





 , Bin Deng 1,2,3,4,†, Tian Sun 1,2,3,4, Yongqin Wang 1, Wei Dong 1, Junfei Jin 1,2,3,4,*
, Bin Deng 1,2,3,4,†, Tian Sun 1,2,3,4, Yongqin Wang 1, Wei Dong 1, Junfei Jin 1,2,3,4,* , Zhixiong Pan 1,2,3,4,*
, Zhixiong Pan 1,2,3,4,*1 Guangxi Key Laboratory of Molecular Medicine in Liver Injury and Repair, The First Affiliated Hospital of Guilin Medical University, 541001 Guilin, Guangxi, China
2 Guangxi Health Commission Key Laboratory of Basic Research in Sphingolipid Metabolism Related Diseases, The First Affiliated Hospital of Guilin Medical University, 541001 Guilin, Guangxi, China
3 China-USA Lipids in Health and Disease Research Center, Guilin Medical University, 541001 Guilin, Guangxi, China
4 Laboratory of Hepatobiliary and Pancreatic Surgery, The First Affiliated Hospital of Guilin Medical University, 541001 Guilin, Guangxi, China
†These authors contributed equally.
Abstract
Exosomes are specialized secreted vesicles for intercellular communication and signaling pathways as specialized secreted vesicles. Multiple studies have suggested the potential roles of hepatocyte-derived exosomes as biomarkers of liver injury and facilitators of hepatocyte proliferation and liver regeneration.
By utilizing murine models of hepatic ischemia-reperfusion injury (IRI), we examined the impact of hepatocyte-derived exosomes on mitigating hepatic IRI.
Our experiments have demonstrated that significantly lower levels of alanine transaminase, aspartate transaminase, and lactate dehydrogenase in mice treated with hepatocyte-derived exosomes compared with mice treated with phosphate-buffered saline (PBS). Furthermore, hepatocyte-derived exosomes inhibited hepatocyte apoptosis, reduced levels of inflammatory cytokines, and suppressed the entry of inflammatory cells into the liver following hepatic IRI. Complement 3d (C3d) expression showed a notable decrease in exosome-treated mice compared with PBS-treated mice, suggesting that hepatocyte-derived exosomes effectively inhibited complement activation during hepatic IRI. Blocking the fusion of exosomes with cells using Annexin V weakened the protective effects of the exosomes against hepatic IRI.
Our findings highlight the ability of hepatocyte-derived exosomes to mitigate hepatic IRI by inhibiting complement activation. These results reveal a novel role for exosomes in blocking complement activation, suggesting a potential new therapeutic avenue for preventing hepatic IRI.
Keywords
- exosomes
- hepatic ischemia reperfusion injury
- complement
- apoptosis
Liver ischemic-reperfusion (IR) injury (IRI) may occur during liver surgery, such as partial resection and transplantation [1, 2, 3]. IRI has been appreciated as a critical risk factor related to poor postoperative outcomes [4]. Given the technical aspects of these surgical procedures, dealing with liver IRI is a common challenge [5, 6]. Understanding the pathogenesis of hepatic IRI may inform potential strategies for improving clinical outcomes after liver surgery [7, 8]. During hepatic IR, a complex interaction between endoplasmic reticulum stress, oxidative stress, and inflammation takes place, resulting in a significant loss of liver cells during reperfusion and ultimately leading to hepatic IRI [9, 10, 11, 12]. Dying liver cells release damage-associated molecular patterns (DAMPs) during liver IRI [13, 14], which when bound to pattern-recognition receptors (PRRs) trigger innate immune cells and promote inflammatory responses [3, 13].
Exosomes are small extracellular vesicles surrounded by a lipid bilayer, with a diameter typically in the range between 30 and 150 nm [15, 16]. They contain different parts of the cell, including DNA, RNA, lipids, metabolites, and proteins found in the cytoplasm and on the cell surface. Extracellular matrix receives exosomes when the multivesicular body (MVB) merges with the cell membrane. Playing a key role in cellular signaling, exosomes primarily facilitate communication [17, 18]. Recent research has focused on exploring the potential role of exosome components and their regulation of cell activity as intercellular communication in receptor cells [19, 20]. For example, it has been shown that miRNAs from tumor exosomes lacking p53 communicate with sensory nerves, leading to their transformation into adrenergic nerve types that invade the tumor [21]. Additionally, exosomes containing programmed cell death ligand 1 (PD-L1) promote the growth of malignant tumor cells by modulating immune-cell polarization and inducing immune suppression in the tumor microenvironment [22]. Exosomes have also been demonstrated to facilitate tissue repair after pulmonary fibrosis-induced lung injury [23]. Furthermore, recent research underscores the importance of exosomes in tissue repair, regulation of signal transduction, and dialogue between renal tubules and macrophages in diabetic nephropathy [24]. Dll4-containing exosomes have been identified as regulators of renal tubule-macrophage dialogue in diabetic nephropathy [25]. Hepatocyte-derived exosomes have shown potential as regulators of liver injury [26, 27]. It has also been shown that exosomes play a crucial role in decreasing the quantities of immunosuppressive mRNA and immunostimulation DAMPs in the overall circulation [28]. In summary, exosomes perform a vital role in maintaining immune balance under stressful conditions.
In this study, we focused on assessing the role of hepatocyte-derived exosomes in hepatic IR. According to our results, exosomes released from hepatocytes reduce complement activation and protect against hepatic IRI. The results of this study suggest that hepatocyte exosomes may be considered for preventing hepatic IRI.
The SLAC Laboratory (HUNAN, CHINA) provided adult male C57BL/6 mice. The mice were maintained in a specific pathogen–free environment (temperature: 20–26 °C; relative humidity: 50%–60%; air changes: 10–20 times/hour) with a 12-hour/12-hour light–dark cycle. The mice utilized in the study were about 8 weeks old and weighed between 20 and 24 g. All animal experiments were conducted in line with the guidelines for the animal care and use of the Guilin Medical University Animal Ethics, ensuring minimal animal usage and reducing discomfort (GLMC-IACUC-2023025).
The mice were randomly allocated to receive hepatic IR or sham surgery. Following anesthesia with sodium pentobarbital (50 mg/kg, P3761, Sigma-Aldrich) and positioning in a supine manner, a midline laparotomy was performed to unveil the livers. The portal vein and hepatic artery, which supply blood to the liver, were bluntly isolated. A noninvasive microvascular clamp was promptly used to occlude the portal vein and hepatic artery so as to stop the blood flow. The sham group received identical anesthesia and midline laparotomy without vascular blockage. After 30 minutes of liver ischemia, the clamps were removed to start reperfusion. Exogenous Exo (200 µg per mouse), Annexin Ⅴ (A5, 230-30121, Sigma, NY, USA) (20 µg per mouse), and Exo + A5 mixture were injected intravenously. After a reperfusion period of 6 hours, the mice were anesthetized via intraperitoneal injection of sodium pentobarbital (50 mg/kg, P3761, Sigma-Aldrich), subsequently blood collection and cervical dislocation. Following the surgical procedure, liver tissues and serum samples were collected for further analysis.
The AML12 cell line was cultured in DMEM/F12 medium (C11330500BT, Gibco, Thermo
Fisher, Paisley, UK) containing 3.2 g/L of glucose, 5 mg/mL of
Insulin–Transferrin–Selenium-X (ITS, 13146, Sigma, NY, USA) each (Sigma-Aldrich), 40
ng/mL of dexamethasone (D8040, Solarbio, LIFE SCIENCES, Beijing, China), and 10% Exo-FBS (Bovine
Exosome-depleted FBS, 50A-1, System Biosciences, Shanghai, China). Exosomes were obtained from the
medium that has cultured AML12 cells using the exoEasy Maxi Kit (76064, QIAGEN, , Shanghai, China) (Supplementary Table 1)
from QIAGEN in accordance with the provided instructions. The cell culture
supernatants were filtered to exclude particles larger than 0.8
The size of exosomes was measured using an H-7650 transmission electron microscope (HT-7700, Hitachi, Japan), and exosome purity was evaluated by ZetaView PMX 110 (8.04.02 SP2, Meerbusch, Germany). The presence of surface markers on exosomes, including Tsg101, CD9, and CD63, was verified using western blot (Supplementary Fig. 1).
A 30-minute period of ischemia was followed by a 6-hour reperfusion period. Exosomes (Exo)-treated mice received exosomes (200 µg per mouse) intravenously before reperfusion. The corresponding control group received A5 (20 µg per mouse) intravenously. Additionally, exosomes were incubated with A5 for 30 minutes, and the mixture was intravenously administered to Exo + A5–treated mice.
For histopathological analysis, a sample of the liver was taken at the time of
euthanasia and placed in a solution containing 10% formalin (F5554,
Sigma-Aldrich, Milwaukee, USA). The fixed tissues were then embedded in paraffin, deparaffinized
with xylene, rehydrated with an ethanol series, and subsequently stained with
hematoxylin and eosin (H&E). Utilizing a fluorescent microscope (DM4, Leica, Wetzlar,
Germany), images were acquired with consistent acquisition settings. The extent
of necrosis was examined in five random fields on each slide under 40
The TdT-mediated dUTP nick end labeling (TUNEL) kit (11684817910, Roche, Basel, Switzerland) was utilized to assess cellular apoptosis in the liver tissue slices in accordance with the provided guidelines. The sections were initially dewaxed, hydrated, and subjected to protease K treatment for protein digestion. Subsequently, they were exposed to TUNEL reaction mixtures under humidified conditions at 37 °C for 1 hour in a light-free environment. Subsequently, DAPI staining (1:1000; Invitrogen, CA, USA) was carried out for 3 minutes at room temperature. The visualization and observation of TUNEL-positive cells was conducted with a fluorescence microscope (DM4, Leica, Wetzlar, Germany).
For immunohistochemical staining, the liver specimens were first dewaxed and then incubated with 1% hydrogen peroxide (H2O2, 230314, CNPP, Shanghai, China) for 20 minutes to inhibit endogenous peroxidase. Next, they were blocked with 5% bovine serum albumin (BSA, A8020, Solarbio, Beijing, China) at room temperature for 1 hour, followed by overnight incubation at 4 °C with primary antibodies against cluster of differentiation 11b (CD11b) (Cat No. Ab133357; 1:4000, Abcam, Cambridge, MA, USA), lymphocyte antigen 6 complex locus G6D (Ly6G) (Cat No. Ab238132; 1:2000, Abcam), and C3d (Cat No. AF2655-SP; 3 µg/mL, R&D Systems, Minneapolis, MN, USA). The sections were then exposed to secondary antibodies for one hour at 37 °C and underwent 3,3′-diaminobenzidine (DAB; 8059S, Zhongshan Golden Bridge) staining. Hematoxylin was used as a counterstain, and examination of the stained samples was performed using a light microscope (DM4, Leica, Wetzlar, Germany).
Statistical analysis was performed using GraphPad Prism 5.0 (9.0.1, GraphPad Software, CA, USA). Differences
among the groups were assessed through one-way analysis of variance (ANOVA).
Next, pairwise inter-group comparisons were based on the t-test or
Tukey’s post hoc test. All data were reported as mean
To investigate the potential involvement of hepatocyte-derived exosomes in hepatic IRI, intravenous injection of hepatocyte-derived exosomes was administered before reperfusion. After 6 hours of reperfusion, signs of severe liver damage were observed, including increased infiltration by inflammatory cells, hepatocyte necrosis, and elevated levels of serum alanine aminotransferase (ALT) (Fig. 1C), aspartate aminotransferase (AST) (Fig. 1D), and lactate dehydrogenase (LDH) (Fig. 1E) in the phosphate-buffered saline (PBS)-treated mice compared to the mice without IR. In contrast, treatment with exosomes significantly mitigated liver damage, as evidenced by reduced necrotic area, inflammatory cells, and serum ALT, AST, and LDH levels. A5, which has been shown to inhibit the fusion of exosomes with target cells, was used in this study to block the interaction between exosomes and target cells. While A5 alone did not affect hepatic IRI, it diminished the protective effect of exosomes against hepatic IRI (Fig. 1A–E). These findings suggest that hepatocyte-derived exosomes may confer protection against hepatic IRI through their interaction with target cells.
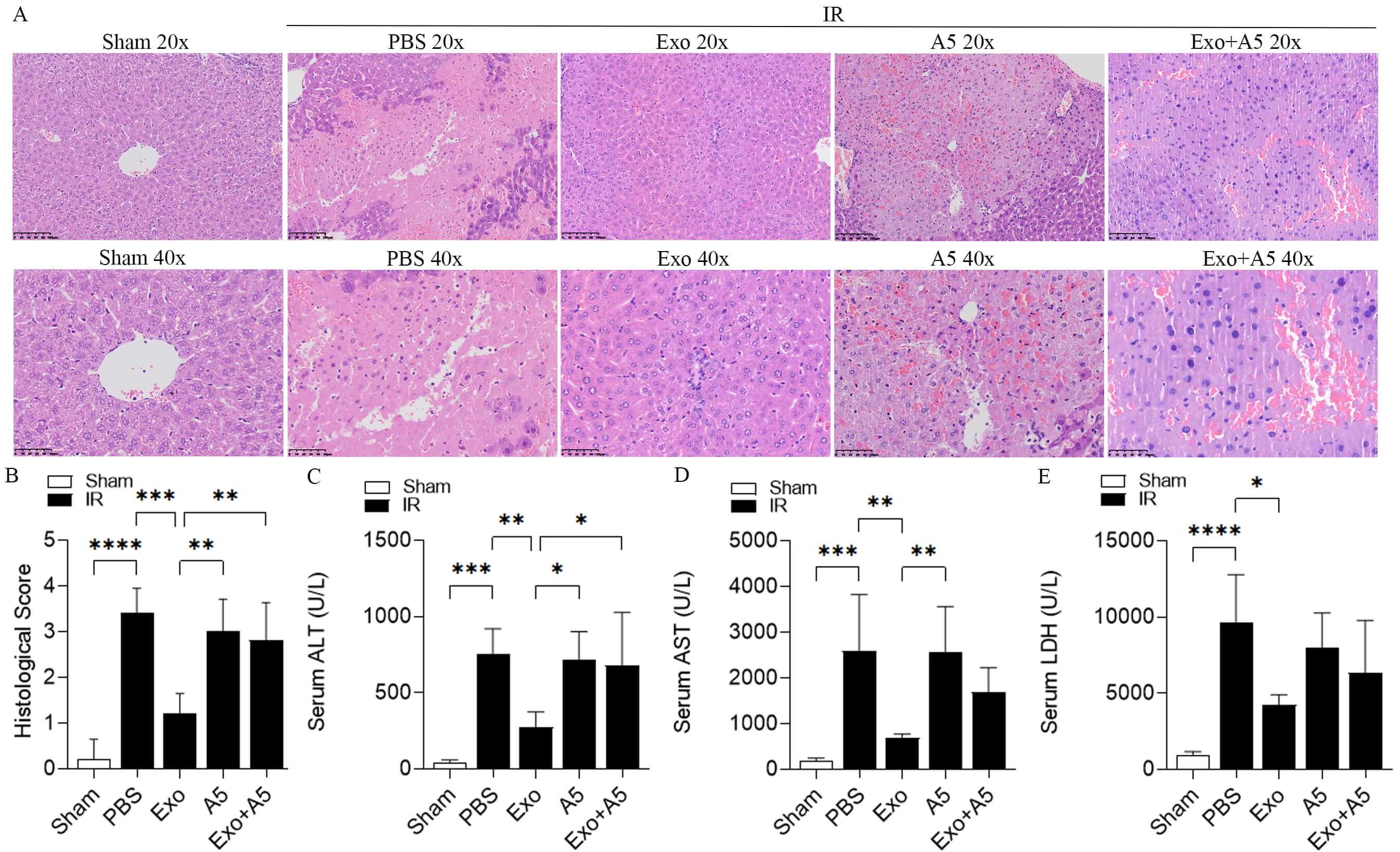 Fig. 1.
Fig. 1.
Hepatocyte-derived exosomes ameliorate hepatic IRI. In the
C57BL/6 mice treated with exosomes (Exo) and the appropriate control group, H&E
staining (A), histological score (B), and levels of ALT (C), AST (D), and LDH (E)
were measured after 30-minute hepatic ischemia followed by 6-hour reperfusion.
The Exo-treated mice were injected intravenously with exosomes (200 µg per
mouse) before reperfusion. The control group was injected intravenously with
Annexin V (A5, 20 µg per mouse). Additionally, exosomes were incubated with
A5 for 30 minutes, and the mixture was intravenously administered to the Exo +
A5–treated mice. The 20× scale bar indicates 100 µm. The 40× scale bar
indicates 50 µm. Data are displayed as mean
Hepatocyte cell death is the initial event leading to liver injury after hepatic IR [29, 30, 31]. To evaluate hepatocyte apoptosis, TUNEL staining was performed. As expected, there was a notable rise in the number of TUNEL-positive cells in the liver of the mice subjected to hepatic IR compared to the control mice without IR. However, under IR conditions, the Compared to the mice treated with PBS, Exo-treated mice showed a significant reduction in the number of TUNEL-positive cells in the liver (Fig. 2A,B). Additionally, A5 alone did not alter the count of TUNEL-positive cells, but it did inhibit the effect of exosomes. In addition, cleaved caspase-3 protein expression showed consistent results, cleaved caspase-3 is significantly increased after IRI and significantly reduced in Exo-treated animals (Fig. 2C,D).
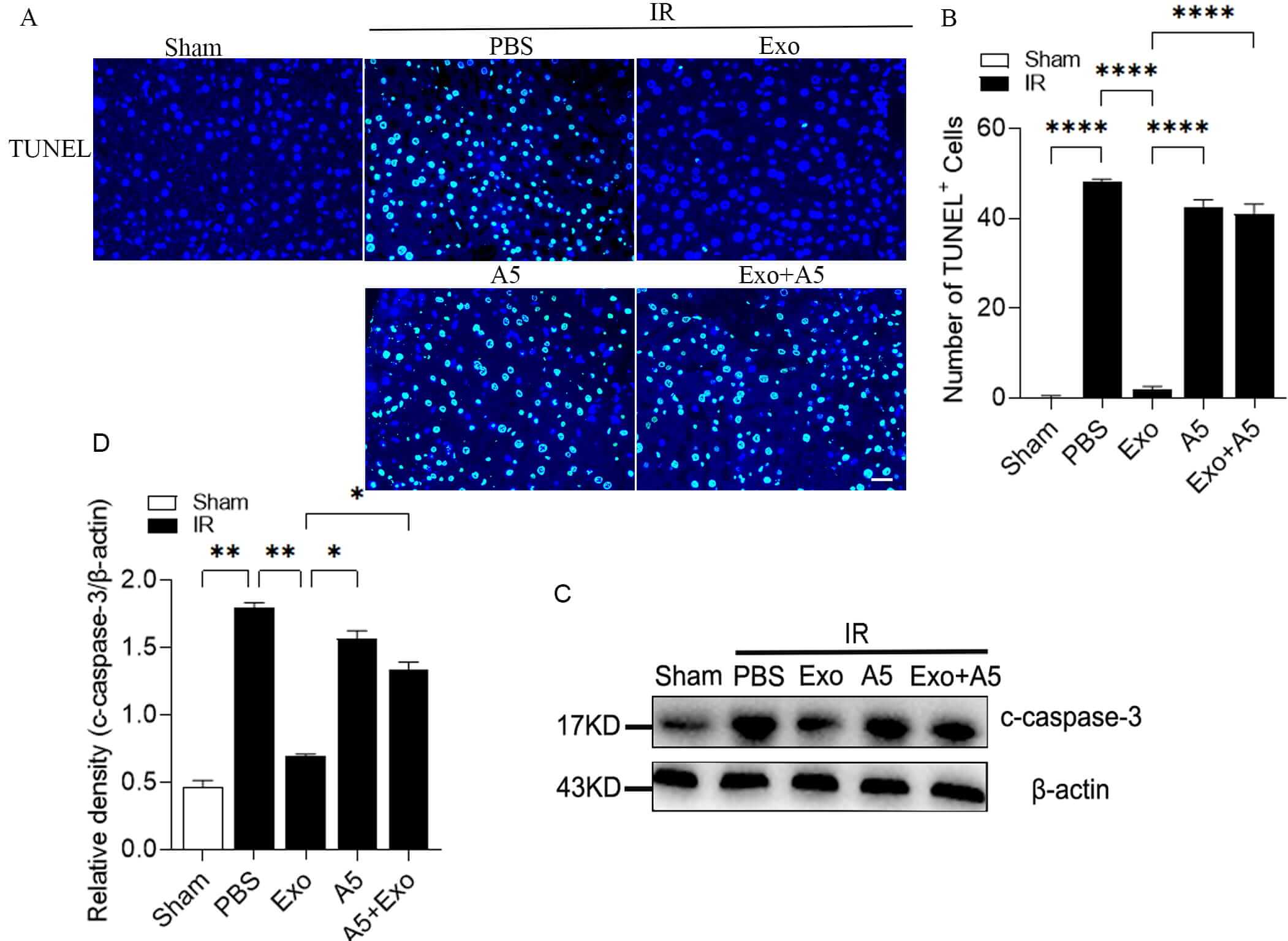 Fig. 2.
Fig. 2.
Hepatocyte-derived exosomes reduce hepatocyte apoptosis during
hepatic IRI. Analysis of TUNEL staining (A) and its quantification (B),
c-caspase-3 protein expression (C) and its quantification (D) by WB in the livers
of the mice treated with Exo and the corresponding control groups, exposed to 30
minutes of hepatic ischemia and 6 hours of reperfusion. The scale bars indicate
50 µm. Data are expressed as the mean
In light of the crucial role of inflammation in hepatic IRI, as well as the
potential therapeutic impact of exosomes on hepatic IRI [32], we next
investigated the relationship between exosomes and inflammation in this specific
context. Following the induction of hepatic IRI in the mice treated with PBS,
there was a noticeable increase in the levels of interleukin-6 (IL-6), tumor
necrosis factor
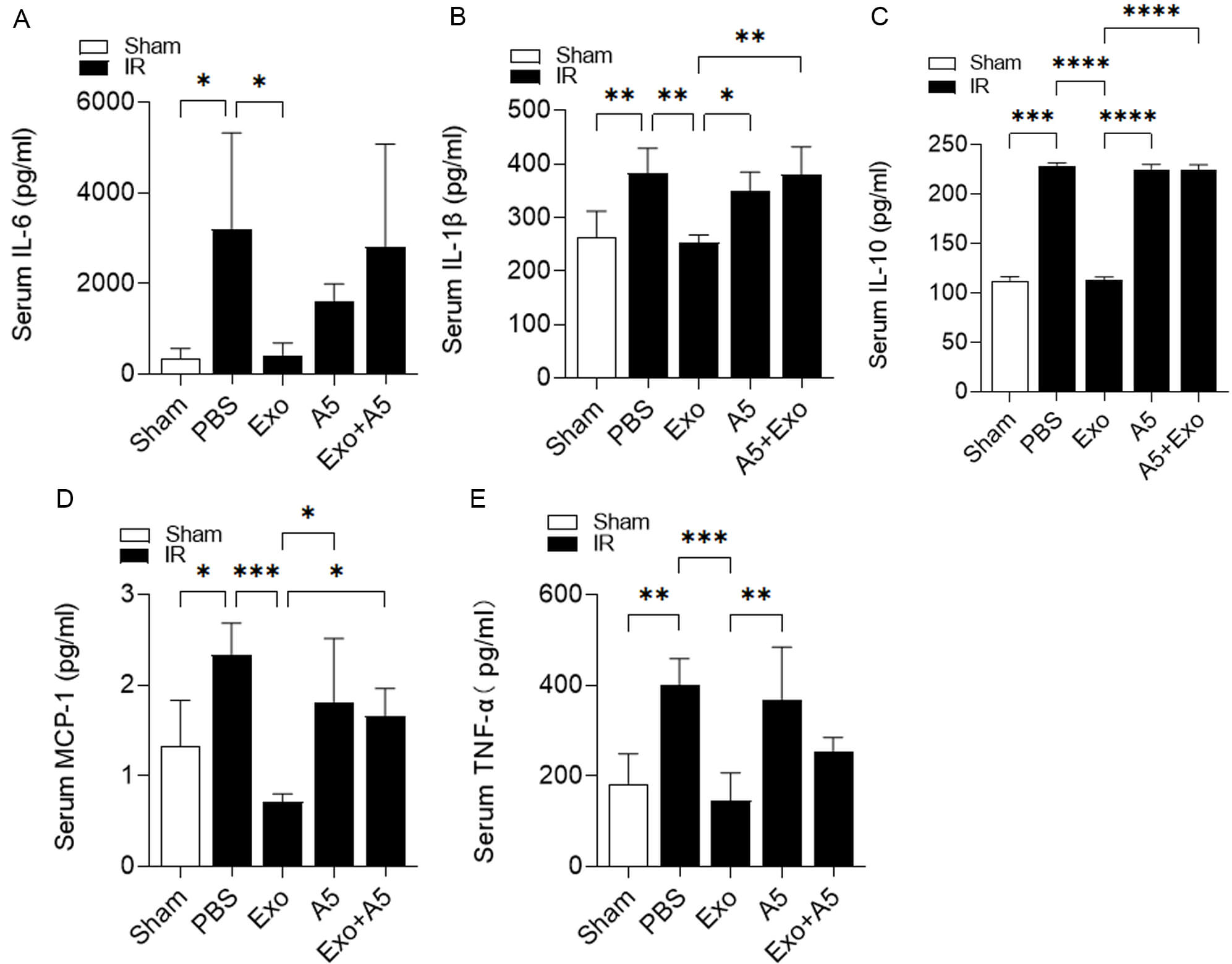 Fig. 3.
Fig. 3.
Hepatocyte-derived exosomes ameliorate inflammation during
hepatic IRI. Serum levels of IL-6 (A), IL-1
Consistent with the aforementioned Enzyme linked immunosorbent assay (ELISA) results, the immunohistochemical analysis revealed an increase in Ly6G-positive cells and CD11b-positive inflammatory cells in the livers of the mice treated with PBS after hepatic IRI compared to the control group without IR (Fig. 4A,B). In contrast, there was a significant decrease in inflammatory cell count in the livers of the mice receiving Exo treatment during the hepatic IR challenge (Fig. 4A,B). Additionally, although A5 alone did not influence the infiltration by inflammatory cells, there was a notable rise in the inflammatory cell population in the livers of the mice treated with Exo + A5 following hepatic IRI (Fig. 4A,B).
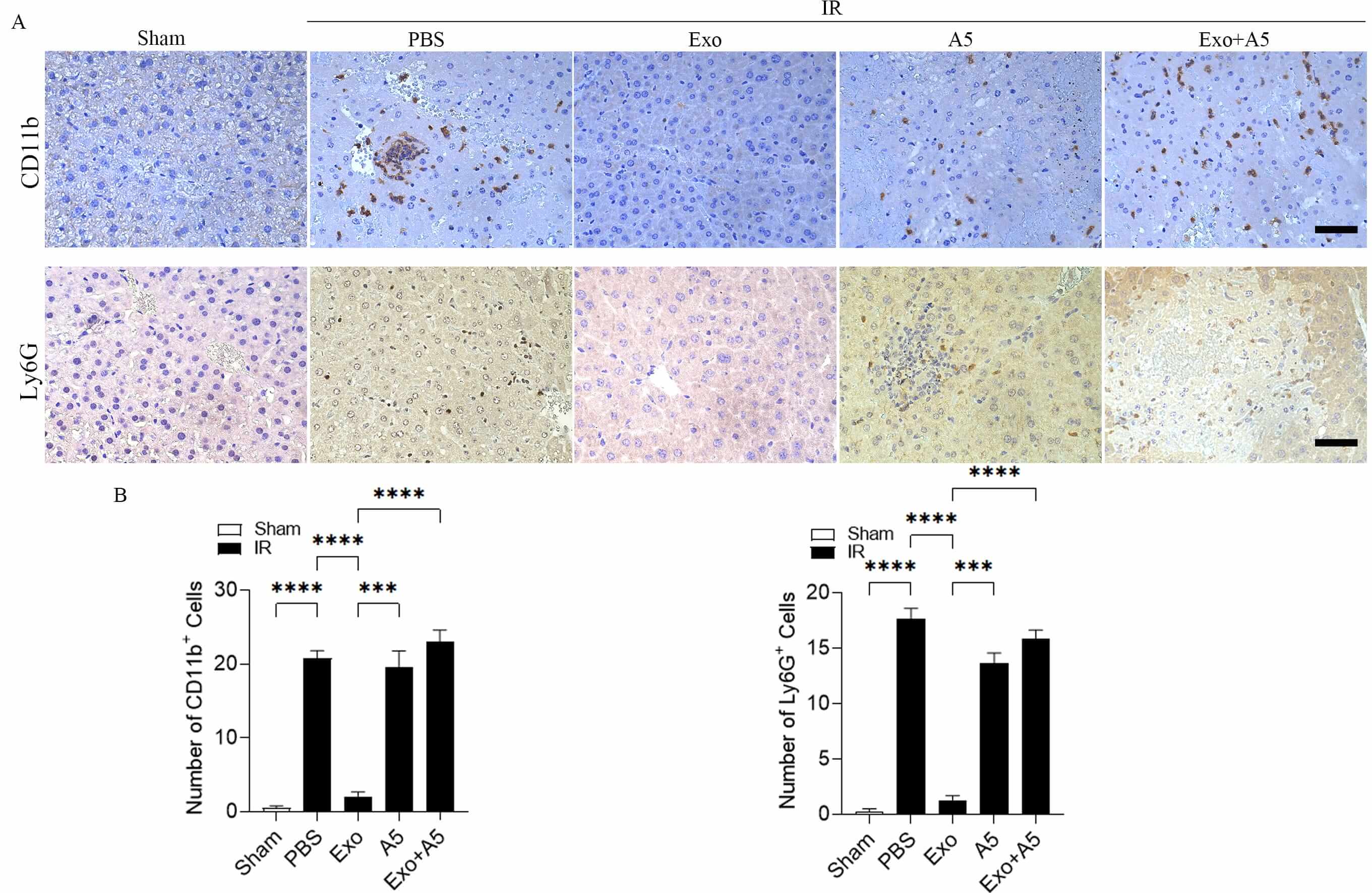 Fig. 4.
Fig. 4.
Hepatocyte-derived exosomes suppress the inflammatory cell of
inflammatory cells in hepatic IRI. High-quality immunohistochemical staining (A)
and evaluation (B) of CD11b-positive and Ly6G-positive cells infiltrating the
livers of the mice treated with Exo and the control groups subjected to 30
minutes of hepatic ischemia and 6 hours of reperfusion. The scale bars denote 50
µm. Data are expressed as mean
After hepatic IR, the systemic immune response activates the complement cascade, leading to the occurrence of hepatic IRI. Consequently, we examined whether exosomes from the liver cells could decrease liver IRI by blocking the activation of the complement system. It is well known that C3d deposition is a marker of complement activation. C3d deposition was significantly enhanced after hepatic IRI in the PBS-treated mice compared to the sham-treated mice. C3d deposition was remarkably reduced in the livers of the Exo-treated mice compared to the PBS-treated mice (Fig. 5A,B). As expected, A5 alone had no effect on C3d deposition; however, it significantly increased C3d deposition in the livers of the Exo-treated mice that underwent hepatic IRI.
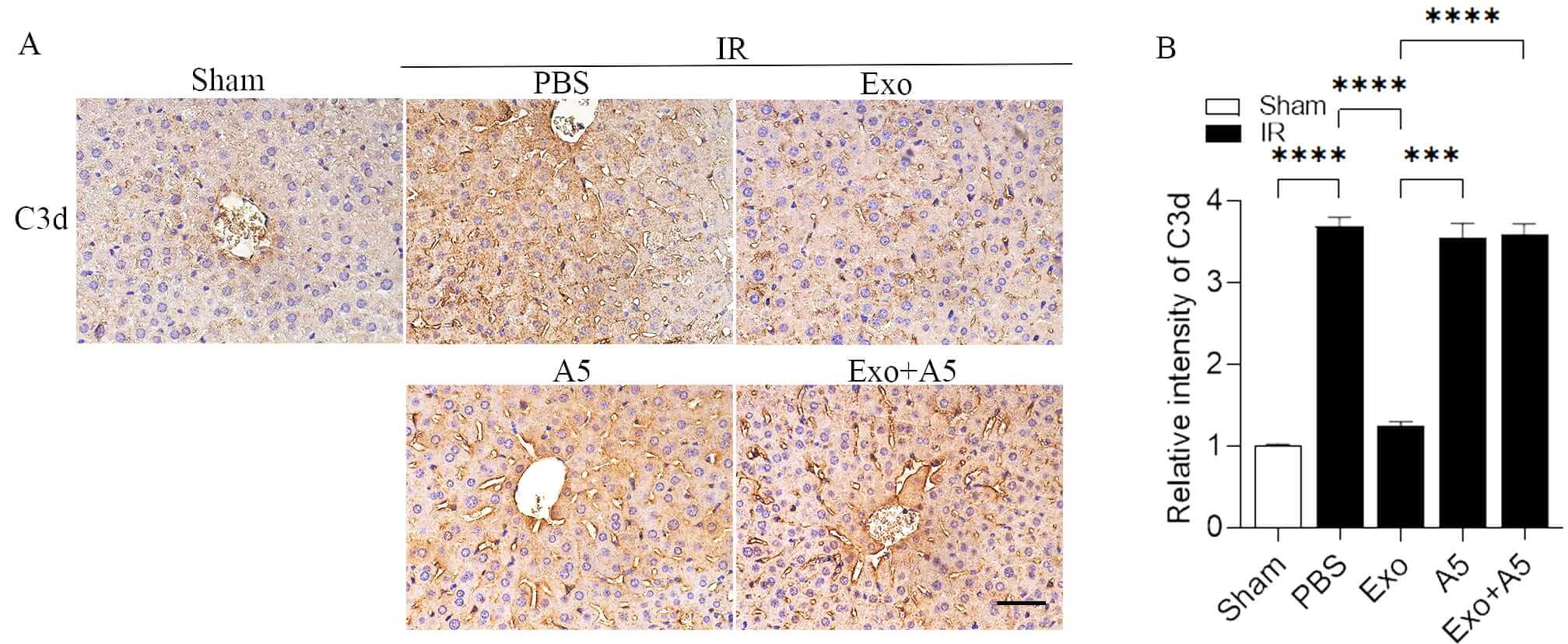 Fig. 5.
Fig. 5.
Hepatocyte-derived exosomes inhibit complement activation during
hepatic IRI. Impeccable immunohistochemical staining (A) and measurement (B)
showing the existence of C3d deposition invading the livers of the mice treated
with Exo and the control groups after 30 minutes of hepatic ischemia and 6 hours
of reperfusion. The scale bars denote 50 µm. Data are displayed as mean
Taken together, these findings indicate that hepatocyte-derived exosomes may reduce liver injury caused by the lack of blood flow and subsequent reperfusion by blocking the activation of complement, which in turn decreases inflammation and cell apoptosis.
The current study revealed an important role of hepatocyte-derived exosomes in reducing hepatic IRI and demonstrated that the underlying mechanism involves the prevention of complement activation. Our findings improve the understanding of the influence of hepatocyte-derived exosomes and indicate that these exosomes may protect against different types of liver damage. Additionally, our results suggest that these exosomes could offer a new strategy for treating liver injury.
A previous study demonstrated an increase in circulating exosome levels
following hepatic IRI [27]. In the present study, we illustrated the crucial
involvement of hepatocyte-derived exosomes in ameliorating hepatic IRI. Under
hepatic IR, the mice treated with hepatocyte-derived exosomes showed less liver
damage than those treated with PBS. In addition, the mice treated with
hepatocyte-derived exosomes showed reduced serum levels of ALT, AST, and LDH, as
well as a significant reduction in TUNEL-positive cells. Furthermore, the
Exo-treated mice exhibited significant reductions in IL-6, IL-1
Previous research has shown that Annexin V inhibits phosphatidylserine on the external layer of exosomes [33]. The objective of this study was to examine how preincubation of exosomes with Annexin V could affect their ability to protect against hepatic IRI. Our results revealed significant suppression of the protective effects in exosomes preincubated with Annexin V. The mice treated with Annexin V alone did not exhibit notable enhancements in liver function, prevention of hepatocyte cell death, decrease in inflammatory cytokines, or reduction in inflammatory cell infiltration in the liver following hepatic IR compared with the mice treated with PBS. Yet, the mice that received exosomes preincubated with Annexin V showed higher levels of hepatocyte apoptosis, inflammatory cytokines, and inflammatory cell infiltration in the liver during hepatic IRI. Furthermore, Annexin V alone did not alter C3d deposition but significantly increased C3d deposition in the liver of the mice treated with exosomes during hepatic IRI. These findings collectively indicate that Annexin V alone does not affect hepatic IRI, but hinders the protective role of exosomes by blocking phosphatidylserine.
In this study, we also investigated whether hepatocyte-derived exosomes could
reduce hepatic IRI by inhibiting complement activation. Under hepatic IR
treatment, we observed a significant decrease in hepatic C3d deposition following
treatment with the exosomes compared with PBS, indicating the ability of these
exosomes to suppress complement activation. Previous studies have shown that
exosomes can modulate inflammation and immune responses in specific contexts
[28, 34]. Furthermore, exosomal miR-21-3p, miR-146a, and miR-146b have been shown
to inhibit the TLR/NF-
Hepatocyte-derived exosomes have the potential to alleviate hepatic IRI by suppressing complement activation. This discovery reveals a new function of exosomes in reducing liver damage and suggests exosomes as a potential treatment for various liver diseases.
Exo, exosomes; IRI, ischemia reperfusion injury; A5, Annexin Ⅴ; C3d, complement 3d; TUNEL, TdT-mediated dUTP nick end labeling; IL-6, Interleukin-6; IL-1
There are no data that are publicly available or shared. All the necessary data to assess the findings of the present study can be found within the paper. The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
Conceptualization: ZP (lead) and JJ (supporting). Data curation & Formal Analysis: ZY, BD, TS. Methodology & Investigation: ZY, BD, TS, and YW. Funding acquisition & Resources: JJ and ZP. Supervision: JJ, TS. Project administration: ZP, ZY, and YW. Software & Visualization: WD, BD. Validation & Wrote manuscript: ZP, ZY. Writing — review & editing: all. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All animal experiments were conducted in accordance with the institutional guidelines of the Guilin Medical University Animal Ethics Committee (approval number: GLMC-IACUC-2023025).
We thank Dr. Chong Zhang for technical assistance and helpful suggestions.
This present study was partially funded by the Natural Science Foundation of Guangxi (2020GXNSFDA238006), by the Special Fund of the Central Government Guiding Local Scientific and Technological Development by Guangxi Science and Technology Department (GuikeZY21195024), the third batch of Lijiang Scholar Award in Guilin (2022-5-07), the Science and Technology Planned Project in Guilin (20210102-1), the Medical High Level Talents Training Plan in Guangxi (G202002005), the Construction Fund of Guangxi Health Commission Key Laboratory of Basic Research in Sphingolipid Metabolism Related Diseases (the Affiliated Hospital of Guilin Medical University, ZJC2020005), and the Guangxi Medical and Health Key Discipline Construction Project (2021-8-4-3).
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBL46946.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.