





 , Rita Mohammad 1, Yuzhe Wang 1, Wenyu Xue 1, Philipp Malakhov 1, Viktor Nekrasov 1, Elizaveta Kontareva 1, Zain Nofal 1, Vyacheslav Saburov 3, Dmitry Kolesov 4,5, Andreyan Osipov 1,2, Sergey Leonov 1,6
, Rita Mohammad 1, Yuzhe Wang 1, Wenyu Xue 1, Philipp Malakhov 1, Viktor Nekrasov 1, Elizaveta Kontareva 1, Zain Nofal 1, Vyacheslav Saburov 3, Dmitry Kolesov 4,5, Andreyan Osipov 1,2, Sergey Leonov 1,61 Institute of Future Biophysics, 141701 Dolgoprudny, Russia
2 State Research Center - Burnasyan Federal Medical Biophysical Center of Federal Medical Biological Agency, 123098 Moscow, Russia
3 A. Tsyb Medical Radiological Research Center-Branch of the National Medical Research Radiological Center of the Ministry of Health of the Russian Federation, 249031 Obninsk, Russia
4 Laboratory of Scanning Probe Microscopy, Moscow Polytechnic University, 107023 Moscow, Russia
5 Institute of General Pathology and Pathophysiology, 125315 Moscow, Russia
6 Institute of Cell Biophysics of Russian Academy of Sciences, 142290 Pushchino, Russia
Abstract
The implementation of proton beam irradiation (PBI) for breast cancer (BC) treatment is rapidly advancing due to its enhanced target coverage and reduced toxicities to organs at risk. However, the effects of PBI can vary depending on the cell type. This study aimed to explore the effects of PBI on two BC cell lines, MCF7 and MDA-MB-231.
The relative biological effectiveness (RBE) of PBI was assessed using a clonogenic assay. DNA double-strand break (DSB) repair, epithelial–mesenchymal transition (EMT), and filamentous actin (F-actin) were evaluated using immunofluorescence analysis. The extent of entosis and the senescence-associated β-galactosidase (SA-β-gal) activity were estimated by cytochemistry analysis. The influence of the extracellular matrix was evaluated by cultivating cells in both adherent two-dimensional (2D) environments and within 3D fibrin gels of varying stiffness. The metastatic propensity of cells was investigated using migration tests and the cell encapsulation of carboxylate-modified fluorescent nanoparticles. The comparative tumorigenic potential of cells was investigated using an in vivo model of the chick embryo chorioallantoic membrane (CAM).
PBI demonstrated superior efficacy in eliminating MCF7 and MDA-MB-231 cells with RBE 1.7 and 1.75, respectively. Following PBI, MDA-MB-231 cells exhibited significantly lower clonogenic survival compared to MCF7, which was accompanied by the accumulation of phosphorylated histone H2AX (γH2AX), p53-binding protein 1 (53BP1) and Rad51 foci of DNA DSBs repair proteins. After surviving 7 days post-PBI, MCF7 cells exhibited 2.5-fold higher levels of the senescence phenotype and entosis compared to the MDA-MB-231 offspring. Both PBI-survived cell lines had greater capability for 2D collective migration, but their metastatic potential was significantly reduced. A significant influence of extracellular matrix stiffness on the correlation between F-actin expression in PBI-survived cells—an indicator of cell stiffness—and their ability to uptake nanoparticles, a trait associated with metastatic potential, was observed. PBI-survived MDA-MB-231RP subline exhibited a hybrid EMT phenotype and a 70% reduction in tumor growth in the in vivo model of the chick embryo CAM. In contrast, PBI-survived MCF7RP cells exhibit mesenchymal-to-epithelial transition (MET)-like features, and their in vivo tumor growth increased by 66% compared to parental cells.
PBI triggers various cellular responses in different BC cell lines, influencing tumor growth through mechanisms like DNA damage repair, stress-induced premature senescence (SIPS), and alterations in the stiffness of tumor cell membranes. Our insights into entosis and the effect of extracellular matrix stiffness on metastatic propensity (nanoparticle uptake) enhance the understanding of the role of PBI in BC cells, emphasizing the need for more research to optimize its therapeutic application.
Keywords
- breast cancer
- proton therapy
- DNA damage repair
- extracellular matrix (ECM) stiffness
- stress-induced premature senescence (SIPS)
- entosis
- stiffness of tumor cell membranes
- mechanosensation
- chick embryo chorioallantoic membrane (CAM) model
Breast cancer is the most common cancer type among women worldwide and is the second leading cause of cancer-related deaths in this demographic globally [1]. Radiation therapy (RT) remains a cornerstone in the treatment of early [2] and metastatic [3] breast cancer (BC) as recommended by the European Society for Medical Oncology (ESMO) guideline committee. RT types are classified according to radiation quality measured by linear energy transfer (LET), which calculates energy deposited per unit length of a particle’s path. Low-LET-based radiation (including X-rays, gamma radiation, and electrons) deposits lower amounts of energy per unit length and thus induces sparse and mostly single-strand DNA breaks (SSBs). Its usage in BC treatment can lead to cardiotoxicity, which is a significant side effect. The introduction of hadron therapy (high-LET), such as accelerated proton beams, offers significant advantages over conventional low-LET-based RT [4]. High-LET protons deposit higher concentrations of energy in a shorter distance when traversing tissue and induce highly localized clusters of DNA double-strand breaks (DSBs) [5]. Due to unique physical properties, proton beams enhance target coverage and enable the precise delivery of radiation doses. This improves cancer cell-killing efficacy while safeguarding healthy tissues and minimizing damage to organs at risk, such as the heart, lungs, and skin. Nevertheless, proton therapy (PT) presents certain challenges, notably its higher costs and limited accessibility compared to established photon irradiation techniques. Systematic reviews of clinical trials evaluating PT for BC indicate that it may not significantly improve overall treatment outcomes compared to photon therapy. However, these studies consistently report a substantial reduction in treatment-related toxicities, with radiation dermatitis being the most notable improvement [6, 7]. Therefore, in clinical practice, it is crucial for radiation oncologists to meticulously evaluate patients before recommending PT to ensure that the dosimetric advantages translate into meaningful clinical outcomes [8].
Based on the expression of hormone receptors, i.e., the estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor-2 (HER-2) receptors in the tumor, BC is classified into five subtypes: luminal types A and B, luminal B/Her-2+, triple negative/basal, and Her-2+ [9]. The triple-negative breast cancer (TNBC) subtype, characterized by negative ER, PR, and HER2 receptor status, poses a significant clinical challenge for oncologists due to its aggressive nature and the lack of effective therapies. Several authors have reported controversies with respect to the use of conventional RT for patients diagnosed with TNBC [10]. Since PT is an effective treatment for cancers located in hard-to-reach areas and those resistant to conventional radiotherapy, it is crucial to investigate whether PT can be a viable therapeutic option for TNBC.
In clinical practice, one of the most critical challenges is selecting individuals undergoing treatment with high- or low-LET radiation, based on the tumor’s radiobiological properties which determine its radiosensitivity [11]. Analyzing differential gene expression profiles in primary and BC cell lines suggests that key factors influencing radiosensitivity include the activation of DNA repair pathways, the presence of cancer stem-like cells [5, 12, 13], hypoxia, and the complexity of the tumor microenvironment. To address this challenge effectively, it is essential to identify the precise molecular and phenotypic characteristics of tumor cells in response to RT. This deeper understanding will serve as the foundation for designing highly effective clinical treatment strategies. In the era of targeted therapies, recognizing biomarkers that define cancer cell responses is crucial for making informed therapeutic decisions. Transcriptional alterations specific to radiation types have been sporadically examined [14, 15, 16, 17]. However, to our knowledge, there has been no systematic effort to characterize the molecular (at the protein level) and biological effects of high-LET radiation and radiation doses in BC cells with varying susceptibility to conventional low-LET RT. Establishing this knowledge base would be instrumental in differentiating the adverse side effects of high-LET RT from its beneficial cytotoxic and immunogenic properties.
In general, high LET radiation has a higher relative biological effectiveness
(RBE). Recent study suggests a linear correlation between the doses of X-rays and
protons required to achieve a specific fraction of surviving cells [18]. However,
this relationship needs to be tested in specific BC cell lines. RBE varies
depending on cell line-specific biological factors, such as their ability to
repair DNA damage [19]. High-LET radiation typically induces a greater number of
DSBs per dose unit and generates more complex, clustered lesions compared to
low-LET radiation types [5]. This is due to its ability to deposit large amounts
of energy over a short distance. Different forms of DNA damage activate distinct
components of the DNA damage response (DDR) system, which plays a crucial role in
repair. Several key DDR proteins are involved in this process and are commonly
used to assess DNA damage. One of the most critical proteins modified near DNA
damage sites is phosphorylated histone H2AX (
Cancer cell migration is essential for invasion and metastasis, as it involves changes in cell shape and rigidity to navigate through tight tissue spaces [29]. During this process, cells often acquire increased invasiveness and resistance to anoikis, a form of programmed cell death [30]. Research on resistance mechanisms has not clearly explained how migration in confined spaces relates to the development of therapeutic resistance.
Research shows that irradiation can change tumor aggression by either reducing or increasing the invasiveness of surviving cancer cells, which depends on the cell line and type of radiation [31]. However, the effects of proton irradiation on the characteristics of tumor cells that survive and migrate through porous tumor tissue have not been fully studied. Understanding these changes is essential for developing effective treatments that target the migratory and invasive capabilities of cancer cells. Proton irradiation may inhibit cellular migration, as studies show it reduces cell motility in melanoma cells and alters the expression of migration and adhesion proteins [32, 33]. However, the impact of proton irradiation on cancer cell migration and resistance, especially in TNBC, needs more research to uncover its therapeutic potential.
In this study, we aimed to assess the differential biological effects of high-
and low-LET radiation doses on the clonogenic survival rates of MCF7 and
MDA-MB-231 cell lines, widely used in vitro models representing
ER-positive and TNBC/Claudin-low breast tumors, respectively. These models
exhibit distinct metastatic phenotypes and varying degrees of susceptibility to
conventional low-LET RT [34]. As a key component of the BC cell survival
response, the molecular reaction to DNA DSBs was evaluated through quantitative
high-content immunofluorescence analysis of radiation-induced foci (RIF) of
BC cell lines, MDA-MB-231 and MCF7 were sourced from ATCC (Manassas, VA, USA). MDA-MB-231 cells are characterized by their triple-negative status (ER-/PR-/HER2-) and TP53 deficiency, while MCF7 cells are estrogen receptor-positive (ER+) and progesterone receptor-positive (PR+), with intact TP53 function. Both cell lines were cultured in DMEM medium (PanEko, Moscow, Russia) containing 4.5 g/L glucose and 1% L-glutamine, supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, 100 U/mL streptomycin, and sodium pyruvate (PanEko, Moscow, Russia). The cells were maintained at 37 °C in a humidified atmosphere with 5% CO2. Cell line authenticity was verified through STR profiling, and they tested negative for mycoplasma contamination using the MycoReport Mycoplasma Detection Kit (Cat. # MR001, Evrogene, Russia).
Cells were irradiated at the Russian Proton Accelerator “Prometheus” of A.
Tsyb Medical Radiological Research Centre (MRRC, Obninsk, Russia). Irradiation
was carried out at the middle spread-out Bragg peak (SOBP, 15 mm width), with
doses of 2, 4, and 6 Gy, with 70
Cells were seeded in T25 tissue culture flasks 24 h prior to irradiation at 3–5
PE and survival fraction (SF) were subsequently calculated using the formula:
RBE is defined as the ratio of the absorbed dose of the reference radiation (gamma-irradiation, Со60, Dref) to the absorbed dose of the test radiation (proton Beam, D) required to achieve the same biological effect (Eqn. 3).
The biological endpoint is assessed through in vitro clonogenic survival, which is typically modeled using the linear quadratic model (LQM).
where the surviving fraction S as a function of the absorbed dose D,
Thus, the RBE was evaluated as a function of the surviving fraction S (RBES) using Eqn. 5:
where
Prior to irradiation, cells were seeded at a density of 0.1
Following incubation for 1, 12, 24, and 48 hours, cells were fixed in 4%
paraformaldehyde in PBS (pH 7.4) for 20 minutes at room temperature. After two
PBS rinses, cells were permeabilized with 0.3% Triton-X100 (PanEko, Moscow,
Russia) in PBS supplemented with 2% BSA to prevent nonspecific antibody binding.
Cells were then incubated with primary antibodies against
The proportion of cells expressing senescence-associated
Cells were seeded in 3D fibrin gel using the protocol described previously [36]. Fibrin gels were fabricated using human fibrinogen and thrombin (NPO RENAM, Moscow, Russia). The fibrinogen powder was dissolved in sterile 0.9% saline solution at 37 °C to get 20 mg/mL concentration, aliquoted and stored at –80 °C. Thrombin stock (0.1 U/µL) was prepared by dissolving 1000 U thrombin powder in 10 mL T7 buffer (50 mM Tris, 150 mM NaCl, pH 7.4) and stored at –80 °C in 150 µL aliquots. Gel of 90 and 1050 stiffness was prepared according to Table 1.
| Gel stiffness (Pa) | Fibrinogen (µL) | T7 buffer (µL) | Cells mixture/culture medium (µL) | Thrombin (µL) | Total (µL) |
| 90 | 12.5 | 112.5 | 125 | 5 | 255 |
| 1050 | 100 | 25 | 125 | 5 | 255 |
For live 3D colony growth monitoring, 5000 MCF7 cells and 8000 MDA-MB-231 cells
were seeded into 3D fibrin gels per well in a 24-well plate. Two wells were made
for each condition and at least 20 images of colonies were captured in each well
every two days on an EVOS M5000 fluorescent microscope (Thermo Fisher Scientific,
Waltham, MA, USA) in a transmitted light channel with a 20
Cells were seeded in 384-well plates at a concentration of 2.5
To prepare the cells for analysis of filamentous actin (F-actin), they were
fixed using a solution containing 4% paraformaldehyde diluted in 1
Non-irradiated and irradiation-survived MCF7 and MDA-MB-231 cells were seeded at
a density of 5
where WA0 h is the initial wound area, measured immediately after creating the
wound, and WA
The test was performed according to our previously published protocol [37]. Briefly, carboxylate-modified fluorescent 200 nm in diameter particles (excitation/emission 505/515 nm) (Invitrogen life technologies, Carlsbad, CA, USA) were used to evaluate the adhesion and encapsulation efficiency of the cells. Cells were seeded in 96-well plates (Corning Incorporated, Corning, NY, USA) at a concentration of 5000 cells per well. Nanoparticles (NPs) were added to the cells to achieve the final concentration of approximately 2000 particles per cell. Nuclei were stained with Hoechst 33342. Cells were incubated for 1 hour at 37 °C, then washed with PBS and treated with trypsin to remove non-internalized particles. Cell viability was confirmed using a live/dead nuclei fluorescent stain. Negative controls, consisting of cells without nanoparticles, were also evaluated under the same conditions. Data were obtained from 3 independent experiments, each sample contained at least 3 replicates.
For the analysis of cell migration through physically confined environments cells were serum-starved overnight and then seeded in the upper compartment of Transwell inserts (Cat.# 3428 , EDM Milli-pore, Billerica, MA, USA) containing 0.1 mL of serum-free medium specific to the cell type. The inserts had 8 µm pores and were placed in wells with serum-supplemented growth medium in the lower chamber, which served as a chemoattractant. Cells were incubated for 72 hours at 37 °C and 5% CO2. Following incubation, cells from both compartments were collected by trypsinization and counted using a hemocytometer after neutralization with a serum-containing medium. For each cell type, the experiment was repeated 6 to 9 times, with three replicates per experiment, to determine the percentage of cancer cells that migrated through the membrane.
Scanning ion-conductance microscopy (SICM) was employed to evaluate the mechanical stiffness of cell membranes. This advanced technique utilizes a nanoscale glass capillary filled with an electrolyte solution, which performs hopping movements to effectively map the topographical and mechanical properties of biological samples adhered to a substrate also immersed in the same solution.
The measurement of ion current occurs through the capillary tip, which is positioned between two AgCl electrodes that are submerged in the electrolyte solution, both inside and outside the capillary probe. This configuration allows for precise control over the interaction between the probe and the sample surface. For topographical mapping, a setpoint for an ion current decrease of 0.4% was utilized, while 1% and 2% were applied to assess stiffness. The Young’s modulus was subsequently calculated using the methodology established by Rheinlaender and Schäffer [38].
The SICM measurements were conducted using an instrument from ICAPPIC Ltd. (Moscow, Russia), which was equipped with a 1.2 mm borosilicate glass capillary that had been shaped using a P-2000 laser puller (Sutter Instrument, Novato, CA, USA). The inner diameter of the capillary tip was approximately 50 nm, as determined by measuring the ion current in a buffer solution.
For these SICM assessments, cells were seeded at a density of 100,000 cells per 35 mm Petri dish and incubated overnight at 37 °C in a CO2 incubator. Prior to conducting measurements, the culture medium was replaced with an HBSS buffer solution that served as the electrolyte.
Fertilized specific pathogen-free (SPF) eggs were purchased from a local SPF
hatchery. The surface of the eggs was cleaned with paper towels and sterile
water, and then placed horizontally in an incubator set to 37 °C and
70% relative humidity, with the automatic rotation function activated. This day
was designated as embryonic day 0 (EMD 0). On embryonic day 3 (EMD 3), the eggs
were removed, and the air sacs and major blood vessels were identified and marked
using an egg candler. Approximately 3 mL of egg white was then withdrawn, and
upon observing the chorioallantoic membrane (CAM) drop, a 2 cm diameter circle
was drawn at the drop location. The eggshell was cut along the marked edges using
a mini chainsaw, and the window was sealed with 3M semipermeable film before the
eggs were returned to the incubator, with the automatic rotation function turned
off. On EMD 9, tumor cells were seeded. The eggs were removed, the semipermeable
membrane was peeled off, and an O-ring (polytetrafluoroethylene (PTFE) O-ring,
with an inner diameter of 6 mm and an outer diameter of 9 mm) was placed over the
window near the densely vascularized area of the CAM. Twenty-five µL
of a serum-free cell suspension containing 5
The CAM xenograft experiments in this study were conducted in strict compliance with the guidelines and regulations of the Institutional Animal Care and Use Committee (IACUC). Chicken embryos are not classified as living animals until 17 days of incubation [39, 40], and our experiments with CAM were completed by embryonic day 16 (E16), which were exempt from IACUC regulation. Additionally, in adherence to the Russian Animal Experiment and Welfare Guidelines, ethical approval was not required. Daily assessments were carried out to ensure embryonic viability.
Statistical analysis was performed using Student’s t-test, the
Mann–Whitney U-test, and two-way ANOVA with Tukey’s post-hoc test for multiple
comparisons, all facilitated by GraphPad Prism 8 software. Results are presented
as means
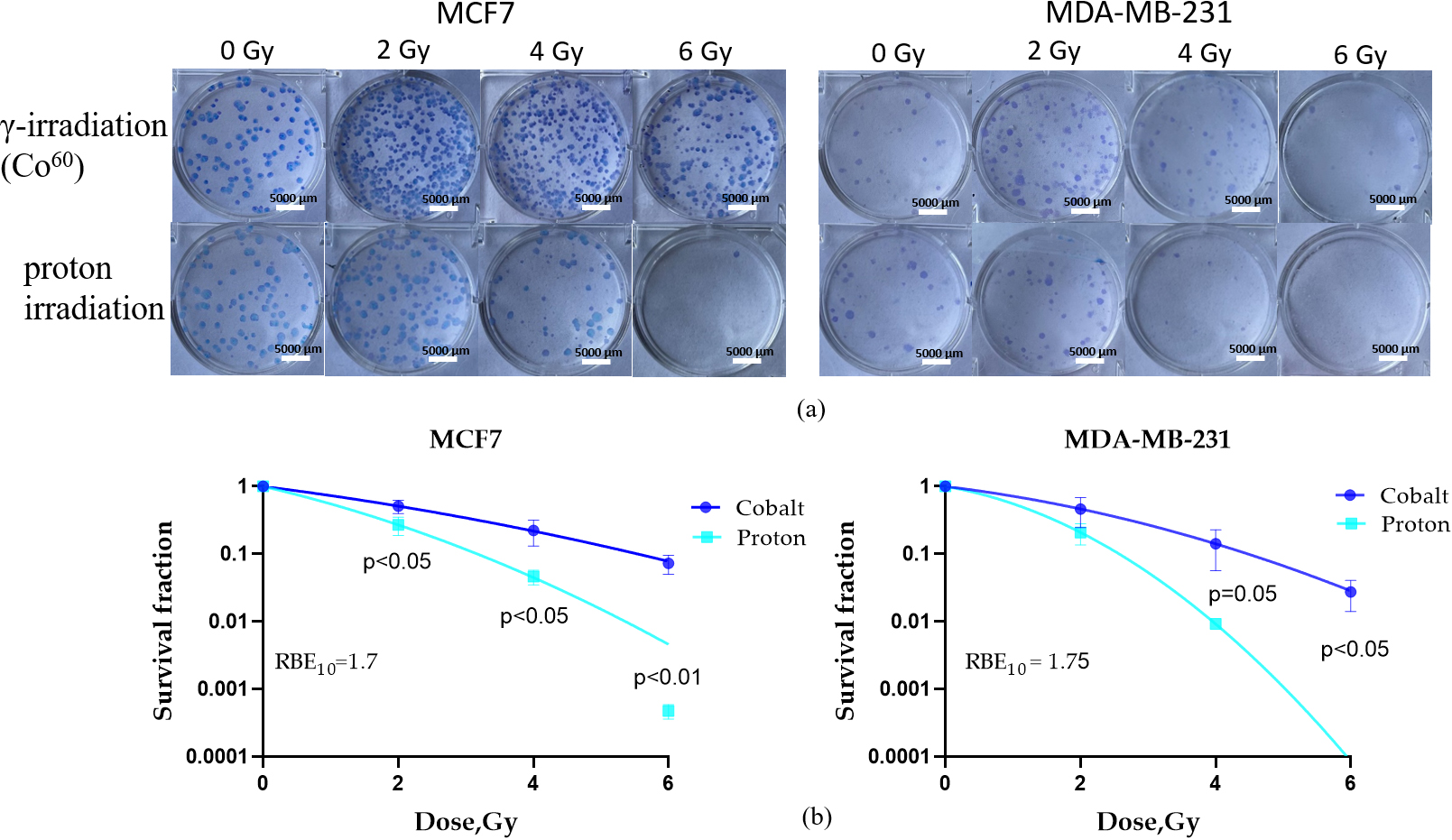
In this study, we first investigated the RBE of PBI for two BC cell lines, MCF7 and MDA-MB-231 (Fig. 1a), using clonogenic assay. Our goal was to examine the unique interplay between biological factors influencing physical parameters controlling the LET effect and innate radiosensitivity in determining a cell’s reaction to protons.
 Fig. 1.
Fig. 1.
Cell survival clonogenic assay was performed to evaluate the RBE
of proton beam irradiation. Representative pictures of colonies grown after each
irradiation dose of gamma and proton beam irradiation of MCF7 and MDA-MB-231 (a)
cells. Survival curves of MCF7 and MDA-MB-231 (b) cell lines after proton and
gamma (Co60 source) irradiation at doses of 2, 4, and 6 Gy were calculated.
Data are means
RBE is a fundamental concept in radiation biology that quantifies the
effectiveness of different types of ionizing radiation in inducing biological
effects, typically in comparison to standard reference radiation, such as X-rays
or gamma rays [41]. Using gamma irradiation from the Co60 source as a
reference, RBE was calculated based on the linear quadratic model (LQM), the most
widely used model for predicting cell survival in radiation therapy. Following
PBI, the radiosensitivity of MCF7 cells, as indicated by survival fraction,
significantly decreased compared to gamma irradiation (p
After a continuous cultivation period of 3 to 4 weeks in T-25 tissue culture flasks, we observed that a distinct subset of cells that survived high-LET radiation from each parental cell line successfully generated viable progeny. These progeny were designated as the MDA-MB-231RP and MCF7RP sublines for further investigation (see follow-up Sections 3.4–3.8). All subsequent experiments involving irradiation-survived progeny were conducted within 10 passages.
After radiation exposure, DNA damage response (DDR) proteins are rapidly recruited and/or modified at DSB sites leading to their accumulation at the damage site. This results in the formation of nuclear foci, known as radiation-induced foci (RIF), which are detectable under fluorescent microscopy. RIF quantification serves as an indirect method for assessing DNA damage and repair by monitoring the molecular response to DNA alterations, ultimately reflecting the resolution of RIF. Typically, RIF measurements are performed on fixed samples at defined time points following ionizing radiation (IR) exposure [42].
Our observed differences in the sensitivity of TNBC (MDA-MB-231) and ER-positive
(MCF7) cell lines to PT (Fig. 1) raised important questions regarding the DDR
protein signatures associated with cell sensitivity to high- versus low-LET
radiation. We further aimed to explore the molecular mechanisms underlying the
heightened biological impact of protons, as well as potential proton-specific
resistance mechanisms. Using quantitative immunofluorescence analysis, we
calculated the kinetics of RIF formation for key DDR proteins,
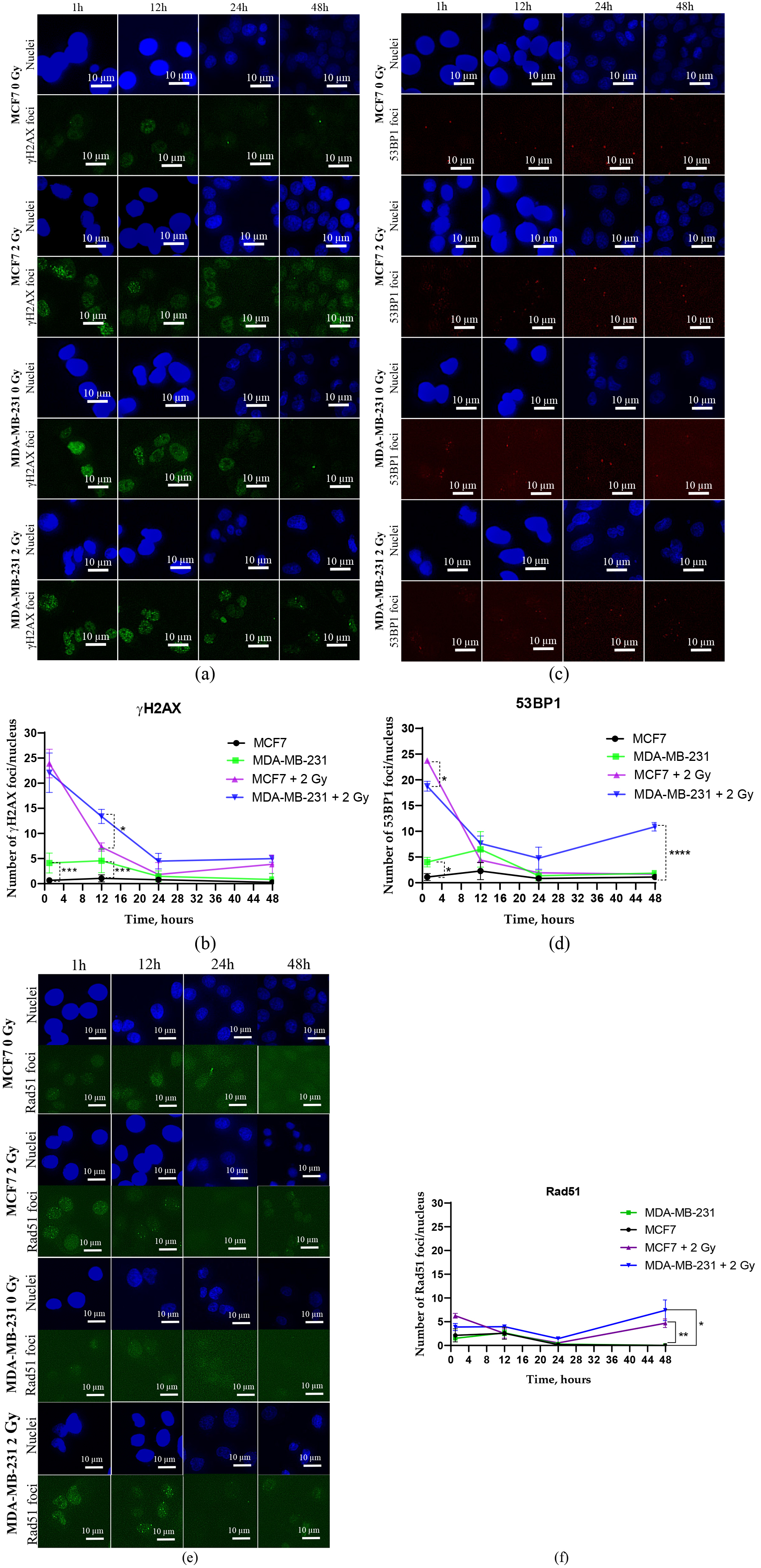 Fig. 2.
Fig. 2.
Immunofluorescence images and quantitative
immunofluorescence data illustrating the kinetics of DNA DSBs repair proteins.
Quantitative immunofluorescence analysis was performed to calculate the kinetics
of RIF numbers of key DDR proteins, namely
MCF7 cells exhibited a significantly higher number of 53BP1 RIF at the initial
time point (1 hour) post-irradiation compared to MDA-MB-231 cells (p
In irradiated MCF7 cells, the initial peak of Rad51 RIF, a key marker of the
homologous recombination (HR) pathway, reached approximately 5 foci per nucleus
(Fig. 2e) and gradually declined to baseline levels by 24 hours (Fig. 2f).
Increasing the incubation time to 48 hours resulted in more Rad51 RIF in both
irradiated cell lines. In irradiated MDA-MB-231 cells, the number of Rad51 RIF
remained above control levels at all examined time points (Fig. 2f). Notably, at
48 hours post-irradiation, Rad51 RIF levels were significantly higher compared to
control levels (p
The emergence of IR-induced cell senescence, a form of stress-induced premature senescence (SIPS) [43], represents a potential method to assist cancer cells in overcoming RT, and it may exacerbate the biological behavior of tumor cells after IR treatment [44]. Therefore, we sought to determine whether SIPS also contributes to the differences in radiosensitivity and DDR network functionality induced by high-LET radiation between MCF7 and MDA-MB-231 cells. Specifically, we aimed to investigate whether SIPS is associated with the inaccurate repair of DNA damage following high-LET irradiation.
To address this, we analyzed the proportion of senescent cells on days 1, 3, and
7 post-irradiation using analysis of the senescence-associated (SA)
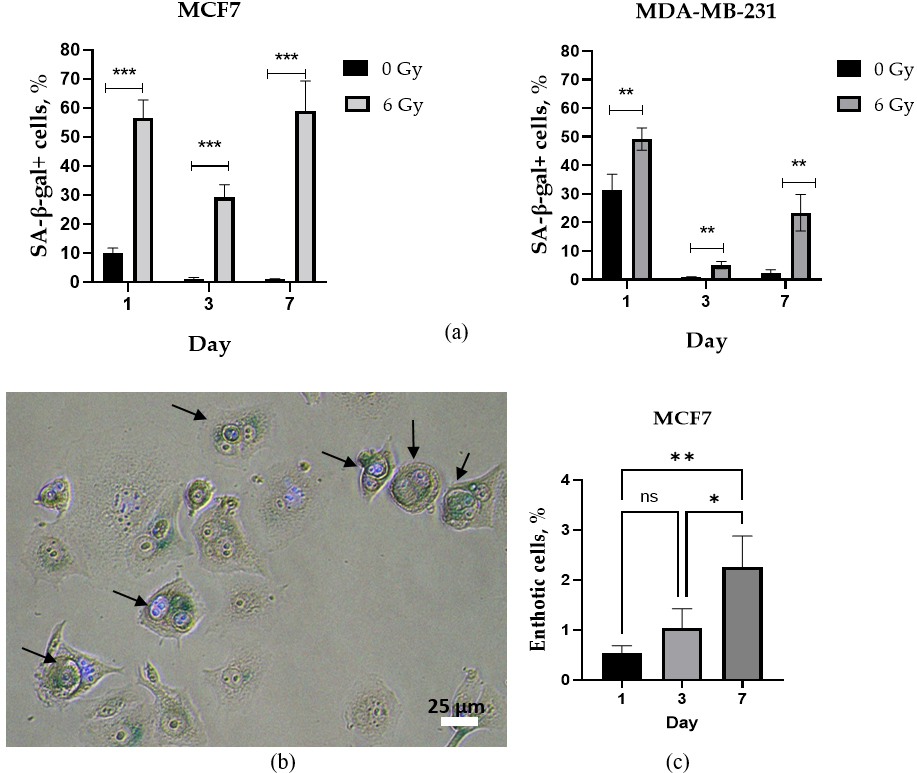 Fig. 3.
Fig. 3.
Quantitative analysis of senescence-associated
PBI caused a significant increase in the proportion of SA-
Entosis has recently been proposed as another potential survival mechanism in cancer cells exposed to radiation-induced stress [46]. BC has been found to have a high level of entosis competence, with entotic figures being observed in both primary tumors and metastatic lesions [47, 48, 49]. Despite this, little is currently known about the extent and effectiveness of the entosis process in human BC cell lines following proton irradiation.
Non-irradiated MCF7 cells exhibited a very low fraction (
Thus, the lower levels of the SA-
The remarkable biophysical properties of metastatic migrating cells, such as their exceptional motility and deformability, enable them to navigate through physical confinements created by neighboring cells or extracellular matrix (ECM) [29, 50]. The “scratch” test, also known as the wound healing assay, is a widely employed in vitro technique serving as a valuable tool allowing for the real-time visualization and quantification of 2D cell migration inside a monolayer cultured in serum-depleted medium (1% FBS) for 24 hours post-wound formation.
Non-irradiated parental MDA-MB-231 cells exhibited nearly double the migration within a monolayer as compared to non-irradiated MCF7 cells (Fig. 4a,b). Both MDA-MB-231RP and MCF7RP cell sublines demonstrated enhanced wound healing abilities compared to their non-irradiated parental cell lines (Fig. 4b).
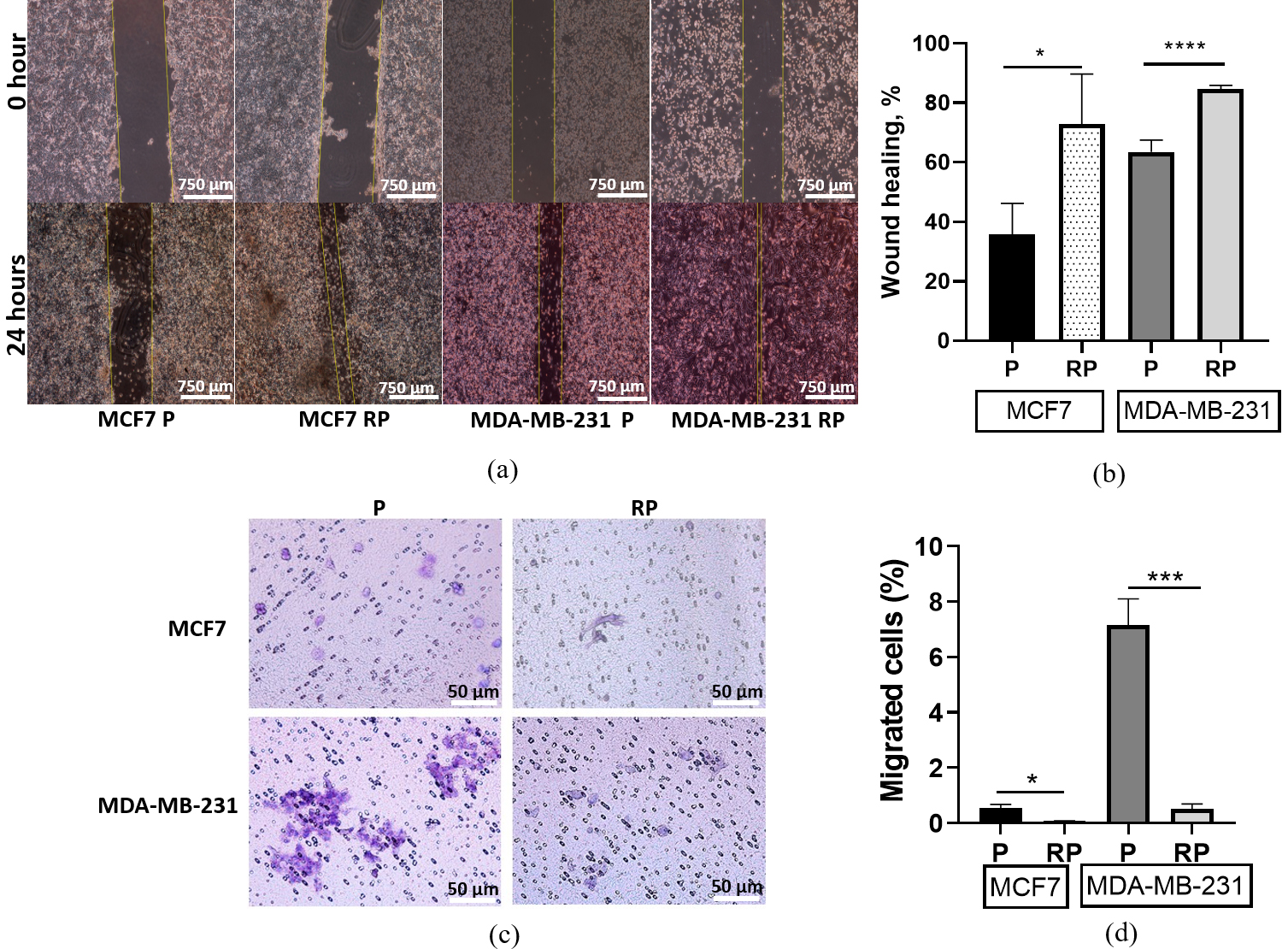 Fig. 4.
Fig. 4.
Collective (wound healing assay) and confined (through 8
µm pores of Boyden chamber membranes) migration of parental (P) and
proton irradiation-survived (RP) MCF7 and MDA-MB-231 cell lines. Representative
images at 24 hours after wound formation (a). Scale bar 750 µm.
Bar-graphs of changes in wound healing percentage (b). Representative pictures of
Giemsa-stained cells on the outer surface of the Boyden chamber membranes at 48
hours of confined migration (c). Scale bar 50 µm. Bar graphs of the
fraction of cells migrated into the lower Boyden chamber (d). Data are means of
three replicates
Using the classical transwell or Boyden Chamber assay, we evaluated a fraction of cancer cells that were confined in a physically restricted microenvironment. These cells were allowed to migrate through a membrane with 8 µm pores, which are significantly smaller than their size (19–21 µm), while following a gradient of serum concentration formed between the upper and lower chambers (Fig. 4c). In concordance with the 2D monolayer motility data, the non-irradiated MCF7 cells possess a significantly lower fraction of confined-migrating cells compared to MDA-MB-231 cells (Fig. 4d). In contrast to the 2D monolayer motility data, the MDA-MB-231RP subline, which survived high-LET exposure, exhibited a most prominent (14-fold) decrease in the number of confined-migrating cells compared to the MCF7RP (10-fold) subline (Fig. 4d).
EMT is a crucial biological process involved in various physiological and pathological conditions including cancer metastasis and invasion. EMT is characterized by the transition of epithelial cells into a mesenchymal phenotype, leading to changes in cellular morphology, gene expression, and functional properties. During classical EMT, epithelial markers such as E-cadherin are downregulated, while mesenchymal markers, including N-cadherin, Vimentin, and SNAIL are upregulated [51].
Unexpectedly, high-LET exposure led to an increase in E-cadherin expression in
MCF7RP cells, while no significant effect was observed in MDA-MB-231RP cells
compared to their respective non-irradiated parental controls (Fig. 5a,
Supplementary Fig. 1a). Additionally, we
observed a significant decrease in N-cadherin expression in MCF7RP cells
(p
 Fig. 5.
Fig. 5.
Analysis of EMT marker expression in parental (P) and proton
irradiation-survived (RP) MCF7 and MDA-MB-231 cells. The expression changes of
E-cadherin (a), N-cadherin (b), Vimentin (c), and SNAIL (d) were analyzed using
high-content immunofluorescent imaging and analysis. Statistical significance
calculated by Student’s t-test. * p
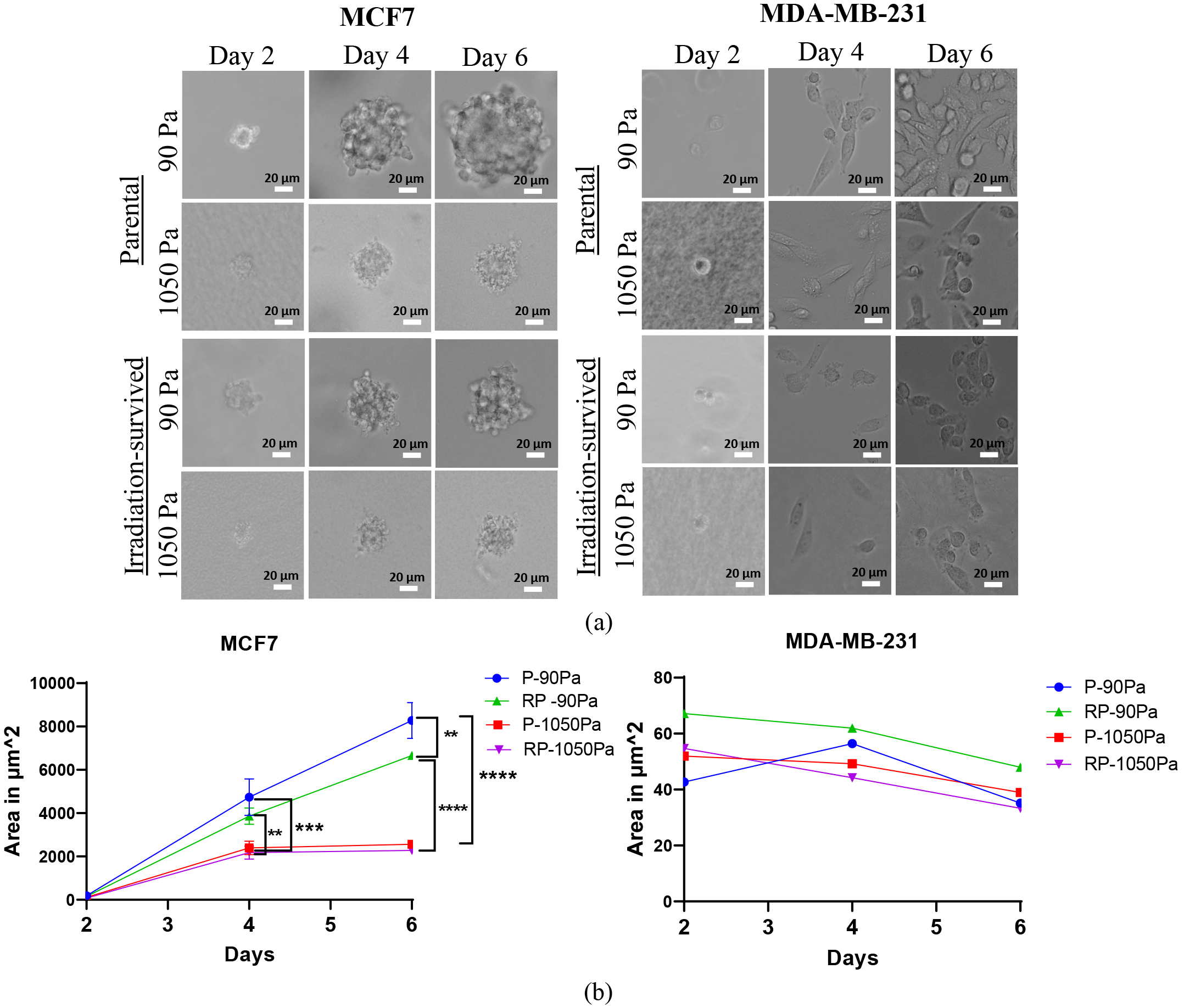
Soft 3D fibrin gels can select highly tumorigenic cells and support their growth without differentiation. Only soft tumor cells, not stiff ones, can form spheroid colonies [53]. In this study, untreated parental MCF7 and MDA-MB-231 cells, along with their isogenic proton irradiation-survived descendants were seeded in 3D fibrin gels with stiffness levels of 90 Pa and 1050 Pa. Previous research has demonstrated that stiffness of 90 Pa promotes cancer stemness [54], whereas 1050 Pa approximates the stiffness of healthy breast tissue [55]. Bright-field digital images of growing spheroids were captured under a microscope on days 2, 4, and 6 post-seeding (Fig. 6a). The area of each spheroid colony was subsequently measured using ImageJ software.
 Fig. 6.
Fig. 6.
Analysis of tumor spheroid growth in a 3D fibrin matrix. Images
(a) and quantification of the area (b) of multicellular tumor spheroids grown
within 90 and 1050 Pa 3D fibrin gels during the culture course of parental (P)
and proton irradiation-survived (RP) MCF7 and MDA-MB-231 cells on days 2, 4, and
6. Statistical significance calculated by two-way ANOVA with Tukey correction for
multiple comparisons. ** p
The mean area of compact MCF7 spheroids (of both irradiation-survived and
parental cells) increased with time becoming significantly (p
Meanwhile, MDA-MB-231 cells, both non-irradiated parental and irradiation-survived, formed only small diffuse spheroid colonies that remained almost unchanged in size over time, regardless of fibrin gel stiffness (Fig. 6b). By day 4 post-seeding, these cells had even degraded the fibrin gel and were predominantly found adherent to the bottom of the 6-well plate. Since cancer cells are known to secrete matrix metalloproteinases (MMPs) to degrade the basement membrane and facilitate invasion into the stromal matrix, we hypothesized that MDA-MB-231 cells mediate protease-driven degradation of the fibrin gel. However, the addition of the MMP inhibitor Marimastat on the first day of culturing had little to no effect on fibrin gel degradation in both parental and irradiation-survived MDA-MB-231 cells (data not shown).
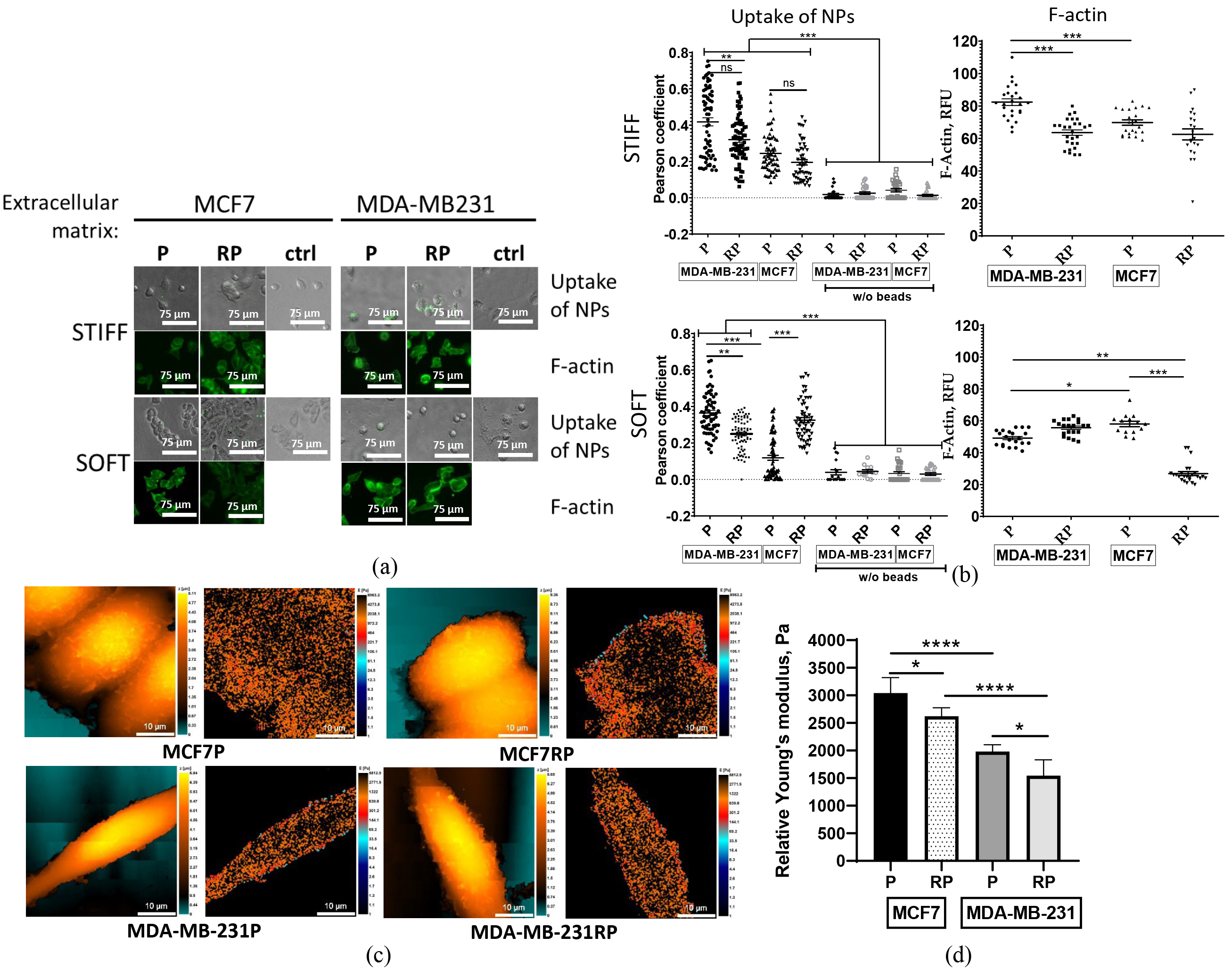
After observing the differing abilities of proton irradiation-survived MCF7RP and MDA-MB-231RP cells in 2D migration in a monolayer and confined migration (Fig. 4), we aimed to further evaluate their metastatic potential in comparison to their parental counterparts. One emerging method for assessing metastatic potential is the NP test, which examines cellular nanoparticles (NP) uptake. This approach explores the relationship between cell migration and cancer cell endocytosis, emphasizing the role of the actin cytoskeleton [37].
To investigate the well-documented influence of ECM on cytoskeletal changes,
cells were either pre-cultivated on rigid plastic surfaces (
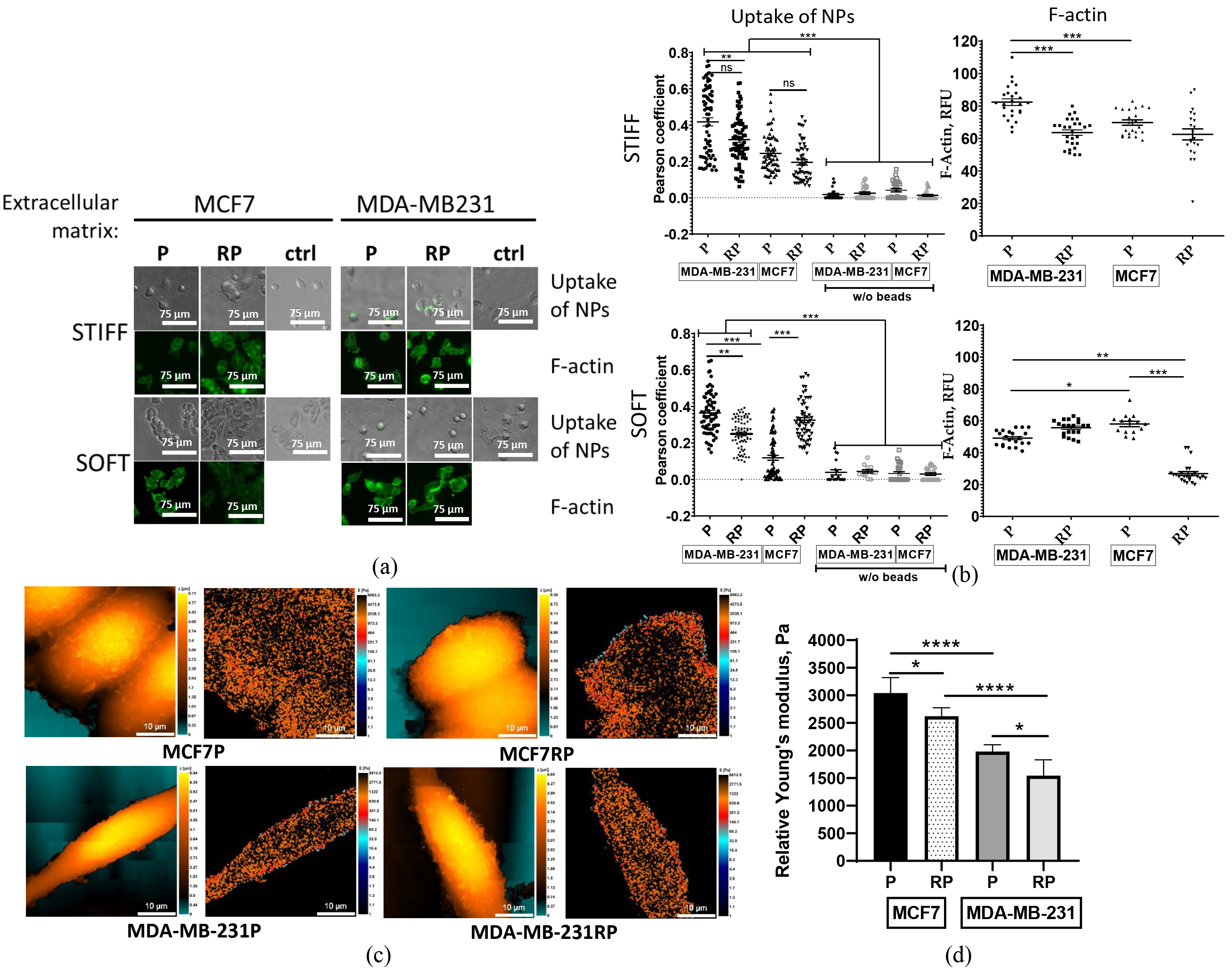 Fig. 7.
Fig. 7.
Analysis of nanoparticles association and F-actin
expression by parental (P) and proton irradiation-survived (RP) MCF7, and
MDA-MB-231 cells seeded on 2D stiff plastic or a soft fibrin (90 Pa) layer
following their pre-cultivation on either 2D stiff (plastic) or 3D soft fibrin
gel, respectively. Representative pictures of cellular nanoparticles (NP) association test and
F-actin phalloidin staining (a). The quantitative comparison of cellular NP
uptake efficiency was calculated based on quantitative co-localization (Pearson’s
and overlap coefficients, scatter slope) of fluorescent nanoparticles, and
integrated fluorescence intensities (RFUs) of F-actin filaments (b). Scale bar 75
µm. Topography (left) and stiffness (right) mapping images obtained
using scanning ion-conductance microscopy (SICM) on breast cancer cell lines (c). Scale bar 10 µm. Average
relative Young’s modulus calculated as a weighted mean for each cell and then
averaged across all measured cells (d). Statistical significance calculated by
Student’s t-test. * p
Regardless of microenvironment stiffness during pre-cultivation and NP
association, the highly metastatic parental MDA-MB-231 cells exhibited
significantly higher NP association efficiency compared to non-metastatic
parental MCF7 cells (Fig. 7b). In contrast, the efficiency of NP association in
PBI-survived MCF7RP and MDA-MB-231RP cells displayed a clear dependence on
microenvironment stiffness. MDA-MB-231RP cells exhibited a marked reduction in
NPs association compared to their parental counterparts (p
Next, we investigated the hypothesis that the microenvironment stiffness experienced by BC cell lines during culture influences their own cytoskeletal changes. Given that filamentous actin (F-actin) plays a crucial role in determining cell stiffness, one of the important biophysical features of malignant cells [56], we undertook a comprehensive analysis utilizing phalloidin staining of F-actin.
Both parental (P) MDA-MB-231 and proton irradiation-survived (RP) MCF7 cells cultured on a stiff plastic (Fig. 7a,b upper panels) exhibited significantly greater levels of cellular F-actin expression than those grown on a soft fibrin layer (Fig. 7a,b lower panels). High-LET irradiation-survived MCF7RP cells demonstrated a notable increase in softness compared to their parental cells, as evidenced by a significant reduction in F-actin expression, but only within a softer microenvironment (Fig. 7b). Conversely, MDA-MB-231RP cells exhibited greater stiffness in comparison to their parental counterparts, although the difference was not statistically significant when maintained in the same microenvironment (Fig. 7b, lower panels).
SICM was used to confirm the mechanical properties of the cell membrane by obtaining both topographical and stiffness mapping images of parental and PBI-survived cell sublines (Fig. 7c). We did not observe significant morphological changes following PBI exposure. The characteristic Young’s modulus revealed that parental MCF7 is significantly stiffer than MDA-MB-231 cells, which corroborates previous studies [57, 58]. Furthermore, both the PBI-survived cell sublines MCF7RP and MDA-MB-231RP exhibited a statistically significant decrease in stiffness following irradiation (Fig. 7d). This finding aligns with our data on NP uptake and F-actin expression in cells cultured within a stiff microenvironment (Fig. 7b, upper panel), suggesting a correlation between reduced stiffness, F-actin expression and increased 2D collective migration of PBI-survived cells (Fig. 4b).
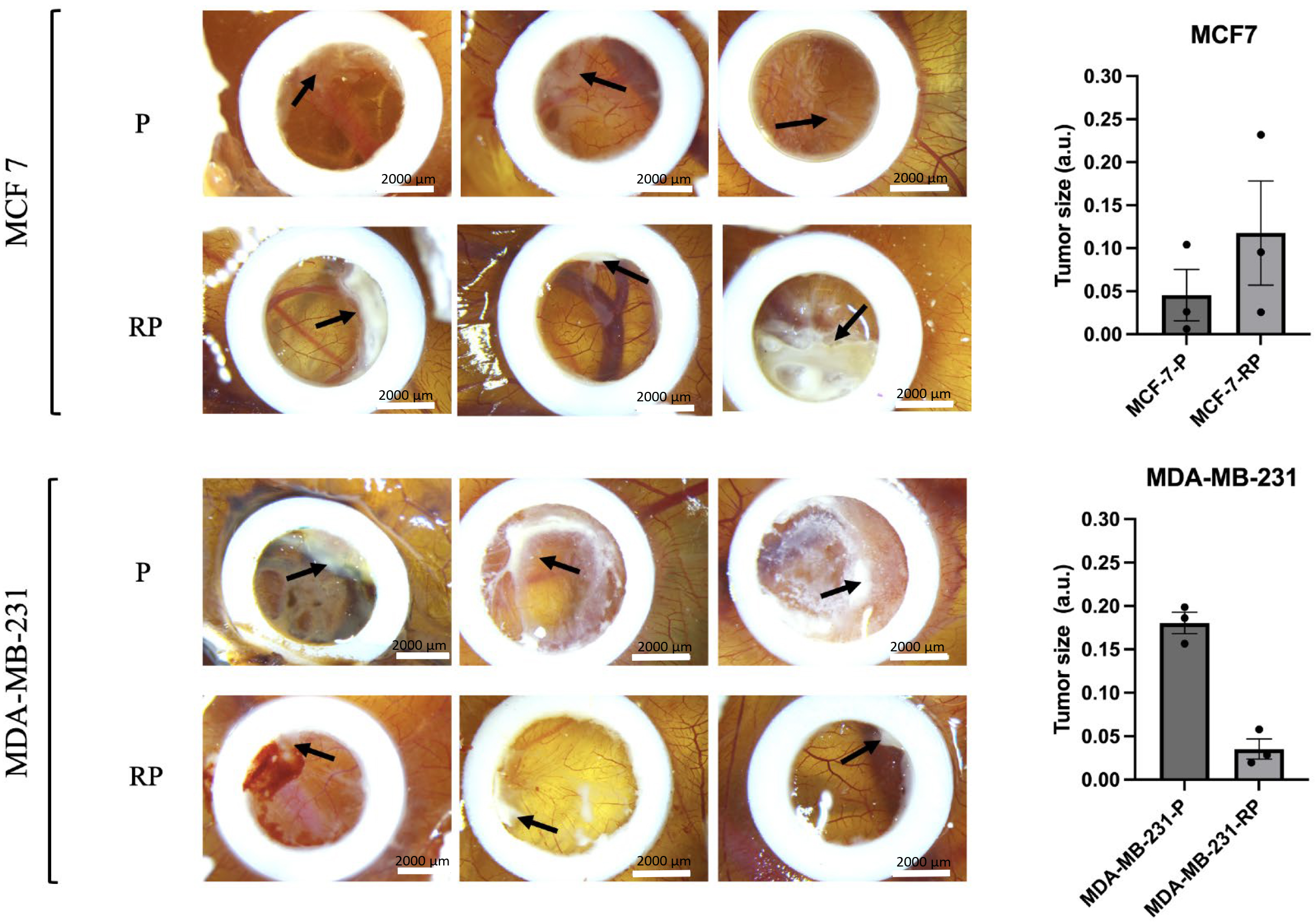
The chick embryo CAM model presents a compelling in vivo alternative for tumor engraftment. Its vascularized membrane fosters robust tumor growth while the chick embryo’s immune system remains underdeveloped until embryonic day 16 (E16), making it an ideal environment for such research [59, 60, 61]. In this in vivo model, we sought to investigate the comparative tumorigenic potential of parental (P) MDA-MB-231 and MCF7 cells alongside their proton irradiation-survived counterparts (RPs).
By E16, the parental (P) MDA-MB-231 cells formed a substantially larger tumor mass organization characterized by a well-organized tumor–stroma that exhibited a white-opaque appearance and marked revascularization through CAM vessels (see Fig. 8, lower panel, marked by the black arrow). The implanted parental MCF7 cells exhibited a well-organized structure within small clusters, particularly near the CAM vessels. There was little indication of tumor mass organization, accompanied by surrounding mesenchymal and inflammatory reactions (refer to Fig. 8, upper panel).
 Fig. 8.
Fig. 8.
The chick embryo chorioallantoic membrane (CAM) model was
employed to assess the tumorigenic potential of BC cells. Representative images
depict tumors that developed by embryonic day 16 in an in vivo CAM
tumorigenicity assay, comparing parental (P) MDA-MB-231 and MCF7 cells with their
proton irradiation-survived counterparts (RPs). Black arrows indicate tumor
formation sites. Scale bar 2000 µm. The right panel illustrates the
comparative tumor sizes between the P and RP groups. Statistical significance
calculated by the Mann–Witney U-test. Values represent data from individual
tumors with the mean
Fig. 8 clearly illustrates that the proton irradiation-survived (RP) variant of MDA-MB-231RP cells displayed only minimal tumor growth (indicated by black arrow), highlighting a ~6-fold decrease in their tumorigenic potential in vivo compared to parental MDA-MB-231 cells. On the other hand, MCF7RP cells exhibited a more pronounced (~3-fold higher size) organization of tumor mass compared to their parental cell line. These data suggest that high-LET proton irradiation substantially impacts the tumorigenic ability of these cells when compared to their syngeneic parental counterparts.
BC is the most common cancer type among women globally and the second most common overall, making it a major contributor to cancer-related mortality among women worldwide [62]. While PT shows a promising approach for the treatment of BC, there is still a substantial knowledge gap regarding the cellular and molecular responses of BC cells to proton irradiation. In this study, we investigated the RBE of PBI for two BC cell lines, MCF7 and MDA-MB-231 (Fig. 1a), using a clonogenic assay. The RBE of PBI in MCF7 and MDA-MB-231 (Fig. 1b) cell lines for the 10% cell survival fraction (i.e., SF = 0.1) was found to be 1.7 and 1.75, correspondently, demonstrating the superior efficacy of PBI in eliminating cancer cells compared to conventional photon irradiation. Notably, the MDA-MB-231 cells demonstrated the lowest clonogenic survival rates after high LET irradiation when compared to the MCF7 cell line. This finding is consistent with previous observations [34]. The large differences in the slopes of D10% for proton and D10% for X-ray curves in both BC cell lines show that both physical and biological factors are important in determining the sensitivity of BC cells to radiation based on LET.
High-LET radiation, such as PT, induces complex clustered DNA lesions including non-DSB damage surrounding DSB. These lesions are often inefficiently repaired or remain unrepaired, contributing to the greater mutagenic and cytotoxic effects observed in irradiated cells [63, 64]. Our observed differences in the sensitivity of TNBC (MDA-MB-231) and ER-positive (MCF7) cell lines to PT raise questions regarding the DDR protein signatures associated with cellular sensitivity to high- versus low-LET radiation. The expression changes of these DDR proteins following ionizing radiation exposure have been extensively studied by us and other researchers [65, 66, 67, 68, 69]. Earlier transcriptomic studies identified several key genes involved in BC cell radiosensitivity to high-LET radiation including Rad51 [70], along with four additional effectors, recently confirmed [71].
The present study demonstrates that PBI induces significantly different effects
on DNA DSBs in two BC cell lines, as evidenced by the immunofluorescent analysis
of
Tumor repopulation and metastasis following IR remain major challenges in the successful effective elimination of cancer. To address this, we aimed to investigate the characteristics of BC cells that survive PBI, focusing on their ability to grow and spread. MCF7 and MDA-MB-231 cell lines were exposed to a single lethal dose (6 Gy) of PBI and allowed to recover for 3–4 weeks. All subsequent experiments (refer to Sections 2.6–2.12) were conducted to assess the survival characteristics of their progeny, now designated as the MDA-MB-231RP and MCF7RP sublines, in comparison to their parental isogenic counterparts.
Even when DNA damage remains unaddressed, tumor cells can evade programmed cell
death and survive proton-based treatment by entering various forms of dormancy
including senescence. Despite being permanently arrested in proliferation and
lacking the ability to divide, senescent cells exhibit resistance to apoptosis,
sustained metabolic activity, an SIPS phenotype, and the ability to alter their
surrounding microenvironment [72]. Studies, including our own, have demonstrated
that the IR dose plays a crucial role in determining whether irradiated cancer
cells undergo senescence or apoptosis [67, 73, 74, 75, 76]. This phenomenon is closely
associated with the extent of DNA damage and the functionality of the DDR
network. Entosis has recently been proposed as another potential survival
mechanism in cancer cells exposed to radiation-induced stress [46]. Through this
process, tumor cells compete with the winning cells engulfing and eliminating
weaker neighboring cells. Additionally, there is evidence suggesting that stress
conditions can trigger a fatal form of entosis, where internalized cell undergoes
lysosomal digestion. This has been observed in approximately 30% of MCF7 cells
grown in suspension, while under the same conditions, no entotic events were
detected in MDA-MB-231 cells [77]. However, under fully adherent conditions,
certain cancer cells may undergo a vital form of entosis instead [46]. BC is
known to exhibit a high level of entosis competence, with entotic figures
observed in both primary tumors and metastatic lesions [47, 48, 49]. Here, for the
first time, we report that the entotic program is exclusively activated in proton
irradiation-survived MCF7 cells (Fig. 3c) under fully adherent conditions. In
contrast, this phenomenon remains absent in MDA-MB-231 cells grown under
identical conditions. Remarkably, a majority of entotic MCF7 cells were positive
for SA-
Our observed significant differences in clonogenic survival, DDR, motility, and invasiveness in response to PBI raise the question of whether the key molecular effectors driving these processes are also differentially engaged in MCF7RP and MDA-MB-231RP sublines. Recent transcriptomic studies have indicated that following PBI, the TP53 signaling pathway was identified as the most statistically significant pathway activated in MCF7 cells [71, 81]. This activation emphasizes its crucial role in balancing cell survival and death after irradiation. In contrast, the EMT pathway was among the top five activated pathways in MDA-MB-231 cells following PBI [12, 71]. Our findings collectively indicate that high-LET radiation induces a divergent, either non-classical (hybrid) EMT or mesenchymal-to-epithelial transition (MET) phenotypes in irradiation-survived sublines. Based on classical marker expressions, MCF7RP (TP53-competent) cells exhibited more likely a MET phenotype, characterized by lowering N-cadherin and Vimentin levels and a significant increase in E-cadherin expression. Conversely, MDA-MB-231RP (TP53-deficient) cells intensified their mesenchymal phenotype, showing significant upregulation of N-cadherin and a marginal increase in SNAIL, despite unchanged levels of Vimentin and E-cadherin expressions. This hybrid EMT phenotype induced by high-LET irradiation in MDA-MB-231RP sublines significantly enhanced 2D collective migration in monolayer (Fig. 4b), but not the 3D confined migration (Fig. 4d) compared to their parental counterparts. This raises the question of whether there is a direct correlation between the EMT-MET phenotype, migration, and invasion of cancer cells. For instance, previous study have demonstrated that EGFR inhibition in prostate cancer epithelial cells reduced their migration but not invasion [82]. Additionally, defining EMT solely based on a limited set of classical markers, as was carried out in the current study, may not provide a comprehensive characterization of the process. Initially studied in the context of embryonic development, EMT research has since evolved into a major focus in cancer biology. However, the variability and adaptability of EMT programs have not been adequately addressed [39]. Our study highlights that during the EMT–MET process, cancer cells often co-express epithelial and mesenchymal markers, frequently failing to undergo complete EMT transition. This suggests that partial (or hybrid) EMT states are more common than previously thought, reinforcing the idea that traditional EMT markers, such as E-cadherin, N-cadherin, and Vimentin, may not fully capture the complexity of EMT in cancer cells [39].
The unique biophysical properties of metastatic migrating cells, including their exceptional motility and deformation capabilities, enable them to navigate through the physical constraints imposed by adjacent cells and the ECM [29, 50]. Solid tumors are influenced by mechanical forces from the surrounding tissue, which induce tumor cells’ deformation, significantly impacting their migration, division, and survival [83]. A strong correlation exists between cancer cell deformability and malignancy, with a reduction in cancer cell stiffness, as measured by the Young’s modulus [84]. Stiffness defined as the ratio of stress to deformation describes a cell’s ability to resist shape changes. In various cancer types, undifferentiated tumorigenic cells are generally softer than their differentiated counterparts [85]. Notably, metastatic cancer cells isolated from the pleural fluids of BC patients exhibited over 70% lower stiffness compared to benign reactive mesothelial cells [86]. During entosis, highly deformable soft cells are more efficient in engulfing and outcompeting stiffer neighboring cells, a process driven by RhoA and actomyosin activity [87]. Our current findings indicate that high-LET radiation induces entosis in MCF7 cells (Fig. 3c), while it does not affect MDA-MB-231 cells. The difference suggests inherent variations in cellular stiffness, which was confirmed by our SICM measurements of the two cell lines. Interestingly, the tendency for entosis did not show any correlation with cell stiffness as assessed by SICM (Fig. 7c,d). Of note, the descendants of PBI-survived both MCF7 and MDA-MB-231 cells further decreased their cellular stiffness.
ECM stiffness plays a crucial role in various cellular processes, including growth, proliferation, migration, immunity, malignant transformation, metabolic reprogramming, and apoptosis [88]. In the present study, we investigated the effects of ECM stiffness on MCF7 and MDA-MB-231 cells. Our findings reveal a compelling inverse relationship between the volume of parental MCF7 spheroids and the stiffness of the fibrin gel used for encapsulation (Fig. 7a,b). This observation aligns with a recent study that reported similar results using MCF7 cells cultured on a linear stiffness gradient of GelAGE [89]. For the first time, we demonstrate that proton irradiation affects the spheroid-forming (tumorigenic) abilities of MCF7 cells, as revealed by spheroid culturing within a 3D soft (90 Pa) fibrin gel microenvironment (Fig. 6). Liu et al. [53] previously showed that softer fibrin gels (90 Pa) promote tumorigenicity and stemness features in melanoma cells, leading to primary and secondary tumor formation in animal models. In contrast, a microenvironment that mimics the mechanical properties of healthy breast tissue (1050 Pa) [90] likely plays a protective role in maintaining cell properties, making it difficult to distinguish MCF7RP from parental cells based on spheroid area. Our data also highlight the limitations of using soft fibrin gels to differentiate highly deformable and potentially more metastatic cancer cells, particularly when these cells (e.g., both parental and proton irradiation-survived MDA-MB-231 cells) secrete gel-degrading factors that compromise ECM stiffness (Fig. 6). In our current study, we did not aim to investigate the nature of fibrin gel-degrading activity. The role of matrix metalloproteases (MMPs) was not supported, as using Marimastat, a broad-spectrum inhibitor of MMPs, did not affect our study results. Indeed, plasmin, a serine protease, can break down cross-linked fibrin multimers within a clot in the peripheral circulation. Aprotinin, a proteolytic enzyme inhibitor, has been previously demonstrated to slow fibrinolysis [91]. Aprotinin is a natural serine protease inhibitor that blocks various proteases, such as plasmin, urokinase, protein C, elastase, cathepsin, and kallikrein. Aminocaproic acid, another plasmin-specific inhibitor, could be further utilized in experiments involving MDA-MB-231 cells grown in fibrin. Consequently, the influence of these plasmin inhibitors warrants further investigation. Together, these findings prompted us to explore alternative methods for comparing the mechano-biological characteristics that underlie differences between high-LET radiation-survived BC cell lines.
F-actin is a critical component of the cytoskeleton playing a key role in determining cell the mechanical properties of cells, particularly stiffness [56]. Disrupting actin stress fibers eliminates variations in cell stiffness, chromatin stretching, and gene upregulation in response to mechanical forces [92]. Previous studies have shown that beads coated with a synthetic peptide containing the Arg-Gly-Asp (RGD) sequence bind to cells via integrin–actin linkages. When tethered by tense F-actin bundles (stress fibers), these bundles primarily resist bead rotation or movement under stress, serving as an index of cell stiffness. Disrupting of F-actin bundles reduces tethering, leading to decreased resistance to bead rotation and significantly lower cell stiffness [93, 94]. To examine how ECM stiffness affects cytoskeletal (F-actin) changes and their link to metastatic potential, we conducted a nano-bead association assay to assess the ability of MCF7RP, MDA-MB-231RP, and their parental cell lines to encapsulate carboxylate-modified fluorescent NPs (see Fig. 7a,b). This encapsulation was used as an indicator of their pro-metastatic phenotype [37]. Regardless of the microenvironment stiffness, the highly metastatic parental MDA-MB-231 cells exhibited significantly higher association efficiency than non-metastatic parental MCF7 cells (Fig. 7b), consistent with previous findings [37]. Next, we found that both parental cell lines demonstrated mechanosensation—the ability to perceive ECM stiffness and translate that information into F-actin expression (Fig. 7b, right upper vs. lower panels). Our results show that high-LET irradiation alters the mechanosensation of proton-irradiated MCF7RP and MDA-MB-231RP cells, affecting their perception of microenvironment stiffness. This, in turn, leads to substantial modifications in their mechano-biological characteristics, ultimately impacting their metastatic potential. This finding aligns with recent observations in non-metastatic MCF7 cells and confirms earlier predictions [95]. The F-actin cytoskeleton is a highly organized composite structure composed of cortical actin and F-actin stress fibers, both of which play a crucial role in cell mechanics, including division and migration [96]. We present novel evidence that MCF7RP and MDA-MB-231RP cells that survived high-LET proton irradiation exhibit an inverse correlation between F-actin expression—an indicator of cell stiffness—and NP uptake, a marker of metastatic potential. Our findings underscore the significant impact of relationships. In soft microenvironments, high-LET irradiation survivors from the highly aggressive and metastatic MDA-MB-231 cell line exhibit a significant reduction in metastatic potential, as manifested by lowering NP uptake in vitro. Conversely, high-LET irradiation survivors from the non-metastatic MCF7 cell line appear to enhance this trait under the same microenvironment. These unexpected in vitro results prevented us from drawing definitive conclusions regarding the tumorigenic and metastatic potential of the proton irradiation-survived sublines.
This prompted us to directly compare the tumorigenicity of MDA-MB-231RP and MCF7RP cell sublines using an in vivo model. The chick embryo CAM, in ovo model, is a promising alternative in vivo system, well-suited for tumor engraftment due to its extensive vascularization, which supports tumor growth, and the lack of a fully developed immune system in the chick embryo until E16 [59, 60, 61]. This model has been widely used to study cancer biology in various contexts, including tumorigenicity, angiogenesis, metastasis, and anticancer drug testing, and serves as a robust alternative to simple organoids and costly patient-derived xenograft (PDX) mouse models [97]. In ovo experiments offer a low-cost, time-efficient approach, allowing tumor growth assessment within a relatively short 16-day period, with being for further analysis [59].
In this model, the parental highly aggressive and metastatic MDA-MB-231 cells
formed substantial cell clusters characterized by a well-organized tumor–stroma
that exhibited a white-opaque appearance and marked revascularization through CAM
vessels (see Fig. 8, the black arrow in the upper P panel). In stark contrast,
when we grafted parental MCF7, we observed minimal evidence of organized tumor
mass formation, alongside notable angiogenic and vasculogenic effects in the
surrounding tissue (refer to Fig. 8, the black arrow in the upper panel). The
data show strong consistency with previously published findings from other
research groups [98, 99]. Consistent with our in vitro NP uptake
findings under soft microenvironments, MDA-MB-231RP cells displayed only minimal
tumor growth (Fig. 8, the black arrow in the lower RP panel), indicating a
substantial decrease in their tumorigenic potential in vivo. In
contrast, MCF7RP cells generated larger tumors compared to their parental cell
line (Fig. 8, the black arrow in the upper RP panel), a result that also aligns
well with our in vitro findings regarding nanoparticle uptake in softer
microenvironments. It is important to consider that the effects observed in MCF7
cells may be specific to either cell type or the species, as parental MCF7 cells
exhibited significant alterations in their molecular profile after grafting onto
the CAM, leading to the acquisition of a more aggressive phenotype. Notably, they
transitioned to a triple-negative status and initiated EMT, as indicated by the
expression of vimentin and the concurrent loss of
The effects of ionizing radiation on the cytoskeleton-mediated mechanobiological properties of tumor cells are well-established and thoroughly researched [102]. The investigation into how PBI affects these properties has just started, leading to new insights and discoveries. While mechanosensation of the tumor microenvironment (TME) is usually performed through the activation of surface mechanosensors such as integrins [103], transient receptor potential (TRP) ion channels [104], and the yes-associated protein (YAP) and the transcriptional coactivator with PDZ-binding motif (TAZ) molecular complex [105], actin cytoskeleton plays an essential role as a mechanotransducer. In this role, it converts physical stress into biochemical signals, which affect cell actions like division, adhesion, and migration. X-ray irradiation induced a depolymerization of the actin filaments and, consequently, a decrease in their elastic modulus that ultimately led to the augmentation in the migratory and invasive abilities of tongue squamous cell carcinoma (TSCC) cells [106]. Alterations in the cytoskeletal network were observed, indicated by a reduced signal intensity of actin fibers. These data correlate well with our results. Furthermore, our study clearly indicates that PBI also affects mechanosensation, so that highly metastatic tumor cells surviving proton irradiation (MDA-MB-231RP) are more likely to metastasize (indicated by NP uptake) in a rigid TME compared to when they are in a softer TME. While this study did not focus on the direct impact of PBI on mechanosensor functions, it clearly requires further investigation.
PT for BC is characterized by higher costs and limited accessibility compared to traditional photon therapy. Clinical trials have not consistently shown that PT is better than photon therapy, so radiation oncologists must evaluate patients carefully. This evaluation is crucial to ensure that the dosimetric advantages of proton therapy translate into meaningful clinical benefits.
There are no systematic reviews or clinical trials comparing proton therapy’s effects on different molecular subtypes of BC. Studies have explored how breast cancer cell lines react to proton irradiation, particularly regarding DNA repair [107] and molecular pathways [108]. These studies examine how various cell lines respond to PT, but they do not address clinical outcomes for different breast cancer subtypes.
Our research indicates that PT may effectively treat TNBC by studying cancer recurrence in live models. In particular, our results indicate that highly metastatic tumor cells surviving proton irradiation (MDA-MB-231RP) are more likely to metastasize (indicated by NP uptake) in a rigid microenvironment compared to when they are in a softer environment. From a clinical standpoint, combining treatments that soften tumor tissue with proton therapy may reduce the risk of metastases in TNBC patients following radiation therapy. However, this requires further confirmation in vivo models. There is still a strong need for thorough reviews and clinical trials to compare the outcomes of PT in various BC subtypes. Such studies would help determine whether PT provides unique benefits for specific subtypes and inform personalized treatment strategies.
PBI appears to be a key factor in modulating the mechanosensation of two distinct BC cell lines. In softer ECM environments, survivors of non-metastatic and less aggressive MCF7 cells tend to acquire pro-metastatic characteristics. Conversely, post-PBI descendants of highly metastatic and more aggressive MDA-MB-231 cells exhibit a marked reduction in these traits under similar conditions. Our findings emphasize the limitations of using 2D environments such as rigid plastic culture dishes in distinguishing highly deformable cancer cells with increased metastatic potential and tumor repopulation capacity. This becomes particularly evident with parental MDA-MB-231 and MCF7RP cells acquiring “softness” when cultured in soft conditions such as a 90 Pa fibrin layer, a trait that correlates with the larger tumor sizes observed in vivo. A limitation of this study is the use of only two BC cell lines, representing just two of the four recognized molecular subtypes. However, the observed association between their distinct molecular phenotypes and differential responses to PBI suggests a potential avenue for developing more personalized treatment strategies tailored to patients with specific molecular profiles and tumor stiffness. Further studies are needed to examine the response of additional BC molecular subtypes to PBI, either alone or in combination with chemotherapy. Potential treatment enhancements could include recently suggested STAT3-targeting inhibitors [109] or natural compounds such as ellagic acid [110], which may improve antitumor efficacy while minimizing adverse effects. Overall our research demonstrates that high-LET proton irradiation significantly alters the tumorigenic potential of BC cell lines in vivo, particularly when compared to their syngeneic parental counterparts. Results from our clonogenic (in vitro) and CAM (in vivo) assays not only validate the superior effectiveness of proton beams but also endorse their targeted application in certain cancer therapies.
All data reported in this paper will be shared by the lead contact upon request.
MP—original draft preparation, project management, methodology and conceptualization. RM, YW, WX, PM, VN, EK, ZN, DK—formal analysis, research, visualization. VS—irradiation. SL, and MP—funding acquisition. MP—writing. AO, SL—writing-review and editing and conceptualization. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All animal experiments in this study were conducted in strict compliance with the guidelines and regulations of the Institutional Animal Care and Use Committee (IACUC). Chicken embryos are not classified as living animals until 17 days of incubation, and our experiments with CAM were completed by embryonic day 16 (E16), which were exempt from IACUC regulation. Additionally, in adherence to the Russian Animal Experiment and Welfare Guidelines, ethical approval was not required.
The authors wish to express their sincere appreciation to the “Proton radiation complex” group at the A. Tsyb Medical Radiological Research Centre (MRRC, Obninsk, Russia) for providing the irradiation experiments essential to this study.
This research was funded by the Ministry of Science and Higher Education of the Russian Federation (State Task) 075-03-2024-117/113, Project No. FSMG-2024-0045.
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBL36415.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.