



1 Department of Health Sciences and Sport Medicine, Hungarian University of Sports Science, 1123 Budapest, Hungary
2 Department of Sports Medicine, Semmelweis University, 1122 Budapest, Hungary
Abstract
The Piezo2 transmembrane proteins were identified by Ardem Patapoutian and his team. They also found that Piezo2 is the principal mechanosensory ion channel responsible for proprioception. Even before the Nobel Prize was awarded to him, it was proposed that these Piezo2 channels could sustain acquired microdamage at the proprioceptive somatosensory terminals under allostatic stress. Moreover, the principality of Piezo2 is suggested to extend beyond its physiological function, highlighting its relevance in the context of microdamage as well. Hence, acquired Piezo2 channelopathy is proposed to constitute one principal gateway to pathophysiology underpinned by proton affinity, energy metabolism and a proprioceptive pathway switch. The differentiating incomparable hallmark of Piezo2 is theorized to be a low-frequency semiconductor Schottky barrier diode-like feature that provides proton handling for quantum tunnelling and ultrafast long-range signalling to the hippocampus. Accordingly, even the proposed acquired Piezo2 channelopathy is also enigmatic by causing the impairment of this Piezo2-initiated ultrafast proton-based long-range signalling and proper synchronization to the hippocampus. The revealing of this protonic word and the ultrafast long-range signalling within the nervous system and its microdamage brings an entirely new perspective in medicine with the interpretation of the quad-phasic non-contact injury model. This is why this Piezo2 microdamage has been coined as the primary damage or the root cause of ageing. Paired-associative electromagnetic stimulation appears to be a promising treatment method and heart rate variability detection could be used for diagnosing autonomic nervous system disbalance as one symptom of this proposed Piezo2 channelopathy.
Keywords
- Piezo2
- channelopathy
- proprioception
- proton
- ultrafast long-range neurotransmission
- quad-phasic non-contact injury model
The Piezo2 transmembrane proteins were demonstrated to be the principal mechanotransduction channels for proprioception [1]. However, the principality of Piezo2 ion channels may also be reflected in the proposition that Piezo2 containing somatosensory neurons are carrying “special genetic signature” [2]. Recently it was addressed that these Piezo2 channels could go through a microdamage in an acute stress response (ASR) time interval or under allostatic stress. This may evolve when unaccustomed and/or strenuous repetitive forced lengthening (eccentric) contractions result in hyperexcitation, neuro-energetic depletion and eventually in the mechano-energetic impairment of somatosensory neuron terminals contributing to proprioception [3].
The realization that terminal arbour degeneration (TAD) like lesion [4] of proprioceptive terminals could be analogous to an ASR induced neural mitochondria energy depletion associated functional microdamage of the Piezo2 channels, lead to the theory of acquired Piezo2 channelopathy or the primary damage [3, 5]. Accordingly, this intrafusal primary damage induced impaired proprioception results in a more severe secondary extrafusal microdamage if the forced lengthening contractions are prolonged. Consequently, it leads to harsher peripheral tissue microdamages in a bi-phasic manner and to axonopathy [3, 5]. Hence, the loss of primary proprioceptive protection is like the loss of a protective airbag function.
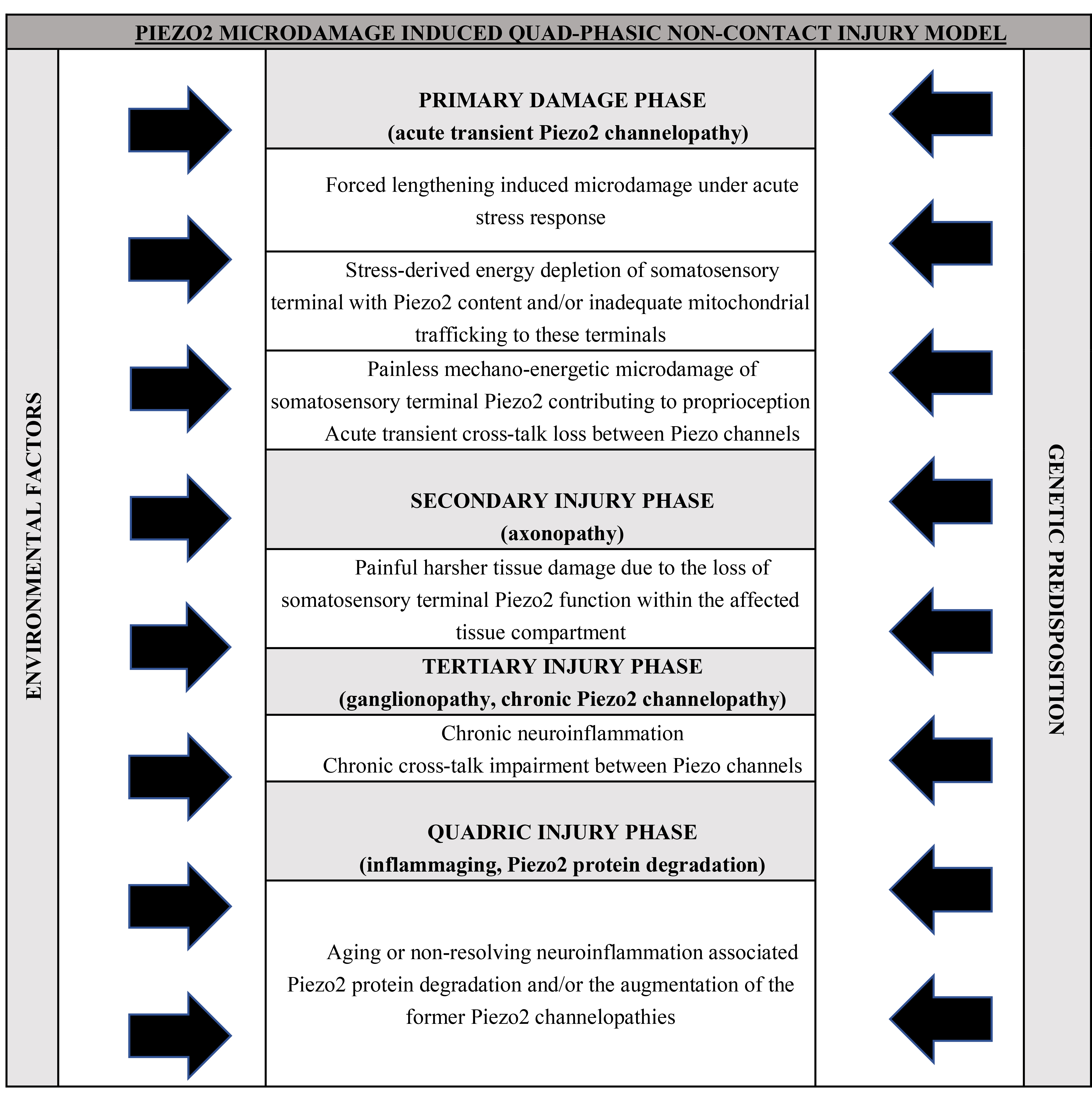
Later on, this bi-phasic bi-compartmental non-contact microinjury model, involving both the selective barrier protected intrafusal space and the extrafusal compartment as first suggested in delayed onset muscle soreness (DOMS) [3], was extended to the concept of the quad-phasic non-contact injury model (Table 1, Ref. [6, 7]). In this model the tertiary damage phase is the repeated microdamage of Piezo2 ion channel, suggested to be represented in a ganglionopathy (Table 1). The quadric stage is the impact of inflammaging or the ageing associated augmented low-grade neuroinflammation in the central nervous system (Table 1) [6, 7]. This quad-phasic non-contact injury model (Table 1) could explain numerous clinically unexplained conditions and diseases. These are mostly emerged in the 20th century as epidemics of modern times due to our insatiable appetite for growth, overreaching behaviour and resultant life style changes that created perplexing challenges to our cells, tissues, organs and to our body, but most importantly to our nervous system.
Not to mention that the principality of Piezo2 may also be presented in its acquired channelopathy, since it is theorized to impaire the unaccounted ultrafast long-range oscillatory synchronization between the periphery and the hippocampus [8, 9]. In addition, this acquired Piezo2 channelopathy on somatosensory terminals contributing to proprioception is also suggested to be a principal transcription activator [7], underpinning the suspected “special genetic signature” feature [2]. Furthermore, this primary damage could be depicted as the breach of the limits of remodelling [9] or proper wound healing, by initiating the inflammatory and gateway reflexes [10]. Indeed, Piezo ion channels are at need to ensure effective inflammation and maintenance of selective barrier integrity in support of proper wound closure [11]. Moreover, wound healing last longer with Piezo1 than without it [12] and the involvement of neurons in the wound healing process will lead to a more sophisticated process [13]. Accordingly, it has been theorized that even the channelopathy of Piezo1 could eventually induce somatosensory Piezo2 channelopathy under allostatic stress [6].
Emerging Piezo related researches are pointing to the direction that the aforementioned quad-phasic non-contact injury model (Table 1) could describe the mechanism of several unexplained conditions and diseases. Hypertension [14, 15, 16], diabetes [17], autoimmune conditions [3, 10] and degenerative conditions, leading to osteoarthritis [18], cardiovascular [19, 20] or neurodegenerative diseases [9] and even cancer [21, 22, 23] could fit into this Piezo2 implicated model. However, it should not be unnoticed that these primary non-contact microinjuries of Piezo2 exerted a concomitant beneficial neural training and adaptation effect on our body in order to satisfy our increasing life-style demands. In support, the mechanism underpinning this impairment is learning and memory related and construct the basis for remodelling and regeneration [9].
After all, the current paper puts into perspective that going overboard beyond the limits of proprioception, or more precisely the proper functionality of Piezo2 channels, is the equivalent of dancing on the edge of one principal crossroad between physiology and pathophysiology or the gateway to primary damage. Noteworthy, the unresolved “primary damage” is considered in science as the root cause of the ageing process [24].
This perspective review paper does not have the intention to dispute any former theories. It is rather about reversing the eyesight. Hence, looking at the pathomechanism of certain medical conditions and diseases from a neurocentric view may provide us with a better understanding than the formerly used view in reference to peripheral tissues. Accordingly, tissues like muscles, bones, skin and joints are ‘only’ neuromodulators in this interpretation. In contrast, the non-contact injuries are primarily initiated by autonomously acquired Piezo2 microdamage of the innervating somatosensory terminals contributing to proprioception. After all these mechanosensitive ion channels are likely the first ones under certain circumstances, like ultradian events, to translate external physical cues into inner chemical and biological ones. Correspondingly, ultradian events likely activate proprioceptive Piezo2 ion channels first in order to serve the readiness for unpredictable changes [9].
Moreover, the allostatic stress related microlesion of the Piezo containing sensory nerve afferent terminals innervating smooth muscles and myocardial muscles could also serve as principal gateways to pathophysiology through the impairment of the cross-talk between Piezo ion channels [8]. Noteworthy that a cross-talk between Piezo2 and Piezo1 has been suspected earlier [6, 25, 26], but lately it is suggested to be cross-coupled by Huygens synchronization of mitochondria and protons [8, 9]. Notable, that this interplay may exist even on single neurons, since Piezo1 is co-expressed with Piezo2 in small diameter peripheral pain neurons [27].
The gateway is suggested to be the acquired Piezo2 channelopathy and this is the locus where low grade neuroinflammation could be initiated by the inducement of the inflammatory and gateway reflexes, and potentially escort us throughout life. This low grade neuroinflammation could be augmented and may become progressive depending on repeated microdamage of Piezo2, environmental risk factors and genetic predisposition, as proposed by the aforementioned quad-phasic non-contact injury model (Table 1) [6, 7].
In summary the current paper introduces how the repeated microdamage of Piezo2 in an acquired fashion could cause the new epidemic of non-communicable diseases of our times.
Ardem Patapoutian and his team [28] discovered Piezo ion channels and he received the Nobel Prize for this significant finding. Piezo proteins are the largest transmembrane proteins known so far with numerous transmembrane segments [29]. However, the precise topology and function of these Piezo proteins is far from entirely understood [29]. The currently known physiological functions of Piezo ion channels in mechanotransduction could be acquainted from the comprehensive review of Bailong Xiao [30] who was also a former member of the Nobel laureate’s team. Ardem Patapoutian and his group also identified that Piezo2 is the principal ion channel responsible for proprioceptive mechanotransduction [1]. Noteworthy that Piezo2 is not the sole proprioceptive ion channel, however it is the only one suggested to initiate the earlier mentioned unaccounted long-range protonic ultrafast oscillatory neurotransmission to the hippocampus [8, 9]. In support, proprioceptive neural signalling is not abolished even in the absence of Piezo2 initiated ultrafast signaling, but taken over for example by Nav1.1 ion channels in a miswired fashion [31].
The in-depth mechanism theory of Piezo2 channelopathy could be acquainted from earlier papers [3, 5, 7, 9]. Noteworthy that ion channel microdamages or channelopathies could be either inherited or acquired. The examples for the inherited PIEZO2 channelopathies are like distal arthrogryposis type 5 with a gain-of-function mutations on the PIEZO2 gene [32], but Gordon syndrome and Marden-Walker syndrome are also inherited channelopathies of Piezo2 [33]. However, the subject of this manuscript is rather about the theoretical acquired form of Piezo2 channelopathy that was first suggested in a strange DOMS related syndrome, called post orgasmic illness syndrome (POIS) [3]. This non-contact or autonomous Piezo2 channelopathy could develop transiently, as is suggested in the primary damage phase of DOMS, but may evolve into a chronic state as well [3]. Two years later a research team involving the aforementioned Nobel laureate Ardem Patapoutian confirmed that Piezo2 indeed essential for sexual function [34], further underpinning Piezo2’s theoretical role in POIS. Moreover, another study also showed that excessive mechanotransduction on Piezo2 containing sensory neurons causes joint contracture [35]. This finding could be analogous to the theoretical acquired Piezo2 channelopathy induced exaggerated contractions in DOMS [3]. Later, even a third form of acquired Piezo2 channelopathy was proposed, the irreversible and progressive one, in amyotrophic lateral sclerosis (ALS) pathomechanism [7]. After all, the suggested acquired Piezo2 channelopathy could evolve into an acute transient, a chronic or even to an irreversible progressive protein microdamage depending on repeated microdamage, environmental risk factors and genetic predisposition [7].
Nevertheless, there is no unquestionable direct electrophysiological evidence of this structural and/or functional microdamage of Piezo2, but the indirect findings are emerging. For years, the scientific community was sceptical whether these proteins could endure a microdamage, as was suggested first in 2021 [3], but lately more and more scientists raise this possibility [17, 36]. Furthermore, a recent paper stressed that the protonation of Piezo2 should be considered [37]. Indeed channel inactivation is stabilized with the protonation of Piezo1 [38]. In addition, a former genetically engineered double mutation research demonstrated that translocating arginine with high proton affinity characteristics from extracellular to intracellular domains of the cellular membrane on particular structural sites eliminates Piezo1 function [39]. This translocation is translated as a “proton affinity switch”, leading to proton reverse flux to intracellular domains [37] that in fact exists in nature as an autonomous light-driven proton pump of bacteriorhodopsin [40]. Moreover, a chemotherapeutic agent, called oxaliplatin, may also instigate a similar “proton affinity switch” on Piezo2 in an acquired fashion, resulting in the channelopathy of Piezo2 and neuropathy, since platinum also has high proton affinity [37].
Accordingly, Waxman et al. [41] theorized earlier that alterations of voltage gated Na+ (Nav) channels could follow after nerve injury. This theory was based on their findings that nerve damage altered the transcription of various Nav channels. One key hallmark of Piezo2 is its burst activation that instigates proprioceptive signalling in primary sensory afferents. The instantaneous activation of Piezo2 contributes to maintained and amplified consistent firing of Nav1.1 channels [31]. Consequently, Nav1.1 is required for suitable proprioceptive signalling when Piezo2 is inactivated under prolonged muscle stretch [31]. When low frequency power of heart rate variability decreases close to unnoticeable level during acute intensive exercise is hypothesized to be such a Piezo2 inactivation moment. It has been also suggested, as referred earlier, that Nav channels could gain control over proprioception in a lessened way on primary afferents in the absence of Piezo2 activity [8]. In support, likely pathogenic variants of Nav1.1 ion channel encoding genes were identified in ALS where Piezo2 channelopathy is proposed to be irreversible [42]. Noteworthy that earlier research showed that mechanosensitive channels are indeed vulnerable to irreversible loss of rapid kinetics caused by membrane-cytoskeleton decoupling due to mechanical over stimulation [43, 44].
Important consideration of the acquired Piezo2 channelopathy theory is the role of auxiliary subunit proteins to Piezo2. MyoD-family inhibitor proteins are such proteins [45] and the conformational alteration of this protein-protein interaction might also have a role why Piezo2 is coined as being a principal transcription activator [9]. Another one is the transmembrane protein 120A (TMEM120A), or TACAN, that inhibits Piezo2 [46]. TACAN is associated with leakage currents, even after stretch inducement, when they are transfected with Piezo ion channels [47], not to mention it is implicated in lipid metabolism and innate immune response regulation [46]. The inducement of these leakage currents may have relevance when protein-protein interaction goes through conformational changes under shear stress induced microdamage, possibly leading to the structural and functional dissociation from Piezo2 [9]. Correspondingly, the Piezo2 channelopathy theory indeed involves the inducement of imbalanced subthreshold leakage currents [3] and the dysregulation of lipids and the innate immune system [3, 48].
Not only protein-protein interaction could be altered under allostatic stress, but protein-lipid interaction as well. It has been observed that negatively charged membrane lipids are densely neighbouring the unique propeller blade structure of Piezo2 [49, 50, 51]. The depletion of phosphatidylinositol 4,5-bisphosphate (PIP2) induced activation of phospholipase C (PLC) and resultant transient receptor potential cation channel subfamily V member 1 (TRPV1) activation inhibits Piezo channels [52]. Moreover, phospholipase D (PLD) produces phosphatidic acid and it is also shown to inhibit Piezo2 [53]. It is known that mechanical shear stress could disrupt the nanodomains of lipid rafts, leading to the interplay between PLD and activated PIP2 [54]. Furthermore, it should not be forgotten that PLD and phosphatidic acid might have a role in the biogenesis and cargo loading of extracellular vesicles [55], since the Piezo2 channelopathy theory entails the impairment of the glutamate vesicular release machinery [3, 7].
These protein-lipid interaction findings are noteworthy because it seems that PLD might have role in the inhibition of Piezo2 on Type Ia proprioceptive terminals in the intrafusal space and PLC may have an inhibitory role of both Piezo1 and Piezo2 in the extrafusal space. In support, a recent paper emphasized the role of Piezo2 and atypical hippocampal-like metabotropic glutamate receptors coupled to PLD containing proprioceptive primary afferent terminals in the Piezo2 initiated proton signalled long-range ultrafast synchronization to hippocampal theta rhythm (Fig. 1, Ref. [56]) [8, 9].
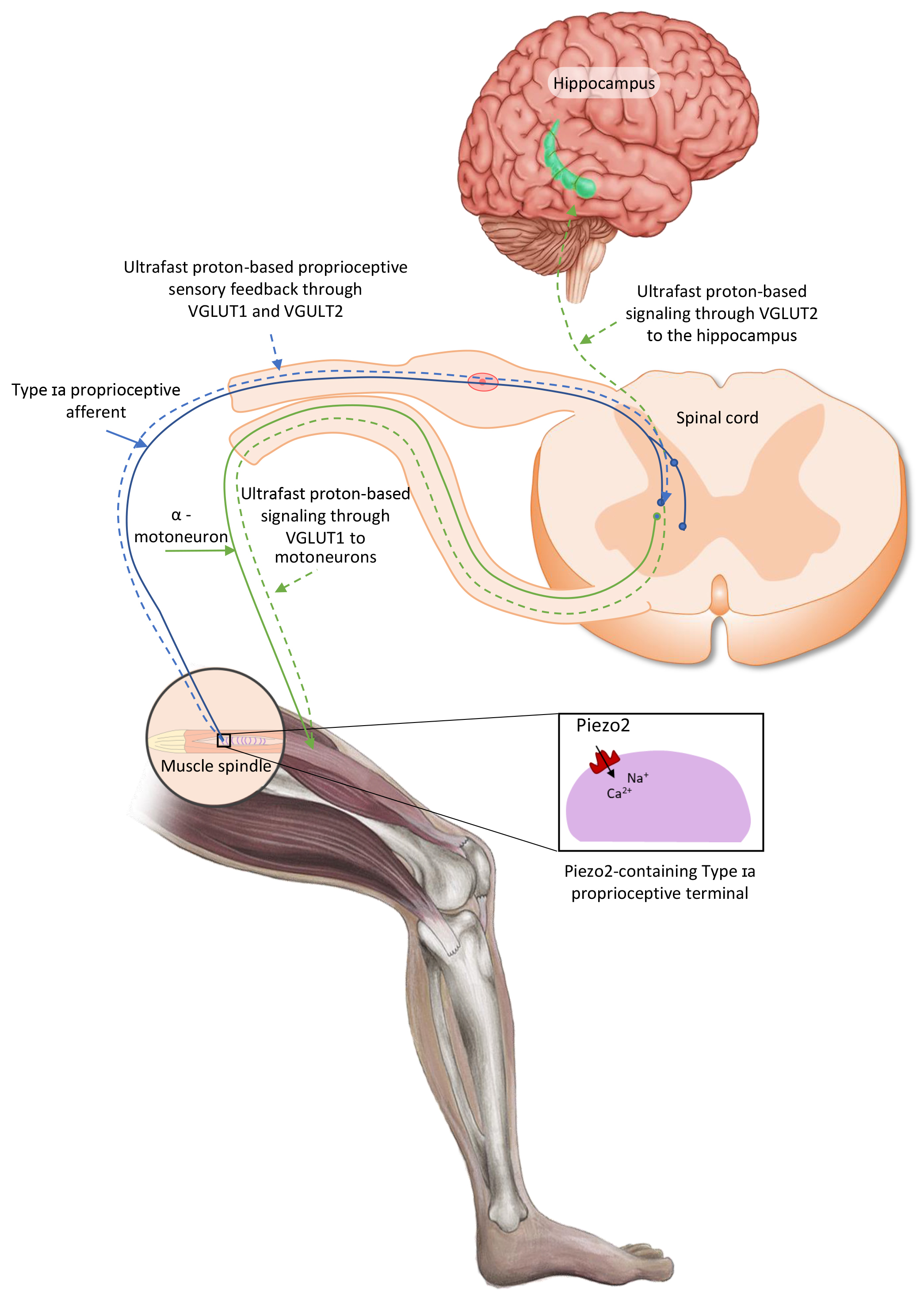 Fig. 1.
Fig. 1.
Proposed intrafusal proprioceptive terminal Piezo2 initiated ultrafast proton-based long-range synchronizational signalling to hippocampal theta rhythm through vesicular glutamte transporter 2 (VGLUT2). The current figure is an English adaption of the figure from Magyar Reumatológia based on their permission [56].
The suggested acquired Piezo2 channelopathy impairs this ultrafast long-range proton-based neurotransmission, as is suggested in DOMS and ALS [8, 9]. Indicative of the consequential proprioceptive miswiring that one research shows that submaximal eccentric contractions causes immediate voluntary activation deficits and alters movement sense [57]. Da Silva et al. [57] interpreted their results as these eccentric contractions induced central alterations that impacted somatosensory function. However, the current author proposes that this observation is rather the result of acquired Piezo2 channelopathy induced impaired ultrafast long-range proton-based oscillatory synchronization meant to arise from the muscle spindle to the hippocampus. Moreover, this Piezo2 channelopathy could be the initiating peripheral impaired input source that drives central sensitization of spinal nociceptive neurons [58, 59].
Significant finding that paired associative transcranial and peripheral electromagnetic stimulation is shown to alleviate DOMS effect and improve force generation [60]. Even more interesting that peripheral electromagnetic stimulation alone was inefficient as a treatment method in DOMS [61]. This non-invasive neuromodulation treatment highlighted the neurogenic pathophysiology of DOMS, as was theorized by its acute compression axonopathy theory [5]. It is suspected that a common receptor exists for the detection of electromagnetic field induced oscillating energy and this receptor could activate biological pathways at various range of frequencies and interfacial water has a central role in this modulation [62]. This common receptor is theorized by the current author to be the Piezo2 ion channel at somatosensory nerve terminals with capabilities of providing proton handling, in collaboration with acid-sensing ion channel 2 (ASIC2) [9], for quantum tunnelling in support of the proposed ultrafast signalling [8, 9].
However, Piezo2 channels and its auxiliary proteins and surrounding lipids may go through conformational changes under allostatic stress. These secondary structure formations are capable of regulating transcriptional or post-transcriptional processes, as suggested by Perez et al. [62], in association with the aforementioned undesired functional disruptions or structural dissociations [8, 9]. Worthy to note again that this is in line with the Piezo2 channelopathy theory that states it is a principal transcription activator besides being the primary damage [7].
Moreover, the critical cell membrane stress buffering role of heat shock protein 70 (HSP70) should be noted [63]. Repeated electromagnetic stimulation activates heat shock transcription factor 1 (HSF1) and as a result HSP70 chaperon level is increased in order to prevent cell death [62]. In support, the increase of HSP70 level has relevance in DOMS pathomechanism [64] and HSF1 transcription factor is protective in neurodegenerative diseases, like Alzheimer and Parkinson’s disease [62] or ALS [65]. Remarkable that HSF1 is a long non-coding RNA [53] and long non-coding RNAs are contributing largely to the genetic background of non-familial neurodegenerative diseases and they seem to initiate a remodelling problem [9]. This remodelling problem could be the basis of Piezo2 channelopathy induced impaired Piezo crosstalk and unfinished wound healing [6].
The Piezo2 channelopathy theory entails that this type of microinjury could evolve in axon terminals where the energy demand is high, hence increased abundance of mitochondria is needed [3]. Proprioceptive terminals under unaccustomed and/or strenuous forced lengthening contractions are proposed to be such loci under hyperexcited mechanotransduction induced allostatic stress. Moreover, the conformational changes of these packed mitochondria in these terminals are also part of this theory [8]. Accordingly, “crystal-like” pattern of mitochondria may have a role in high frequency oscillation generation induced by Huygens synchronization mechanism [8]. Huygens synchronization is crucial on pacemaker cells of the hippocampus in order to generate theta oscillation [66]. In addition, mitochondria could also take a radically symmetric pattern and these mechanisms are coupled [8]. However, it is theorized that excessively prolonged proprioceptive mechanostransduction under allostatic stress induced Piezo2 channelopathy, or the undesired intracellular proton leak, may perturb the energy supply of mitochondria to the point that is not sufficient to operate the Piezo2 ion channel properly vice versa [8].
Noteworthy that depletion of phospholipids has relevance not only in the vicinity of Piezo2 in the proprioceptive terminal membrane, but in mitochondrial phospholipid transport as well [67]. Since these mitochondria are not capable of synthesizing adequate amounts of phospholipids, therefore their supply is dependent on phospholipid import from interorganellar contact sites through phospholipid import pathways [67]. This coexistence is proposed to be symbiotic to the point when excessively prolonged proprioceptive mechanostransduction under allostatic stress dysregulate this homeostasis. As a result, the depletion of proprioceptive terminal membrane cholesterol and phospholipids [7], and the impairment of phospholipid transport pathways in mitochondria may evolve. This impairment could be a charge altering switch in this micro/nanomilieu, since these phospholipids contribute to the negatively charged membrane lipids in a homeostatic state.
The referred structural dissociation phenomenon is in line with the Hofmeister effect by a salting-out mechanism. This effect increases the surface tension of interfacial water, destabilizes proteins due to the insufficient stabilizing phospholipid bilayer [68]. As a result, the Piezo2 channels may not be able to provide the theorized low-frequency Schottky barrier semiconductor diode-like function that could be needed for the ultrafast long-range Piezo2 initiated, proton handling, quantum tunnelling, frequency coupled synchronization to hippocampal theta rhythm from proprioceptive terminals [8, 9]. Indicative research findings are arising like, Piezo2 presents a voltage-block in cell membranes and it is highly attuned by membrane voltage [69]. Therefore, the aforementioned conformational changes of negatively charged membrane lipids in the close vicinity of Piezo2 may have high relevance not only in proton handling, but in the voltage-block under allostatic stress. Noteworthy again, that Piezo2 is theorized with a low-frequency Schottky barrier semiconductor diode-like feature [8] and the negative voltage across diodes is important in order no current could flow. Piezo2 resembles to Schottky diodes in its very fast switching action, too. The role of Nav channels in the membrane voltage modulation should be considered as well. However, the loss of the negative charge neighbouring Piezo2 may impair this voltage-block function.
Another supportive finding showed that the supplementation of eicosapentaenoic acid rich fish has a beneficial effect against DOMS evolvement, motoneuron and muscle damage [70]. This beneficial effect of eicosapentaenoic acid was devoted theoretically to the preservation of proper Piezo2 function [5] and later proven by research [26]. It is also highly indicative of the Piezo2 channelopathy theory that linolenic acid supplementation in animal models of Angelman syndrome, a neurogenetic disorder with a mutation on another gene than Piezo2, improves Piezo2 dysfunction [71] or the proposed acquired Piezo2 channelopathy.
Further important consideration in the activation of Piezo2 is the increased blood flow induced shear stress and arterial pressure pulsation. An interesting new study shows that Piezo2 may induce an intrinsic oscillatory interoceptive mechanism through a pressure pulsation transduction pathway that modulates olfactory bulb activity in arousal and it is synchronized to brain activities [72]. This Piezo2 initiated distant synchronization was shown earlier within the brain [73] and theorized between the periphery and the brain [8]. The finding of Jammal Salameh et al. [72] not only shows the importance of Piezo2 in interoceptive arterial pressure pulse detection, but the oscillatory synchronization mechanism is analogous to the aforementioned theory that may play a role in DOMS and ALS pathomechanism [8]. Moreover, it has been known in reference to DOMS mechanism that blood flow restriction could induce muscle damage and repeated bout effect (RBE) [74]. Noteworthy that RBE is suggested to be the tertiary injury phase of DOMS according to the Piezo2 channelopathy theory [3, 5]. The author of this manuscript suggests that the muscle damage could be explained by the blood flow restriction induced lost interoceptive pressure pulse detection and lost Piezo2 initiated oscillatory synchronization mechanism. As a result of this Piezo2 associated intrafusal proprioceptive protection loss and resultant lost oscillatory synchronization and hypoxia, the secondary damage phase could be experienced in the form of extrafusal muscle damage.
This interoceptive pressure pulse induced Piezo2 activation may have high relevance under acute intensive exercise activities when the blood flow is significantly redistributed towards muscles and the visceral organs are less supplied. The current author suggests that this mechanism could explain the Janus-faced feature of the acquired Piezo2 channelopathy, namely Piezo2 channelopathy is also a blood flow restriction dependent mechanism. In support the same Piezo2 channelopathy is suspected as the primary damage of DOMS [3, 5] and for example in psoriasis as well. Correspondingly, blood flow restriction could have high relevance in the Piezo2 channelopathy of visceral organs, like in Piezo2 containing enterochromaffin cells of the colon.
In essence there is a great need for the in depth understanding of the conformational changes of the protein-protein and protein-lipid interaction induced proton affinity switch and the charge alteration in the close vicinity of Piezo2 under allostatic stress. The structural alterations of interfacial water, protein crystals, membrane lipids, proton affinity and the protonation state changes could be of special interest. Hence, electrostatic change effects on protein-protein interaction, negatively charged membrane lipids and on glycoproteins of the extracellular matrix, like, e.g., syndecans [9], should be investigated and modelled. Non-contact injuries, like non-contact anterior cruciate ligament injury or DOMS [5], are common classification in medicine, but after all external physical cues do seems to matter at cellular, micro or even nano levels when these physical inputs are translated to biological and chemical signals during proprioceptive mechanotransduction. Piezo2 is suggested to be a principal player in this process and the direct or indirect overexertion of its functional and structural boundaries of this protein under allostatic stress could lead to the proposed initiating primary damage.
Piezo2 channelopathy may not only come with a proprioceptive miswiring or switch [7], but may also come with the impairment of the astrocyte-neuron lactate shuttle (ANLS) like mechanism in association with the impairment of the glutamate vesicular release [75]. The inducement of the ANLS in the central nervous system is favourable for neurons under allostatic stress. It has been hypothesized that an analogous ANLS-like mechanism exists intrafusally as well in order to nourish proprioceptive neurons when glucose is in shortage [75]. However, when Piezo2 channelopathy evolves then it may also impair the glutamate vesicular release system and the ANLS-like machinery, leading to glutamate excitotoxicity [3, 75]. After all, this is why lactate might have a role in DOMS inducement within the muscle spindle [75] and glutamate excitotoxicity is implicated in ALS pathomechanism. In line with this energy metabolism switch, it was demonstrated in DOMS that the higher was the muscle damage, the greater was the insulin resistance, but the lipid markers showed a positive response [76]. These metabolic alterations could be invoked by RBE as well in a smaller scale [76]. However, the current author proposes that prolonged frequently repeated damage of Piezo2 under allostatic stress without any regeneration periodization could lead to further dysregulation of this energy metabolism switch and may culminate in insulin resistance and metabolic syndrome in the presence of environmental risk factors and genetic predisposition. Important to note that under stress a metabolic switch could facilitate fructose production from glucose. Moreover, salts may also induce this metabolic switch [77]. The current author suggests that these mechanisms may contribute for example to DOMS pathomechanism in an unaccounted way leading to proposed impaired proton-based long-range ultrafast neurotransmission. Accordingly, the Piezo2 channelopathy induced lost Piezo cross-talk may increase insulin secretion transiently from beta cells of pancreas in a compensatory fashion [8, 9]. However, the chronic form of Piezo2 channelopathy, as the tertiary damage phase, may instigate insulin resistance and metabolic syndrome as a common underlying risk factor for the aforementioned unexplained non-communicable disease epidemic of our times. Furthermore, the acutely lost Piezo2-Piezo1 cross-talk might disrupt the homeostatic osmotic pressure change regulation [6], likely contributing to the salting-out of the before mentioned Hofmeister effect around Piezo2. Nevertheless, the chronic Piezo2 channelopathy may facilitate lipid accumulation at the affected loci in order to preserve water due to the dysregulated osmotic pressure change regulation. This metabolic switch and the consequential metabolic dysregulation on the chronic path, not only could explain ALS [9] and the common denominator of neurodegenerative diseases, depending on the evolvement loci of Piezo2 channelopathy, but could explain the one common initiating cause for the unexplained recent epidemic of non-communicable disease or the primary damage induced accelerated ageing.
Finally, the current author proposes that the aforementioned critical energy generation switch and the associated impairment of the ANLS-like mechanism at Type Ia proprioceptive terminals in DOMS [9] is likely underpinned by an alternative, evolutionarily lower, energy generating pathway than the mitochondrial oxidative phosphorylation system (OXPHOS). OXPHOS energy generation yields adenosin-5′-triphosphate (ATP) production due to energy from transfer of electrons and proton motive force. Correspondingly, when the aforementioned auxiliary subunits of Piezo2 may undergo conformational changes under allostatic stress leading to Piezo2 channelopathy then the inward leakage of extracellular protons prevails [37]. Therefore, this primary damage impairs the low-frequency Schottky barrier semiconductor diode-like function of Piezo2 [8], not to mention it also impairs the intracellular electrochemical proton gradient [37]. Despite neural glutamine uptake, as part of the ANLS-like machinery, is at no need for proton availability, but the low pH [78] and the impaired intracellular and transmembrane proton gradient could switch and load mitochondrial energy production from evolutionarily superior OXPHOS and glutamine respiration pathways to evolutionarily lower mitochondrial glucose and even more importantly to glutamine fermentation pathways. These respirofermentation pathways are often paralleled in fast growth milieu, like in cancer or immune cell activation [79]. Sonkodi et al. [48] emphasized this distinct metabolic programming of natural killer T (NKT) cells and their dysregulated increase in response to DOMS inducing exercise.
In summary the impairment of the aforementioned ANLS-like mechanism at Piezo2 containing Type Ia proprioceptive terminals in the primary damage phase of DOMS mechanism [75] may be explained by the aforementioned energy generating deficit, propelled by glutamine fermentation. The energy production derailment is supported by the damage of the skeletal muscle mitochondria-related genes in DOMS [80]. However, the onset of mitochondrial damage is likely initiated at the intrafusal proprioceptive terminals in the primary damage phase of DOMS, because the proprioceptive switch involves more neurons and muscle cell activation in circuitries as a compensatory mechanism, hence demanding more energy due to the primary damage.
Piezo2 channelopathy is suggested to initiate a low grade neuroinflammation [3, 5] that is evident in the aforementioned unexplained conditions and diseases of our modern times, like in DOMS [5]. Moreover, the representation of Piezo2 expression is often associated with reflexes, hence Piezo2 channelopathy is a good candidate to induce the so-called inflammatory reflex transiently or the gateway reflex on the chronic path [10]. Noteworthy, the gateway reflexes are neuroimmune homeostatic gatekeepers through neural circuits in organs with blood barriers [81], like it could be the case in muscle spindles with their selective barrier [5].
Furthermore, Piezo2 channelopathy is proposed to affect the neuroimmune response or the initial steps of the innate immune response, with the likely involvement of TACAN [46], leading to the elevation and imbalanced control of NKT cells in its acute form [48]. However, chronic Piezo2 channelopathy depletes these NKT cells, leading to autoinflammatory conditions and autoimmune diseases [10]. In contrast in the third, progressive and irreversible form of Piezo2 channelopathy, suggested in ALS, increases NKT cell production due to the progressive nature of the disease trajectory, however with lost regulation [9]. Moreover, Piezo2 channelopathy induced impaired Piezo2-Piezo1 cross-talk could activate dendritic cells with Piezo1 downregulation and satellite/astrocyte cells with Piezo1 upregulation in the affected organs or compartments with selective barriers [8].
Another important consideration that Piezo2 channelopathy is also proposed to dysregulate the autonomic nervous system [3] due to the impairment of the Piezo2-Piezo2 and Piezo2-Piezo1 cross-talk [6]. Correspondingly, it is theorized that the low frequency power of heart rate variability substantially represents the Piezo2 activity level in the baroreceptors [8]. Indeed, a pilot study showed that orthostatic tolerance is impaired in DOMS, not to mention that this orthostatic impairment resembles to the one experienced in diabetes, indicating the aforementioned metabolic switch [82]. Notable that all the implicated Piezo2 channelopathy induced conditions and diseases come with dysautonomia. Promising recent finding showed in young athletes that paired-associative electromagnetic stimulation favourably impacts the autonomic nervous system and facilitates recovery in response to DOMS effect without causing any disarray in the typical DOMS recovery mechanism [83].
Finally, transient Piezo2 channelopathy without the secondary damage phase is suggested to be a pain free condition [3, 5]. The primary Piezo2 microdamage in association with the secondary damage phase, entailing a harsher tissue damage, suggested to cause delayed onset of pain sensation with the involvement of the wide dynamic range neurons (WDR neuron) on the spinal dorsal horn and other ion channels [7], like proposed in DOMS [3, 5]. Furthermore, it is important to note again that this Piezo2 channelopathy could be the initiating peripheral input source that drives central sensitization of spinal nociceptive neurons [58, 59]. Hence, central sensitization independent of peripheral input is likely not the autonomous pain generator [58, 59]. The chronification of Piezo2 channelopathy proposed to construct a pain spectrum with the before mentioned involvement of WDR neurons and other ion channels [7, 10]. However, the progressive and irreversible form of Piezo2 channelopathy could completely disrupt these dysregulated pain pathways [7, 8, 9].
Cognitive demand induced repetitive forced lengthening under allostatic stress could microdamage the functionality of Piezo2 channels at the somatosensory terminals that are contributing to proprioception. This allostatic stress related overexertion associated autonomous mechano-energetic Piezo2 channelopathy is proposed to be one principal gateway to pathophysiology or the primary damage. Moreover, the repetitive inducement of the primary damage without regeneration periodization potentially leads to a quad-phasic non-contact injury trajectory (Table 1) and that could be the pathway to the development of non-communicable diseases of our times. Important to emphasize, that exposure to environmental factors and genetic predisposition could be risk factors that might accelerate the proposed quad-phasic non-contact pathophysiological roadmap leading to accelerated ageing (Table 1).
This model is a reverse clinical view of the currently used on the periphery by looking at the pathophysiology from a neurocentric angle, as opposed to the view that is focusing only on particular tissue or organ dysfunction. Accordingly, Piezo1 content of peripheral cells are ‘only’ neuromodulators of Piezo2 channels of the innervating sensory terminals within a compartmental micromilieu where the gateway to pathophysiology is located, and the inflammatory and gateway reflex with associated low-grade neuroinflammation is initiated.
The current perspective outlook might allow to apply an interdisciplinary approach for the following reason: (1) it should be considered that mechanotransduction is the conversion of physics to chemistry and biology from a cellular to whole body level, (2) the mechanotransduction system is hierarchical [84], (3) the Piezo ion channels are evolutionarily highly conserved and (4) Piezo ion channels are burst activating mechanosensory channels [85]. Correspondingly, it is suggested that Piezo2 at somatosensory terminals are the first to be microdamaged if we apply a critical path method when forced repetitive lengthening induced non-contact injuries under allostatic stress evolves. The current author suggests that Piezo2 activation is a response to ultradian events that serves the readiness for unpredictable changes in adaptation and use the ultrafast long-range proton-based oscillatory neurotransmission in order to respond to these challenges instantaneously [8, 9]. Indicative that one recent study found that human physical activity is organized on a high-level of hierarchy [86], as the mechanotransduction system is hierarchical as well [84].
The current perspective outlook and model, substantiated by indirect findings, not only could promote Piezo related research, but critical path method mapping and mathematical algorithm formation are needed for the mathematical and computational modelling of the proposed potentially quad-phasic non-contact injury mechanisms (Table 1). This gateway will be a crossroad where mathematicians, engineers and programmers could and will help the clinicians’ work immensely as it is happening already in certain conditions and diseases, like in neurodegenerative diseases. However, these mathematical and computational modellings should be extended towards this principal gateway to pathophysiology in order to bring light clinically to the ‘no men’s land’ of peripheral Piezo2 containing somatosensory nerves. By doing so, numerous unresolved pathomechanism will be expected to be more enlightened, like DOMS, non-contact injuries, mild and blast traumatic brain injury, scoliosis, osteoporosis, asthma, hypertension, diabetes, obesity, immune conditions like POIS, non-coeliac gluten sensitivity, autoimmune conditions and diseases like psoriasis, rheumatoid arthritis, systemic lupus erythematosus, ankylosing spondylitis, not to mention cardiovascular diseases, neurodegenerative diseases or even cancer.
At the end, this proposed gateway to pathophysiology has not posed a clinical challenge, despite the primary damage is long-suspected, in the past since neither diagnostics, nor treatment methods were available. However, the demand for the prevention and treatment of the new epidemic of non-communicable diseases of our times is unprecedently on the rise. Moreover, the internet of things are generating immense longitudinal data in respect to the risk factors of the aforementioned diseases and data owners have an insatiable appetite for scientifically substantiated interpretations. Good news that paired-associative electromagnetic stimulation seems to be a promising treatment method to clinicians for the restitution of the suggested acquired Piezo2 induced impaired ultrafast proton-signalled long-range oscillatory synchronization to the hippocampus. Moreover, in depth evaluation of heart rate variability will likely gain more importance in the diagnosis of Piezo2 channelopathy due to its capability of detecting the disbalance of the autonomous nervous system.
ALS, amyotrophic lateral sclerosis; ANLS, astrocyte-neuron lactate shuttle; ASIC2, acid-sensing ion channel 2; ASR, acute stress response; ATP, adenosin-5′-triphosphate; HSF1, heat shock transcription factor 1; HSP70, heat shock protein 70; NKT, natural killer T; OXPHOS, oxidative phosphorylation system; PIP2, phosphatidylinositol 4,5-bisphosphate; PLC, phospholipase C; PLD, phospholipase D; POIS, post orgasmic illness syndrome; RBE, repeated bout effect; TAD, terminal arbor degeneration; TMEM120A, transmembrane protein 120A; TRPV1, transient receptor potential cation channel subfamily V member 1; WDR neuron, wide dynamic range neuron.
The single author had to the sole role in designing, data collection, analysis, and manuscript writing.
Not applicable.
Not applicable.
This research received no external funding.
The author declares no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.