




1 Department of Surgery, Laboratory of Tumor Immunology and Immunotherapy, Morehouse School of Medicine, Atlanta, GA 30310, USA
Abstract
Immunology advances have increased our understanding of autoimmune, auto-inflammatory, immunodeficiency, infectious, and other immune-mediated inflammatory diseases (IMIDs). Furthermore, evidence is growing for the immune involvement in aging, metabolic and neurodegenerative diseases, and different cancers. However, further research has indicated sex/gender-based immune differences, which further increase higher incidences of various autoimmune diseases (AIDs), such as systemic lupus erythematosus (SLE), myasthenia gravis, and rheumatoid arthritis (RA) in females. On the other hand, reproductive-age females also show a more potent immune response against infections and vaccines than their age-matched males—furthermore, some immune-based therapies, including immune checkpoint inhibitors (ICIs), show gender-based efficacy and adverse events. Metabolic demands are different in males and females. Immune cell function and polarization are also governed by their metabolic reprogramming, called immunometabolism and immunometabolic reprogramming (IR). Therefore, sex/gender-associated immune differences and their involvement in immune-mediated diseases and immune-based therapeutics indicate the demand for gender-based IR studies to increase the efficacy of immune-based precision medicine.
Keywords
- sex/gender
- immunometabolism
- immunometabolic reprograming
- immune cells
- PRRs
The use of immune-based therapies, such as vaccines, immune-checkpoint inhibitors (ICIs), and antibody- and cell-based immune therapies to prevent and treat infectious diseases, cancers, autoimmune and immunodeficiency disorders, and immune-mediated inflammatory diseases (IMIDs) has increased, indicating immunological advances in the disease management. However, along with several other factors, gender/sex is a critical factor for their efficacy and adverse events as sex hormones, along with X chromosome dosage compensation, critically impact immune functioning. The effective immune responses and immune cell polarization to pro-inflammatory, anti-inflammatory, and immunoregulatory phenotypes are critically regulated by the immune cell metabolism, called immunometabolism. The immunometabolism can be categorized into cellular and tissue immunometabolism and systemic immunometabolism. Therefore, gender-based immune differences, including immunometabolism and its reprogramming, must be in mind for immune-based precision medicine.
Immune response has evolved to protect the host from internal and external threats. The immune system is classified into innate and adaptive immunity. The innate immune system is the most primitive form of the immune response, which is non-specific and very robust. The signaling components of innate immunity called pattern recognition receptors (PRRs), such as cytosolic guanosine monophosphate (GMP)-adenosine monophosphate (AMP) synthase (cGAS)-like receptors (cGLRs) comprising vertebrate cGAS/stimulator of interferon gene (STING) signaling pathway are evolutionarily most ancient and have evolved in bacteria as a defense against bacteriophages at least 600 million years ago (MYA) (Fig. 1) [1, 2]. At the same time, the adaptive immunity evolved 500 MYA with the evolution of jawed vertebrates (gnathostomes) [3]. However, now we know that the oldest known animal sex chromosome, called Octopus-Z, has evolved in octopuses 380 MYA (Fig. 1) [4]. Thus, from an evolutionary perspective, the immune system evolved before the evolution of animal sex chromosomes, determining animal sex for sexual reproduction.
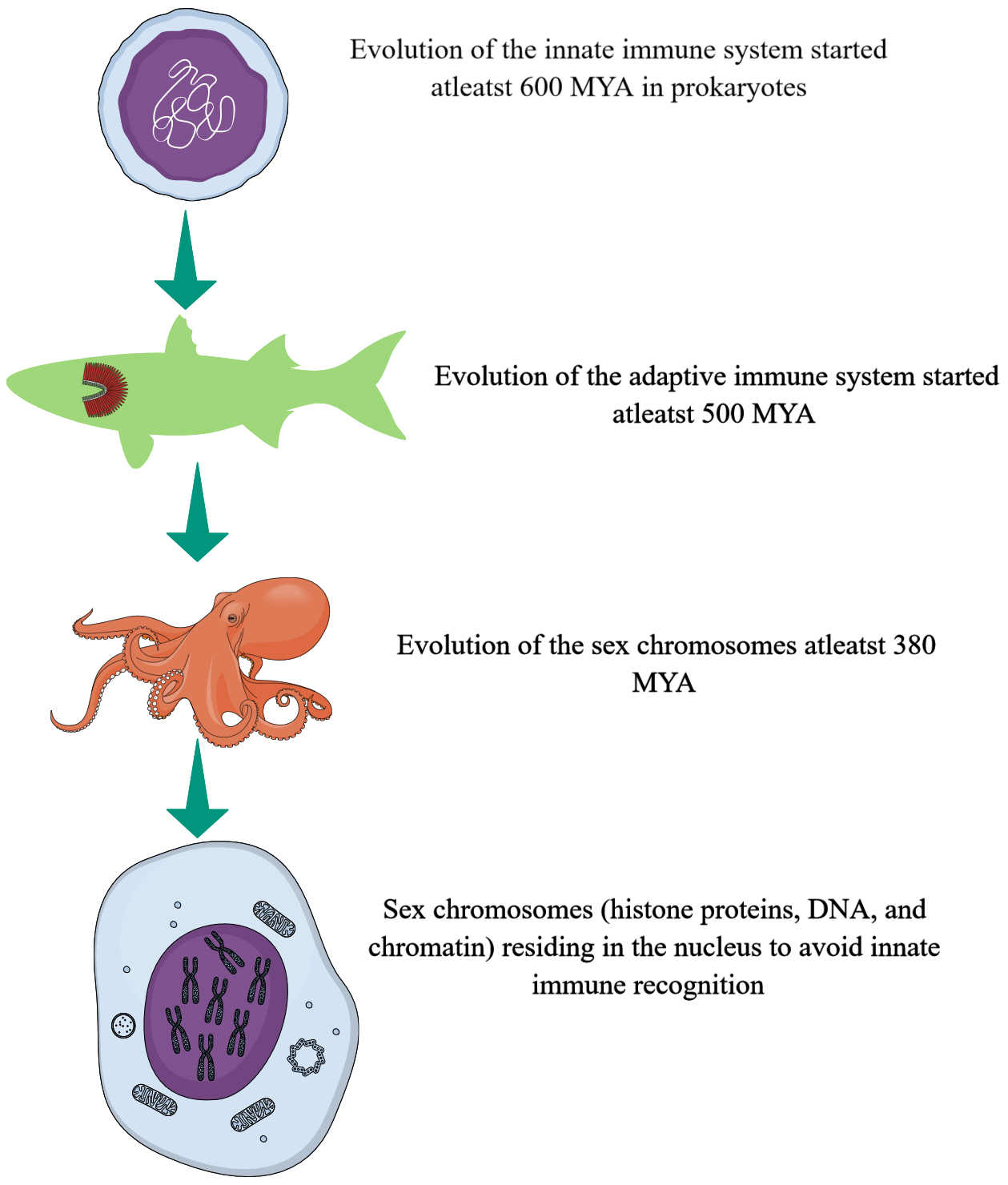 Fig. 1.
Fig. 1.
Time frame of immune system and sex chromosome evolution. The innate immune system (cGLRs) first evolved in prokaryotes (bacteria) at least 600 million years ago (MYA) and the adaptive immune system first evolved in gnathostomes at least 500 MYA. The sex chromosome has evolved at least 380 MYA. cGLRs, cytosolic guanosine monophosphate (GMP)-adenosine monophosphate (AMP) synthase (cGAS)-like receptors. Created with mindthegraph.com.
Furthermore, the localization of sex and other somatic chromosomes inside the nucleus prevents their immune recognition. However, genetic stress seen during infections and other inflammatory insults releases free DNA into the cytosol, which initiates a PRR-dependent pro-inflammatory immune response [5]. Thus, the location of the sex chromosome (inside the nucleus, which provides an immunologically inert site for intact DNA and chromosomes) indicates that the evolution of sex might have required an immune escape. We need further studies to investigate the evolutionary relationship between sex, sex chromosomes, and immunity. For example, cGAS, a potent PRR for double-stranded DNA (dsDNA) comprising cGLRs, is present in the nucleus but remains inactive due to its binding to nucleosomes and continuous competition with chromatin architectural components [6, 7, 8].
Furthermore, the ubiquitin-proteasome system (UPS) degrades nuclear cGAS in
cells undergoing cell cycling signaling events SplA/Ryanodine Receptor Domain And SOCS Box Containing 3 (SPSB3) as the cGAS-targeting
substrate receptor associates with the cullin—RING ubiquitin ligase 5 (CRL5)
complex to ligate ubiquitin onto nuclear cGAS [9]. Notably, free or soluble cGAS
can only recognize the virus and host-derived naked DNA to initiate the
pro-inflammatory innate immune response. Interestingly, the soluble nuclear cGAS
(cGAS not attached to the nucleosome or the chromatin) generated during viral
infections, such as herpes simplex virus-1 (HSV-1), recognizes viral DNA, generates
cyclic guanosine monophosphate (GMP)- adenosine monophosphate (AMP) (cGAMP), and
type-1interferon (IFN)-dependent pro-inflammatory immune response [10]. Hence,
the nucleus serves as a better place for chromosomes, including the sex
chromosome, for their cellular localization, which is an immunologically inert
location. However, evolutionarily, the cGAS system evolved first in the bacteria
as cGAS-DncV (dinucleotide cyclase in Vibrio cholerae)-like
nucleotidyltransferase (cyclic dinucleotidyltransferase (CD-NTase), a
structurally and evolutionarily distinct subset of the DNA polymerase
Bacteria have circular genomes comprising of naked DNA supercoiled DNA, which is present in nucleoids (a false nucleus-like structure), and their CD-NTases are encoded in conserved operons in mobile genetic elements (MGEs), such as plasmids [11]. For example, genes encoding identical CD-NTases are often found in specific strains of unrelated bacterial species, reflecting that these genes are members of the “mobilome” [11]. The evolution of eukaryotic linear chromosomes from the prokaryotic circular genome has yet to be well understood [13]. Still, their localization in the eukaryotic true nucleus and evolution of escape mechanisms from the nuclear cGAS recognition, such as the inactivation of nuclear cGAS by several mechanisms and formation of actual chromosomes (DNA + histone bodies forming nucleosomes + chromatin fibers), decreased the chances of host-derived naked DNA in the nucleus, which the free soluble cGAS in the nucleus could recognize. Hence, the nucleus served as a perfect site for the location of sex and autosomal chromosomes to escape from the host’s immune response.
Advances in immune and sex-based studies have further indicated the immune
response variation in males and females against different infections and
incidence of autoimmunity [14, 15]. Recent coronavirus disease-2019 (COVID-19)
pandemic research data has further indicated that females had a lower incidence
of severe COVID-19 than age-matched males due to differences in immune response
[16]. However, females have a higher incidence of certain autoimmune diseases
(AIDs), such as systemic lupus erythematosus (SLE), rheumatoid arthritis (RA),
and Sjogren’s syndrome (SS). Furthermore, recent trends in cancer incidences have
also indicated an increase in different cancers (lung, breast, and colorectal) in
reproductive-age women (
Every living cell requires energy for its growth, survival, division, and function to maintain homeostasis. The energy generation and expenditure by the cell or host to maintain homeostasis is governed by metabolic processes or metabolism. However, for several years, metabolism was taught in the context of carbohydrate, lipid, and protein metabolism as general, not cell-type specific. Immunological advances have now suggested that immune cell metabolic processes are critical for maintaining immune homeostasis, leading to the development of immunometabolism, and further exploration is needed to advance immune-based precision medicine. Different PRRs’ activation affects IR to control pro-inflammatory immune response differently [21]. Furthermore, the immunometabolism research is complete, undertaking the gender-based immune cell-specific immune response. For example, hormonal and metabolic energy differences in reproductive-age women and men should be critically considered when targeting IR. Vaccination and natural infections have indicated higher protective pro-inflammatory immune responses in reproductive-age women than in age-matched men. Gender-based immunological laboratory data has also supported these findings. Promoters of several innate immunity genes have androgen and estrogen response elements (AREs and EREs), indicating their direct impact on the innate immune response depending on gender. Activation of the innate immune response is the primary determinant of generating the adaptive immune response.
For example, generally, the lipopolysaccharide (LPS) or interferon-
Further, females respond differently to stress due to different organization and functional effects, including the release of neurotransmitters exerted by sex hormones on human brains [28, 29]. For example, females show sluggish cortisol feedback on the brain and less or delayed containment of the stress response than males, which can affect the immune system/response differently (Fig. 2) [28, 30, 31]. Whereas, once puberty has completed, male sex hormone levels in healthy men do not fluctuate regularly, but female sex hormone levels fluctuate periodically over a much more extensive range than men (Fig. 2). Furthermore acute immune response to stress is well documented in adults [32]. Moreover, there we go for neuroimmunology, which can lead to endoneuroimmunology (endocrine control of neuroimmunology) to study the impact of sex hormones on neuroimmunology. Therefore, pre-existing stress that can be anxiety disorders or disease (cancer or infection)-associated stress can impact energy metabolism, including immunometabolism or IR, differently in females than men (Fig. 2). For example, immunometabolism or IR is critical in cancer pathogenesis and tumor immunity or immune microenvironment [33]. Furthermore, a gender-based mice study has indicated the protective action of invariant natural killer T (iNKT) cells in male mice during high fat and choline-deficient (HFCD) diet-induced steatohepatitis as compared to females [34]. iNKT cells of male and female mice exhibit noteworthy sex/gender-associated disparities as indicated by the production of pro-inflammatory cytokines, iNKT cell gene expression, and immune cell profiles impacting the steatohepatitis-associated immune response. This further suggests that gender-based hormonal differences impact immune cell functioning and their IR, which needs further exploration to delineate the mechanisms of differential outcomes of different immune-based therapies in two genders.
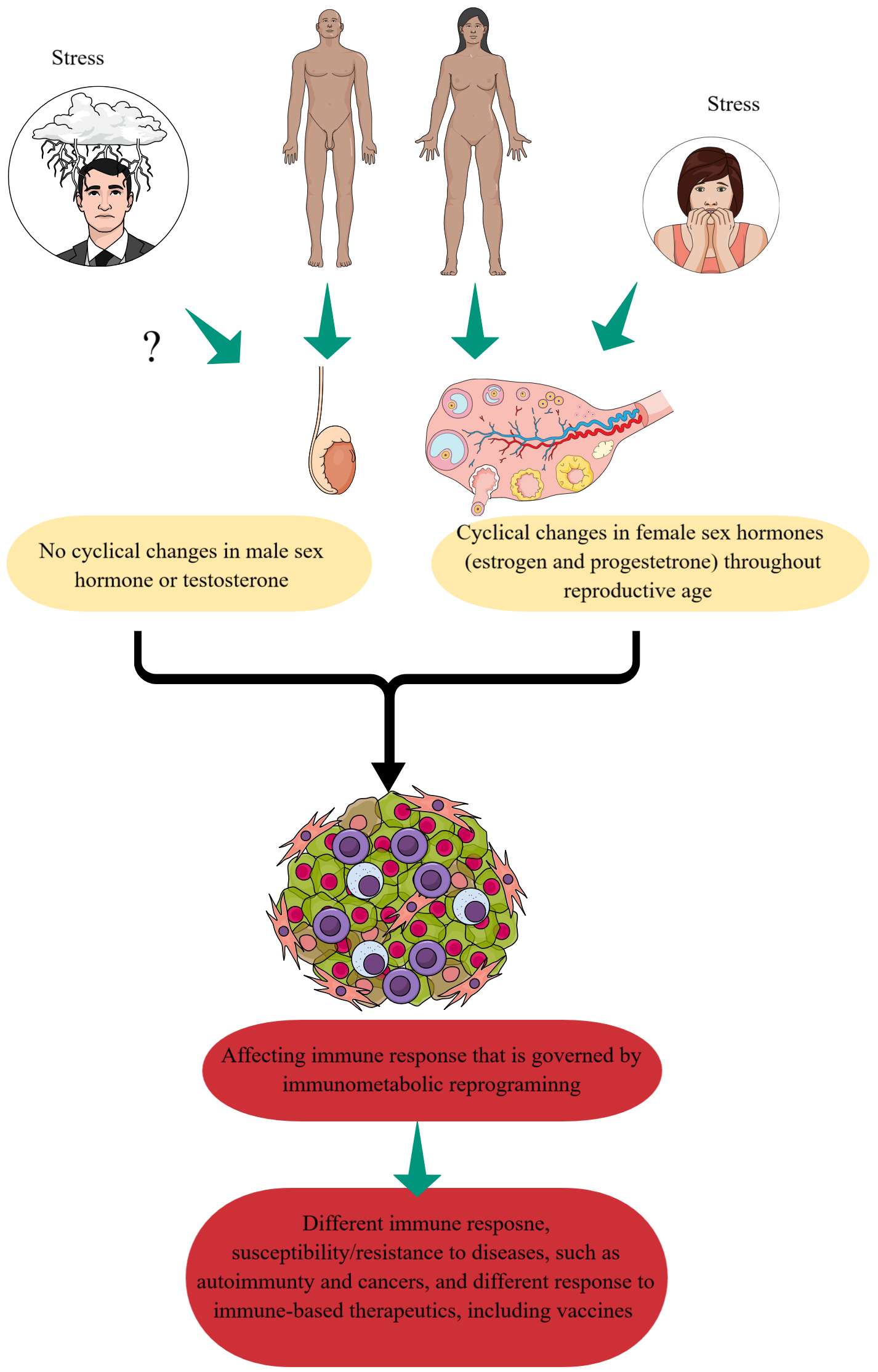 Fig. 2.
Fig. 2.
Adult male and female response differently to stress that impacts their immune response. Males and females respond differently to stress and cyclical sex hormones fluctuations are present in human females throughout their reproductive period. Chronic stress may dysregulate their sex hormones, which impact immune cell functioning governed by immunometabolic reprograming (IR). This may affect their immune response to different inflammatory and infectious conditions and vaccinations. Created with mindthegraph.com.
Furthermore, the clinical trial of the drug lecanemab (a high affinity humanized
Immunoglobulin G1 (IgG1) monoclonal antibody against amyloid-beta (A
Evolutionary forces first created the immunologically inert environment for the
immune escape of the host’s genetic material (DNA) in the form of chromosomes
localized in the nucleus as cGLRs, such as cGAS (evolved more than six hundred
MYA before the evolution of sex chromosomes), recognize naked DNA released into
the cytosol as a result of nuclear and genotoxic damage. Thus, DNA in the intact
chromosome and proteins comprising nucleosomes escape from the innate immune
response in the nucleus due to their intact structure and inactive cGAS.
Furthermore, evidence has emerged indicating differences in the immune responses
between the two sexes to fight against infections, responding to immune-based
therapies, and higher incidences of AIDs and increased adverse events to
immune-based therapies in reproductive age females than males. Therefore, we must
recognize gender-based immune response and IR to design precision medicine for
immunological disorders and IMIDs. For example, females generate profound
inflammatory immune responses to antigens, infections, and vaccines where immune
stimulation is needed. Still, they also exhibit more severe immune-mediated
adverse events than men [41, 42]. Nevertheless, they are less potent than men in
response to immunosuppressive immunotherapeutics such as anti-tumor necrosis factor-alpha (TNF-
Therefore, studies on gender-based cellular and tissue immunometabolism are urgently needed to design better immunotherapies and lower the gender disparity in immune-mediated diseases and therapies. Furthermore, immunometabolism research has seen significant advances in the last ten years that further need the inclusion of sex/gender to demystify hidden mechanisms of diseases and different outcomes of immune-based therapies in two sexes.
VK was responsible for the conception of ideas presented, writing, and the entire preparation of this manuscript.
Not applicable.
Not applicable.
This research received no external funding.
Given his role as the Guest Editor and Editorial Board member, Vijay Kumar had no involvement in the peer-review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process for this article was delegated to Graham Pawelec.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.