicine and Biomedical Technologies, Tyumen Medical University, 625023 Tyumen, Russia
3 Tyumen Cardiology Research Center, Tomsk National Research Medical Center, Russian Academy of Sciences, 634009 Tomsk, Russia
4 Department Engineering and Architecture, Institute of Building Technology and Energy, Lucerne University of Applied Sciences and Arts, 6048 Horw, Switzerland
5 Halberg Chronobiology Center, University of Minnesota, Minneapolis, MN 55455, USA
Abstract
In discussing the interplay between light exposure/blue light exposure (LE/BLE) and circadian health, we emphasize the role of light hygiene and its effects on maintaining sleep, mood, and metabolic health, among other physiological processes. We define compromised circadian light hygiene as low dynamic range and/or irregular 24-hour patterns of LE. Poor light hygiene interferes with circadian entrainment and weakens circadian robustness alignment, thereby increasing health hazards. We provide an overview of the complex molecular pathways underlying light perception and downstream signaling. Given that genetic polymorphisms influence key elements within these light signaling pathways, we propose that personalized light hygiene approaches be designed for populations affected by compromised circadian LE or at risk of light-induced circadian disruption.
Keywords
- circadian
- light
- light hygiene
- melatonin
- mood
- metabolism
The circadian system orchestrates a plethora of physiological processes, including the sleep-wake cycle, metabolism, cognitive function, mood, and performance [1, 2]. Driven by a self-sustained endogenous clock and its associated molecular machinery, the circadian system is entrained by environmental cues, light serving as principal synchronizer, aligning internal rhythms to the 24-h environmental light-dark cycle. While exposure to natural daylight promotes circadian synchrony and robustness, light at night and irregular light patterns disrupt melatonin secretion and sleep, leading to circadian misalignment. Since robust rhythms are a hallmark of health [1], circadian disruption is associated with a heightened disease risk and the actual presence of numerous diseases. Circadian light hygiene is the purposeful design of the pattern of exposure to light [2]. It stands for the strategic management of dynamic light exposure (LE) patterns to ensure robust rhythmicity and circadian health in both contexts of primary prevention and countermeasure or treatment [3]. Beyond their direct effect on circadian rhythms, lighting interventions can influence behavior in healthcare settings [4]. To effectively implement circadian light hygiene and realize its potential benefits for health and well-being, its underlying molecular mechanisms need to be fundamentally understood (Fig. 1) [5, 6]. Mimicking natural daylight with alternative light sources is vital where natural light is inadequate, such as in winter at high latitudes.
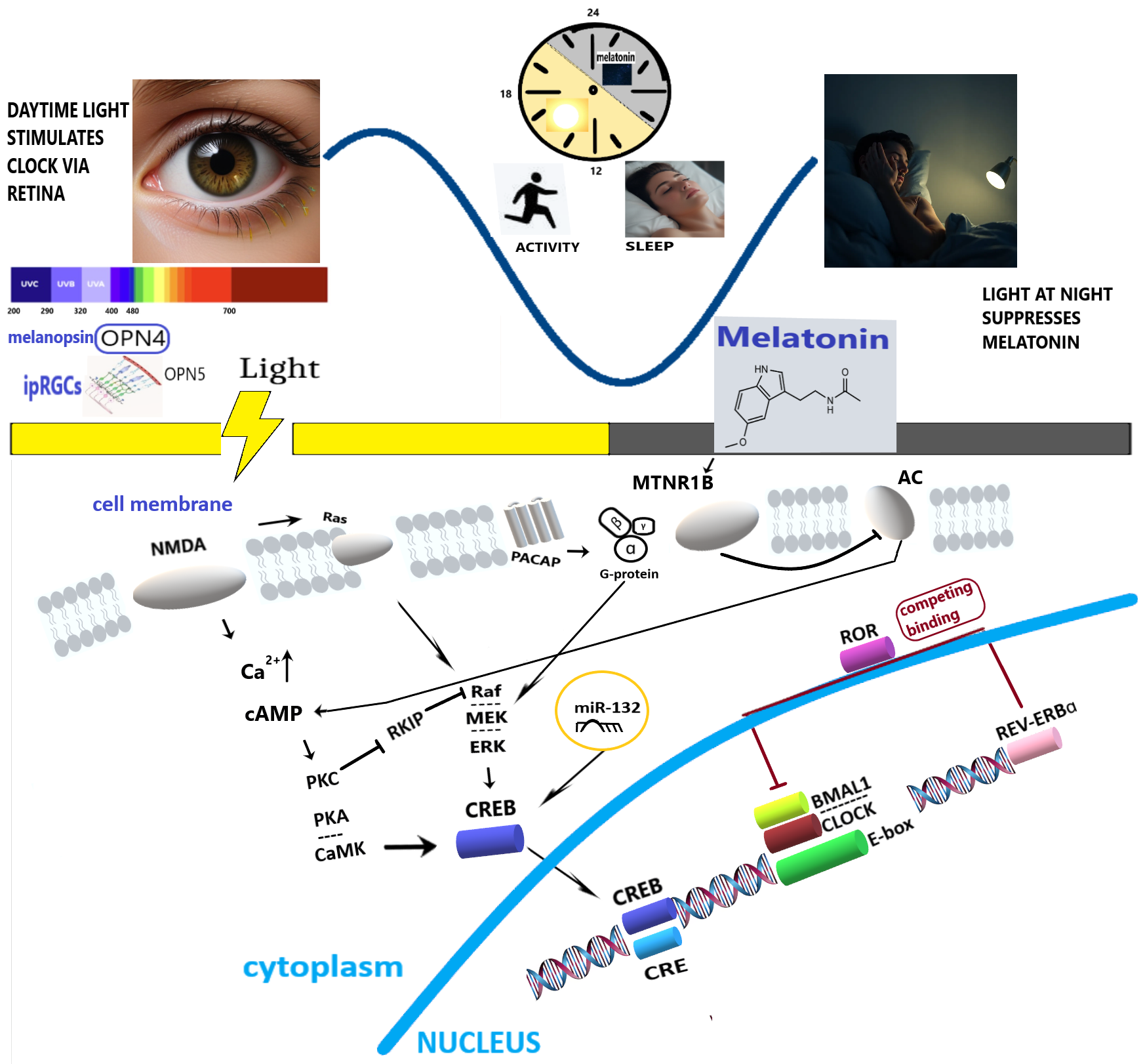
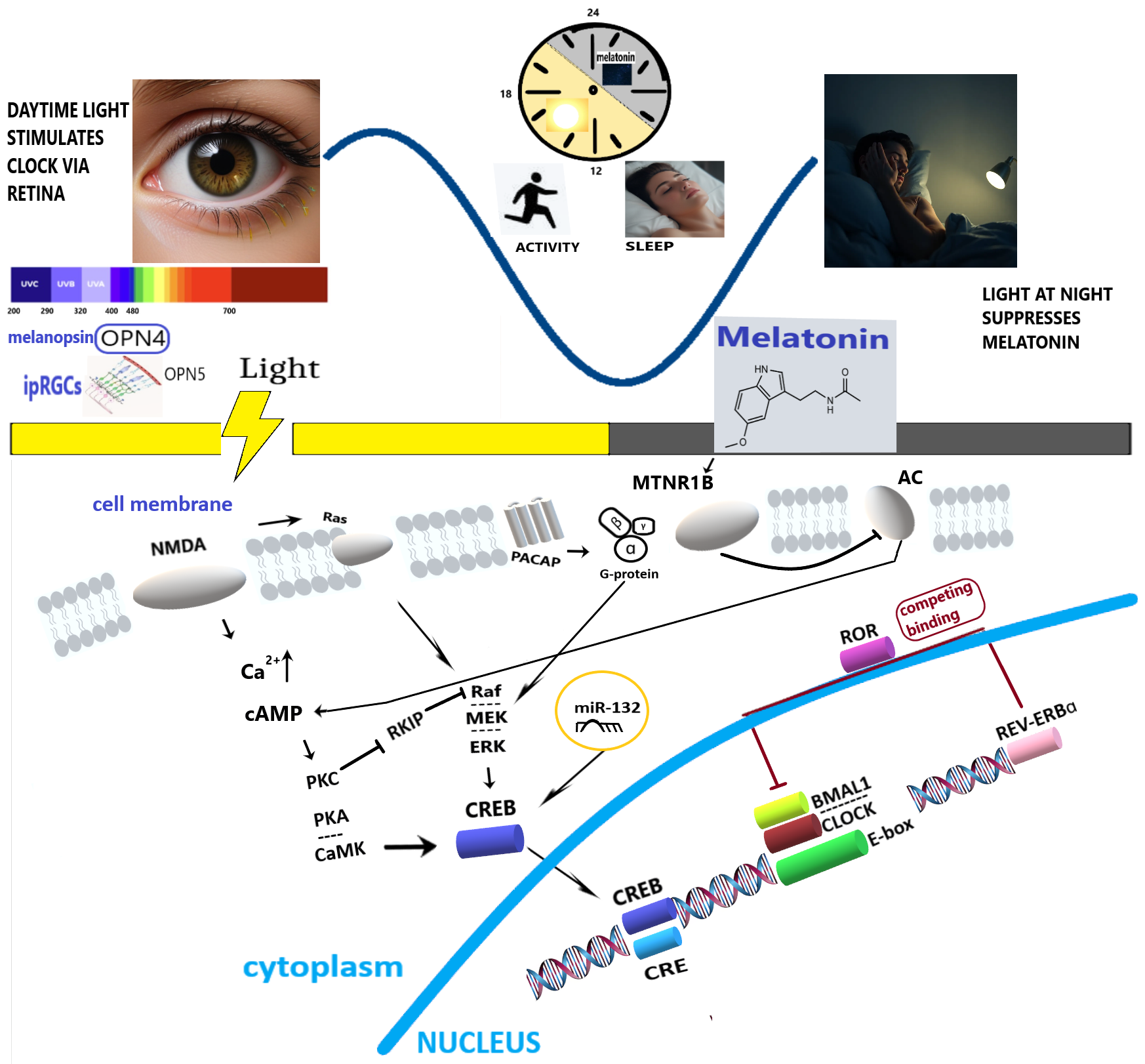 Fig. 1.
Fig. 1.
Simplified molecular pathways mediating the effect of light-dark alternation on human circadian rhythms and health. OPN4, melanopsin; OPN5, neuropsin; ipRGCs, intrinsically photosensitive Retinal Ganglion Cells; NMDA, N-Methyl-D-Aspartate receptor; PACAP, Pituitary Adenylate Cyclase-Activating Polypeptide; MTNR1B, Melatonin Receptor Type 1B; AC, Adenylate cyclase; cAMP, cyclic Adenosine Monophosphate; PKC, Protein Kinase C; PKA, Protein Kinase A; Rafs-Raf-MEK-ERK, Ras/Raf/Mitogen-activated Protein Kinase/ERK Kinase (MEK)/Extracellular-signal-Regulated Kinase (ERK); CaMK, Ca2+/Calmodulin-Dependent Protein Kinase; RKIP, Raf Kinase Inhibitory Protein; CREB, cAMP Response Element-Binding protein; miR-132, microRNA 132; ROR, Retinoic acid receptor-related Orphan Receptor; REV-ERBɑ, Nuclear Receptor Subfamily 1 Group D Member 1; CRE, cAMP Response Element; BMAL1, Brain and Muscle Arnt-Like 1; CLOCK, Circadian Locomotor Output Cycles Kaput; E-box, Enhancer box.
Light, clocks, and sleep. Knowing which types of light and how they signal at the molecular level is important to develop light hygiene. Light perception begins in the retina, where opsins, light-sensitive proteins of the G-protein-coupled receptor superfamily, reside within specialized cells. The ability to sense light relies on rhythmic cyclic guanosine monophosphate signaling and the expression of melanopsin (OPN4) and neuropsin (OPN5). Melanopsin, a pigment of intrinsically photosensitive retinal ganglion cells (ipRGCs), is the primary mediator of light-driven circadian entrainment in the suprachiasmatic nucleus (SCN), with peak sensitivity to blue light (approximately 480 nm) [7]. LE suppresses melatonin production in the pineal gland via the melanopic pathway, with substantial inter-individual variability in sensitivity [8]. A meta-analysis has retrospectively shown a correlation between light, melanopsin activation, and both sleep and vigilance [9]. However, a recent clinical trial showed that evening melatonin suppression induced by artificial blue light exposure (BLE, ending 50 minutes before habitual bedtime) did not result in statistically significant immediate changes in subjective sleepiness, sensory processing, or quantitative sleep parameters [10]. The color of the light source did not affect non-visual functions in humans either [11]. These functions are primarily mediated by melanopsin photoreception, specifically through the melanopic equivalent daylight illuminance (melanopic EDI) of the light source. Melanopic EDI is an internationally recognized standard used to quantify the impact of light on melanopsin, the photoreceptor that plays a crucial role in mediating the non-visual effects of light [3, 12]. These effects include acute alertness, melatonin suppression, and circadian phase shifting. These results are supported by recent findings that low-melanopic light can effectively reduce the adverse effects of use of electronic displays at night, confirming the key role of melanopsin in setting the circadian clock [13]. Retinal ganglion cells (RGCs) loss in diseases such as glaucoma affects light signaling, with consequences for circadian rhythms [14]. Extreme seasonality of LE patterns at high latitudes modulates the circadian amplitude and phase of melatonin [15]. Macronutrients [16] and physical activity [17] act in tandem with LE to facilitate circadian entrainment and robustness.
Metabolic effects of light. They likely differ between diurnal and nocturnal species due to differences in activity patterns and their relation to the timing of melatonin secretion, a hormone intrinsically linked to darkness. In humans, the distribution of LE throughout the 24-h cycle, rather than its absolute quantity, appears as the dominant factor influencing the circadian amplitude and phase of melatonin, and sleep [14]. Individual metabolic effects of LE may be uncoupled from sleep disturbances, but linked to melatonin receptor polymorphism and distinct 24-h patterns of skin temperature [18]. LE affects adaptive thermogenesis in brown adipose tissue (BAT) via the retina–supra-optic nucleus–BAT axis that inhibits BAT thermogenesis and reduces glucose tolerance [19, 20]. We hypothesize the existence of chronobiological windows of susceptibility: “hot spots” vs. “blind spots” with greatest vs. lowest sensitivity/molecular responsiveness to light within the 24-h cycle. For a metabolic response, hot spots likely cluster around sleep-wake transitions, as observed in our study [18]. Such circadian gating can be modulated by genetic polymorphisms in melatonin receptors [21] that may also affect circadian aspects of thermoregulation and metabolism, albeit according to principally different ways between nocturnal and diurnal species [22]. It suggests that the genetic background that governs circadian preference in humans (chronotype) can modulate light-medicated effects on thermoregulation and metabolism. Genetic polymorphisms, such as the melatonin receptor type 1B (MTNR1B) variant rs10830963, may augment metabolic and thermogenic responses to low-intensity evening BLE at “hot spot” times such as three hours before midnight for individuals with regular sleep (e.g., 22:30–06:50), as observed in actigraphy studies [18]. Melatonin receptors share a common evolutionary ancestor with opsins, the primary light-sensitive chromophores in organisms [23]. In humans, BLE modulates glucose tolerance at temperatures where BAT is active [19]. Human metabolism is negatively affected by later mealtimes that result in elevated glucose concentrations during the period of active melatonin production [24, 25]. The mechanism involves the interference by melatonin of insulin secretion, a disruption that may be further amplified in carriers of the MTNR1B rs10830963 polymorphism [26]. The MTNR1B rs10830963 variant was also related to the inter-individual variability in the glucose tolerance test’s response to the quantity and duration of daytime LE [27]. Disrupted circadian rhythms are increasingly recognized as a significant contributor to metabolic disorders, including obesity, cardiovascular disease, hepatic steatosis, and type 2 diabetes [28].
Genetic background. While core clock genes are essential for timekeeping, genome-wide association studies show that many genes, which influence light sensitivity and chronotype are not traditional clock genes, suggesting that diverse biological processes affect individual light responses and sleep timing [6, 8, 29, 30, 31]. That MTNR1B encodes the melatonin receptor, a G-protein coupled receptor, suggests a potential convergence of light-sensitive pathways and metabolic regulation. A systematic gene-property analysis of light sensitivity loci identified significant over-representation of genes crucial to the G protein-coupled glutamate receptor signaling pathway and genes specifically expressed in Per2-positive hypothalamic neurons, highlighting the convergence of glutamatergic signaling and circadian rhythm regulation in the modulation of light sensitivity [6]. This study also linked light sensitivity to the ARL14EP gene and associations with circadian traits. The ARL14EP gene codes for a protein that interacts with ADP-ribosylation factor-like GTPase 14 (ARL14), a key regulator of protein trafficking, especially for Major Histocompatibility Complex class II molecules in immune cells. These molecules are essential for presenting antigens to T lymphocytes, launching an immune response. Our data also reveal an association between higher daytime LE and coronavirus disease 2019 (COVID-19) negative status [32], while a history of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection appears to amplify the negative impact of poor circadian light hygiene on circadian rhythms [33]. While interesting, any association of ARL14EP with COVID-19 susceptibility will need further study. Whether ARL14EP or other mechanisms underlie this enhanced vulnerability to light-related circadian disruption in post-COVID-19 individuals also warrants further investigation.
Molecular clockwork machinery. Light perceived by ipRGCs
projects to the SCN via the retinohypothalamic tract (RHT), releasing glutamate
and pituitary adenylate cyclase-activating polypeptide (PACAP) [34]. These
ligands activate N-methyl-D-aspartate receptors and Pac1 receptors, respectively,
triggering Ca2+ influx and subsequent activation of Ca2+-calmodulin kinase II and
cyclic adenosine monophosphate (cAMP) response element binding protein (CREB)
[34]. The SCN orchestrates rhythmic gene expression, including genes encoding
core clock proteins (period circadian regulator (PER) 1,2,3, cryptochrome
circadian regulator (CRY)1,2, circadian locomotor output cycles kaput (CLOCK),
brain and muscle ARNT-like 1 (BMAL1)) that form a major oscillator driven by
transcriptional feedback loops. Timed LE is essential to entrain and maintain
robust oscillations. Circadian rhythms within the SCN that feedback with the
pineal gland could be fine-tuned by melatonin, which acts via the cerebrospinal
fluid on the SCN and pars tuberalis, influencing transcriptional feedback loops
and proteasomal degradation of clock proteins [35]. Such interaction of melatonin
with the proteasome can be important to adjust the SCN clock response to
dynamically changing light conditions [35]. Furthermore, the nuclear receptors
REV-ERBs (REV-ERB
Post-transcriptional and post-translational mechanisms. The molecular response to light is further fine-tuned post-transcriptionally and post-translationally via microRNAs (miRNAs) [54]. LE can modulate the availability of clock mRNAs through miRNA-mediated interference with AU-rich elements. For instance, light-sensitive miR-132 provides an additional layer of regulation on clock gene expression [55, 56], including seasonal variations in LE [56]. The expression of miR-132 is linked to the transcription factor CREB, which is activated by light signaling. A recent study identified novel specific light-responsive miRNAs that target clock genes, particularly the expression of cryptochrome genes in zebrafish [57], suggesting potential therapeutic targets for human circadian rhythm disorders and applied strategies to correct such disorders by manipulating LE.
Light therapy. Phototherapy is currently used to treat mood disorders (depression, seasonal affective disorder), sleep disorders (insomnia), skin conditions (psoriasis, eczema, acne), and neonatal jaundice. Recent studies [55, 58, 59] document possible new usages in other fields, notably in cardiology: the application of intense light was used as anticoagulant therapy and as a means to reduce infarct sizes following myocardial ischemia by enhancing the circadian amplitude of PER2. Beyond retinal pathways, light can also influence circadian rhythms via non-retinal mechanisms, such as through the skin. Within the context of circadian rhythmicity, ultraviolet B (UVB) radiation drives vitamin D synthesis [60, 61], while photobiomodulation, employing infrared (IR) light [62, 63], directly modulates cellular processes, such as mitochondrial activity or gene expression.
Light hygiene for cardiometabolic health. Modern lifestyles, characterized by inadequate exposure to natural daylight and excessive exposure to artificial light, significantly affect appetite and meal timing [64, 65]. This chronic disruption of circadian rhythms is associated with an increased risk of metabolic and cardiovascular disorders [28, 66]. Specifically, poor circadian light hygiene is associated with cardiometabolic dysfunction, increased body mass index, altered blood glucose and lipid profiles [2, 14, 15, 18, 26, 27, 67], contributing to the concept of “circadian syndrome”, a refinement of “metabolic syndrome” [68, 69]. While understanding the molecular mechanisms underlying circadian rhythms is crucial, clinical implications are paramount. Obesity, a pandemic comorbidity closely related to circadian disruption, highlights the importance of considering the broader physiological effect of light hygiene, beyond its cellular mechanism. Obesity is often linked to hormonal and cardiovascular complications, including altered circadian patterns of hormones which are in phase with melatonin, e.g., leptin, ghrelin and growth hormone secretion [70], further impacting metabolic health and cardiac function [71]. Improving circadian light hygiene to align with recommendations for 24-hour light exposure [2, 3] represents a promising strategy for mitigating cardiometabolic disorders and addressing the global challenge of obesity and its related comorbidities.
The heterogeneity in genetic susceptibility to light sensitivity supports the development of personalized interventions for light hygiene. Elucidating these intricate molecular pathways in further research should lay the foundation for personalized light hygiene approaches aimed at preventing and correcting circadian rhythm disorders, ultimately promoting overall health, well-being, and possible uses in sleep medicine, metabolic diseases and mental health.
LE, light exposure; BLE, blue light exposure; OPN4, melanopsin; OPN5, neuropsin; ipRGCs, intrinsically photosensitive Retinal Ganglion Cells; NMDA, N-Methyl-D-Aspartate receptor; MTNR1B, Melatonin Receptor Type 1B; AC, Adenylate cyclase; cAMP, cyclic Adenosine Monophosphate; PKC, Protein Kinase C; PKA, Protein Kinase A; Ras-Raf-MEK-ERK, Ras/Raf/Mitogen-activated Protein Kinase/ERK Kinase (MEK)/Extracellular-signal-Regulated Kinase (ERK); CaMK, Ca2+/Calmodulin-Dependent Protein Kinase; RKIP, Raf Kinase Inhibitory Protein; CREB, cAMP Response Element-Binding protein; miR-132, microRNA 132; ROR, Retinoic acid receptor-related Orphan Receptor; REV-ERBɑ, Nuclear Receptor Subfamily 1 Group D Member 1; SCN, Suprachiasmatic Nucleus; PACAP, Pituitary Adenylate Cyclase-Activating Polypeptide.
Conceptualization, DG; writing—original draft preparation, DG; draw figure, writing—review, revision and discussing, DG, OS and GC; supervision and resources supply, DG; funding acquisition, DG. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
The study was supported by the West-Siberian Science and Education Center, Government of Tyumen District, Decree of 20.11.2020, No. 928-rp.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.