



 , Hengyan Zhang 2,†, Heguo Yan 3,4, Pingxing Han 1, Jing Zhang 1,*
, Hengyan Zhang 2,†, Heguo Yan 3,4, Pingxing Han 1, Jing Zhang 1,* , Yangwen Liu 4
, Yangwen Liu 41 Department of Reproductive Medicine, Zhaotong Hospital of Traditional Chinese Medicine, 657000 Zhaotong, Yunnan, China
2 Department of Dermatology, Zhaotong Hospital of Traditional Chinese Medicine, 657000 Zhaotong, Yunnan, China
3 Department of Clinical Medicine, Yunnan University of Chinese Medicine, 650500 Kunming, Yunnan, China
4 Department of Endocrinology, Zhaotong Hospital of Traditional Chinese Medicine, 657000 Zhaotong, Yunnan, China
†These authors contributed equally.
Abstract
Male infertility represents a major health concern, accounting for approximately 50% of all infertility cases in couples. This condition arises from multiple etiologies, with oxidative stress gaining increasing attention in recent studies. During the final stages of sperm maturation, the majority of the cytoplasm is discarded, leaving sperm with a diminished antioxidant defense system, which makes them highly susceptible to the detrimental effects of reactive oxygen species (ROS). ROS can be generated from both intrinsic and extrinsic sources. Intrinsically, ROS are primarily produced by mitochondrial activity, while extrinsic factors include alcohol consumption, smoking, circadian rhythm disruption, gut microbiota imbalance, and leukocyte infiltration. Excessive ROS production leads to DNA damage, apoptosis, and epigenetic modifications in sperm, ultimately impairing sperm motility and contributing to infertility. This review provides a comprehensive examination of ROS sources and examines the mechanisms by which ROS induce sperm damage. Furthermore, it explores the therapeutic potential of antioxidants in mitigating oxidative stress and improving sperm quality.
Keywords
- male infertility
- oxidative stress
- reactive oxygen species
- sperm function
- antioxidants
Male infertility is defined as the inability to achieve pregnancy in a female partner after one year or more of regular unprotected intercourse, attributed to male factors [1]. Approximately 50% of infertility cases in couples are due to male infertility [2]. The causes of male infertility are varied, including: (1) spermatogenic dysfunction, such as oligoasthenoteratozoospermia, (2) genetic disorders, such as Y chromosome microdeletions and androgen insensitivity syndrome, (3) varicocele, (4) ejaculatory disorders, and (5) environmental factors, such as exposure to high temperatures and heavy metals [3, 4, 5]. Semen analysis is essential for the evaluation of male infertility, with research showing significantly elevated levels of reactive oxygen species (ROS) in the sperm from infertile men, suggesting that oxidative stress may play a role in the pathogenesis of male infertility [6]. ROS have been implicated in the peroxidative damage to sperm plasma membranes in infertile men [7]. Such damage impairs sperm motility and membrane fusion capabilities, both of which are critical for fertilization [8].
In 1943, MacLeod [9] first demonstrated that human sperm could generate ROS, specifically hydrogen peroxide. Subsequent research identified the superoxide anion as the primary ROS produced by human sperm, which is rapidly converted to hydrogen peroxide by the enzyme superoxide dismutase (SOD) within the cell [10]. In addition to hydrogen peroxide and superoxide anions, other ROS, such as hydroxyl radicals and nitric oxide radicals, also play significant roles in sperm function [11]. Low levels of ROS are essential for normal sperm activities, including fertilization capability (acrosome reaction, hyperactivation, capacitation, and chemotaxis) and sperm motility [12]. However, as shown in Fig. 1, excessive ROS levels can induce to increased sperm apoptosis, thereby elevating the risk of male infertility [13]. Sperm are particularly sensitive to oxidative stress, as they lose most of their cytoplasm during the later stages of differentiation, resulting in a significant reduction in [14]. This deficiency in the antioxidant system not only heightens oxidative stress but also leaves sperm highly vulnerable to ROS-induced damage, including mitochondrial and nuclear DNA damage, DNA fragmentation, telomere shortening, and Y chromosome microdeletions [15], which further exacerbate the risk of male infertility [16]. Additionally, sperm cell membranes are rich in polyunsaturated fatty acids (PUFAs), making them highly susceptible to ROS-induced damage [17]. External factors, such as aging, air pollution, smoking, high-temperature environments, and alcohol consumption, further contribute to oxidative stress, thereby increasing the risk of male infertility [18, 19, 20, 21].
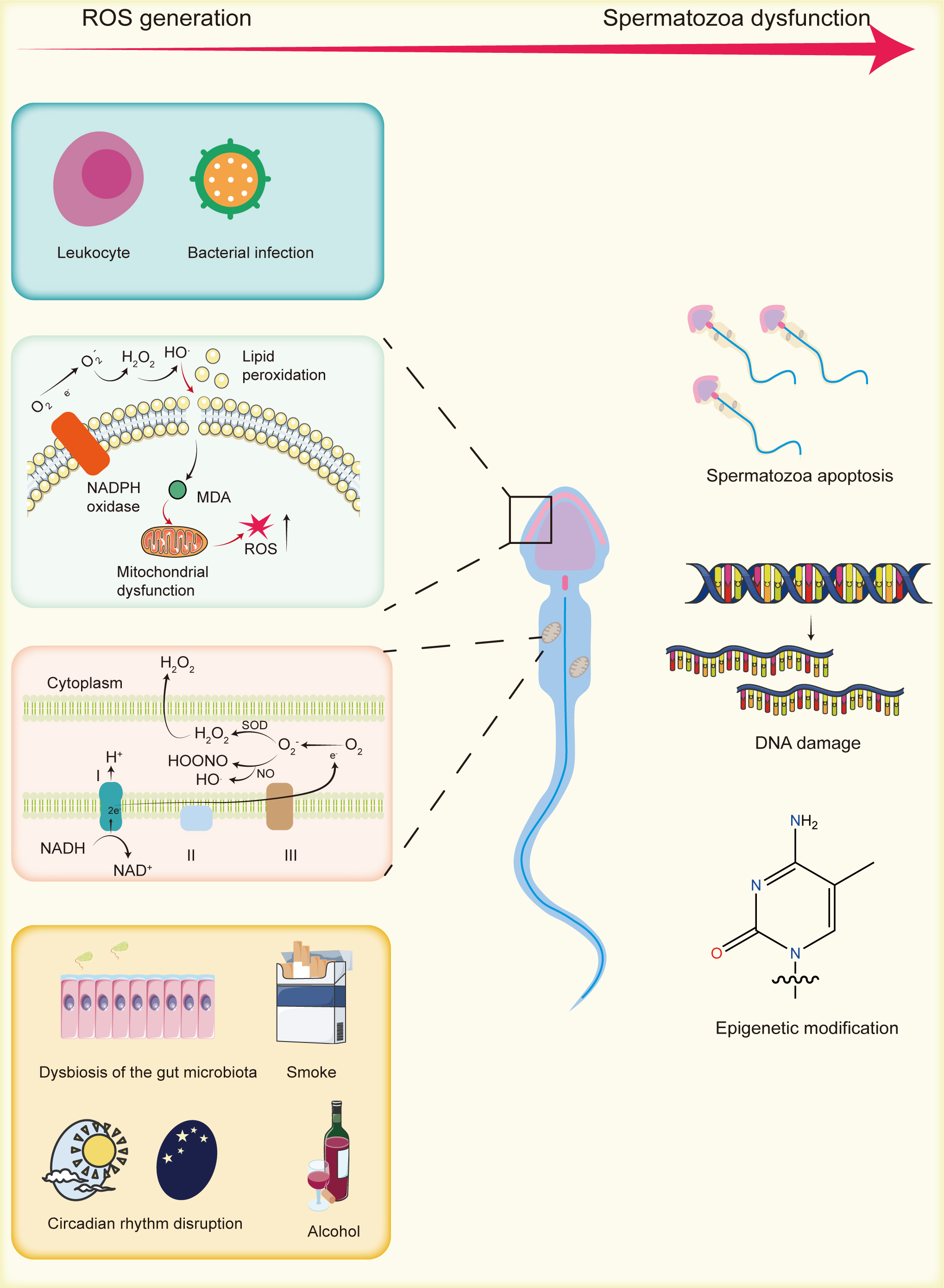 Fig. 1.
Fig. 1.
The generation of reactive oxygen species (ROS) and their impact on sperm. Endogenous ROS primarily originate from mitochondria, lipid peroxidation, and white blood cells. Exogenous sources include gut microbiota alterations, smoking, circadian rhythm disruptions, and alcohol consumption. Excessive ROS can cause sperm DNA damage, epigenetic modifications, and apoptosis, thereby contributing to male infertility. MDA, malondialdehyde; NADPH, nicotinamide adenine dinucleotide phosphate; NADH, nicotinamide adenine dinucleotide; SOD, superoxide dismutase. Created by using Adobe Illustrator (version 28.7.1, Adobe Systems Incorporated, San Jose, CA, USA).
This review provides a comprehensive analysis of the sources of ROS in sperm, addressing both endogenous and exogenous origins. Endogenous ROS include those produced by mitochondria and through lipid peroxidation, while exogenous sources stem from factors such as gut microbiota, inflammation, and lifestyle habits (e.g., smoking, circadian rhythm disruption, and alcohol consumption). The review also explores how ROS contribute to reduced sperm motility by inducing DNA damage, inducing apoptosis, and alterations in histone modifications and DNA methylation. Furthermore, it examines the potential role of antioxidants in enhancing sperm motility, offering valuable insights into oxidative stress, male infertility, and sperm vitality, while advocating for the application of antioxidants in male infertility treatment.
Oxidative stress arises from an imbalance between pro-oxidants and antioxidants, favoring oxidizing agents. In the reproductive system, oxidative stress occurs when free radical production surpasses the body’s antioxidant defense capacity. Research has demonstrated that the levels of superoxide anions and peroxynitrite in the semen of infertile men are significantly elevated compared to fertile men, with a positive correlation between oxidative stress levels and sperm DNA fragmentation [22]. Sperm DNA fragmentation is linked to various adverse clinical outcomes, including reduced fertility and higher miscarriage rates [23]. Therefore, ROS levels are closely associated with fertilization rates and sperm physiological functions [24]. The sources of oxidative stress in sperm can be categorized into endogenous and exogenous origins. Endogenous sources involve the oxidative stress caused by an excess of ROS generated from intracellular metabolic activities and immune responses. The sources primarily include the mitochondrial electron transport chain, lipid peroxidation, and leukocytes.
Mitochondria, in addition their role in adenosine triphosphate (ATP) production through oxidative phosphorylation, participate in various intracellular physiological processes, including ROS generation [25]. Research has indicated that human sperm motility depends more on glycolysis than on mitochondrial ATP production [26]. However, damage to the mitochondrial structure or the mitochondrial genome significantly impacts sperm motility. Further study has shown that sperm mitochondria is closely linked to ROS production [27]. During cellular respiration, the mitochondrial electron transport chain (ETC), located on the inner mitochondrial membrane, plays a key role in energy production. As electrons traverse the ETC, they can leak and interact with oxygen molecules to form superoxide anions, primarily occurring at Complex I and Complex III. ROS generated by Complex I are the main contributors to oxidative damage in human sperm [28]. Additionally, flavoenzymes in the mitochondrial electron transport chain, such as electron-transferring flavoprotein, have also been implicated in ROS production [29]. Changes in environmental conditions can also induce mitochondrial ROS production. Sperm experience significant oxidative stress during freezing and thawing, leading to a decline in sperm quality [30]. Studies suggest that cryopreservation results in osmotic changes, with mitochondria being more susceptible to damage from osmotic shifts than the sperm plasma membrane, further triggering ROS production [31, 32]. Due to their charged nature, superoxide anions have limited ability to cross mitochondrial membranes. Within mitochondria, SOD converts superoxide anions into hydrogen peroxide [33]. Hydrogen peroxide can oxidize the thiol groups at the active site of glyceraldehyde-3-phosphate dehydrogenase (GAPDS), a key enzyme in the glycolytic pathway, explaining the observed decline in ATP production in sperm cells as ROS levels rise [34]. Moreover, superoxide anions can react with nitrogen oxides to form peroxynitrite, which can dissociate into nitrogen dioxide and hydroxyl radicals [35].
Nicotinamide adenine dinucleotide phosphate (NADPH) oxidase, a membrane-bound enzyme, is present on the sperm membrane. Under the influence of NADPH oxidase, NADPH acts as an electron donor, transferring its electrons to oxygen, which leads to the generation of superoxide anions and other ROS, including hydrogen peroxide and hydroxyl radicals [36]. These ROS further oxidize the lipids in the sperm plasma membrane, initiating a cascade of lipid peroxidation reactions [37]. The sperm plasma membrane is rich in PUFAs, which contain multiple carbon-carbon double bonds, offering numerous sites for radical oxidation. Additionally, the bis-allylic methylene positions in PUFAs have low C-H bond dissociation energies, making them particularly susceptible to oxidation. ROS produced by NADPH oxidase oxidize PUFAs, generating lipid radicals, which then react with oxygen to form peroxyl or alkoxyl radicals. These radicals stabilize by abstracting hydrogen atoms from adjacent carbon atoms, perpetuating the chain reaction of lipid peroxidation. This process results in the formation of various reactive aldehydes, such as 4-hydroxynonenal and malondialdehyde [38]. These aldehydes can bind to mitochondrial proteins, altering their conformation, leading to electron leakage and further ROS production [37].
In addition to ROS generated by sperm, semen ROS levels are also influenced by
the presence of leukocytes. An increase in leukocyte concentration in semen
raises nitrite levels, thereby diminishing the semen’s antioxidant capacity [39].
WHO guidelines define leukocytospermia as a leukocyte
In addition to the intrinsic ROS production, various external environmental factors, such as shifts in the microbiota, microbial infections, tobacco use, disturbances in circadian rhythms, and alcohol intake, can contribute to elevated ROS levels. These external factors generate free radicals, collectively termed extrinsic ROS. Together, they contribute to extrinsic oxidative stress, where these harmful environmental influences enhance free radical production, exceeding the capacity of the body’s antioxidant defense systems and resulting in oxidative damage. Research indicates demonstrated that male infertility arises from a combination of environmental influences and genetic factors. Therefore, understanding the relationship between environmental influences and ROS is essential for advancing therapeutic strategies for male infertility.
Recent research has increasingly focused on the microbiota’s role in human health. Evidence suggests that men with male infertility exhibit significant imbalances in the composition and functionality of the microbiota in both the gastrointestinal and reproductive tracts [44]. The gut, acting as a crucial conduit between the body and the external environment at both the oral and anal openings, serves as an “endocrine organ” influencing various physiological functions. Notably, research by Tremellen et al. [45] has demonstrated that compromised gut mucosal barrier integrity can lead to the translocation of gut microbiota into systemic circulation, which may reduce testosterone levels in men of reproductive age. Modulating the gut microbiota with oligosaccharides has been shown to improve lipid metabolism, which in turn ameliorates sperm quality and the testicular microenvironment. Concurrently, an enhancement in the antioxidant capacity within the testes and blood has been observed [46]. These observations highlight the gut microbiota’s ability to modulate the redox balance in both the blood and the testicular microenvironment, a process known as the gut-testis axis [47].
Gut microbes and their metabolites, such as lipopolysaccharide (LPS), can enter
the systemic circulation as microbial-associated molecular patterns (MAMPs) and
subsequently reach the testes via the testicular arteries. In the
testes, they are recognized by Toll-like receptors (TLRs) on sperm, triggering
the production of pro-inflammatory cytokines, including TNF-
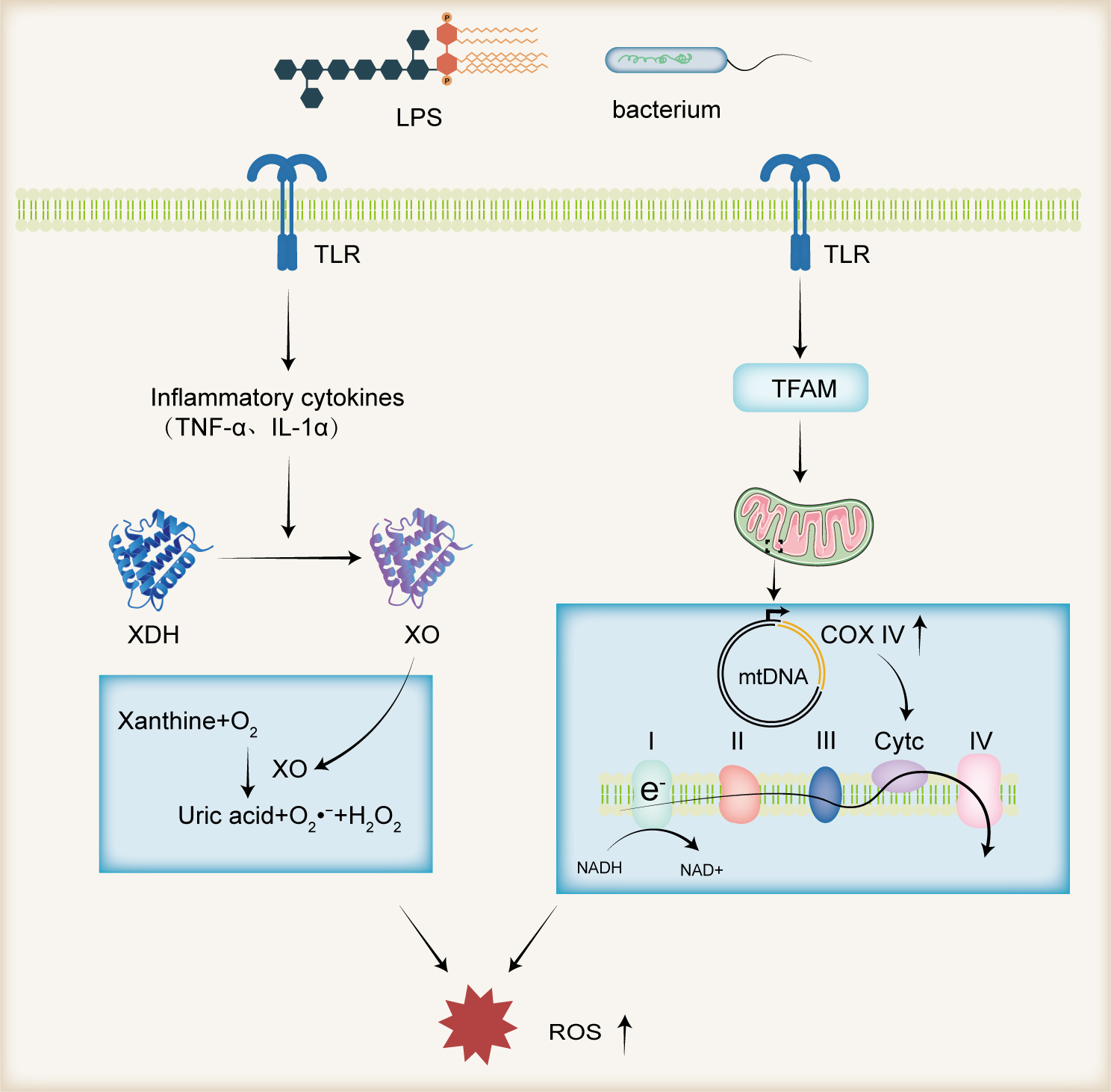 Fig. 2.
Fig. 2.
Gut Microbiota and ROS Production. Gut microorganisms and their
metabolite LPS function as MAMPs, which are recognized by TLRs on the surface of
sperm. This recognition induces the production of pro-inflammatory cytokines such
as TNF-
In addition to the influence of the gut microbiota, approximately 15% of male infertility cases are associated with microbial infections within the urogenital system [54]. Clinically, the presence of more than 1000 CFU/mL of semen in aerobic culture conditions is considered indicative of bacterial prostatitis. Gram-negative urethral pathogens have been identified as agents that diminish sperm motility and viability, compromise the integrity of the sperm plasma membrane, and escalate intracellular ROS levels [55, 56, 57]. Alongside the elevation of ROS via LPS, an increase in leukocyte counts has been observed in the semen of patients with bacterial prostatitis [58], further contributing to ROS production in the semen.
Although the precise effect of smoking on male fertility remains uncertain, research indicates that smoking impairs sperm motility and antioxidant capacity [59]. A meta-analysis of 20 studies involving 5865 participants revealed that smoking adversely affects semen parameters, notably reducing sperm count and motility, with the correlation intensifying as smoking intensity increases [60]. Additionally, even smokeless cigarettes exhibit a dose-dependent decline in sperm parameters [61]. Cigarettes contain a variety of toxic chemicals, including significant amounts of reactive free radicals and ROS, which damage lipids, proteins, and nucleic acids, contributing to infertility [62]. Moreover, secondhand smoke exposure has been shown to elevate ROS levels in tissues, resulting in DNA and methylation damage [63]. Increased oxidative stress from smoking may also correlate with reduced seminal plasma zinc levels. Zinc, a coenzyme for numerous enzymes, plays a critical role in antimicrobial activities and inhibition of cellular ROS production [64]. Kumosani et al. [65] found that smoking reduces zinc levels in seminal plasma, heightening oxidative stress and negatively affecting sperm density and motility. Furthermore, Arabi and Moshtaghi [66] demonstrated that, in addition to directly increasing ROS production, sperm from smokers exhibit increased sensitivity to peroxides, rendering them more vulnerable to oxidative damage.
Sleep deprivation has been shown to negatively affect male fertility. In an experiment involving rats subjected to sleep deprivation for either 4 days or 7 days, Choi et al. [67] found a significant decrease in sperm motility in the group deprived of sleep for 7 days, although this association was not seen in the other groups. However, all groups exhibited a significant decline in testosterone levels [67], which has been reported to lead to sexual dysfunction [68]. These observations underscore the close relationship between sleep and male fertility. Sleep plays a crucial role in ROS elimination, with ROS being produced during the day and cleared during sleep. Insufficient sleep significantly elevates ROS levels in the body, triggering oxidative stress [69]. In a Wistar rat model, peripubertal sleep restriction (21 days, 18 h/day) significantly increased lipid peroxidation, glutathione levels, and total radical-trapping antioxidant potential, decreased neutrophil migration and epithelial compartment size, and ultimately impaired epididymal development and sperm motility [70]. To explore the relationship between sleep patterns and male fertility, Lu et al. [71] conducted on a Mendelian randomization study, highlighting the potential influence of chronotype on testosterone secretion, a key hormone in male reproductive health. In contrast, no substantial causal associations were found between other sleep-related factors, such as sleep duration or insomnia, and male fertility potential [71]. This distinction suggests that circadian preferences play a more critical role in fertility than other sleep characteristics. It is worth noting that circadian rhythm disruptions have also been reported to be associated with decreased female fertility [72]. These findings indicate that circadian rhythm imbalance, rather than sleep duration, is likely the primary factor affecting male fertility e lipid peroxidation and oxidative stress.
Case report demonstrates that long-term severe alcohol intake (approximately 165 g of alcohol per day, over a period of 10 years) significantly impacts male fertility and ultimately leads to azoospermia [73]. Akang et al. [74] demonstrated that alcohol-treated mice exhibited substantially elevated levels of testicular malondialdehyde and DNA fragmentation, along with reduced SOD activity and glutathione content, indicating that alcohol induces infertility by increasing oxidative stress. Alcohol consumption can also cause mitochondrial DNA damage, disrupting mitochondrial function and exacerbating mitochondrial ROS production [75]. This effect may arise from alcohol’s ability to alter the structure of mitochondrial ribosomes, impairing mitochondrial protein synthesis [76] and affecting the translation of oxidative phosphorylation-related proteins. As a result, mitochondrial DNA undergoes oxidative modifications, antioxidant defenses are depleted, and glutathione levels are reduced, ultimately leading to cell apoptosis [77]. Additionally, chronic alcohol consumption induces the expression of cytochrome P450 enzymes, particularly CYP2E1, which increases NADPH oxidase activity in cells, thereby enhancing ROS production [78].
Over the past few decades, obesity resulting from high-fat and high-calorie
diets has been shown to significantly impair male fertility [79]. Research
indicates that the increase in adipose tissue promotes insulin resistance, which,
in turn, reduces sperm uptake and metabolism of glucose [80]. Disruption of
glucose homeostasis leads to an increase in leptin concentrations in seminal
plasma, which impairs sperm quality by promoting the generation of ROS [81].
Furthermore, excessive adiposity has been reported to enhance the activity of
aromatase, which facilitates the conversion of testosterone to estradiol, thereby
reducing testosterone levels. Lower testosterone levels have been associated with
mitochondrial dysfunction in Leydig cells [82]. Mitochondrial dysfunction leads
to oxidative damage to mitochondrial DNA and further promotes ROS production
[82]. Additionally, high-energy diet-induced pre-diabetic Wistar rat model
demonstrated a reduction in testicular antioxidant capacity and mitochondrial DNA
copy numbers [83]. This study also demonstrated that high-calorie diets reduce
the expression of testicular PGC-1
An increasing body of evidence suggests that a sedentary lifestyle predisposes
men to hypogonadism and reduced sperm motility [87], whereas physically active
men exhibit a more favorable anabolic hormonal environment and healthier semen
quality [88]. Moderate physical exercise has been shown to alleviate inflammation
and DNA damage in sperm [89]. For instance, a study involving 159 infertile men
and 143 fathers revealed that a lack of physical activity and excess body fat
relative to age-related norms were positively correlated with infertility [90].
Men engaging in outdoor activities (
One of the major sources of environmental ROS is the heat stress effect on the reproductive organs [93]. Study on cryptorchid mice has shown that exposure of the testes to abdominal temperature results in an increase in ROS production [94]. Research by Ikeda et al. [95] demonstrated that after exposing testicular cells to 43 °C for 1 hour, followed by a 23-hour incubation at 32.5 °C, intracellular peroxide levels were elevated, and H2O2 could attenuate heat stress-induced apoptosis. These findings indicate that heat stress can elevate testicular oxidative stress, leading to apoptosis of testicular cells. The increase in environmental temperature raises the metabolic rate of the testes, which consequently increases the oxygen demand of the tissue, resulting in higher ROS production [96]. Furthermore, the study by Paul et al. [97] demonstrated that heat-stressed mouse testes exhibited enhanced HIF1A expression, which in turn promoted the upregulation of antioxidant enzymes, such as heme oxygenase 1, to counteract the increased oxidative stress.
Airborne pollutants have also been shown to affect sperm membrane integrity, increasing the generation of free radicals and promoting oxidative stress, which negatively impacts sperm parameters [98]. Phthalates, commonly found in food packaging materials and personal care products, have been reported to induce oxidative stress and lead to sperm DNA damage [99]. Pant et al. [100] observed that the levels of di(2-ethylhexyl) phthalate in semen were negatively correlated with sperm quality and positively correlated with mitochondrial depolarization, increased ROS production, lipid peroxidation, and DNA fragmentation. This suggests that phthalates may deteriorate semen quality through ROS, lipid peroxidation, and mitochondrial dysfunction [100]. Another environmental pollutant, heavy metals, has been implicated in oxidative stress. Previous research has demonstrated that mice injected intraperitoneally with lead acetate exhibit a significant increase in peroxidative potential within testicular tissue, accompanied by reduced testicular weight, an increased incidence of abnormal sperm, and a decreased total sperm count [101]. Additionally, the concentrations of lead and cadmium in seminal plasma have been significantly correlated with biomarkers of oxidative DNA damage, such as 8-hydroxy-2′-deoxyguanosine (8-OHdG), indicating that these metals exacerbate oxidative stress in sperm and contribute to sperm DNA damage [102].
ROS originating from sperm cells or the external environment can disrupt the redox balance within sperm cells, resulting in increased oxidative stress and subsequent DNA damage. Sperm DNA damage can be categorized into two types based on its location: intratesticular and post-testicular. Intratesticular DNA damage occurs during the chromatin remodeling and apoptosis processes in sperm maturation, serving as a primary factor affecting sperm DNA integrity. Post-testicular DNA damage occurs as sperm traverse the reproductive tract. Both types of damage are associated with increased oxidative stress-induced damage to nuclear and mitochondrial DNA in sperm. Oxidative stress-induced sperm DNA damage encompasses DNA fragmentation, mitochondrial DNA damage, telomere shortening, Y chromosome deletions, and epigenetic abnormalities [15]. These alterations lead to genomic instability, replication errors, and transcriptional arrest [103]. ROS-induced DNA damage also exacerbates sperm apoptosis, reducing male fertility [104].
A significant correlation exists between ROS levels and sperm chromosomal
telomere DNA length. Under severe oxidative stress (ROS
Programmed cell death of germ cells is vital for normal development. Apoptosis eliminates abnormal or excess cells, enabling growth factors to promote the growth of neighboring cells and maintaining the stability of germ cell populations [119]. However, dysregulated apoptosis can result in sperm cell death, exacerbating male infertility [120].
Oxidative stress induces apoptosis through multiple pathways. As illustrated in Fig. 3, excessive ROS cause DNA damage, which triggers the phosphorylation of p53. Due to the limited DNA repair capacity in sperm cells, damaged DNA remains unrepaired, leading to further phosphorylation and activation of p53. This, in turn, activates the pro-apoptotic protein Bax, initiating the mitochondrial-dependent (intrinsic) apoptosis pathway [121]. In this pathway, cytochrome C within the mitochondria plays a pivotal role in initiating intrinsic apoptosis [122]. Emokpae and Chima [123] observed that the total antioxidant capacity of semen in infertile men declines significantly with age, while apoptotic markers such as cytochrome C and caspase-3 show a positive correlation with increasing age. In response to apoptotic signals such as ROS, pro-apoptotic proteins from the Bcl-2 family, including Bax and Bak, translocate from the interior of the mitochondria to the outer mitochondrial membrane. These proteins oligomerize or interact with the mitochondrial permeability transition pore to form channels [124]. These channels allow small molecules like cytochrome C to pass through the inner mitochondrial membrane into the cytoplasm, where cytochrome C binds to and activates Apaf1. This, in turn, activates caspase-9 [125]. Activated caspase-9 triggers a cascade of downstream caspases, including caspase-3, -6, and -7, which degrade intracellular proteins and ultimately lead to apoptosis [126].
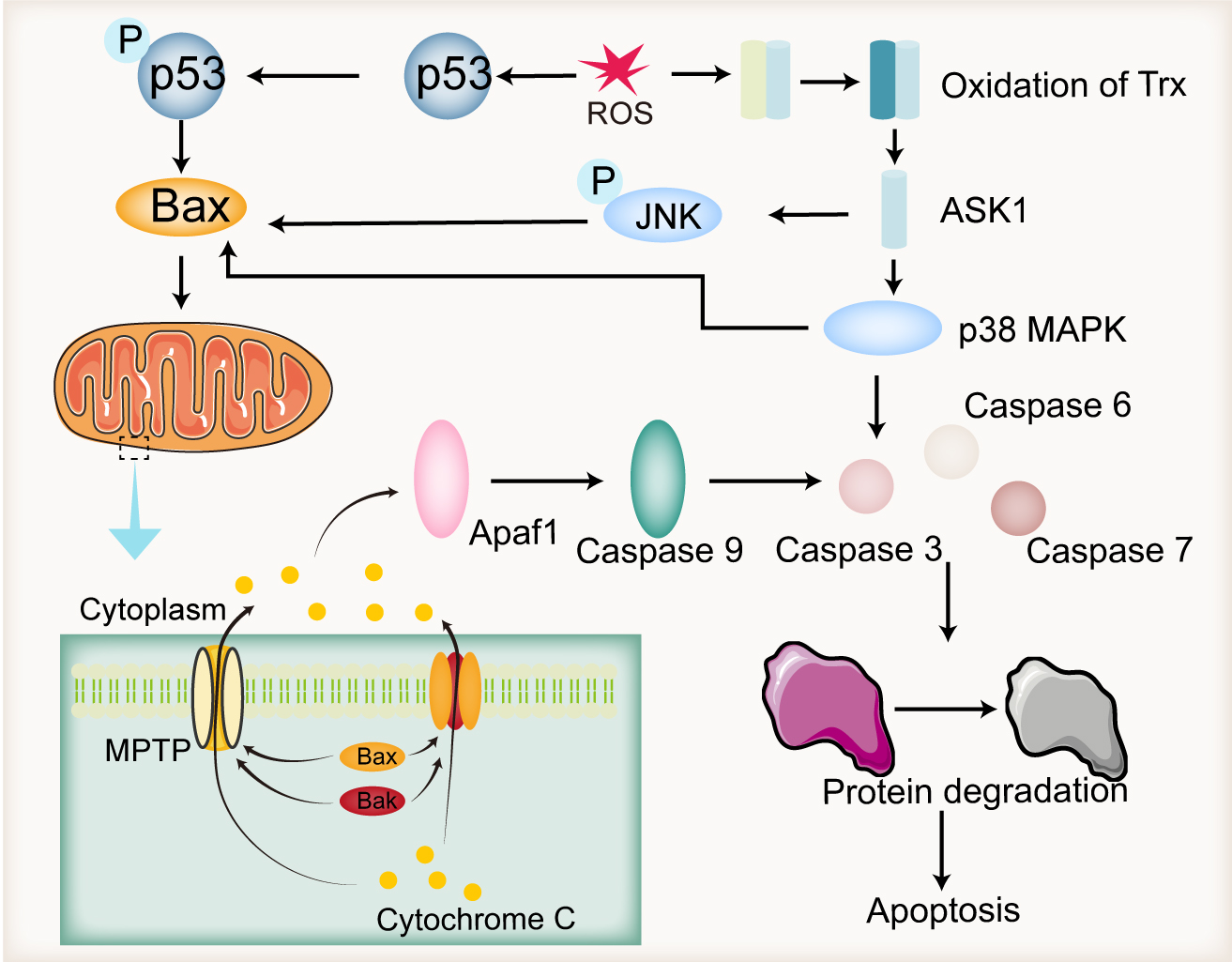 Fig. 3.
Fig. 3.
Endogenous apoptotic pathways induced by ROS. ROS-induced DNA damage leads to the phosphorylation of p53, which activates Bax, resulting in the release of cytochrome C from mitochondria. The released cytochrome C binds to Apaf1, activating it, which subsequently triggers caspase 9 activation, followed by the activation of caspase 3, caspase 6, and caspase 7. These caspases further degrade intracellular proteins, leading to apoptosis. Additionally, ROS can also induce apoptosis by oxidizing Trx, releasing ASK1, and activating the JNK and p38 MAPK pathways. Apaf1, apoptotic protease activating factor 1; ASK1, apoptosis signal-regulating kinase 1; Trx, thioredoxin; MPTP: mitochondrial permeability transition pore; Bax, Bcl-2-associated X protein; Bak, Bcl-2 antagonist/killer; MAPK, mitogen-activated protein kinase; JNK, c-Jun N-terminal kinase. Created by using Adobe Illustrator (version 28.7.1, Adobe Systems Incorporated, San Jose, CA, USA).
ROS can also promote apoptosis through the MAPK pathway. Under normal physiological conditions, ASK1 is inhibited by its binding to Trx, preventing stress-induced apoptosis [127]. However, in the presence of ROS, Trx undergoes oxidation, leading to the release of ASK1. As a member of the MAPK family, ASK1 phosphorylates downstream c-Jun N-terminal kinase (JNK) and p38 MAPK [128]. JNK induces apoptosis by activating pro-apoptotic Bcl-2 family members, Bax and Bak [129]. p38 MAPK regulates apoptosis by modulating the balance between anti-apoptotic (Bcl-2, Bcl-xL, Mcl-1) and pro-apoptotic (Bak, Bax, Bad, Bid) Bcl-2 family members [130], and also by activating caspase-3, thereby promoting cell death [131]. In infertile men, elevated levels of BAX and significantly reduced levels of BCL2 have been reported in semen [132].
In addition to the mitochondrial-dependent pathway, ROS also contribute to in the extrinsic apoptosis pathway, which is mediated by the activation of death receptors such as Fas. Upon binding with FasL, Fas trimerizes and recruits the adaptor protein FADD through its death domain [133]. FADD then recruits initiator caspase-8, forming the death-inducing signaling complex (DISC). DISC directly cleaves and activates caspase-3 to initiate apoptosis [134], and also cleaves the pro-apoptotic Bcl-2 family protein BID, thereby activating the mitochondrial-dependent apoptosis pathway [135].
Epigenetic changes involve modifications in gene activity without altering the underlying DNA sequence, thereby affecting cellular functions and developmental processes [136]. Among these, histone modifications represent a key epigenetic regulatory mechanism, which can be influenced by oxidative stress [137, 138]. In male germ cells, genomic DNA is organized into nucleosomes, which are octameric complexes composed of histones H2A, H2B, H3, and H4 [139]. Various post-translational modifications (PTMs) on histones influence the stability of the nucleosome octamer, driving chromatin conformation changes critical for spermatogenesis [140]. Methylation, a prominent PTM, typically occurs on lysine and arginine residues, with key sites including H3K4, H3K9, and H3K27 [141]. ROS have been shown to regulate histone methylation by activating the expression of SET7, a histone H3K4 methyltransferase [142]. The NADPH oxidase (NOX) family, a primary source of intracellular ROS, catalyzes the production of superoxide anions. Zhou et al. [143] demonstrated that the NOX inducer, succinate, enhances H3K4 trimethylation and H3K9 dimethylation while decreasing H3K27 trimethylation. Recent studies indicate that histone modifications in human sperm cells are critical for transmitting the epigenetic information necessary for embryonic development. Aberrant methylation levels at H3 sites have been associated with impaired spermatogenesis and defective epigenetic reprogramming in sperm [144, 145]. Furthermore, transgenic mice overexpressing lysine demethylase 1A (KDM1A), a lysine-specific demethylase, exhibit reduced H3K4 and H3K9 methylation levels in sperm, with 25% of their offspring dying within 21 days after birth [146]. KDM1A generates H2O2 as a by-product of demethylation, which can further inhibit its enzymatic activity by forming disulfide bonds between Cys600 and Cys618 residues, thus impairing demethylation [147]. These findings underscore the pivotal role of ROS in regulating histone methylation and demethylation by modulating the activities of methyltransferases and demethylases like KDM1A, ultimately impacting offspring development and survival. In addition to methylation, phosphorylation is another critical post-translational modification of histones. Brunner et al. [148] identified phosphorylation modifications at the T9 and T120 sites of H2B in mature mouse sperm, suggesting that these modifications may play a role in epigenetic traits underlying intergenerational inheritance. Metafora et al. [149] demonstrated that SV-IV, an immune-regulatory, anti-inflammatory, and sperm immune-protective protein derived from rat seminal vesicle epithelium, can compete with histone H1 as a substrate for protein kinase C phosphorylation. Additionally, the phosphorylation of SV-IV enhances the activity of Glutathione peroxidase (GPX) and horseradish peroxidase, indicating that histone modifications may be involved in the regulation of ROS. Phosphorylation of histone H3 plays a crucial role in the G2/M transition of the cell cycle and serves as an important marker for chromosome condensation. ROS can activate ERK and promote increased phosphorylation of histone H3 [150]. The phosphorylation of histone H3 is a prerequisite for the ubiquitination and degradation of excess histones, suggesting that ROS may facilitate the removal and degradation of intracellular histone H3 through phosphorylation, thereby regulating sperm DNA packaging [151]. Phosphorylation of H2AX is an early marker of the DNA damage response and aids in the recruitment of DNA repair factors [152]. Li et al. [153] demonstrated that hydrogen peroxide can induce phosphorylation of H2AX in human sperm cells in a time- and dose-dependent manner. Histone acetylation is also a significant post-translational modification. Salehi et al. [154] found that frozen sperm cultured with Beltsville extender exhibited elevated levels of ROS and H3K9 acetylation, along with reduced sperm parameters, suggesting that ROS may promote H3K9 acetylation, thereby decreasing sperm motility.
DNA methylation is another key epigenetic mechanism that plays a critical role in regulating gene expression. Abnormal sperm DNA methylation patterns have been frequently observed in patients with male infertility [155, 156]. Research by Tunc and Tremellen [157] demonstrated a significant inverse correlation between ROS levels and sperm DNA methylation, with antioxidant supplementation shown to restore normal methylation patterns. This suggests that ROS disrupt with the normal DNA methylation process in sperm [157]. DNA methylation primarily occurs at CpG islands located in the 5′ region of genes, catalyzed by the DNA methyltransferase (DNMT) family, which includes DNMT1, DNMT2, DNMT3A, DNMT3B, and DNMT3L, resulting in the formation of 5-methylcytosine [158]. Different DNMT enzymes operate at distinct stages of DNA methylation. While DNMT3B knockout has minimal impact on sperm, DNMT1 knockout leads to abnormal DNA methylation in mature sperm [159]. DNMT1 contains cysteine residues, making it highly susceptible to oxidative damage. ROS can oxidize DNMT1, impairing its methylation activity and leading to DNA methylation defects [160]. Furthermore, inactivation of DNMT1 has been shown to activate the p53 pathway, triggering apoptosis [161].
These studies underscore the pivotal role of ROS in modulating both histone and DNA methylation. ROS not only upregulate the expression of histone methyltransferases but also suppress the activity of histone demethylases. Through these mechanisms, ROS orchestrate the epigenetic landscape of sperm cells, influencing spermatogenesis and the epigenetic programming necessary for offspring development and survival. Additionally, ROS can inactivate DNMT1, a key member of the DNA methyltransferase family, by facilitating the formation of intramolecular disulfide bonds. This inactivation reduces DNA methylation, triggers apoptosis, and ultimately impairs sperm motility and fertility.
Under normal physiological conditions, spermatogenesis is an iron-dependent process, and disruption of iron metabolism can impair spermatogenesis and lead to reproductive dysfunction [162]. When ferritin is degraded under the guidance of the Nuclear receptor coactivator 4 (NCOA4) receptor, iron is released into the labile iron pool (LIP), which then triggers ferroptosis in testicular cells [163]. Ferroptosis is a form of cell death induced by iron-dependent oxidative stress, characterized by excessive intracellular iron accumulation and lipid peroxidation [164]. The iron in the LIP participates in the Fenton reaction, catalyzing the conversion of hydrogen peroxide into hydroxyl radicals, which in turn lead to the peroxidation of PUFAs on the cell membrane, causing membrane rupture and dysfunction [165]. Furthermore, iron-dependent oxidases, such as NADPH oxidase, can further promote ROS production, thus exacerbating ferroptosis [166]. Elevated iron and ROS levels have been observed in the semen of asthenozoospermic patients, indicating that ferroptosis contributes to sperm dysfunction [167]. Additionally, increased levels of ferroptosis, iron ions, and ROS have been detected in the semen of infertile smokers, with higher iron and ROS levels correlating with decreased sperm motility [168]. Research by Zhao et al. [169] demonstrated that ferroptosis inhibitors and deferoxamine could suppress ferroptosis and restore sperm concentration and motility in Busulfan-treated mice. These findings suggest that maintaining cellular iron balance is crucial for spermatogenesis, as excessive iron can induce ferroptosis and cause male reproductive disorders.
As a key regulator of ferroptosis, GPX4 plays a central role in alleviating lipid peroxidation and protecting cells from oxidative damage. Under the action of GPX4 and its cofactor glutathione (GSH), oxidized PUFAs are reduced to hydroxy phospholipids, thereby protecting the cell membrane [170]. In infertile men diagnosed with oligospermia, GPX4 expression in sperm is significantly reduced [171], and GPX4 knockout mice show a significant decrease in sperm count and exhibit infertility [172]. The activity of GPX4 depends on selenium and GSH [173], and selenium supplementation has been shown to enhance GPX4 activity and promote germ cell proliferation [167]. GSH, an important antioxidant, is synthesized with the involvement of system Xc-, which exchanges extracellular glutamate for cystine. Cystine is then reduced to cysteine inside the cell, which is crucial for GSH synthesis [174]. This is consistent with the study by Hamashima et al. [175], which showed that system Xc- knockout mice had an increased number of immature spermatogenic cells in the cauda epididymis. Furthermore, increased GSH levels have been shown to improve sperm quality and testicular histomorphology in diabetic mice [176].
Increased oxidative stress is a major cause of decreased sperm motility. Consequently, the use of antioxidants is considered one of the primary effective treatments for male infertility [177]. Antioxidant therapy has been shown to significantly reduce ROS level and improve sperm motility [178]. Recent reports also suggest that certain plant extracts can enhance sperm activity through their antioxidant properties [179]. The effects of antioxidants on sperm are summarized in Table 1 (Ref. [180, 181, 182, 183, 184, 185, 186, 187, 188, 189, 190, 191, 192, 193, 194, 195, 196, 197, 198, 199, 200, 201, 202]).
| Antioxidant | Effects | References |
| Glutathione | Reduce sperm DNA fragmentation and morphological anomalies while enhancing sperm count restoration. | [180] |
| N-Acetylcysteine | Diminish apoptosis in reproductive cells to bolster sperm vitality. | [181, 182] |
| vitamins C | Ameliorate the integrity of sperm DNA by reducing fragmentation and concurrently improving sperm vigor. | [183] |
| Carnitine | Enhance sperm kinematics and energetic potential. | [184, 185] |
| Edaravone | Reduce the prevalence of immature spermatozoa. | [186] |
| Aqueous extract of celery | Enhance sperm numerical density. | [187] |
| Cornus mas fruit extract | Elevate spermatozoal antioxidant defenses alongside sperm quantity. | [188] |
| Satureja Khuzestanica essential oil | Upgrade sperm quality and augment fecundity indices. | [189] |
| Ginger and cinnamon | Optimize sperm viability through enhanced motility. | [190] |
| 1 |
Prevent diabetes-induced decline in testicular antioxidants, protecting the reproductive system and maintaining sperm count and motility. | [191] |
| Tea powder | Increase testicular antioxidants, reduces MDA, alleviates oxidative damage, and improves testicular index. | [192] |
| Curcumin nanomicelle | Improve antioxidant capacity, reduces MDA, and boosts sperm count, concentration, and motility in infertile men. | [193] |
| Eruca sativa aqueous extracts | Reduces testicular MDA levels, increases thiol levels, and enhances the activity of SOD, CAT, and GPx, preventing testicular damage in rats, and improving sperm density, motility, and viability. | [194] |
| Ethanol extract of Ionidium suffruticosum (L.) Ging leaves | Increase testicular SOD and CAT levels, promotes spermatogenesis, increases sperm count, and reduces sperm aggregation in rats. | [195] |
| Pomegranate juice | Restore the decline in testicular SOD, CAT, GPx, GST, GR, and GSH levels, and improves the degeneration of germ cells and Leydig cells, as well as sperm morphological. | [196] |
| Lycopene | Enhance total sperm antioxidant capacity, increases ejaculate volume, total sperm count, sperm concentration, and total motility. | [197] |
| Aqueous leaf extract of Moringa oleifera | Reduce sperm reactive oxygen species levels, improve sperm DNA fragmentation, and maintain sperm function. | [198] |
| Camellia sinensis | Enhance the CAT and SOD enzyme activities and glutathione levels in rat sperm, thereby protecting against oxidative stress-induced sperm damage. | [199] |
| Aspalathin linearis | Increase the SOD enzyme level, reduce the MDA level, and enhance sperm concentration in rats. | [200] |
| Lactobacillus rhamnosus CECT8361, Bifidobacterium longum CECT7347 | Mitigate DNA fragmentation to reinforce sperm motility. | [201] |
| FamiLact | Enhance sperm concentration and motility, attenuate sperm morphological abnormalities, oxidative stress, and DNA fragmentation indices. | [202] |
MDA, malondialdehyde; SOD, superoxide dismutase; CAT, catalase; GPx, glutathione peroxidase; GST, glutathione S-Transferase; GR, glutathione reductase; GSH, glutathione.
Glutathione, a thiol-containing tripeptide, is a potent antioxidant activity. Research has shown that reduced levels of glutathione in seminal plasma are associated with impaired sperm viability and motility [203]. Chemotherapeutic agents such as bleomycin, etoposide, and cisplatin, commonly used in the treatment of testicular cancer, can induce the ROS production [204]. The use of glutathione has been reported to significantly mitigate bleomycin, etoposide, and cisplatin-induced sperm DNA fragmentation and morphological abnormalities in rats [180]. N-Acetylcysteine (NAC), a precursor of glutathione, effectively, scavenges ROS effectively as an antioxidant. In vitro study has demonstrated that NAC inhibits apoptosis in human testicular germ cells [181]. Clinical trials have shown that NAC significantly improves sperm oxidative stress indices and motility in infertile men [182]. Vitamin C neutralizes hydroxyl radicals, superoxide anions, and hydrogen peroxide, reducing oxidative stress. The concentration of vitamin C in seminal plasma is more than ten times higher than in blood plasma, indicating its critical antioxidant role in semen [205]. A marked decrease in vitamin C levels and total antioxidant capacity has been observed in patients with asthenozoospermia, indicating a significant positive correlation between lower vitamin C levels and male infertility [206]. However, clinical outcomes regarding vitamin C supplementation remain mixed. Rolf et al. [207] reported that high doses of vitamin C and E did not improve sperm concentration and motility in patients with asthenozoospermia. Nonetheless, recent study suggests that a combination of zinc, vitamin C, and vitamin E can effectively reduce DNA fragmentation, improve sperm motility, and decrease oxidative stress in infertile patients [183]. Further investigation is needed to establish the therapeutic efficacy of vitamin C in male infertility. Additionally, low concentrations (100 mmol/L) of the antioxidant carnitine have been shown to improve sperm motility and viability in vitro. However, this effect appears to be mediated through the regulation of specific gene expressions (Vasa, Dazl, Acr, and Prm1) rather than direct ROS modulation [184, 185]. Edaravone, a drug with notable antioxidant and neuroprotective properties, functions primarily through free radical scavenging. Recent study has shown that edaravone reduces the proportion of immature sperm in busulfan-induced azoospermia and improves azoospermia by scavenging free radical and regulating autophagy in testicular tissue [186].
Numerous plant extracts, rich in antioxidant compounds, present promising
solutions for addressing male infertility. For example, celery, which is rich in
phenols and coumarins, has been shown to significantly increase sperm count in
mice treated with celery extract [187]. Similarly, Safarnavadeh and Rastegarpanah
[189] demonstrated that Satureja Khuzestanica essential oil, extracted from
Satureja Khuzestanica Jamzad, contains up to 90.8% carvacrol, which effectively
enhances sperm quality and fertility by acting as an antioxidant [208]. This
suggests that the active components in carvacrol can significantly improve sperm
quality by reducing oxidative stress, though further clinical study is required
to substantiate its efficacy. In addition to carvacrol, other plants also exhibit
notable antioxidant properties. Zarei et al. [188] demonstrated that
Cornus mas extract significantly prevented the reduction of total
antioxidant capacity induced by methotrexate treatment in mice. Additionally, it
effectively mitigated methotrexate-induced sperm DNA damage and the decline in
sperm maturation [188]. Using a diabetic mouse model, Khaki demonstrated that
both individual and combined use of ginger and cinnamon significantly improve
serum antioxidant levels (TAC, SOD, GPX, and catalase) while markedly enhancing
sperm vitality and motility [190]. Curcumin, extracted from the rhizomes of
Curcuma longa, possesses potent antioxidant properties, with its antioxidant
activity being 300 times stronger than that of vitamin E [209]. A study by
Alizadeh et al. [193] demonstrated that daily supplementation with 80 mg
of curcumin nanoparticles significantly improved total antioxidant capacity,
reduced MDA levels, and enhanced sperm count, concentration, and motility in
infertile men. Research by Hamden et al. [191] showed that the combined
use of 1
Probiotics can enhance testicular function and promote sperm regeneration through antioxidant effects and modulation of the gut microbiota. Valcarce et al.’s research [213] demonstrated that the antioxidant probiotic strains Lactobacillus rhamnosus CECT8361 and Bifidobacterium longum CECT7347 improved sperm quality and offspring survival rates in zebrafish. Subsequent human trials confirmed these effects, revealing that the intake of these strains reduced hydrogen peroxide levels in sperm cells by approximately 3.5-fold, decreased DNA fragmentation, and enhanced sperm motility [201]. This finding supported by Abbasi et al. [202], who administered the probiotic supplement FamiLact (containing Lactobacillus rhamnosus, Lactobacillus casei, Lactobacillus bulgaricus, Lactobacillus acidophilus, Bifidobacterium breve, Bifidobacterium longum, and Streptococcus thermophilus) to male subjects with idiopathic infertility. Significant increases in sperm concentration and motility were observed, along with reductions in abnormal sperm morphology, oxidative stress, and DNA fragmentation [202]. Further study indicated that FamiLact administration in patients post-varicocelectomy resulted in improved sperm concentration and a higher proportion of normal sperm [214]. Additionally, a study on male Japanese quails supplemented with Bifidobacterium longum and mannan oligosaccharides found that probiotic and prebiotic supplementation significantly upregulated the expression of SOD and catalase in testicular cells, thereby enhancing male fertility in juvenile quails [215].
Male infertility affects approximately half of infertile couples, with elevated ROS levels in sperm indicating that oxidative stress may plays a pivotal role in this condition. ROS can arise from both endogenous and exogenous sources. Mitochondria serve as the primary endogenous source of ROS within sperm, where electron leakage from the mitochondrial electron transport chain generates superoxide anions, thereby exacerbating oxidative stress. This heightened oxidative stress promotes lipid peroxidation, and its by-products of this process can damage mitochondrial proteins, further enhancing electron leakage. Exogenous factors including gut microbiota dysbiosis, circadian rhythm disruptions, smoking, leukocyte-derived ROS, and alcohol consumption, also contribute to excessive ROS production. These external sources increase oxidative stress in sperm, resulting in DNA damage and fragmentation the risk of male infertility. Additionally, oxidative stress activates both mitochondria-dependent and death receptor-dependent apoptotic pathways, leading to sperm apoptosis and further contributing to infertility. Moreover, oxidative stress can also induce epigenetic modifications by modulating the expression of histone methyltransferases and demethylases, leading to altered histone methylation patterns, which may impact offspring development. It also disrupts sperm DNA methylation by impairing DNA methyltransferase activity, thereby reducing sperm motility.
Antioxidants, including small molecule antioxidants, plant extracts, and probiotics, have shown considerable potential in alleviating oxidative stress in sperm and enhancing motility. However, the beneficial effects of these treatments have primarily been demonstrated at the cellular and animal levels, and further clinical studies are necessary to validate their therapeutic efficacy in male infertility. In addition to histone and DNA methylation, epigenetic regulation involves various mechanisms, such as miRNA expression and histone acetylation, which also influence sperm function. Future research should focus on the interplay between ROS, epigenetic modifications, and sperm motility to enhance our understanding of oxidative stress and its impact on male fertility. Abnormal sperm function is a key contributor to male infertility, with sperm abnormalities are often linked to reproductive tract disorders, including varicocele, vas deferens obstruction, cryptorchidism, and orchitis. Future studies should explore the relationship between oxidative stress and reproductive tract disorders to further advance the clinical application of antioxidants in treating male infertility.
YZ, HZ, HY, PH, JZ, and YL contributed to the literature review and manuscript writing. YZ and HZ also critically reviewed and revised the manuscript for important intellectual content. All authors read and approved the final version of the manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This work was supported by Yunnan Provincial Department of Science and Technology Yunnan University of Traditional Chinese Medicine Applied Basic Research Joint Special Project (202301AZ070001-166).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.