

 , Maria Rita Assenza 2, Francesca Sciarra 3, Giorgia D’Addato 1, Francesca Gioia Klinger 1, Mary Anna Venneri 3, Andrea M. Isidori 3, Federica Campolo 1,3,4,*
, Maria Rita Assenza 2, Francesca Sciarra 3, Giorgia D’Addato 1, Francesca Gioia Klinger 1, Mary Anna Venneri 3, Andrea M. Isidori 3, Federica Campolo 1,3,4,*
1 Departmental Faculty of Medicine, Saint Camillus International University of Health Sciences, 00133 Rome, Italy
2 Innate Lymphoid Cells Unit, Bambino Gesù Children’s Hospital, IRCCS, 00146 Rome, Italy
3 Department of Experimental Medicine, Sapienza University of Rome, 00161 Rome, Italy
4 Department of Anatomy, Histology, Forensic Medicine and Orthopedics, Unit of Histology and Medical Embryology, Sapienza University of Rome, 00161 Rome, Italy
Abstract
Phosphodiesterases (PDEs) are a huge superfamily of enzymes that fine-tune the intracellular levels of cyclic nucleotides —cyclic adenosine monophosphate (cAMP) and cyclic guanosine monophosphate (cGMP)—thus playing a pivotal role in the control of many cellular processes. While traditionally studied in the context of cardiovascular and neurological systems, mounting evidences highlight a crucial involvement of PDEs in metabolic homeostasis. This review explores the expanding landscape of PDEs function beyond classical cyclic nucleotide degradation, focusing on their roles in glucose and lipid metabolism and their implications in metabolic disorders, including obesity, type 2 diabetes (T2DM), and metabolic syndrome (MetS). Starting from an overview of the PDE superfamily, this work deeply examines the compartmentalized actions of cAMP-dependent protein kinase A (PKA) and cGMP-dependent protein kinase G (PKG) signaling pathways in key metabolically active tissues integrating PDE activities across different organs and disease states to offer a holistic view of their metabolic relevance. Special attention is given to the therapeutic relevance of PDE inhibitors (PDEi), distinguishing between established applications and emerging strategies targeting specific PDE isoforms in metabolic disease contexts to underscore the evolving concept that PDEs act as dynamic regulators of metabolic signaling networks. Understanding their isoform-specific and tissue-specific actions could thus open new avenues for therapeutic intervention in complex metabolic disorders.
Keywords
- cyclic nucleotide phosphodiesterases
- phosphodiesterase inhibitors
- obesity
- diabetes mellitus
- metabolic syndrome
Metabolic disorders, including obesity, type 2 diabetes (T2DM), and metabolic syndrome (MetS), represent a growing global health burden with profound clinical, social, and economic implications [1]. According to recent epidemiological estimates, over 1 billion people worldwide are affected by overweight or obesity, and the prevalence of T2DM continues to rise, particularly in low and middle-income countries [2]. These conditions are not only closely linked to increased cardiovascular morbidity and mortality but also contribute to a wide range of comorbidities including hepatic steatosis and renal dysfunction [3].
At the core of these disorders lies a complex interplay of molecular and cellular mechanisms, with insulin resistance and chronic low-grade inflammation playing central roles in the pathogenesis of metabolic dysregulation [4]. Although lifestyle interventions such as caloric restriction and physical activity remain the cornerstone of prevention and treatment, their long-term efficacy is often limited by poor adherence and compensatory metabolic adaptations [5, 6]. Pharmacological agents such as metformin, Glucagon-Like Peptide-1 (GLP-1) receptor agonists, and thiazolidinediones have demonstrated benefits in improving glycemic control and insulin sensitivity, yet the need for safer and more durable therapies remains largely unmet [7, 8].
In recent years, cyclic nucleotides signaling pathways have emerged as critical regulators of metabolic homeostasis [9, 10]. The second messengers cyclic adenosine monophosphate (cAMP) and cyclic guanosine monophosphate (cGMP) govern a broad array of physiological processes, including mitochondrial biogenesis, lipolysis, insulin secretion, inflammation, vascular tone, and thermogenesis across metabolically active tissues such as adipose tissue, liver, skeletal muscle, and heart [11]. The intracellular levels of cAMP and cGMP are tightly controlled by phosphodiesterases, a superfamily of enzymes that hydrolyze these second messengers [12].
Among the phosphodiesterase (PDE) families, several isoforms, most notably PDE3, PDE4, and PDE5, have been identified as key modulators of metabolic signaling cascades [13, 14, 15]. These enzymes contribute to the spatial and temporal compartmentalization of cyclic nucleotide responses, providing tissue- and context-specific regulation of metabolic pathways. Pharmacological inhibition of PDEs offers a unique opportunity to selectively amplify beneficial cAMP- or cGMP-dependent signaling events. Notably, PDE5 inhibitors, historically used for the treatment of erectile dysfunction and pulmonary hypertension, have shown favorable effects on insulin sensitivity, endothelial function, and lipid metabolism in both preclinical and clinical studies [16, 17, 18, 19]. Similarly, PDE3 and PDE4 inhibitors have been investigated for their potential to modulate inflammation, lipolysis, and adipocyte function [20].
Finally, beyond their classical role in cyclic nucleotide degradation, both PDE3 and PDE4 have been shown to assemble into multiprotein macrocomplexes with A-kinase anchoring proteins (AKAPs), sarcoplasmic reticulum Ca2⁺-ATPase 2 (SERCA2), phospholamban (PLB), and the 5-hydroxytryptamine 4 (5-HT4) receptor, thereby modulating distinct signaling pathways in a tissue-specific manner [21, 22, 23].
In this context, targeting cyclic nucleotide signaling through PDE inhibition could represent a promising therapeutic avenue for the management of metabolic diseases. This review aims to explore the molecular mechanisms by which the cAMP-dependent protein kinase A (PKA) and cGMP-dependent protein kinase G (cGMP-PKG) pathways regulates glucose and lipid metabolism, assess the current clinical evidence supporting their use, and discuss emerging insights into isoform-specific and tissue-targeted pharmacological strategies.
Different databases, including PubMed, Web of Science, Scopus and Google Scholar, were used for the keywords of “Phosphodiesterases, Obesity, Type 2 Diabetes, Metabolic syndrome, Phosphodiesterases Inhibitors, Phosphodiesterases knockout mouse models”.
Inclusion criteria included all of the following keywords: “PDEs and obesity, PDEs and T2DM, PDEs and MetS, PDEi and metabolic disorders, PDEs knockout mouse models and metabolic disorders”.
Exclusion criteria were metabolic diseases apart from “obesity, T2DM and MetS”.
The cAMP–dependent PKA signaling cascade is a central regulatory pathway in cellular energy metabolism, integrating hormonal signals to coordinate glucose and lipid homeostasis across metabolically active tissues [24].
Signal initiation begins with the activation of a G protein-coupled receptor (GPCR), which stimulates adenylyl cyclase (AC) activity, catalyzing the conversion of adenosine triphosphate (ATP) into cAMP. Rapid increase in intracellular cAMP levels activates PKA, which phosphorylates a broad range of downstream effectors, thereby modulating lipolysis, gluconeogenesis, mitochondrial function, and thermogenesis [25] (Fig. 1).
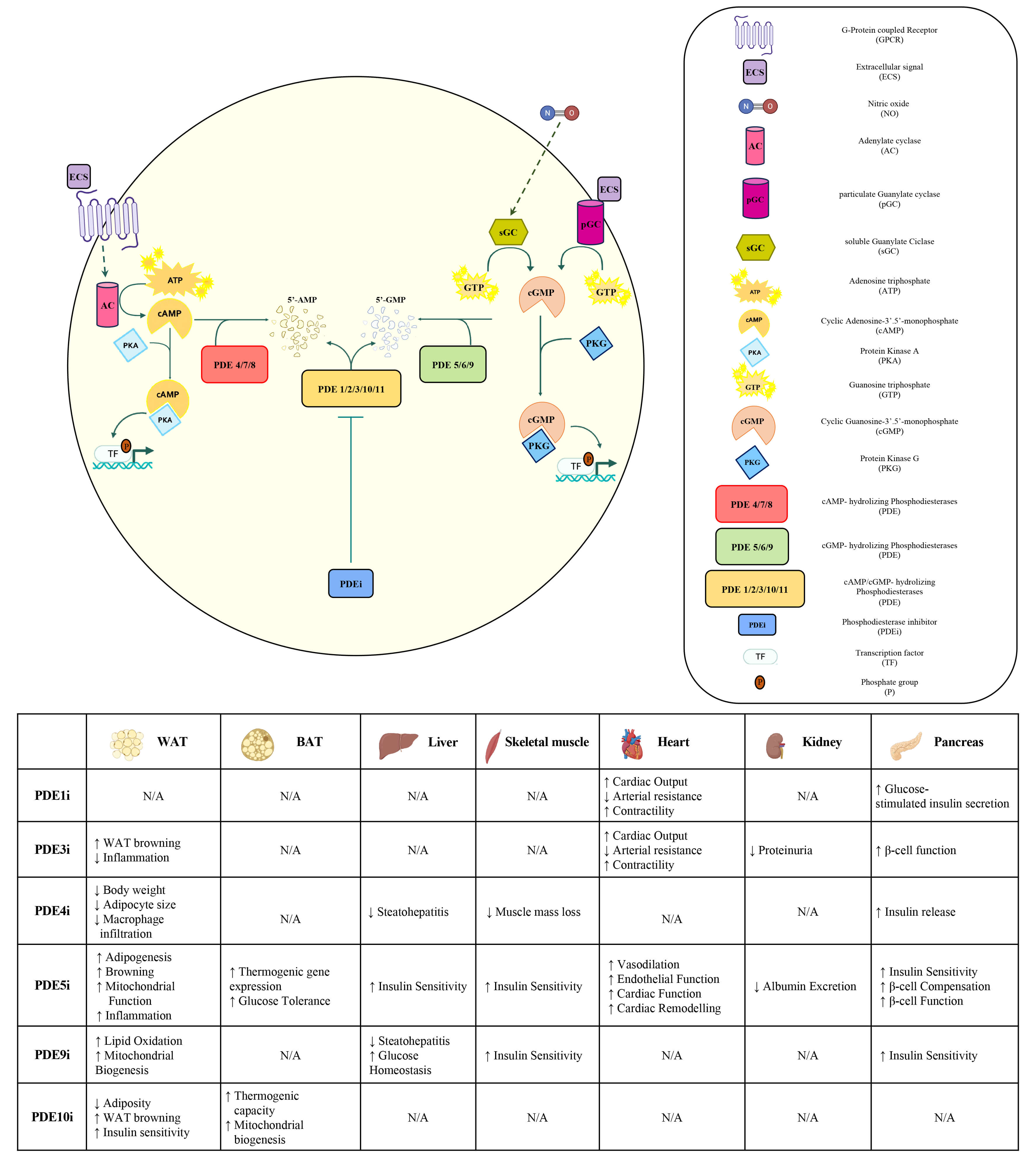 Fig. 1.
Fig. 1.
Overview of the cAMP and cGMP signalling pathways and known
effects of PDEi on key metabolically active tissues. The activation of the cAMP
signalling pathway is initiated by the binding of an extracellular ligand (e.g.,
peptide hormones, catecholamines, neurotransmitters) to a GPC receptor which
activates AC that in turn catalyses the conversion of ATP into cAMP. The
resulting increase in intracellular cAMP levels leads to the activation of the
PKA, a tetrameric enzyme that exerts its biological function through the
serine/threonine phosphorylation of specific downstream protein substrates,
including TFs, thereby modulating specific cellular responses and gene
expression. PDEs catalyses the hydrolysis of cAMP, resulting in its degradation.
PDEi act by selectively blocking the catalytic activity of PDEs, thereby
prolonging and compartmentalizing cAMP levels and potentiating the downstream
effects of PKA. In the cGMP signalling pathway, the binding of NO activates sGC,
and the binding of extracellular ligands activates pGC. Both sGC and pGC catalyse
the conversion of GTP into cGMP. Increased intracellular cGMP activates PKG,
which in turn phosphorylates downstream targets. PDEs catalyse the hydrolysis of
cAMP to 5′-AMP and cGMP to 5′-GMP, resulting in their degradation. Known effects
of PDEi on key metabolically active tissues are summarized in the table.
To ensure signaling specificity, cAMP–PKA activity is compartmentalized into subcellular domains, primarily orchestrated by AKAPs. These scaffold proteins tether PKA in close proximity to both substrates and key regulators, including AC and PDEs, such as PDE4 [24].
In parallel to the canonical PKA pathway, cAMP also activates exchange protein directly activated by cAMP (EPAC), a guanine nucleotide exchange factor that regulates small GTPases [26]. This PKA-independent pathway has been implicated in the modulation of key metabolic processes, including glucose metabolism, insulin secretion, and energy homeostasis [27].
Data obtained from mouse models have demonstrated that targeted disruption of
specific PKA subunits confers protection against diet-induced obesity and
metabolic dysfunction. In this regard, knockout of PKA regulatory subunit
RII
In the liver, the cAMP–PKA axis plays a crucial role in promoting glucose production during fasting [30]. Upon glucagon stimulation, elevated cAMP levels activate PKA, which phosphorylates and activates key transcription factors such as cAMP response element–binding protein (CREB). This induces the expression of gluconeogenic genes including phosphoenolpyruvate carboxykinase (PEPCK) and glucose-6-phosphatase (G6Pase), often in concert with the CREB regulated transcription coactivator 2 (CRTC2), thereby enhancing hepatic gluconeogenesis and contributing to systemic glucose availability during fasting states [31].
PKA also phosphorylates and inhibits acetyl-CoA carboxylase (ACC), reducing malonyl-CoA levels and promoting fatty acid oxidation (FAO), while simultaneously suppressing de novo lipogenesis [32].
Glucose production can be influenced by PKA also through post-translational mechanisms, including phosphorylation of bifunctional enzymes that modulate glycolysis and gluconeogenesis: dysregulation of these pathways contributes to hyperglycemia in T2DM, highlighting its potential as a therapeutic target [33].
In white adipose tissue (WAT), the cAMP-PKA signaling cascade exerts dual
regulatory control over both adipogenesis and lipolytic processes [34].
PKA-dependent phosphorylation triggers the activation of activating transcription
factor (ATF)/CREB transcription factors, which subsequently drives the
transcriptional induction of key adipogenic master regulators, including
peroxisome proliferator-activated receptor gamma (PPAR
Brown adipose tissue (BAT), by contrast, utilizes cAMP signaling to regulate
thermogenesis. Cold-induced sympathetic stimulation activates
Interestingly, cAMP also contributes to “WAT browning”, a transdifferentiation process by which white adipocytes adopt BAT-like features becoming beige/brite adipocytes, representing a promising strategy to fight obesity [39, 40].
Although less pronounced than in adipose and hepatic tissue, cAMP-PKA signaling also contributes to muscle glycogenolysis through phosphorylation of phosphorylase kinase, which activates glycogen phosphorylase to induce glucose release from glycogen stores during exercise. Moreover, PKA signaling has been implicated in enhancing glucose uptake through insulin-independent pathways under certain physiological contexts, although this remains less well-characterized [41]. In skeletal muscle, cAMP is also involved in acute regulation of contraction via modulation of calcium handling and long-term adaptation through enhanced glycolytic capacity and myofiber hypertrophy [42]. Moreover, epinephrine-induced cAMP production facilitates glycogenolysis during exercise and may help mitigate muscle atrophy under pathological conditions [43].
In pancreatic
Given the widespread metabolic impact of the cAMP pathway, its regulation is subject to intricate control mechanisms. A pivotal mechanism integrating the intracellular dynamics of cyclic nucleotides is the well-characterized cross-talk between the cAMP and cGMP pathways, primarily mediated by the dual-substrate phosphodiesterase 2 (PDE2). This enzyme hydrolyzes cAMP and is allosterically activated by cGMP, thereby establishing a robust negative feedback loop where elevated cGMP levels effectively limit cAMP accumulation [44, 45].
Such regulatory interplay has been extensively investigated in cardiac physiology, embryonic development, and vascular homeostasis, where tightly controlled cyclic nucleotide dynamics are crucial for functional integrity [46, 47].
Notwithstanding the established regulatory capacity of the PDE2-mediated cross-talk, the enzyme’s specific role in metabolic homeostasis remains a significant knowledge gap. Current evidence suggesting PDE2 involvement in metabolic regulation is tenuous, primarily stemming from a singular report that documented its expression and modulation within adipose tissue, correlating with obesity-related phenotypes in rodent models [48]. Crucially, this preliminary investigation did not include functional metabolic assessments, lacking data on the effects of PDE2 inhibition on key processes such as lipolysis rates, glucose uptake, or insulin sensitivity, thus leaving its direct contribution to adipocyte function largely unsubstantiated.
Together, these findings underscore the critical role of PKA in maintaining metabolic homeostasis and support its potential as a therapeutic target for the treatment of obesity and related metabolic disorders.
The cGMP-PKG signaling pathway plays a pivotal role in regulating energy homeostasis and glucose and lipid metabolism [10].
Upon cGMP elevation, PKG is activated and phosphorylates downstream effectors that influence energy balance, mitochondrial function, adipogenesis, and insulin sensitivity across multiple tissues [10, 12] (Fig. 1).
Emerging evidence indicates that modulation of this pathway influences both the
development and thermogenic activity of adipocytes, with direct implications for
metabolic health and resistance to obesity [15]. Loss of PKG impairs brown
adipose tissue thermogenic function, as evidenced by decreased UCP1 expression
and reduced mitochondrial content in PKG-I knockout mice [49]. Brown
preadipocytes isolated from these mice also show defective differentiation with
downregulation of thermogenic markers including PPAR
Recent findings suggested that activation of the cGMP-PKG pathway in adipocytes
promotes lipolysis, adiponectin secretion, and WAT browning, leading to enhanced
thermogenesis and energy expenditure [15, 52].In addition, natriuretic peptides promote mitochondrial biogenesis and
expression of UCP1 via PKG-dependent activation of p38 MAPK and PGC-1
Although the cAMP-PKA signaling pathway holds a preeminent role in governing central metabolic functions, including the rapid regulation of lipolysis, glycogenolysis, and thermogenesis, emerging literature suggests that cGMP-PKG signaling is crucial for metabolic adaptation, exerting a key influence on processes such as mitochondrial biogenesis, glucose uptake, and the modulation of insulin action. This necessitates an integrated view wherein the two signaling systems converge to ensure systemic metabolic flexibility. The convergence of these pathways is highlighted by a critical heterologous cross-talk mechanism, prominently featuring the cyclic nucleotide phosphodiesterases (e.g., PDE3-PDE5) [15].
Phosphodiesterase inhibitors are a diverse class of pharmacological agents that exert their effects by preventing the enzymatic hydrolysis of the second messengers cAMP and cGMP, thereby prolonging and amplifying cyclic nucleotide–mediated signaling [63]. PDE enzymes are divided into 11 families (PDE1–PDE11) based on their substrate specificity, regulatory mechanisms, and tissue distribution. Each isoform fine-tunes local cyclic nucleotide pools within specific subcellular microdomains, contributing to the spatial and temporal regulation of intracellular signaling networks [64]. Historically, PDEi have been developed and approved for non-metabolic indications, most notably, PDE5 inhibitors for erectile dysfunction and pulmonary hypertension, and PDE4 inhibitors for chronic obstructive pulmonary disease and inflammatory disorders [65, 66, 67].
Recent evidence has highlighted the broad expression of PDEs in key
metabolically active tissues including adipose tissue, liver, skeletal muscle,
pancreatic
Chronic low-grade inflammation within adipose tissue represents a pivotal driver
of the pathophysiology of MetS. Hypertrophic adipocytes and infiltrating immune
cells, particularly pro-inflammatory macrophages, establish a sustained
inflammatory milieu characterized by increased secretion of cytokines such as
TNF-
MetS itself is therefore defined as a cluster of interrelated cardiometabolic abnormalities (insulin resistance, abdominal obesity, dyslipidemia, hypertension, and hyperglycemia) that synergistically increase the risk of cardiovascular events and T2DM [73]. These pathological features are deeply connected to cyclic nucleotide-regulated signaling pathways, positioning PDEs as key molecular targets in these contexts. Preclinical studies investigating the effect of PDE blockade in metabolic disorders are summarized in Table 1 (Ref. [15, 18, 52, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86]).
| Gene | Substrate selectivity | Experimental model | Phenotype/outcomes | Reference |
| PDE3 | cAMP-specific, cGMP-inhibited | C57BL/6J male mice fed with HFD + 0.15% cilostazol for 16 weeks | Improvement in glucose tolerance and lipid profile, WAT browning | [74] |
| Pde3b-/- mice | Increases in respiratory uncoupling and FAO, improvement in energy homeostasis and insulin sensitivity, reduction in circulating FFA levels | [75] | ||
| PDE4 | cAMP-specific | Sprague–Dawley male rats fed with HFD + 1 mg/kg rolipram for 2 weeks | HFD-induced obesity impairs PDE4 increase in response to NA stimulation | [76] |
| C57BL/6J male mice fed with HFD + 21 mg/kg roflumilast for 12 weeks | PDE4 inhibition reduces weight gain, improved glucose tolerance and insulin sensitivity | [77] | ||
| Pde4b-/- mice | Pde4b-/- mice with HFD showed lower weights, smaller adipocytes, and decreased serum leptin levels suppressed macrophage infiltration in white adipose tissue | [78] | ||
| C57BL/6J male mice | In HFD-induced obese mice, PDE4 inhibition decreased the animals’ body weight, visceral adipose tissue weight, and adipocyte size | [79] | ||
| Male mice | In diabetic nephropathy, PDE4 inhibition showed suppressive effects on glycosylated hemoglobin, urinary albumin/creatinine ratio, suppressed pro-fibrotic and pro-inflammatory marker mRNAs and increased anti-reactive oxygen species marker mRNAs in the kidneys | [80] | ||
| Female db/db mice | PDE4 inhibition abolished the increase in blood glucose, reduced the increment in glycosylated hemoglobin, and induced insulin release in primary islets | [81] | ||
| Male Wistar rats | PDE4 inhibition decreased the loss of skeletal muscle mass in diabetic rats | [82] | ||
| PDE5 | cGMP-specific | C57BL/6J male mice fed with HFD + 12 mg/kg Sildenafil for 12 weeks | Chronic PDE5 inhibition improves insulin action and energy balance | [83] |
| Pde5a-/- mice | Improved glucose metabolism and white to beige adipocytes conversion, enhanced thermogenic capacity, reduced hepatic fat content | [15] | ||
| Adult mice | In T2DM model, PDE5 upregulation and lower cGMP levels were associated with insulin resistance and endothelial dysfunction | [84] | ||
| Male CD1 mice | PDE5 inibition expanded tissue anti-inflammatory TEMs, which are known to limit inflammation and promote tissue repair | [18] | ||
| PDE9 | cGMP-specific | Pde9a-/- mice | Pde9a-/- mice were resistant to HFD-induced obesity | [52] |
| C57BL/6N male and female mice | In HFD-induced obesity, PDE9 inhibition reduced body fat, enhanced mitochondrial activity and improved metabolic syndrome markers | [85] | ||
| PDE10 | cAMP-specific, cGMP-sensitive | Adult female CD1 and C57BL/6 mice | PDE10 inhibition in HFD-induced obese mice resulted in weight loss, browning of WAT, and improved insulin sensitivity | [84] |
| C57Bl/6N Tac mice | In HFD-induced obesity, PDE10 inhibition showed lower fat fraction in white and brown adipose tissue an upregulation of marker genes involved in WAT beigeing | [86] |
-/-, knockout; HFD, high fat diet; WAT, white adipose tissue; FAO, fatty acid oxidation; FFA, free fatty acid; NA, norepinephrine; TEM, TIE2-expressing monocytes; T2DM, type 2 diabetes; cGMP, guanosine monophosphate; cAMP, cyclic adenosine monophosphate.
Among PDEi, PDE5 inhibitors (PDE5i) such as sildenafil and tadalafil, promote
nitric oxide (NO)-dependent vasodilation by blocking cGMP degradation in vascular
smooth muscle [87]. Beyond their vascular effects, PDE5i enhance adipogenesis,
induce browning of WAT, and improve mitochondrial respiration [88, 89, 90]. In
preclinical and clinical settings, PDE5i have been demonstrated to improve
insulin sensitivity, reduce systemic inflammation, and enhance endothelial
function, mechanisms of paramount importance in MetS and T2DM [91, 92]. A
randomized controlled trial using sildenafil (25 mg three times daily for three
months) in individuals with pre-diabetes revealed significant improvement in
insulin sensitivity, enhanced fibrinolytic balance, and reduction of urinary
albumin excretion, although without augmentation of glucose-stimulated insulin
secretion [61]. Similarly, 20 mg tadalafil daily administered to obese
insulin-resistant patients improved
PDE3 inhibitors (PDE3i), including milrinone and cilostazol, modulate both cAMP
and cGMP levels. While their approved uses are related to heart failure and
intermittent claudication, PDE3B, highly expressed in adipocytes, liver, and
pancreatic islets, has been implicated in energy homeostasis [95, 96]. Mice
overexpressing Pde3b exhibit glucose intolerance and
PDE1 inhibitors (PDE1i) regulate both cAMP and cGMP via a
Ca2+/calmodulin-dependent mechanism. PDE1C inhibition enhances
glucose-stimulated insulin secretion in pancreatic
PDE4 inhibitors (PDE4i), including roflumilast and apremilast, are potent
anti-inflammatory agents, modulating immune cell activity and cytokine release.
In HFD-fed mice, roflumilast reduced weight gain, enhanced energy expenditure,
improved glucose tolerance and insulin sensitivity, and attenuated
steatohepatitis via PKA/CREB activation and PGC-1
PDE9 inhibitors (PDE9i) represent a novel class targeting natriuretic peptide-derived cGMP signaling and PDE9 is highly expressed in heart and adipose tissue modulating pathways involved in lipid oxidation and mitochondrial biogenesis [106]. Genetic deletion or pharmacological inhibition of PDE9 improves glucose homeostasis, enhances insulin sensitivity, and reduces hepatic steatosis in obese and diabetic mice and these effects are mediated via PKG activation and upregulation of mitochondrial and oxidative genes [52, 85].
Pharmacological blockade of PDE10A using the highly selective PDE10i MP-10 has been shown to activate BAT and enhance thermogenic capacity in vivo. In murine models of diet-induced obesity, chronic treatment with MP-10 promotes body weight reduction through elevated whole-body energy expenditure, concomitant with the induction of browning programs in WAT and an improvement in systemic insulin sensitivity [107].
Clinical studies testing the use of PDEi in metabolic disorders have predominantly focused on selective pharmacological agents that offer mechanistic precision. These include, but are not limited to, PDE5 inhibitors (e.g., sildenafil, tadalafil) and PDE4 inhibitors (e.g., roflumilast), which precisely modulate specific cyclic nucleotide signaling pathways governing lipolysis, insulin sensitivity, and energy expenditure. In sharp contrast, a minority of studies have explored the utility of non-selective or naturally derived modulators, such as caffeine and polyphenols, often tested in combination with other treatments. These natural compounds typically exert a broader, yet milder, spectrum of PDE inhibition alongside inherent thermogenic, antioxidant, and anti-inflammatory properties. While the selective inhibitors provide critical translational insights into the role of specific signaling cascades, the non-selective approach demonstrates that weaker, pleiotropic PDE modulation, particularly when synergized with other biological activities, can still significantly influence metabolic regulation. Both complementary approaches are vital, offering a broader and more nuanced understanding of the therapeutic potential of PDE-targeted strategies against metabolic dysfunction.
As summarized in Table 2, the majority of registered clinical trials (RCTs) investigating PDEi in obesity and related metabolic disorders are early-phase, with relatively small sample sizes and variable study designs. A significant proportion of interventions evaluated indirect modulators of PDE signaling, such as caffeine, aiming to stimulate thermogenesis and enhance energy expenditure, though most failed to demonstrate consistent clinical benefits in terms of weight reduction or body composition (Table 3). More targeted approaches, including the use of pentoxifylline and roflumilast, have shown modest but measurable improvements in insulin sensitivity and inflammatory markers in obese or prediabetic populations, underscoring the translational potential of PDE blockade in metabolic disease.
| ID | Title | Sex | Enrollment | Status | Results | Interventions | Phase | Design | Outcome | Sponsor |
| NCT00685945 | Renin-Angiotensin Aldosterone System and Fibrinolysis Interaction in Humans | Both | 24 adults, older | Completed | Yes | Bradykinin, L-NMMA + bradykinin, Isosorbide + L-NMMA + bradykinin, Sildenafil + L-NMMA + bradykinin | NA | Randomized | t-PA Release | Vanderbilt University |
| NCT01444651 | A Trial of Tadalafil and Glycemic Traits | Both | 73 adults | Completed | Yes | Tadalafil, placebo | 3 | Randomized | Insulin resistance | Dr. Thomas J. Wang |
| NCT01862029 | Effects of Roflumilast on Insulin and Blood Sugar Levels in Prediabetic Overweight and Obese Individuals | Both | 24 adults, older | Completed | Yes | Roflumilast | 2 | Single-group assignment | Insulin Sensitivity | National Heart, Lung, and Blood Institute (NHLBI) |
| NCT02524184 | Sildenafil Activates Browning of White Adipose Tissue and Improves Insulin Sensitivity | Male | 11 adults | Unknown | No | Sildenafil, placebo | 4 | Randomized | WAT browning | Dr. Xiang Guang-da |
| NCT02554045 | Daily Tadalafil on Body Fat and Lean Mass | Male | 20 adults, older | Completed | No | Tadalafil, placebo | 4 | Randomized | Variation in lean and fat mass | La Sapienza University of Rome |
| NCT02595684 | Effect of Tadalafil on Insulin Secretion and Insulin Sensitivity in Obese Men | Male | 18 adults | Completed | Yes | Tadalafil, placebo | 4 | Randomized | Fasting Glucose | University of Guadalajara |
| NCT02819440 | PDE5i for Obesity-Related Cardiometabolic Dysfunction | Both | 141 adults | Completed | Yes | Tadalafil, Placebo | 2 | Randomized | Resting Energy Expenditure | Vanderbilt University Medical Center |
| NCT03905018 | Effect of Tadalafil Administration on Vasodilatation Mediated by Flow in Patients with Obesity Grade I-II | Male | 80 adults | Unknown | No | Tadalafil | 3 | Randomized | Vasodilatation | Centro Universitario de Ciencias de la Salud, Mexico |
| NCT04623840 | Combined Effect of Continuous and Interval in Addition to Tadalafil on Erectile Dysfunction | Male | 60 adults | Unknown | No | Tadalafil | NA | Randomized | Five-Item Version of International Index of Erectile Function | Cairo University |
| NCT04684589 | Effect of PDE5 Inhibition on Adipose Metabolism in Humans | Both | 100 adults | Recruiting | No | Tadalafil, Placebo | 2 | Randomized | Thermoneutral FSF of WAT | Vanderbilt University Medical Center |
| NCT05051436 | The Effects of Mirabegron and Tadalafil on Glucose Tolerance in Prediabetics | Both | 96 adults, older | Recruiting | No | Mirabegron, Tadalafil, Placebo | 4 | Randomized | Oral glucose tolerance test | Philip Kern |
| NCT06125665 | Aminophylline on Perioperative Lung Mechanics in COPD Morbidly Obese Patients Undergoing Laparoscopic Bariatric Surgery | Both | 60 adults | Recruiting | No | Aminophylline-Dexmedetomidine | NA | Randomized | Static lung compliance | Tanta University |
NA, not assigned; tPA, tissue plasminogen activator; L-NMMA, L-NG-monomethylarginine, acetate salt; COPD, chronic obstructive pulmonary disease; FSF, fat signal fraction; WAT, white adipose tissue.
| ID | Title | Sex | Enrollment | Status | Results | Interventions | Phase | Design | Outcome | Sponsor |
| NCT00302289 | B181 Stimulation of Thermogenesis by Bio-Active Food Ingredients | Male | 12 adults | Completed | No | Tyrosine, green tea, caffeine | 1 | Randomized | Energy expenditure, appetite ratings and energy intake | University of Copenhagen |
| NCT00377975 | “Pecos” B-adrenergic and PPAR-G Stimulation Upregulates Lipid Metabolism in Human Subcutaneous Fat | Both | 96 adults | Completed | No | Ephedrine, pioglitazone, caffeine | 2 | Randomized | Fat percentage, UCP1 gene expression, visceral adiposity | Pennington Biomedical Research Center |
| NCT00611416 | The Role of Tea Catechins and Caffeine in Relation to Energy Metabolism | Male | 15 adults | Completed | No | Green tea, mix of catechins and caffeine, EGCG, EGC, Caffeine, Placebo | NA | Randomized | Resting Energy Expenditure | University of Copenhagen |
| NCT00681733 | Pentoxifylline Versus Pioglitazone In Non-Alcoholic Steatohepatitis (NASH) | Both | 20 adults, older | Unknown | No | Pioglitazone, pentoxifylline | NA | Randomized | Improvement in metabolic profile and histology | Govind Ballabh Pant Hospital |
| NCT00692731 | Efficacy of a Tea Catechin Sports Drink for Enhancing Exercise-Induced Fat Loss | Both | 132 adults, older | Completed | No | Catechins, caffeine | NA | Perspective | Body fat mass | Provident Clinical Research |
| NCT00770328 | The Effects of Pentoxifylline on PAI-1 in an Obese Population | Both | 37 adults, older | Completed | Yes | Pentoxifylline, placebo | 4 | Randomized | PAI-1 levels | Vanderbilt University |
| NCT01556321 | Long-term Effects of Green Tea on Gut Flora, Fat Absorption, Body Composition and Resting Energy Expenditure | Both | 70 adults | Completed | No | Placebo, green tea | NA | Randomized | Gut flora composition | Maastricht University Medical Center |
| NCT01596907 | Treatment of Low Metabolic Rate Following Bariatric Surgery | Both | 218 adults, older | Completed | No | Ephedrine sulfate with caffeine, cellulose | NA | Randomized | Resting Energy | Oregon Weight Loss Surgery, LLC |
| NCT01691196 | Inflammation in Peritoneal Dialysis Patients Effect of Obesity | Both | Unknown | Withdrawn | No | Unknown | NA | Perspective | Levels of inflammatory cytokines | University of Illinois at Chicago |
| NCT01710722 | The Effect of Leptin A-200, Caffeine/Ephedrine and Their Combination Upon Weight Loss and Body Composition in Man | Both | 45 adults | Completed | No | Caffeine and ephedrine, Leptin A, caffeine, ephedrine, and leptin A | NA | Randomized | Percent loss of total body fat to visceral fat | Pennington Biomedical Research Center |
| NCT01815203 | Caffein Consumption and Response Inhibition | Male | 21 adults | Completed | No | Caffeine, placebo | NA | Randomized | Insulin Sensitivity | Uppsala University |
| NCT02048215 | Effect on Energy Metabolism at Cellular Level of Diet Plus Treatment with Ephedrine and Caffeine in Obesity | Female | 13 adults | Completed | No | Caffeine, placebo, ephedrine, hypocaloric diet | 3 | Randomized | Resting energy expenditure | Istituto Auxologico Italiano |
| NCT02157974 | Liver and Fat Regulation in Overweight Adolescent Girls | Female | 92 children, adults | Completed | Yes | Byetta exenatide | 3 | Non randomized | Hepatic Glucose Release | University of Colorado, Denver |
| NCT02185638 | Appetite Suppression Effects of an Herbal Combination of Yerba matè, Guarana, Damiana (YGD), and 12 Other Herbal Components, Versus YGD Alone | Female | 19 adults | Completed | No | Yerba maté, guarana and damiana (YGD) and Akavar (A20-50) | NA | Randomized | Appetite sensations | University of Copenhagen |
| NCT02740660 | Body Composition Changes with Albuterol and Caffeine Versus Placebo in Adolescents | Both | 12 children | Completed | Yes | Caffeine, albuterol, placebo | NA | Randomized | Fat Mass | Pennington Biomedical Research Center |
| NCT02751840 | Effects of Caffeine and Coffee on Resting Metabolic Rate, Comparing Normal Weight Men to Obese Men | Male | 33 adults | Completed | No | Caffeine, placebo, decaffeinated | 3 | Randomized | Resting metabolism | Tel Hai College |
| NCT02758990 | Interventional Testing of Gene-environment Interactions Via the Verifomics Mobile Application | Both | 16 adults, older | Terminated | No | Vitamin A, Vitamin B6, Vitamin C, Nicotinamide, Vitamin D3, Vitamin, Broccoli, Spinach, Caffeine, Coffee, Chocolate | NA | Non Randomized | Frequency of correct predictions | Verifomics LLC |
| NCT03512496 | Metabolic and Genetic Impacts of Energy Drinks in Youth | Both | Unknown | Withdrawn | No | Acute energy drink, Placebo Acute energy drink, Caffein Acute energy drink, Decaf | NA | Randomized | Glucose tolerance | University of Calgary |
| NCT03717935 | Oral Amino Acid Nutrition to Improve Glucose Excursions in PCOS | Female | 27 child, adults | Completed | Yes | Essential Amino Acid, Placebo | NA | Randomized | Hepatic Fat Fraction | University of Colorado, Denver |
| NCT04025060 | Reducing Sugar-sweetened Beverage Consumption Among Children | Both | 29 children | Completed | No | Caffeine-free soda, Carbonated water, Regular soda | NA | Randomized | Adherence | George Washington University |
| NCT05445232 | An Interaction (DDI) Study of LY3437943 in Obese Participants | Both | 32 adults, older | Completed | No | Midazolam, Warfarin, Caffeine | 1 | Non randomized | Pharmacokinetics | Eli Lilly and Company |
| NCT06564441 | A Study to Test Whether BI 456906 (Survodutide) Influences the Amount of Bupropion, Caffeine and Midazolam in the Blood in People with Overweight or Obesity | Both | 34 adults | Active not recruiting | No | Survodutide, Bupropion, Caffeine, Midazolam | 1 | Interventional | AUC bupropion in plasma | Boehringer Ingelheim |
NA, not assigned; UCP1, uncoupling protein 1; EGCG, epigallocatechin-3-gallate; EGC, epigallocatechin; PAI-1, plasminogen activator inhibitor-1; PCOS, polycystic ovary syndrome; AUC, area under the curve.
A comparative assessment of the clinical trials involving the use of PDEi for the management of obesity and related metabolic dysfunctions highlights distinct features between selective and non-selective phosphodiesterase inhibition. Selective PDEi, particularly those targeting PDE5 and PDE4, exhibit more consistent and mechanistically defined effects, including improvements in insulin sensitivity, endothelial function, and systemic inflammation. These outcomes likely reflect their well-characterized pharmacokinetic and pharmacodynamic properties and tissue-specific modulation of cyclic nucleotide signaling. Conversely, studies investigating non-selective or naturally-derived PDE modulators, such as caffeine, tea catechins, and polyphenols, report broader, though less specific, metabolic effects often intertwined with antioxidant or thermogenic actions. While these agents show modest efficacy in weight reduction and metabolic improvement, their limited isoform selectivity complicates the interpretation of causal mechanisms.
The examination of RCTs involving the use of PDEi in the context of T2DM (Tables 4,5) highlights a heterogeneous body of evidence, with most studies being small, early-phase investigations focusing on surrogate endpoints such as vascular function, hemodynamic parameters, and markers of end-organ damage (e.g., albuminuria, intima-media thickness). Several randomized controlled trials with PDE5i primarily addressed improvements in endothelial function and cardiac remodeling—surrogate measures of clinical relevance in the T2DM population—supporting the notion of pleiotropic effects via the NO–cGMP pathway and myocardial mechanics. Other compounds, including pentoxifylline and cilostazol, were mainly evaluated for renal and anti-atherothrombotic outcomes, with some trials documenting reductions in proteinuria and favorable modulation of inflammatory biomarkers. Although these findings provide encouraging signals on intermediate endpoints, the methodological heterogeneity—differences in primary outcomes, limited treatment durations, and underpowered sample sizes—substantially limits generalizability and precludes firm conclusions regarding long-term clinical benefit in T2DM.
| ID | Title | Sex | Enrollment | Status | Results | Interventions | Phase | Design | Outcome | S Sponsor |
| NCT00527995 | Acute Effects of Sildenafil on Endothelial Function in People With Diabetes | Male | 40 adults, older | Completed | No | Sildenafil | 3 | Randomized | Improvement of flow mediated dilatation of the brachial artery | Ruhr University of Bochum |
| NCT00645268 | A Multicenter, Double-blind Study to Evaluate the Effect of Pretreatment With a Daily Dose of Sildenafil on the As-Needed Efficacy of Viagra in Men With Erectile Dysfunction and Type 2 Diabetes | Male | 300 adults, older | Completed | No | Sildenafil, Placebo | 4 | Randomized | The IIEF Erectile Function Domain score | Viatris Inc. |
| NCT00692237 | Cardiovascular Effects of Chronic Sildenafil in Men With Type 2 Diabetes | Male | 59 adults, older | Completed | Yes | Sildenafil, Placebo | 4 | Randomized | Left Ventricular Torsion | La Sapienza University of Rome |
| NCT00823849 | Study of Cilostazol and Probucol to Assess Their Effects on Atherosclerosis Related Biomarker | Both | 200 adults, older | Completed | No | Cilostazol, Probucol, Cilostazol + Probucol | 4 | Randomized | Primary Efficacy Evaluation | Otsuka Beijing Research Institute |
| NCT00886574 | Cilostazol Versus Aspirin for Primary Prevention of Atherosclerotic Events | Both | 400 adults, older | Unknown | No | Cilostazol, Aspirin | 4 | Randomized | Maximal and mean IMT of both common carotid arteries | Hanyang University |
| NCT01076478 | Asian Study on Cilostazol Effectivity in Neuropathies of Diabetes Mellitus Type 2-A Pilot Study in the Philippines | Both | 47 adults, older | Completed | No | Cilostazol | 4 | Randomized | Subjective neuropathy assessment by NSS | Otsuka Pharmaceutical, Inc., Philippines |
| NCT01084369 | Effect of Testosterone on Endothelial Function and Microcirculation in Type 2 Diabetic Patients with Hypogonadism | Male | 22 adults, older | Terminated | No | Testosterone | 4 | NA | Improvement in endothelial dependent and endothelial-independent vasodilatation and function | Tameside Hospital NHS Foundation Trust |
| NCT01180283 | Efficacy and Safety of Lodenafil Carbonate in the Treatment of Erectile Dysfunction in Patients with Diabetes | Male | Adults, older | Completed | No | lodenafil carbonate | 4 | Non randomized | Erectile function | Cristália Produtos Químicos Farmacêuticos Ltda. |
| NCT01200394 | A Phase 2, Placebo-Controlled Study to Evaluate The Efficacy And Safety Of PF-00489791 In Patients With Type 2 Diabetes And Overt Nephropathy | Both | 256 adults, older | Completed | Yes | PF-00489791, Placebo | 2 | Randomized | Change From Baseline in UACR | Pfizer |
| NCT01238224 | Effects of PDE-5 Inhibition on Postprandial Hyperglycemia in Type 2 Diabetes | Both | 22 adults, older | Terminated | No | Tadalafil | 1 | Randomized | Capillary recruitment, muscle glucose uptake and circulating glucose levels | Vastra Gotaland Region |
| NCT01566006 | Mycophenolate Mofetil, Carnitine and PDE5 Inhibitor, Three Potential Treatments for Resistant Proteinuria Slowing Diabetic Nephropathy Deterioration | Both | 80 adults, older | Unknown | No | Mycophenolate Mofetil (MMF), Phosphodiesterase 5 inhibitors, Carnitine | NA | Randomized | Proteinuria | The Nazareth Hospital, Israel |
| NCT01803828 | REmodelling in Diabetic CardiOmapathy: Gender Response to PDE5i InhibiTOrs | Both | 120 adults, older | Completed | No | Tadalafil, Placebo | 4 | Randomized | Change from Baseline in Left Ventricular torsion | La Sapienza University of Rome |
| NCT02219646 | Diabetes & Vardenafil | Male | 54 adults, older | Completed | No | Vardenafil, Placebo | 2 | Randomized | Endothelin-1 levels | Azienda USL Modena |
| NCT02266030 | Effect of Cilostazol on Coronary Artery Stenosis and Plaque Characteristics in Patients with T2DM | Both | 100 adults, older | Completed | No | Cilostazol, Aspirin | 3 | Randomized | Coronary artery stenosis | Seoul National University Bundang Hospital |
| NCT02601989 | Effects on Insulin Resistance with Tadalafil in Type 2 Diabetes: a Double-blind, Placebo-controlled Crossover Study | Both | 23 adults, older | Completed | No | Tadalafil, Placebo | 2 | Randomized | Insulin sensitivity | Goteborg University |
| NCT02933788 | Effect of aSpirin Versus CilOstazol for Inhibition of Antiplatelet aggRegaTion in Type 2 DM Patients | Both | 116 adults, older | Unknown | No | Cilostazol, Acetylsalicylic acid | 4 | Randomized | Platelet reactivity testing | Kangbuk Samsung Hospital |
| NCT02983214 | Diabetic Artery Obstruction: is it Possible to Reduce Ischemic Events with Cilostazol? | Both | 826 adults, older | Completed | No | Clopidogrel, Cilostazol | 4 | Randomized | Acute ischemic stroke/TIA, MI, or death from vascular cells | University of Ioannina |
| NCT03248401 | Effect of Cilostazol on Carotid Atherosclerosis Estimated by 3D Ultrasound in Patients with Type 2 Diabetes | Both | 50 adults, older | Completed | No | Cilostazol, Aspirin | 4 | Randomized | Change of carotid artery atherosclerosis, plaque composition | Seoul National University Bundang Hospital |
| NCT03462017 | Pharmacodynamic Study to Assess the Effects of Repeated Dosing of SAR247799 on Endothelial Function in Patients with Type 2 Diabetes Mellitus | Both | 54 adults | Completed | No | SAR247799, Placebo, Sildenafil, Acetylcholine | 1 | Randomized | Change in Flow Mediated Dilation | Sanofi |
| NCT04170790 | Evaluation of Drug Interactions of Saxagliptin with Sildenafil in Healthy Volunteers | Male | 18 adults | Unknown | No | Saxagliptin, Sildenafil | NA | Interventional | Maximum Plasma Concentration | Ain Shams University |
| NCT05487755 | Investigational and Comparative Study in the Management of Diabetic Nephropathy | Both | 90 adults, older | Completed | No | Tadalafil Oral Tablet, Pentoxifylline Oral Tablet | 3 | Randomized | Change in Urinary albumin/creatinine ratio | Tanta University |
| NCT06402747 | Clopidogrel Versus Cilostazol on Vessels | Both | 120 adults, older | Recruiting | No | Clopidogrel, Cilostazol | 4 | Randomized | Changes of carotid intima-media thickness | Seoul National University Bundang Hospital |
| NCT06989697 | Cilostazol-Ginkgo for Cognitive Function in Elderly Diabetes | Both | 80 adults, older | Recruiting | No | Cilostazol, Ginko Leaf Dried Ext., Placebo | 3 | Randomized | Change in MMSE Score | Seoul National University Bundang Hospital |
NA, not assigned; IIEF, international index of erectile function; IMT, intima media thickness; NSS, neuropathy symptom score; UACR, urinary albumin creatinine ratio; TIA, transient ischemic attack; MI, myocardial infarction; MMSE, mini-mental state examination.
| ID | Title | Sex | Enrollment | Status | Results | Interventions | Phase | Design | Outcome | S Sponsor |
| NCT00432887 | Experimental Studies of the Effects of Caffeine on Glucose Regulation | Both | 150 adults, older | Completed | No | Caffeine | 1 | Randomized | Postprandial glucose and insulin, fasting glucose and insulin, insulin resistance, glucose tolerance | National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK) |
| NCT00663949 | Assessment of the Effect of Captopril Versus Combination of Captopril and Pentoxifylline on Reducing Proteinuria in Type 2 Diabetic Nephropathy | Both | 70 adults, older | Completed | No | Captopril, Captopril + Pentoxifylline | 3 | Randomized | Decreasing urinary protein | Shiraz University of Medical Sciences |
| NCT00950898 | The Acute Effects of Coffee on Glucose Metabolism | Male | 11 adults | Completed | No | Caffeine, Placebo | NA | Randomized | Plasma glucose levels | Brooklyn College of the City University of New York |
| NCT01030796 | Quitting Caffeine for Better Glucose Metabolism | Both | 25 adults, older | Completed | No | Caffeine | 1 | NA | Caffeine abstinence | National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK) |
| NCT01382303 | Effect of Pentoxifylline on Proteinuria in Korean Type 2 Diabetic Patients | Both | 174 adults, older | Completed | Yes | Pentoxifylline, Placebo | 4 | Randomized | Percentage Change in Proteinuia, Changes of urine protein to creatinie ratio | Ajou University School of Medicine |
| NCT02291666 | Evaluation of CYP450 Activities in Diabetic Patients vs. Non-diabetic Subjects | Both | 73 adults, older | Completed | No | CRCHUM-MT cocktail | 4 | Non randomized | Metabolic ratio | Centre hospitalier de l’Universitè de Montrèal (CHUM) |
| NCT02929901 | The Effects of Coffee Main Constituents (Caffeine and Chlorogenic Acid) Supplementation on Inflammatory, Metabolic Factors, Hepatic Steatosis and Fibrosis in NAFLD Patients with T2DM | Both | 200 adults, older | Completed | No | Caffeine and chlorogenic acid, caffeine, chlorogenic acid, placebo | 3 | Randomized | Hepatic steatosis | National Nutrition and Food Technology Institute |
| NCT03006952 | Add-on Pentoxifylline to Losartan Versus Increasing Dose of Losartan on NT-PRO BNP in Type 2 Diabetics with Nephropathy | Both | 59 adults, older | Completed | No | Pentoxifylline, Losartan | 4 | Randomized | NT-pro BNP levels | Tehran University of Medical Sciences |
| NCT03664414 | Pentoxifylline Effect in Patients with Diabetic Nephropathy. | Both | 196 adults, older | Unknown | No | Pentoxifylline | 4 | Randomized | Change in the glomerular filtration rate | Maria Eugenia Galván Plata |
| NCT04243850 | REGROUP: Renohemodynamic Effects empaGliflozin in vaRiOUs Populations | Both | Adults, older | Withdrawn | No | Empagliflozin, Placebo oral tablet, Caffeine | 4 | Randomized | Glomerular filtration rate | Amsterdam UMC |
| NCT04504045 | Metformin’s Effect on Drug Metabolism in Patients with Type 2 Diabetes | Both | 10 adults, older | Terminated | No | Metformin, Caffeine, Efavirenz, Losartan, Omeprazol, Metoprolol, Midazolam | 1 | NA | Change from Baseline in Metabolic Rate | University of Southern Denmark |
| NCT04910178 | Follow-up of NAFLD Patients With MRI-PDFF | Both | 80 adults, older | Completed | No | Empagliflozin, Ursodeoxycholic acid, Pentoxifylline 400, placebo | 4 | Randomized | Liver fat content | Asmaa Abdelfattah Elsayed |
NA, not assigned; NT-pro BNP, N-terminal pro-B-type natriuretic peptide.
Comparison of the clinical studies analyzing the effect of PDEi for the treatment of T2DM reveals notable differences in the clinical impact and mechanistic scope of selective versus non-selective phosphodiesterase inhibition in T2DM management. Selective PDEi, particularly PDE5 and PDE3 inhibitors, have demonstrated significant benefits in improving endothelial function, insulin-mediated glucose uptake, and peripheral perfusion, reflecting their targeted modulation of cGMP or cAMP signaling in metabolic tissues. These agents also show favorable effects on cardiovascular risk markers, supporting their potential as adjunctive therapies for metabolic and vascular comorbidities in diabetes. In contrast, non-selective or naturally derived PDE modulators, such as methylxanthines, polyphenols, and catechins, exert milder but broader metabolic actions, often associated with enhanced mitochondrial function, antioxidant capacity, and lipid metabolism. However, their heterogeneous composition and lower PDE selectivity limit the ability to attribute observed benefits to specific cyclic nucleotide pathways. Overall, while selective PDEi provide stronger mechanistic and clinical evidence of metabolic improvement, non-selective modulators continue to offer valuable insights into the systemic regulation of energy metabolism and redox balance in diabetic conditions.
Most available clinical studies dissecting the effects of PDEi in MetS are early-phase, single-center investigations with relatively small sample sizes and variable primary outcomes, spanning from vascular function and arterial stiffness to insulin signaling and erectile function (Tables 6,7).
| ID | Title | Sex | Enrollment | Status | Results | Interventions | Phase | Design | Outcome | Sponsor |
| NCT00573950 | Effects of Cilostazol on Plasma Adipocytokine and Arterial Stiffness | Both | 48 adults | Unknown | No | Cilostazol, Placebo | 4 | Randomized | The effect on pulse wave velocity | Korea University Anam Hospital |
| NCT00738400 | Study of Vardenafil in Patients Suffering From Erectile Dysfunction and Metabolic Syndrome | Male | 150 adults | Completed | Yes | Vardenafil (Levitra, BAY38-9456), Placebo | 4 | Randomized | Change From Baseline in IIEF-EF | Bayer |
| NCT00750308 | Renin Angiotensin Aldosterone System (RAAS) and Fibrinolysis in Humans: ACEi and PDE5i | Both | 27 adults, older | Completed | Yes | Ramipril, Tadalafil, placebo | NA | Randomized | Beta cells function | Vanderbilt University |
| NCT01106118 | Therapeutic Effectiveness of Vardenafil in Patients With Erectile Dysfunction and Metabolic Syndrome in Daily Clinical Practice | Male | 2289 adults, older | Completed | No | Vardenafil (Levitra, BAY38-9456) | NA | Observational | Improvement in erectile function | Bayer |
| NCT01334554 | Study of Sildenafil Citrate on Insulin Resistance in African American | Female | 46 adults | Completed | Yes | Sildenafil, Placebo | 2 | Randomized | Insulin Sensitivity | Vanderbilt University |
| NCT02129725 | Effect of Prolonged PDE-5 Inhibition on Insulin Signaling in Skeletal Muscle | Both | 15 adults, older | Completed | Yes | Sildenafil citrate, Placebo | 4 | Randomized | Insulin-stimulated AKT Phosphorylation | Vanderbilt University |
| NCT02963454 | Effects of Levosimendan on Cellular Metabolic | Both | 50 adults, older | Unknown | No | Dobutamine, Levosimendan | NA | Randomized | Changes in the concentration of glucose in the extracellular fluid of the skeletal muscle | Military Hospital of Tunisy |
| NCT04383093 | Tadalafil Plus Tamsulosin for Male LUTS and ED | Male | 75 adults, older | Completed | No | Tadalafil | NA | Observational | Lower Urinary Tract Symptoms | University of Florence |
| NCT05292690 | An Assistive Powered Wheelchair: Stage 2 Trial | Both | 17 adults, older | Completed | No | Tadalafil | NA | Observational | Value of an Obstacle Alerting System | East Kent Hospitals University NHS Foundation Trust |
NA, not assigned; IIEF-EF, International Index of Erectile Function - Erectile Function.
| ID | Title | Sex | Enrollment | Status | Results | Interventions | Phase | Design | Outcome | S Sponsor |
| NCT00310323 | Hepatic Drug Biotransformation in Children With Obstructive Sleep Apnea | Both | 69 children | Completed | No | Dextromethorphan, Caffeine | NA | Non Randomized | Caffeine urinary molar ratio, Dextromethorphan urinary molar ratio | Virginia Commonwealth University |
| NCT02963454 | Effects of Levosimendan on Cellular Metabolic | Both | 50 adults, older | Unknown | No | Dobutamine, Levosimendan | NA | Randomized | Changes in the concentration of glucose in the extracellular fluid of the skeletal muscle | Military Hospital of Tunisy |
| NCT03041129 | Post-Prandial Liver Glucose Metabolism in PCOS | Female | 19 children, adults | Completed | Yes | NA | NA | Observational | Hepatic Fat Fraction measured by MRI | University of Colorado, Denver |
| NCT04278404 | Pharmacokinetics, Pharmacodynamics, and Safety Profile of Understudied Drugs Administered to Children Per Standard of Care (POPS) | Both | 5000 children, adults | Recruiting | No | NA | NA | Observational | Clearance | Duke University |
| NCT04383093 | Tadalafil Plus Tamsulosin for Male LUTS and ED | Male | 75 adults, older | Completed | No | Tadalafil | NA | Observational | Lower Urinary Tract Symptoms | University of Florence |
| NCT05687474 | Baby Detect: Genomic Newborn Screening | Both | 6000 children | Recruiting | No | NA | NA | Observational | Acceptability | Centre Hospitalier Universitaire de Liege |
| NCT05713773 | Pharmacodynamics and Pharmacokinetics of 3 New Developed Coated Glucose Beads in 20 Obese Healthy Subjects | Both | 20 adults | Completed | No | Glucose, Caffeine | 1 | Randomized | GLP-1 levels | Aphaia Pharma US LLC |
| NCT06533007 | PDE3B in Metabolic Regulation | Both | 40 adults, older | Not yet recruiting | No | NA | NA | Observational | Lypolysis | Cambridge University Hospitals NHS Foundation Trust |
NA, not assigned; MRI, magnetic resonance imaging; GLP-1, glucagon-like peptide-1.
The comparison between the clinical trials designed to test the use of PDEi for the management of MetS underscores both the mechanistic specificity and the translational heterogeneity of phosphodiesterase inhibition in the context of MetS. Selective PDEi, particularly those acting on PDE5 and PDE4, demonstrate consistent improvements in key features of MetS (e.g., insulin resistance, dyslipidemia, endothelial dysfunction, and systemic inflammation) through targeted modulation of cAMP and cGMP signaling in vascular and metabolic tissues. Their efficacy profiles, supported by controlled pharmacokinetic parameters, suggest a promising therapeutic role in restoring metabolic and vascular homeostasis. Conversely, non-selective or naturally derived PDE modulators, including caffeine, theobromine, and polyphenolic compounds, exhibit broader systemic effects that integrate metabolic, antioxidant, and anti-inflammatory pathways. While their outcomes appear less predictable and mechanistically defined, these agents contribute to a more holistic understanding of cyclic nucleotide signaling in complex metabolic disorders. Taken together, selective PDE inhibitors provide stronger mechanistic evidence and translational potential, whereas non-selective modulators highlight complementary, lifestyle-associated strategies that may synergize with pharmacological interventions in managing MetS.
Notably, the majority of available RCTs investigated the effect of PDE5 inhibitors in MetS or in overlapping cardiometabolic conditions such as T2DM and erectile dysfunction. These studies consistently demonstrated improvements in endothelial function, insulin sensitivity, and erectile function, supporting the concept of systemic metabolic and vascular benefits mediated via the NO–cGMP–PKG axis. However, most trials prioritized surrogate endpoints rather than long-term cardiometabolic outcomes.
Other PDE-targeting compounds have also been tested, albeit with limited translational relevance to MetS. Cilostazol, a PDE3 inhibitor, was evaluated for effects on adipokines and arterial stiffness, while levosimendan (a PDE3/4 modulator with inotropic effects) was tested in relation to skeletal muscle metabolism. More recently, exploratory observational protocols have focused on PDE3B in metabolic regulation, with ongoing studies aiming to delineate its role in lipolysis and energy balance.
Together, all preclinical and clinical data analyzed and discussed above, suggest that PDE inhibitors, by selectively modulating intracellular cyclic nucleotide signaling, can enhance insulin action, stimulate lipid oxidation, suppress adipogenesis, reduce ectopic lipid deposition, and improve endothelial-metabolic cross-talk. Nevertheless, metabolic outcomes vary by PDE isoform, dosing duration, and tissue context, and some unfavorable effects, such as sildenafil-induced glucose intolerance under obesity, underscore the complexity of chronic modulation. Consequently, large-scale, long-term trials with isoform-selective and tissue-restricted PDE targeting are warranted to establish therapeutic efficacy, tolerability, and personalized applications in metabolic disease.
Collectively, these findings underscore the potential of PDEi as versatile
modulators of metabolic homeostasis, capable of fine-tuning adipocyte
thermogenesis,
FC designed the research. NB, MRA, FS, GD, FGK, MAV, AMI and FC performed literature research and analyzed the data. NB and GD contributed to figure design. MRA and FS participate in the design of the tables. All authors critically reviewed and revised the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This research was funded by the Italian Ministry of Research (grant no. P2022CE79J).
The authors declare no conflict of interest. Given her role as the Guest Editor, Federica Campolo had no involvement in the peer review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process for this article was delegated to Xiaolei Tang.
Although ChatGPT-4o was used as a thesaurus and for minor textual refinement during the drafting of this article, the authors carefully reviewed the entire article and take full responsibility for its content.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.