



 , Ye Cheng 2,*
, Ye Cheng 2,*1 Department of Anesthesia Surgery Center, The West China Xiamen Hospital of Sichuan University, 361021 Xiamen, Fujian, China
2 Department of Cardiology, The Xiamen Cardiovascular Hospital of Xiamen University, 361000 Xiamen, Fujian, China
Abstract
Hypertriglyceridemia, characterized by increased triglyceride (TG) concentrations, is considered the most important risk factor for cardiometabolic disorders, including dyslipidemia, atherosclerotic cardiovascular diseases, and non-alcoholic fatty liver disease (NAFLD). Recently, the angiopoietin-like protein (ANGPTL) family, which comprises ANGPTL1 to ANGPTL8, was confirmed to play an important role in modulating lipoprotein lipase (LPL) activity. However, understanding of the underlying mechanisms remains limited. Importantly, emerging evidence has linked several transcriptional and post-transcriptional factors to the potential alteration of TG metabolism via ANGPTL proteins. This review focused on the similarities and differences in the expression, structural features, and modulatory profile of three ANGPTLs: ANGPTL3, ANGPTL4, and ANGPTL8. In addition, the regulatory functions of those three ANGPTLs in modulating LPL were summarized to provide potential therapeutic and clinical strategies for hypertriglyceridemia and its related cardiometabolic disorders.
Keywords
- angiopoietin-like proteins
- triglycerides
- lipoprotein lipase
- metabolism
- hypertriglyceridemia
Hypertriglyceridemia, characterized by increased triglyceride (TG) levels, is considered the most important risk factor for cardiometabolic disorders, including dyslipidemia, atherosclerotic cardiovascular diseases, and non-alcoholic fatty liver disease (NAFLD) [1]. However, understanding of the underlying mechanisms involved in these disorders remains limited. Severe hypertriglyceridemia may occur when TGs in blood circulation cannot be hydrolyzed by lipoprotein lipase (LPL), an essential rate-limiting enzyme that significantly modulates serum TG metabolism and hydrolyzed serum TG-rich lipoproteins (TRLs), such as very-low-density lipoprotein (VLDL) and chylomicron (CM) [2]. Consequently, serum TRLs might produce excessive non-esterified fatty acids (NEFAs) for peripheral tissue uptake [3]. In contrast, the normal activity and biological function of LPL are significant for keeping a well-balanced metabolism of serum TGs or TRLs.
Emerging evidence recently showed that LPL, produced and released by parenchymal cells, could be transferred by glycosylphosphatidylinositol-anchored high-density lipoprotein binding protein 1 (GPIHBP1) from the basolateral epithelium into the capillary lumen to perform its hydrolytic function [4]. During the different stages of this progression, LPL might be influenced by several modulatory factors at post-transcriptional levels, such as apolipoprotein C (ApoC), apolipoprotein E (ApoE), GPIHBP1, and the angiopoietin-like protein (ANGPTL) family.
Notably, three members of the ANGPTL family, including ANGPTL3, ANGPTL4, and ANGPTL8, were extensively analyzed for their important role in modulating TG metabolism by affecting LPL [5]. As demonstrated by previous studies, ANGPTL3, ANGPTL4, and ANGPTL8 share a common structure but differ in tissue expression [6, 7]. Moreover, owing to their established role in modulating TG metabolism and the risk of hypertriglyceridemia, ANGPTLs have also been considered as treatment options for hypertriglyceridemia and its related cardiovascular diseases. This review summarized the current understanding of the association between these three important ANGPTL proteins with LPL and serum TG activity. Furthermore, the emerging results that focused on the novel therapeutic targets in hypertriglyceridemia and related ANGPTL protein-mediated complications in daily clinical practice were also listed.
The ANGPTL protein family members have a similar structure to angiopoietins [8].
According to previous reports, diverse ANGPTL members have similar structures but
different functions. Specifically, ANGPTL3, encoded by the ANGPTL3 gene,
is located on chromosome 1p31 in humans and chromosome 4 in mouse models;
hepatocytes produce ANGPTL3, which is marginally expressed by renal cells [9].
Additionally, the ANGPTL3 protein contains a 460 amino acid peptide, which
includes a distinctive signal peptide sequence, an N-terminal coiled-coil domain,
and a C-terminal globular fibrinogen homology domain [10]. The N-terminal region
is confirmed to upregulate serum TG concentrations by inhibiting LPL activity;
similarly, the fibrinogen-like domain was verified to combine with the integrin
Comparatively, ANGPTL4 was discovered in 2000 and is produced and released by several cells, such as adipocytes, hepatocytes, and intestinal cells [12]. The human gene encoding ANGPTL4 is conserved among diverse species and shares a sequence homology of approximately 80% with the gene in mice. ANGPTL4 is located on chromosome 13.3p in humans and consists of seven exons that encode a 406 amino acid glycoprotein. The full-length ANGPTL4 protein is a higher-order oligomeric structure. Notably, several investigations demonstrated that the extracellular storage of ANGPTL4 could suppress the biological activity of LPL within 3T3-L1 pre-adipocytes, where the oligomers of the ANGPTL4 gene were formed [13]. Nonetheless, another study showed that ANGPTL4 could not significantly inhibit LPL activity [14].
ANGPTL8 is encoded by the Gm6484 gene in mice and the C19 gene in humans and is also produced by hepatocytes and adipocytes [15]. Eight different ANGPTL protein members (ANGPTL1 to ANGPTL7) share a similar structure, which contains a coiled-coil domain region, an N-terminal signal sequence, and a large fibrinogen/angiopoietin domain region [16]. However, the latter domain region is absent in ANGPTL8, resulting in a 22kDa protein, which is less than half the size of ANGPTL3 and ANGPTL4 [17]. Notably, the discordant structure of these ANGPTL proteins induces different functions in modulating TG levels and the risk of hypertriglyceridemia. The diverse structures of the ANGPTL protein family are listed in Fig. 1.
 Fig. 1.
Fig. 1.
Schematic diagram of the ANGPTL3, ANGPTL4, and ANGPTL8 structures. The ANGPTL protein family consists of three domains: the N-terminal signal peptide structural domain, one or two N-terminal coiled-coil domains, and a C-terminal fibrinogen-like structural domain. The members of the ANGPTL protein family differ slightly in composition. Abbreviations: ANGPTL8, angiopoietin-like protein 8; LPL, lipoprotein lipase; ANGPTL3, angiopoietin-like protein 3; ANGPTL4, angiopoietin-like protein 4.
As reported, the ANGPTL3 gene expression is modulated within hepatocytes by hepatocyte nuclear factor-1 (HNF-1) [18]. Conversely, the ANGPTL4 and ANGPTL8 gene expression levels differ among species. In detail, ANGPTL4 is a secreted protein selectively expressed within adipocytes, hepatocytes, and the placenta under fasting conditions [19]. Consistently, the expression of the ANGPTL4 gene was abundant within adipocytes isolated from lean mice [20]; however, in humans, the intracellular ANGPTL4 gene transcript in adipocytes accounts for only approximately 15% within hepatocytes [21].
The ANGPTL8 gene was found in both hepatocytes and adipocytes; however, the expression is relatively higher in hepatocytes than in adipocytes [22]. Two independent research demonstrated that mouse presented higher ANGPTL8 and ANGPTL4 gene expression levels in brown adipocytes than in white adipocytes, which might be responsible for the primary function of brown adipocytes in thermos-modulation and energy balance [23, 24]. In conclusion, different ANGPTL3, ANGPTL4, and ANGPTL8 expression levels in tissues could induce diverse tissue-specific effects in modulating TG metabolism and related cardiovascular diseases.
ANGPTL3, ANGPTL4, and ANGPTL8 expressions have been shown to differ through
tissue-specific regulation. A previous study demonstrated that peroxisome
proliferators-activated receptors (PPARs)-
Other members of the PPAR family, such as PPAR-
Several other nuclear transcriptional factors, including hypoxia-inducible
factor-1 (HIF-1), Toll-like receptor (TLR), and liver X receptor (LXR), were
identified as important modulators of the ANGPTL protein family expressions [37].
In detail, emerging evidence showed that the administration of
lipopolysaccharide, a special activator of TLR-4, could lead to reduced
ANGPTL3 gene expression levels in hepatocytes and increased
ANGPTL4 gene expression levels in adipocytes [38]. In addition,
lipopolysaccharide also induced decreased ANGPTL4 gene expression in
macrophages [39]. It is worth noting that a remarkably increased expression of
the ANGPTL4 gene in macrophages following TLR-4 stimulation further
promoted the risk and development of dyslipidemia, indicating a mechanism through
which ANGTPL4 can modulate dyslipidemia via TLR-4 [40]. On the other hand, it has
been proposed that hypoxia, identified as one of the main inducers, modulates
ANGPTL4 and ANGPTL8 gene expression by influencing the
expression levels of HIF-1 [41]. Following an in-depth investigation,
HIF-1
Notably, LXR is another transcriptional factor that has received substantial attention recently. Using T0901317, a LXR ligand, could promote the risk of dyslipidemia via upregulating the expression levels of the ANGPTL3 gene in hepatocytes, which further suppresses the hydrolysis of serum TG concentrations [45]. Likewise, it was shown that in mammals, LXR could directly combine with ANGPTL3 [46]; furthermore, the LXR ligand and LXR–retinoid X receptor (RXR) complex could also directly combine with the ANGPTL3 gene promoter element. Through this step, diverse ligands could facilitate promoter activity in the ANGPTL3 gene [47].
More recently, increasing evidence has shown that prolonged cold exposure could also regulate ANGPTL4 gene expression levels in brown and white adipocytes, which further induced a relatively increased LPL activity [48]. Additionally, this phenomenon could promote the delivery of lipids for thermogenesis. Since thermogenesis plays an important role in promoting obesity, an essential ANGPTL4 function has been proposed to influence the risk of obesity [49]. Comparatively, the ANGPTL8 gene expression levels were confirmed to be inversely altered under a cold exposure environment [50].
Under fasting conditions, ANGPTL4 is induced early, reducing the delivery extent of TGs into white adipocytes and releasing free fatty acids into the circulation for subsequent uptake by multiple oxidative-related cells [51]. Meanwhile, ANGPTL3, as a hepatic-specific factor, was confirmed to inversely increase the endocytosis process of circulating VLDL-containing TGs into white adipocytes under feeding conditions rather than into oxidative cells [52]. Consequently, an absence of ANGPTL3 might disrupt the balance, indicating that diverse ANGPTL proteins might affect energy intake and consumption.
Nevertheless, it has been shown that TG accumulation was maintained via facilitating glucose uptake, which transforms glucose into free fatty acids in white adipocytes and not hepatocytes isolated from the ANGTPL3 gene-deficient mouse models. Following these in-depth investigations, the authors confirmed that inhibiting ANGPTL3 could upregulate insulin sensitivity, which might explain the modulatory role of ANGPTL3 in glucose metabolism [53]. In conclusion, the results indicated that ANGPTL3 and ANGPTL4 perform diverse functions in modulating serum TG metabolism. Specifically, ANGPTL3 is dominant under feed conditions, whereas ANGPTL4 plays a greater function under fasting conditions [54].
ANGPTL4 was not involved in the bile acid-mediated decrease of the serum TG concentrations; however, ANGTPL4 was confirmed to facilitate the absorption of bile acids via gut microbiota under taurocholic acid supplementation [55]. ANGPTL4 was also verified to play an important role in glucocorticoid-induced TG catabolism. Indeed, an in-depth investigation demonstrated that the ANGPTL4 gene sequence contained a glucocorticoid receptor binding site, which could induce ANGPTL4 and glucocorticoid binding. Via this method, glucocorticoid could upregulate the expression levels of the ANGPTL4 gene; simultaneously, ANGPTL4 is also required for the function of glucocorticoid, which induces the transportation of TGs from white adipocytes to hepatocytes [36]. The features and basic functions of ANGPTL3, ANGPTL4, and ANGPTL8 are summarized in Table 1.
| Name | Chromosome | Tissue expression | Basic functions |
| ANGPTL3 | 1 | Liver | Pro-angiogenesis, lipid metabolism, pro-inflammation, cancer development |
| ANGPTL4 | 19 | Liver, adipose tissue, kidney, intestine, heart | Pro-angiogenesis, lipid metabolism, pro-inflammation, cancer development |
| ANGPTL8 | 19 | Liver, adipose tissue | Lipid metabolism, pro/anti-inflammation, cancer development |
It has been extensively proposed that the ANGPTL proteins could inhibit LPL-induced TG hydrolysis. Nevertheless, the potential mechanisms whereby diverse ANGPTL proteins modulate the LPL-induced TG metabolism remain poorly understood. For instance, it was demonstrated that ANGPTL3 could suppress the biological activity of LPL in combination with the N-terminal coiled-coil domain, which promotes the dissociation of active LPL dimers into monomers. Importantly, the monomers lost their biological function in modulating TG metabolism. In contrast, ANGPTL4 was verified to stimulate LPL degradation in diverse metabolic-related cells, such as adipocytes, macrophages, and hepatocytes, which resulted in reduced intracellular LPL concentrations, suggesting that ANGPTL4 also modulates intracellular LPL metabolism, thereby influencing intracellular TG concentrations [56].
As noted above, the structure of circulating ANGPTL8 was confirmed to be homologous with ANGPTL3 [57]. Consequently, this special structure of ANGPTL8 could be a functional linkage with similar functions to ANGPTL3 in modulating serum TG metabolism. Subsequently, independent research indicated that ANGPTL8 participated in lipid modulation in circulation [58]; however, the research could not detect the accurate function of ANGTPL8 in modulating LPL activity under similar conditions to those used for ANGPTL3 [59]. Intriguingly, independent research exists that demonstrated that ANGPTL8, in combination with ANGPTL3, could facilitate the cleavage of ANGPTL3; furthermore, genetic analysis results in mice suggested that ANGPTL3 could be stimulated by ANGPTL8, indicating that ANGPTL8 and ANGPTL3 might function via a co-dependent association [60]. However, owing to limited published literature, additional basic experiments are required to elucidate the co-functions of ANGPTL3 and ANGPTL8 in modulating TG metabolism within circulation.
The ANGPTL8 gene expression levels can be regulated under feeding conditions [61]; meanwhile, both central and peripheral ANGPTL8 were consistently demonstrated to influence mammalian appetites. Inhibition of the neuropeptide Y-positive neurons in the dorsomedial hypothalamus in the brain was shown to activate the modulatory role of ANGPTL8 [62]. In addition, exogenous injection of ANGPTL8 protein into the C57BL/6J mice reduced the visceral white adipose mass, potentially via appetite inhibition. Nevertheless, central injection of ANGPTL8 protein into C57BL-6 mice reduced the activity of serum LPL and increased white adipose mass compared with the control mice; reduced serum levels of free fatty acids were also observed in these mice [62].
Recently, a relatively abundant expression of the ANGPTL3 gene was observed in hypothalamic neurons, which modulated energy homeostasis via hypothalamic LPL stimulation. Moreover, food intake and body weight decreased [63]. Furthermore, an ANGPTL3-induced reduction in hypothalamic neuropeptide Y-positive neurons and a decrease in agouti-related peptide expression might be responsible for anorexia and weight loss, indicating that ANGPTL3 could modulate appetite, thereby altering TG metabolism and weight [63].
Dyslipidemia, characterized by elevated TG and low-density lipoprotein cholesterol (LDL-C) serum levels and lower high-density lipoprotein cholesterol (HDL-C) serum levels, strongly correlates with obesity, type 2 diabetes mellitus, and cardiovascular diseases [64]. As mentioned above, diverse ANGPTL proteins are considered essential modulators of lipid profile metabolism by influencing LPL activity, indicating that dysfunctional ANGPTLs could induce aberrant LPL-mediated lipid catabolism.
Emerging evidence from genome-wide association studies (GAWS) has indicated that single nucleotide polymorphisms (SNPs) in the ANGPTL3 gene locus strongly correlate with serum TG concentrations [65, 66]. Data from the Dallas Heart Study population showed that the loss-of-function (LOF) ANGPTL3 gene variant also leads to lower serum TG concentrations, increasing the prevalence of hypertriglyceridemia and atherosclerosis [67]. Similar results were observed in research using ANGPTL3 gene-deficient mouse models [54]. On the contrary, injection of REGN1500, as a monoclonal antibody to ANGPTL3, could decrease serum TG concentrations in mouse models and monkey models, indicating that anti-ANGPTL3 antibodies could represent a potential therapeutic strategy for hypertriglyceridemia [68]. A research study focusing on a traditional Chinese medicinal product, named paeoniflorin, showed that this medicine could lower the risk and development of hypertriglyceridemia by upregulating N-acetyl-galactosaminyl transferase 2 (GALNT2) [69]. Given that GALNT-2 is a modulator that suppresses ANGPTL3 activity, we could reasonably speculate that paeoniflorin reduces TG serum concentrations by inhibiting ANGPTL3 and LPL [69].
In addition to the modulatory role of serum TG levels, ANGPTL3 was demonstrated to modulate serum cholesterol catabolism. For instance, nonsense variants in the ANGPTL3 gene lead to lower LDL-C serum concentrations; additionally, the individuals who possess a nonsense mutation in the ANGPTL3 gene presented hypocholesterolemia characterized by extremely lower serum concentrations of LDL-C with increased LDL receptor (LDLR) gene expression [70]. In addition to modulating LDL-C and TG, ANGPTL3 could also modulate serum concentrations of HDL-C by inhibiting endothelial lipase, which promotes the hydrolyzation of HDL particles [71]. Given that lower HDL-C serum concentrations correlate with an increased risk of cardiovascular diseases, this modulatory function of HDL-C by ANGPTL3 might be a potential mechanism through which ANGPTL3 regulates cardiovascular disease risk. Agents that lower TG levels, including probucol, have been shown to induce HDL-C remodeling, consequently increasing HDL-C serum concentrations by lowering serum ANGPTL3 [72]. Consistent with this notion, another anti-ANGPTL3 agent, named WO2012174178, has been shown to reduce the prevalence of cardiovascular diseases by enhancing lipolytic enzymes and lowering TG levels [72]. Similar results have been observed in experiments using apolipoprotein E (ApoE)-deficient mice. These results showed that 3, 4, 5, 6-tetrahydroxyxanthone treatment reduced ANGPTL3 gene expression [73]. Recently, two botanical medicines, named undaria pinnatifida soluble fiber and Kaempferol, have been confirmed to decrease ANGPTL3 gene expression and further prevent hypertriglyceridemia in mice [74]. The results described above could provide evidence for ANGPTL3 as a novel target to improve hypertriglyceridemia and its related cardiovascular diseases in clinical practice.
Similar to ANGPTL3, ANGPTL4 was confirmed by a study to affect serum TG metabolism positively [74]. Importantly, ANGPTL4 gene silencing has been shown to induce elevated LPL-mediated clearance of circulating TG; furthermore, ANGPTL4 absence in brown adipocytes could increase free fatty acid oxidation, promoting glucose tolerance. In humans and under fasting conditions, carriers of ANGPTL4 gene mutations, such as E40K variants, were shown to possess lower TG serum concentrations alongside lower risks of diabetes mellitus [75]. Otherwise, mice lacking the ANGPTL4 gene have also exhibited improved postprandial hypertriglyceridemia, providing a treatment for familial hypercholesterolemia [76]. It is worth noting that ANGPTL4 also reduced the formation of macrophages into foam cells, suggesting that ANGPTL4 acts as a double-edged sword, modulating the risk of atherosclerosis by affecting foam cell formation [77]. These results also propose a novel treatment target for hypertriglyceridemia.
Recently, ANGPTL4 has been shown to influence HDL function in humans by protecting the HDL particles from endothelial lipase hydrolysis [78]. Conversely, in individuals with type 2 diabetes mellitus, ANGPTL4 could facilitate the hydrolysis of HDL particles, promoting free fatty acid production by suppressing the biological activity of endothelial lipase [78]. However, an independent investigation suggested that diverse forms of ANGPTL4 might have different functions in lipid metabolism. To be more specific, the full-length ANGPTL4 was shown to correlate with HDL concentration and metabolism. In contrast, the truncated type of ANGPTL4 was responsible for modulating the function of LDL particles [79].
Due to technological advances, major breakthroughs have been made to explain the association between ANGPTL8 and serum TG concentrations. Indeed, ANGPTL8 was confirmed to be positively associated with serum TG concentrations via downregulating LPL activity, indicating that ANGPTL8 could modulate the risk of hypertriglyceridemia by affecting LPL [80]. In addition to the modulatory role on serum TG levels, ANGPTL8 was also found to be inversely correlated with the cholesterol efflux capacity of HDL particles. Since HDL-induced cholesterol efflux is important in improving dyslipidemia-related cardiovascular diseases, we could infer from these results that ANGPTL8 might also be negatively associated with cardiovascular diseases, independently of HDL-C serum concentrations [81]. Nevertheless, a larger-scale investigation is still needed to explore the relationship between ANGPTL8 and the metabolism of TGs further.
According to previous studies, ANGPTL3 has been strongly linked to playing an important function in modulating adipogenic differentiation, consequently influencing the pathogenic development of obesity [82]. In detail, ANGPTL3 could directly target the surface of adipocytes, which further stimulates the lipolysis process of intracellular TGs. Subsequently, this process facilitates the significant release of free fatty acids and glycerol within circulation by adipocytes [83]. Nevertheless, the authors demonstrated that ANGPTL3-deficient mice fed a high-calorie diet (HCD) presented reduced epididymal adipocyte weights. In contrast, the adipocyte sizes remained similar to those isolated from control healthy mice, suggesting that ANGPTL3 modulates the hyperplasia rather than the hypertrophy of adipocytes [82].
It has been confirmed for several decades that the animal models with relatively higher ANGPTL4 gene expression presented reduced adipose percentages. This finding has been replicated in a study [84]. According to these results, overexpression of the ANGPTL4 gene could inhibit the clearance process of serum TGs and prevent fat accumulation within adipocytes. Moreover, ANGPTL4 activates fat mobilization and facilitates free fatty acid oxidation [84]. Meanwhile, in another study, ANGPTL4 was confirmed to function as a gatekeeper, inhibiting lipid profiles and protecting against intracellular lipid overload [85]. Contrastingly, the mice deficient in the ANGPTL4 gene weighed more than the control mice [85].
By analyzing data from investigations in humans, obese children presented significantly lower serum ANGPTL4 concentrations than children with clinically normal weights. Furthermore, the main measurements relating to overweight–obesity classifications, including body mass index (BMI), waist circumference, and weight, were all shown to be inversely correlated with serum ANGPTL4 concentrations. The individuals undergoing weight loss presented a negative relationship between serum ANGPTL4 concentrations and total cholesterol serum concentrations, indicating that ANGPTL4 could synchronously modulate the risk of obesity and hypercholesterolemia [86]. However, another study reported that the serum concentrations of ANGPTL3, ANGPTL4, and ANGPTL8 were significantly upregulated in patients with diagnosed obesity [87]. Moreover, the serum ANGPTL8 concentrations were positively associated with developing obesity and dyslipidemia, characterized by elevated serum concentrations of TGs and LDL-C alongside the serum HDL-C levels [88]. Consistently, REGN3776 injections, an ANGPTL8 monoclonal antibody, reduced serum TG levels by stimulating LPL activity. Furthermore, an exogenous supplement of ANGPTL8 presented additional effects on body fat concentrations and body weight in animal experiments [89]. Similarly, the mice with ANGPTL8 gene deficiencies displayed reduced body weight, potentially through abolishing TG accumulation in adipocytes and increasing circulating TG clearance. These results strongly propose that serum ANGPTL8 could be a novel therapeutic target for hypertriglyceridemia and cardiometabolic disorders [90].
Ectopic lipid storage can induce the pathogenic mechanism of atherosclerotic-related cardiovascular diseases. Recently, it has been shown that the proinflammatory response occurring within vessel walls could also facilitate the development of atherosclerosis [91]. The important role of diverse ANGPTLs in regulating atherosclerotic-related cardiovascular diseases has begun to gain appreciation since ANGTPLs play a vital role in regulating serum lipid profiles and the process of inflammation.
Emerging evidence has revealed the essential role of ANGPTL proteins in the development of atherosclerotic-related cardiovascular diseases [92]. For instance, ANGPTL4 has been shown to modulate the metabolism of LDL particles and influence the formation of foam cells. Two independent research demonstrated that mice with an ANGPTL4 gene deficiency presented inhibited atherosclerotic progression [92, 93]. Following in-depth research, it was proposed that ANGTPL4 modulated the development of atherosclerotic lesions via, at least partly, lowering serum TG concentrations mediated by LPL and inhibiting foam cell formation [93]. Furthermore, the authors also demonstrated that ANGPTL4 gene silencing significantly inhibits foam cell formation [93]. Consistent with this notion, the potential mechanism whereby ANGPTL4 protected atherosclerotic development could be due to reduced oxidized low-density lipoprotein (ox-LDL) uptake by macrophages. In addition, ANGPTL4 has further been confirmed to reduce lipid accumulation in macrophages, indicating that ANGPTL4 could improve the development of atherosclerosis by inhibiting the foam cells formation [94]. Recent research has provided evidence that ANGPTL4 could inhibit serum LPL activity and concentrations, simultaneously promoting low-density lipoprotein receptor-related protein 1 (LRP1)/LPL complex formation, which results in increased lipid uptake into macrophages, mediated by LPL [95]. Conversely, decreased ANGPTL3 gene expression has been shown to improve the development of dyslipidemia and cardiovascular diseases in ApoE gene-deficient mouse models via upregulating the clearance of isolated TRLs and ANGPTL4 [96].
Given that diverse ANGPTLs could modulate the pathological progression of the inflammatory response, research focuses are shifting toward exploring the underlying mechanisms. As reported, serum concentrations of ANGPTL3 and ANGPTL4 were significantly related to the progression of the inflammatory response; furthermore, by promoting inflammation-induced hepatic impairment and glycol metabolism, ANGTPL3 and ANGPTL4 could cause pathological alterations in atherosclerotic-related cardiovascular diseases [88].
Multiple studies have focused on the effect of ANGPTL4 on modulating serum or intracellular glucose metabolism. According to the published results, ANGPTL4 gene silencing in adipocytes could improve glucose tolerance, indicating that ANGPTL4 could influence glucose metabolism and the prevalence of diabetes mellitus. ANGPTL4 could also prevent excessive lipid accumulation within hepatocytes and myocytes via this mechanism, reducing the inflammatory response or atherosclerotic-related cardiovascular diseases [94]. Moreover, an independent study focused on patients with obstructive sleep apnea syndrome and found that these patients presented upregulated ANGPTL4 gene expression and inactivated LPL, which subsequently modulate the risk of atherosclerotic-related cardiovascular diseases [97]. In addition to the ANGPTL4-mediated effect in modulating atherosclerosis, ANGPTL4 was also demonstrated to influence endothelial junction stabilization through the vascular endothelial growth factor receptor 2 (VEGFR2) signaling pathway. Consequently, exogenous supplementation of the recombinant ANGPTL4 protein could inhibit the progression of the atherosclerotic lesion area in mouse and rabbit models [97].
In conclusion, the results summarized above strongly suggest that ANGPTL4 is a vital modulator of atherosclerotic development by inhibiting LPL.
Both aberrant lipid metabolism and proinflammatory processes are involved in the
pathogenic progression of multiple hepatic diseases, such as NAFLD- and hepatitis
C virus (HCV)-related liver diseases [98]. For instance, the ANGPTL3
gene expression levels were reduced by inhibiting the HNF-1
On the other hand, the serum concentrations of ANGPTL3 and cytokeratin 18, a proinflammatory response biomarker, were upregulated in the patients with NAFLD. Consequently, ANGPTL3 was used to improve the assessment of non-alcoholic steatohepatitis (NASH) and hepatic inflammatory developmental progression [99]. Similar to the alterations in serum ANGPTL3, subjects with hepatic steatosis presented lower concentrations of ANGPTL4. In contrast, the LPL activity increased compared to control individuals. Via this mechanism, ANPGTL4 could further modulate the storage of free fatty acids in hepatocytes [29]. More recently, the authors demonstrated using the antisense oligonucleotide of ANGPTL8 that the mice with NAFLD presented enhanced post-prandial TG uptake in white adipocytes. As a result, these mice also presented improved NAFLD.
Since the function of diverse ANGPTLs in modulating dyslipidemia and cardiovascular diseases has been well demonstrated, the functions of ANGPTL3, ANGPTL4, and ANGPTL8 in affecting other metabolic diseases, such as diabetes mellitus, ketosis, and renal dysfunction have also been given substantial attention in recent years.
ANGPTLs were demonstrated to modulate serum glucose metabolism and the prevalence of diabetes. Specifically, under diabetes mellitus conditions, LPL would be recruited into the vascular lumen and transformed into an active dimeric type. Following exogenous ANGPTL4 supplementation, LPL could be converted into the inactive form and subsequently suppress TG hydrolysis, which might induce hypertriglyceridemia. Otherwise, in diabetic patients, the authors found that the phosphoinositide 3-kinase (PI3K) signaling pathway was vital for insulin to reduce the ANGPTL4 gene expression. Due to this effect, when insulin resistance occurred in diabetic patients, the inhibitory effect of insulin on the ANGPTL4 gene expression was disrupted [100]. Consistent with these findings, the serum ANGPTL4 concentrations were higher in patients with type 2 diabetes mellitus than in healthy control individuals, proposing that ANGPTL4 could play an important role in modulating diabetic development [78]. These results suggest that the association between hypertriglyceridemia and the development of diabetes mellitus could be due to the increased ANGPTL4 concentrations, which inhibit LPL-mediated TG hydrolysis and enhance hepatic TG secretion [78]. Additionally, the subjects who possessed ANGPTL4 loss-of-function gene mutations presented reduced glucose concentrations and increased insulin sensitivity under the fasting condition compared with healthy volunteers. Emerging recent evidence further verified that the ANGPTL4-induced upregulation of serum amino acids played an essential function in increasing serum glucagon concentrations, further promoting the development of diabetes mellitus [101].
It has also been demonstrated that two other members of the ANGPTL protein family, ANGPTL3 and ANGPTL8, play an important role in affecting serum glucose metabolism and the progression of diabetes mellitus. Notably, the patients who possessed ANGPTL3 loss-of-function gene mutations presented relatively lower serum concentrations of insulin and glucose; furthermore, these individuals had improved insulin sensitivity compared with the control individuals [102]. Conversely, the ANGPTL8 gene expression levels were increased significantly in overweight/obese subjects, compared with those within the BMI-matched insulin-sensitive subjects, especially in subjects with NAFLD and diabetes. Consistently, the antisense oligonucleotide of ANGPTL8 was confirmed to play a protective function in modulating insulin resistance and glucose intolerance in mice fed a high-fat diet (HFD) by improving adipocyte lipid uptake and inhibiting lipid storage [103]. These above results indicate the negative relationship between the expressions of the ANGPTL3 and ANGPTL4 genes and the risk and development of diabetes mellitus and the ANGPTL8 gene.
Secondly, as is well described, strictly limiting daily food could induce extremely lower glucose concentrations; subsequently, ketogenesis, using excessive ketone body storage, could provide an alternative energy supplement. Concerning this notion, a study demonstrated that a naturally occurring flavonoid compound, named 1, 3, 5, 8-tetrahydroxyxanthone, targeted the ANGPTL3–LPL signaling pathway, which reduced the ANGPTL3 gene expression levels. Inversely, 1, 3, 5, 8-tetrahydroxyxanthone upregulated the LPL gene expression, which lowers the ketosis process in mouse models [104]. Using momilactone B as a novel inhibitor of ketosis, the authors found that this agent could modulate the process of ketosis by increasing the gene expression of signal transducer and activator of transcription-5b (STAT-5b) and indirectly negatively regulating ANGPTL3 gene expression [105]. Thus, downregulating the ANGPTL3 gene expression has been suggested as a novel target in treating ketosis.
Third, it was shown that patients with chronic renal dysfunction presented
higher serum concentrations of ANGPTL4 than healthy patients. In renal podocytes,
ANGPTL4 overexpression could lead to excessive proteinuria via the
podocyte-secreted ANGPTL4 under the status of nephritic syndrome [106]. Following
in-depth investigations, with the elevated ratio of free fatty acids to albumin
in serum, the extrarenal organs were found to release excessive ANGPTL4 in the
nephritic range [107]. Meanwhile, since the increased serum ANGPTL4
concentrations could promote the development of hypertriglyceridemia, it could be
considered a potential mechanism combining proteinuria and hypertriglyceridemia
as two biomarkers of chronic renal dysfunction [107]. Nevertheless, another
independent research study showed that the upregulated serum concentrations
related to the ANGPTL4 gene might induce acute kidney injury (AKI) due
to excessive TG storage through decreased LPL activity within renal cells. Given
that AKI could be improved by WY-14643, as a peroxisome proliferator-activated
receptor
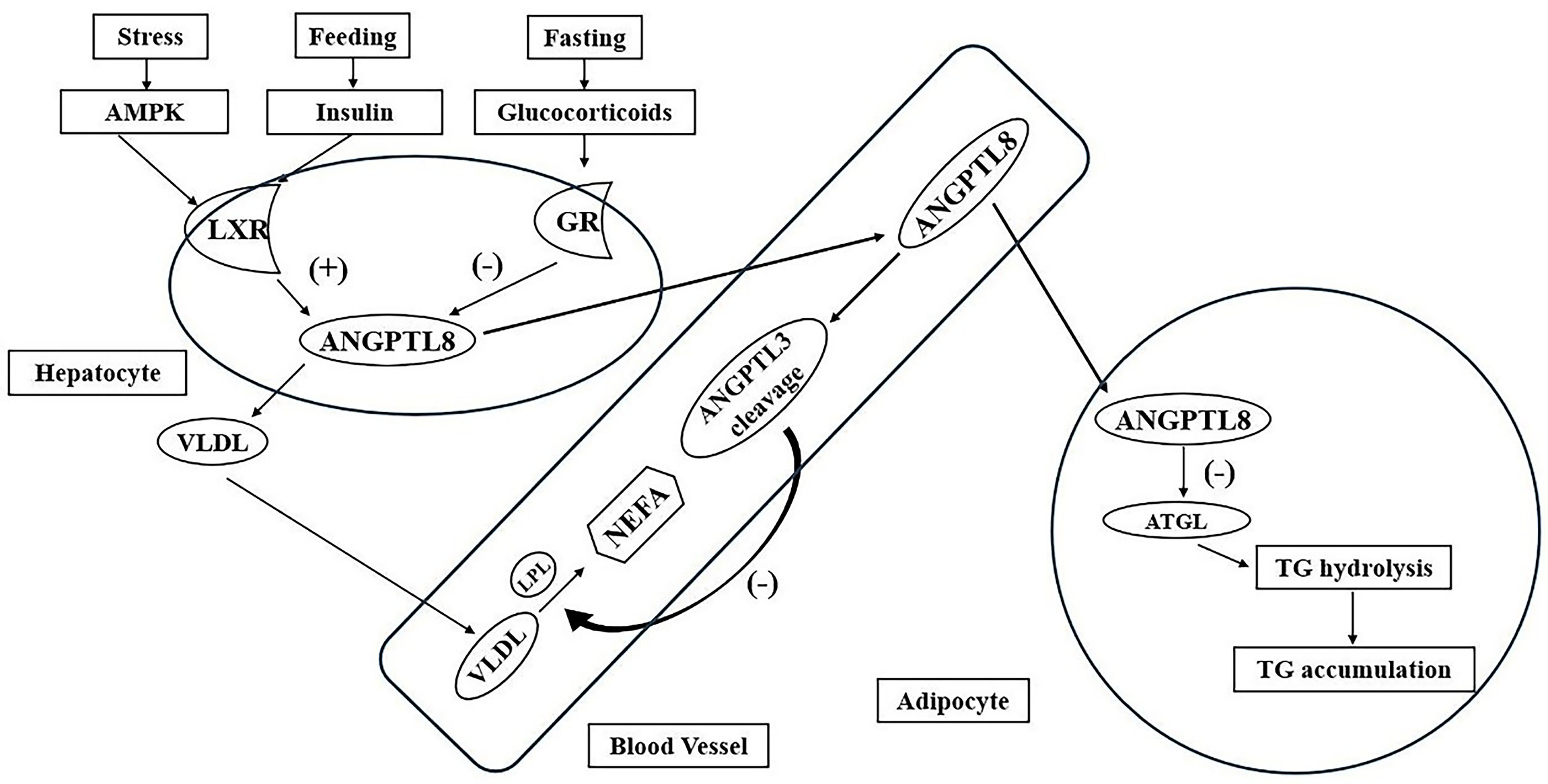 Fig. 2.
Fig. 2.
The roles of the diverse ANGPTL protein family in lipid metabolism. Abbreviation: AMPK, adenosine monophosphate-activated protein kinase; LXR, liver X receptor; GR, glucocorticoid receptor; ANGPTL8, angiopoietin-like protein 8; VLDL, very low-density lipoprotein; LPL, lipoprotein lipase; NEFA, non-esterified fatty acid; ANGPTL3, angiopoietin-like protein 3; ATGL, adipose triglyceride lipase; TG, triglyceride.
The diverse ANGPTL proteins responsible for body energy homeostasis are distributed in various metabolic-related tissues. Specifically, ANGPTL3, ANGPTL4, and ANGPTL8 are the most important members of the ANGPTL protein family, which play an important role in LPL-induced TG metabolism. In addition to the negative effect on modulating the biological activity of LPL, ANGPTL3 is a negative modulator of endothelial lipase, which induces the negative modulation of serum HDL-C and promotes the risk of dyslipidemia. On the other hand, ANGPTL4 has been confirmed to convert LPL enzymes from active dimers to inactive monomers and facilitate the process of LPL degradation, which influences TG metabolism. Recently, ANGPTL8 has been shown to not inhibit LPL activity by itself; however, when ANGPTL8 combines with ANGPTL3, it can suppress LPL activity, which might further modulate the serum TG concentration. On the contrary, ANGPTL3 stimulation is necessary for the ANGPTL8 metabolism.
The comprehensive functions of different ANGPTL proteins in modulating LPL activity in various tissues under fasting and feeding conditions could provide a potential mechanism for hypertriglyceridemia. Furthermore, the interaction between diverse members of the ANGPTL protein family might provide different effects on modulating serum TG metabolism. For instance, the interaction between ANGPTL4 and ANGPTL8 might lead to hypertriglyceridemia, obesity, or diabetes mellitus. As shown above, this review provides evidence that ANGPTLs are novel targets for treating hypertriglyceridemia and its related metabolic disorders in daily clinical practice.
In conclusion, further studies should focus on other ANGPTL proteins, including ANGPTL1, ANGPTL2, and ANGPTL5 to ANGPTL7, and their roles in modulating serum TG metabolism. These subsequent results might explain several controversies regarding the accurate function of different ANGPTLs in inhibiting LPL activity. Furthermore, these results could also stimulate future researchers to determine the potential roles of ANGPTL proteins as therapeutic strategies in dyslipidemia and its related cardiovascular diseases in daily clinical practice. Future studies might provide a much better understanding of the potential mechanisms through which different ANGPTL proteins modulate the risk of dyslipidemia in other species. It will be worth elucidating whether ANGPTLs can induce therapeutic strategies to reduce the prevalence of dyslipidemia-related diseases.
ANGPTLs, angiopoietin-like proteins; TG, triglyceride; GPIHBP1, glycosylphosphatidylinositol-anchored high-density lipoprotein binding protein 1; HNF-1, hepatocyte nuclear factor-1; LXR, Liver X receptor; RXR, LXR-retinoid X receptor; TLR, Toll-like receptor; HIF-1, hypoxia-inducible factor-1; GALNT2, N-acetyl-galactosaminyl transferase 2; LDL-C, LDL cholesterol; LRP1, LDLR-related protein 1.
SY, XXL, and YC contributed to the study design; SY, ML, and XS wrote the manuscript. XS and ML contributed to the literature collection and analysis. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This work was supported by grants from Natural Science Foundation of Xiamen, China (No. 3502Z20227295), the project of Fujian Natural Science Foundation (No. 2024J011429) and the Health Youth Research Project in Fujian Province (No. 2022QNB017).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.