














1 Department of Food Biotechnology and Microbiology, Institute of Food Sciences, Warsaw University of Life Sciences – SGGW, 02-776 Warsaw, Poland
Abstract
This study investigated the selenium-binding capacity of the biomass of two yeast strains, Saccharomyces cerevisiae American Type Culture Collection (ATCC) 7090 and Rhodotorula glutinis CCY 20-2-26.
The studies carried out methods of bioaccumulation by yeast biomass. Inorganic selenium was added to the culture media as an aqueous solution of Na2SeO3 at concentrations ranging from 0 to 40 mg Se4+/L.
The addition of selenium at concentrations >0.5 mg/L significantly reduced biomass yield compared with the control in the case of S. cerevisiae. A significant reduction in the biomass of R. glutinis was observed only at selenium doses >30 mg/L. The study found that for S. cerevisiae, cultivation should occur for 24 h in a medium with an initial selenium concentration of 20 mg/L to achieve the most efficient selenium accumulation by the yeast biomass. Under these conditions, the yeast could accumulate 4.27 mg Se4+/g. For the red yeast R. glutinis, optimal selenium binding conditions were achieved by cultivating for 48 h in a medium with an initial selenium ion concentration of 40 mg/L. This yeast strain was more resistant to high selenium doses, accumulating 7.53 mg Se4+/L at the highest tested dose (40 mg Se4+/L). Selenium supplementation of the medium from 20 mg Se4+/L and cultivation for 72 h caused significant changes in the morphology of S. cerevisiae cells (e.g., increased surface area compared with the control). Selenium doses of 20–40 mg/L after 48 h of cultivation significantly reduced the surface area compared with the control results for R. glutinis cells.
Selenium significantly impacted carotenoid pigment production, with levels decreasing as the selenium concentration in the medium increased. Furthermore, selenium in the tested concentration range increased protein content in the cellular biomass but did not affect intracellular lipid production.
Keywords
- selenium
- Saccharomyces cerevisiae
- Rhodotorula glutinis
- selenium accumulation
- yeast
- supplementation
Selenium is one of the most important and essential trace elements for human and animal health [1, 2]. It plays a crucial role in the functioning of selenoenzymes in the human body, protecting against cardiovascular diseases, infertility and diabetes. It also stimulates the immune system and increases the synthesis of antibodies (IgG and IgM) [3]. Low selenium levels in the soil affect its content in plants used for animal feed and food production, negatively impacting animal and human health [4]. Selenium intake in the diet is insufficient in many parts of the world, such as Europe, New Zealand and China [5, 6]. As a result, food companies have produced selenium-fortified foods and supplements to increase selenium intake levels in humans [7, 8]. An important aspect of selenium supplementation is its bioavailability. Organic selenium compounds are absorbed at about 90–95%, with a human utilization rate of about 75–90% [2, 9]. Thus, selenium-enriched yeast, which biotransforms inorganic selenium forms (SeO32- and SeO42-) into organic forms such as selenomethionine, selenocysteine and selenomethylselenocysteine, are a good source of selenium [10]. Owing to ease of cultivation, non-toxicity and non-pathogenicity, yeast is an excellent tool in biological research [11], including selenium accumulation analyses. Saccharomyces cerevisiae yeast has been and continues to be the main object of research on the biotransformation of inorganic selenium forms into selenomethionine. Nevertheless, researchers are exploring the use of other yeast species for this biotransformation, including Yarrowia lipolytica, Candida utilis and Rhodotorula glutinis [12, 13]. Recent literature highlights increasing research on another yeast strain from the Saccharomyces genus, namely Saccharomyces boulardii, a probiotic microorganism [10, 14].
Research on selenium accumulation by yeast cells remains relevant and provides numerous economic and environmental benefits [2], as each year new publications illustrate the diverse positive impacts of selenium in medicine and industry. In their study, Zhang et al. [4] provided evidence that selenium-enriched yeast added to cereal feed (as an external seleniufm source) can significantly increase the selenium content in chicken eggs (particularly in albumins and yolk globulins). A study presented by Ferrari et al. [2] showed that adding selenium-enriched yeast or selenium nanoparticles to feed resulted in significant improvements in broiler weight gain, increased fish growth efficiency and improved internal organ condition (e.g., intestines). It also led to better rumen fermentation in sheep and better feed utilization.
In medicine, S. cerevisiae yeast is an excellent model for studying cancer metabolism owing to observed similarities in glucose metabolism between yeast and cancer cells [1]. Studies on human cancer report the chemoprotective effects of routinely taken selenium yeast [15, 16], further motivating researchers to continue investigations in this area. Increasing studies are emerging on the prophylactic and therapeutic properties of selenium in combating COVID-19. It has been reported that selenium compounds can halt the progression of lung damage and inhibit systemic complications [17].
The global nanotechnology market has gained significant attention in various fields of science and technology, such as drug delivery systems and materials with antimicrobial and antiparasitic properties [18]. Recently, certain yeast strains, including Yarrowia [19], Saccharomyces [20], Rhodotorula [21] and Pichia [22], have been proposed as new biocatalysts for selenium nanoparticle synthesis (SeNP - selenium nanoparticles). Ashengroph and Tozandehjani [18] described the biotransformation of selenite oxyanion into selenium nanoparticles (both intracellular and extracellular) by the Rhodotorula mucilaginosa R-8441 strain as an alternative to physicochemical synthesis. Field emission scanning electron microscopy (FESEM) results indicated that as selenite oxyanion concentrations increased, particle sizes also increased. Selenium doses ranging from 1 to 4 mM caused the formation of spherical nanoparticles, and those above 5 mM created rod-shaped structures.
This study aimed to determine the impact of inorganic selenium on the physiological activity of S. cerevisiae American Type Culture Collection (ATCC) 7090 and R. glutinis CCY 20-2-26 yeast cells. Changes in biomass yield of the tested yeast strains, the degree of selenium accumulation by yeast cells and the protein and lipid content in the biomass, as well as the carotenoid content and fractions (only for Rhodotorula glutinis CCY 20-2-26), were examined.
Two yeast strains, Saccharomyces cerevisiae ATCC 7090 and Rhodotorula glutinis CCY 20-2-26, were used in this study. The strains were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA) and the Culture Collection of Yeasts (CCY, Bratislava, Slovakia). The strains were stored at 4 °C on solid yeast peptone dextrose (YPD) medium (BLT, Lodz, Poland).
The yeast inoculum was prepared by inoculating 100 mL of liquid YPD medium (2%
glucose, 2% peptone, 1% yeast extract, pH 4.8, BLT, Lodz, Poland) with a 24-h culture of yeast
strain from a Petri dish using an inoculation loop. To determine the degree of
selenium binding by the yeast, the experimental YPD medium with an initial pH of
4.8 was supplemented with an aqueous solution of sodium selenite (IV) (Na2SeO3, Sigma-Aldrich, Warsaw, Poland). All
analyses were performed five times. The aqueous selenium solution was prepared by
dissolving 0.219 g Na2SeO3 in 100 mL of deionized water (final
concentration of Se4+ ions was 1000 mg/L). Both the YPD medium and the
aqueous Na2SeO3 solution were sterilized at 121 °C for 15
minutes (Systec D-45 autoclave, De Ville, Warsaw, Poland). Sterile selenium salt
solution was added to the sterile YPD medium in volumes to achieve final selenium
concentrations in the experimental medium ranging from 0.5 to 40 mg Se4+/L.
The last step involved adding the yeast inoculum (2.3
The biomass yield of yeast cells was determined after centrifugation (Eppendorf 5810 Centrifuge, Hamburg, Germany) (3500 rpm; 10 min) of 30 mL of post-culture fluid in dry, pre-weighed 50 mL Falcon tubes. The supernatant was decanted for selenium content determination, and the sediment was dried (SML Zalmed Dryer, Warsaw, Poland) (80 °C) to a constant weight. Biomass yield was recalculated per 1 L of medium and expressed in grams of dry weight (gd.w./L).
The spectrophotometric method used to determine selenium is based on measuring the absorbance (wavelength 546 nm) of the coloured compound Variamine Blue (VB, Sigma-Aldrich, Warsaw, Poland). The intensity of VB staining depends on the number of its molecules reacting, initially triggered by selenium ions [23]. The method involved taking 1 mL of the supernatant into 50 mL volumetric flasks containing 5 mL of a 2% KI solution (Merck, Warsaw, Poland) and 5 mL of 2 M HCl (Merck, Warsaw, Poland), followed by gentle mixing to release iodine. After iodine release, 2.5 mL of 0.05% VB reagent and 10 mL of 1M sodium acetate were added to each flask. The volume was then made up to 50 mL with distilled water, and the absorbance was measured (BIO-RAD SmartSpec 3000 spectrophotometer, Warsaw, Poland) at 546 nm against a control sample. The average absorbance results were converted to selenium content in the sample using the regression equation (y = 0.0527x – 0.009) (Fig. 1). The obtained value was corrected considering the culture volume. The difference between the initial and final selenium concentration in the medium indicated the amount of selenium accumulated by the yeast cells. The selenium content accumulated per gram of dry weight of the tested yeast strains was calculated based on the biomass yield.
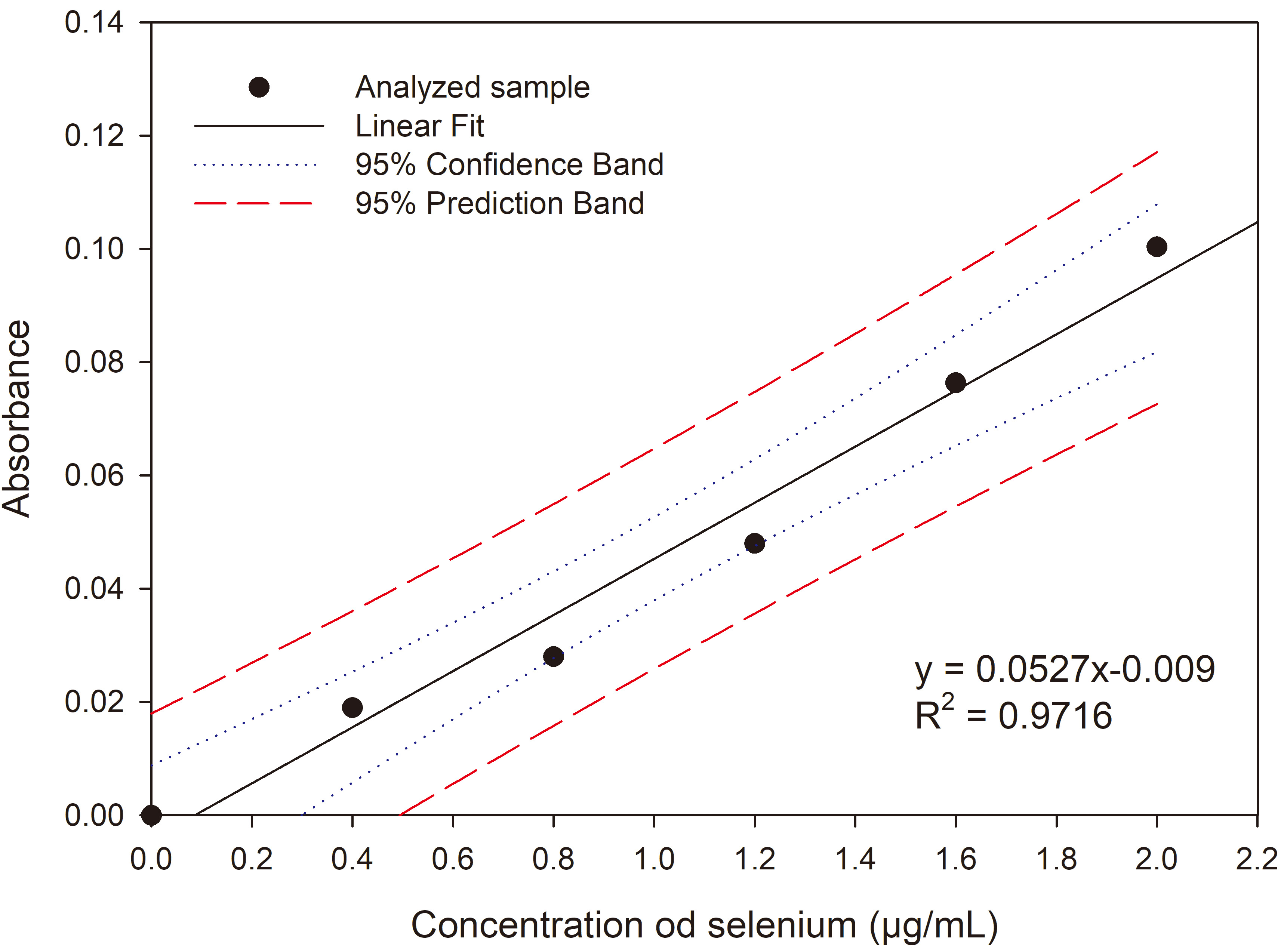 Fig. 1.
Fig. 1.
Calibration curve of absorbance for sodium selenite (IV).
The morphology of the studied microorganisms was observed using an optical microscope (OPTA-TECH, Warsaw, Poland). Measurements of yeast cell length and width were recorded using Opta View 7 software (OPTA-TECH, Warsaw, Poland).
The liquid yeast biomass (1.5 mL) was immersed in 2 mL of dimethyl
sulfoxide (DMSO, Sigma-Aldrich, Warsaw, Poland), to which 0.5 g of glass beads approximately 500 µm in
diameter were added and mixed for 1 h at 70 rpm (Rotator MultiBio RS-24, Biosan,
Riga, Latvia). Then, 2 mL of acetone (Sigma-Aldrich, Warsaw, Poland), 2 mL of petroleum ether (with 0.25% BHT)(Sigma-Aldrich, Warsaw, Poland)
and 2 mL of 20% sodium chloride (Sigma-Aldrich, Warsaw, Poland) were added to each tube, and the samples were
mixed again under the same conditions. To separate the phases, the tubes were
centrifuged at 20 °C at 3000 rpm for 5 minutes (Eppendorf 5810
Centrifuge, Hamburg, Germany). The separated ether phase containing the
carotenoids was transferred to a cuvette, and the absorbance was measured at
To identify the extracted carotenoids, after evaporating the petroleum ether
under nitrogen atmosphere, 1.2 mL of the HPLC mixture consisting of acetonitrile,
isopropanol and ethyl acetate in a ratio of 4:4:2 was added. The mixture was
filtered through a 0.45 µm pore size nylon filter and analysed
chromatographically on a C18 analytical column (Bionacom, 250 mm
The analysis was conducted using the Kjeldahl method to determine the total protein content in the yeast cell biomass. Additionally, changes in protein content in the experimental media were assessed compared with the control sample of the studied yeast strains. One hundred mg of dry yeast cell substance was weighed and mineralized (Buchi Digestion Unit K-435, Germany) in 96% sulfuric acid (VI) in the presence of a catalyst (Kjeltabs CT/3.5 tablets, FOSS Analytical, Hillerød, Denmark). After the process was completed, the solution was cooled, then distilled (KjelFlex K-360, BÜCHI Labortechnik AG, Munich, Germany) and automatically titrated (TitroLine 5000, SI Analytics, SI Analytics GmbH, Mainz, Germany) using 0.1 M acid (Kjeltec 2100, Foos). The nitrogen content was converted to total protein content using the conversion factor 6.25.
To determine the lipid content in the dry yeast biomass, the Bligh and Dyer method was used [25]. To disintegrate the cell wall, 10 mL of 1M HCl was added to 200 mg of dry yeast biomass and incubated for 2 h at 60 °C (Water Bath SBS-TWB-2004, Steinberg Systems, Berlin, Germany). After cooling the samples, lipid extraction was performed using a chloroform (Sigma-Aldrich, Warsaw, Poland) and methanol (Sigma-Aldrich, Warsaw, Poland) mixture (1:1 ratio). After 20 minutes of intense shaking (Rotator MultiBio RS-24, Biosan, Riga, Latvia), 5 mL of a 20% sodium chloride solution (Sigma-Aldrich, Warsaw, Poland) was added to facilitate phase separation. The samples were then centrifuged at 3500 rpm for 10 min (Eppendorf 5810 Centrifuge, Hamburg, Germany). The chloroform phase was transferred to a pre-weighed tube. The chloroform was evaporated under a nitrogen atmosphere (Reacti-therm III #TS-18823, Thermo Fisher Scientific, Waltham, MA, USA), and the fat content was determined gravimetrically.
The obtained research results were analysed using analysis of variance in the
Statistica 13.3 program (StatSoft, Warsaw, Poland). The significance of differences between the means in the
respective groups was verified using Tukey’s honestly significant difference (HSD) test at a significance level of
The growth of S. cerevisiae ATCC 7090, expressed as biomass yield, was dependent on the selenium concentration in the growth medium. After 24 h of cultivation in the control medium, this strain produced 9.67 g/L, and in subsequent days, the values were not statistically significant in the same control medium (Fig. 2). Adding selenium to the medium at a dose of 0.5 mg Se4+/L significantly reduced the biomass yield compared with the control to 6.32 g/L after 24 h of cultivation, and as in the control medium, its values did not change significantly in subsequent days. Similar biomass yield values were noted for selenium doses of 1, 2 and 5 mg Se4+/L. When selenium was added at 10 and 20 mg Se4+/L, the biomass yields after 24 h of cultivation were 5.67 and 4.57 g/L, respectively. The highest doses of selenium (30 and 40 mg Se4+/L) resulted in the greatest inhibition of yeast growth (2.88–3.70 g/L). Additionally, in cultures with selenium doses of 30 and 40 mg/L, the formation of a red precipitate (elemental selenium) was observed at the bottom of the flasks.
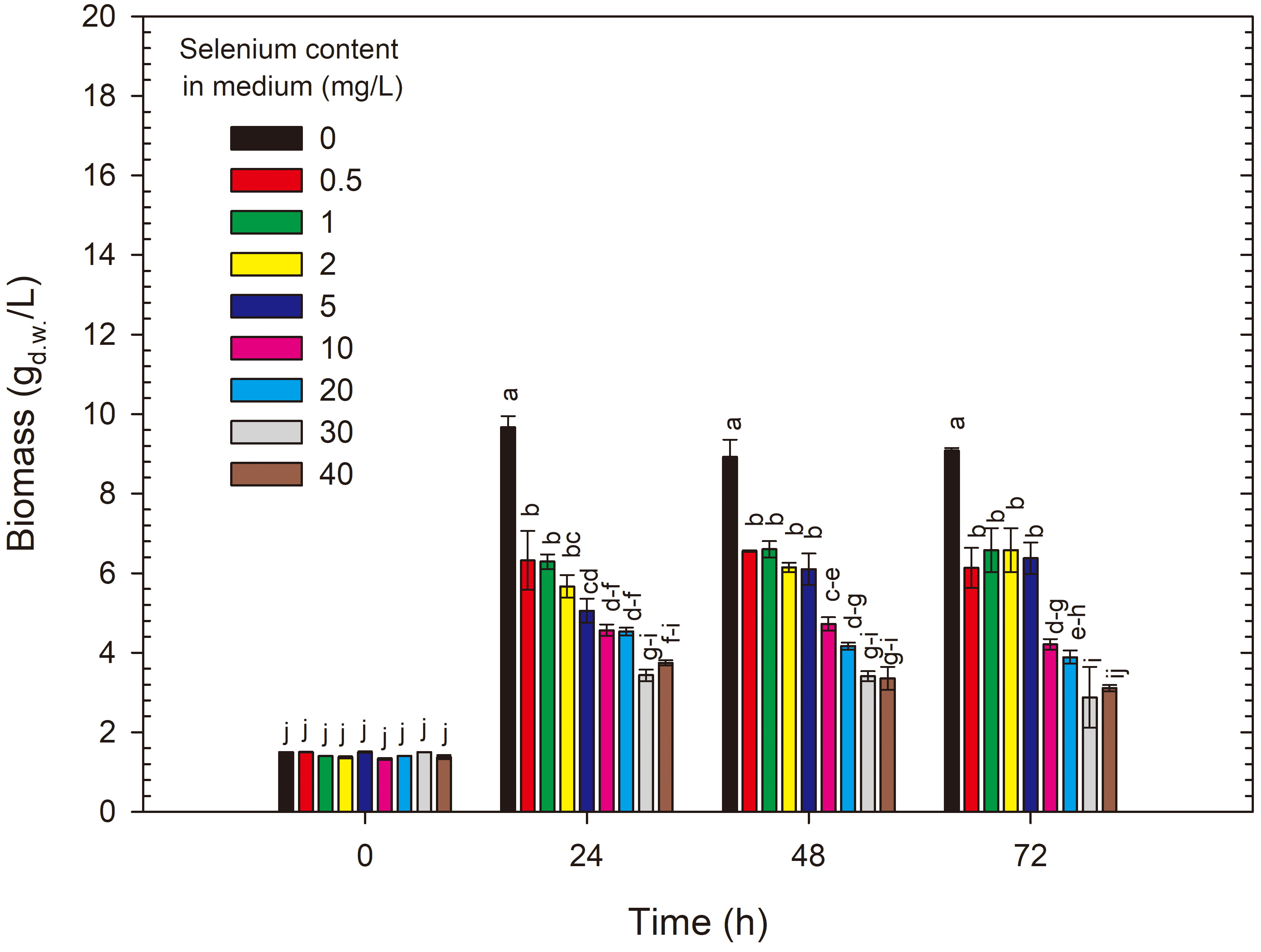 Fig. 2.
Fig. 2.
Changes in yeast biomass yield during 72 h cultivation of S. cerevisiae in control and experimental media enriched with selenium. a-j Means with the same letter did not differ significantly.
The growth results of R. glutinis CCY 20-2-26 differed from those obtained for S. cerevisiae ATCC 7090 (Fig. 3). After 24 h of cultivation, the biomass yield in the control medium without selenium reached 16.57 g/L. This is 71% higher than that obtained for S. cerevisiae ATCC 7090 after the same cultivation time. The biomass yield remained at a similar level throughout the cultivation process. The addition of selenium to the medium in amounts ranging from 0.5 to 5 mg Se4+/L did not significantly reduce biomass yields. The growth limitation compared with the control was noted for cultures conducted in media with 20, 30 and 40 mg Se4+/L, with the greatest limitation observed for the highest doses. After 72 h of cultivation in media with initial selenium concentrations of 20–40 mg Se4+/L, the biomass yields of R. glutinis CCY 20-2-26 ranged from 5.40 to 5.80 g/L. In cultures with the highest selenium doses, the yeast biomass lost its typical red colour. The presence of high selenium doses may cause changes in cellular metabolism. Enzymes and metabolic pathways responsible for pigment synthesis (desaturases) may be inhibited or altered by selenium, leading to reduced pigment production [26]. Selenium can directly react with pigments present in yeast cells. Pigments like carotenoids, which can give yeast a red colour, may undergo chemical modifications in the presence of high selenium concentrations, leading to changes in their colour properties. Similar results were observed by Kieliszek et al. [21] studying the effect of selenium on carotenoid production by the yeast R. mucilaginosa MK1, which reduced carotenoid concentrations in yeast cells.
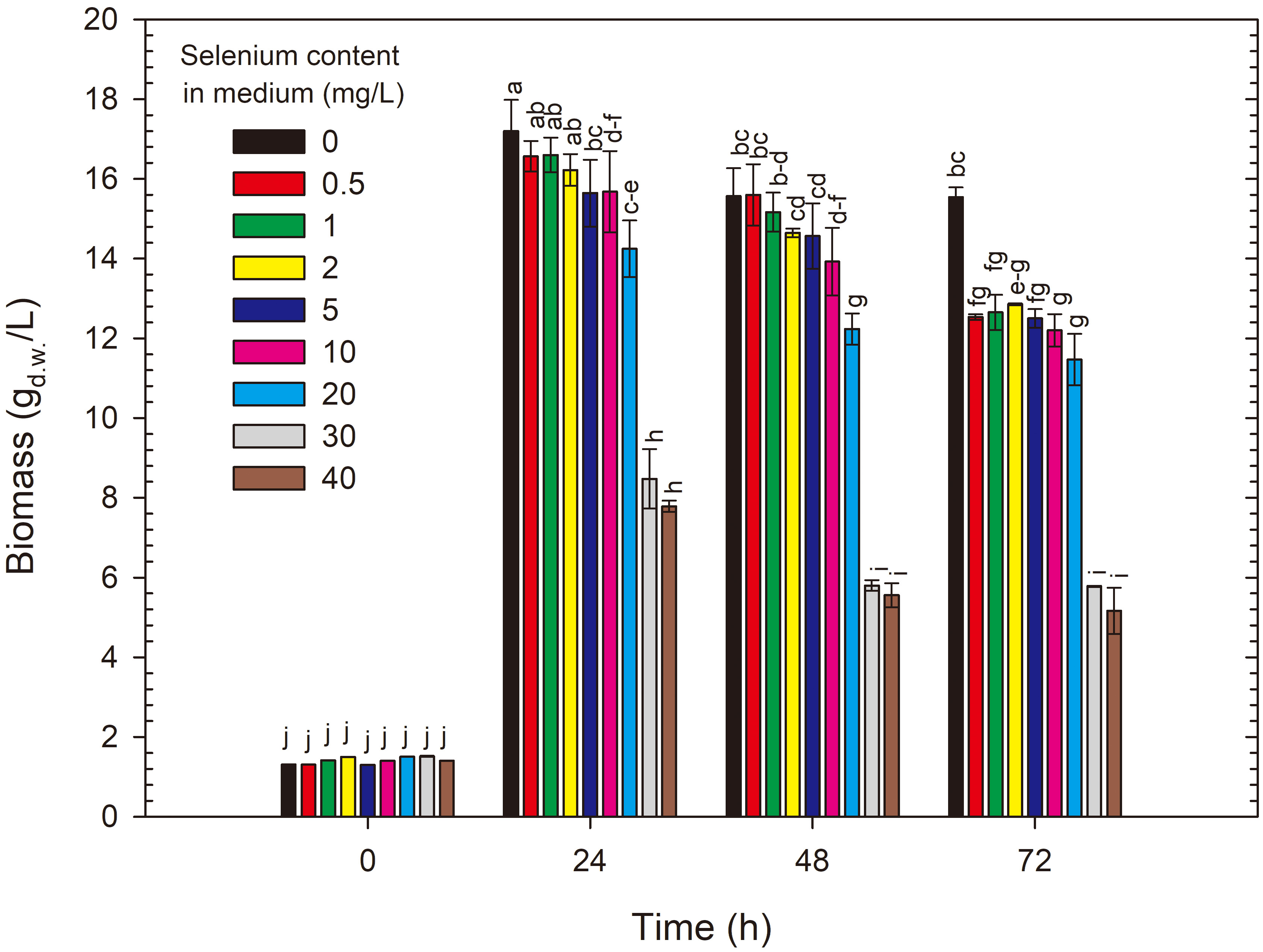 Fig. 3.
Fig. 3.
Changes in yeast biomass yield during 72 h cultivation of R. glutinis in control and experimental media enriched with selenium. a-j Means with the same letter did not differ significantly.
The biomass yield results of the two studied yeast strains show significant differences in cell reactions to selenium-induced stress in the medium. Significant biomass yield reduction compared with the control in S. cerevisiae ATCC 7090 cultivation was caused by the smallest selenium doses, while in R. glutinis CCY 20-2-26, only selenium addition above 20 mg Se4+/L resulted in biomass yield reduction compared with the control at each time point. The results of this stage of the study demonstrated greater selenium tolerance in red yeast compared with the widely studied yeast S. cerevisiae ATCC 7090.
In the study conducted by Kieliszek et al. [23], the selenium binding ability of the biomass of S. cerevisiae ATCC MYA-2200 and C. utilis ATCC 9950 was assessed. Sodium selenite (IV) salts were used in concentrations of 10, 20, 40 and 60 mg Se4+/L. For S. cerevisiae, a dose of 10 mg Se4+/L after 24 h of cultivation significantly affected the biomass yield, reducing it from 9.80 g/L (control medium) to 6.7 g/L (a reduction of 46%). In this study, for S. cerevisiae ATCC 7090, this value decreased from 9.67 g/L (control) to 5.06 g/L (a reduction of 47.7%) under the same cultivation conditions. Comparing the results obtained in Kieliszek et al. [23] for C. utilis ATCC 9950, they resemble those obtained for R. glutinis CCY 20-2-26. After 24 h of cultivation without selenium, the biomass yield of Candida utilis ATCC 9950 was 16.5 g/L, the same as obtained in the control medium of R. glutinis CCY 20-2-26. Selenium supplementation at a dose of 10 mg Se4+/L reduced the yield of C. utilis ATCC 9950 to 15 g/L. The same dose for R. glutinisCCY 20-2-26 caused an identical biomass yield reduction.
Hyrslova et al. [27] conducted a study on the growth of seven different types of bacteria: Streptococcus, Lactococcus and Enterococcus, using various Na2SeO3 concentrations (5, 10, 30, 50 and 100 mg/L). The growth of all four Streptococcus thermophilus CCDM 144, 570, 561 and 129 strains decreased with increasing selenite concentrations. In contrast to streptococci, selenium presence did not significantly affect the growth of Enterococcus faecium CCDM 922A and Lactococcus lactis subsp. cremoris CCDM 72 and CCDM 73 strains. After 24 h of cultivation, all studied strains were able to convert Na2SeO3 to elemental selenium, manifested by a reddish colour, especially in environments with higher selenite concentrations of 10 mg/L or more (30, 50 and 100 mg/L). These results may indicate similar selenium tolerance in bacteria compared with the S. cerevisiae ATCC 7090 strain.
Yang et al. [28] examined the impact of selenium on biomass production in C. utilis SZU 07-01 yeast. They conducted cultivation in a bioreactor (pH 6.0; 30 °C; 20 h; 200 rpm), with a fermentation medium containing 30 g/L glucose, 8 g/L (NH4)2SO4, 3 g/L KH2PO4 and 0.25 g/L MgSO4. Selenium was added as sodium selenite (Na2SeO3) at a concentration of 15 mg Se4+/L in two variants: (I) by adding selenium at the start of cultivation and (II) by adding selenium after 15 h of cultivation, i.e., in the logarithmic growth phase. Biomass yield following control cultivation reached 13.2 g/L, whereas in variant I, it dropped by approximately 33% to 8.9 g/L. Adding selenium to the 15-h culture did not negatively affect yeast growth (13.1 g/L). It can be concluded that selenium toxicity to yeast cells strongly depends on the growth phase, with cells in the adaptation phase being more sensitive to this micronutrient.
The addition of selenium salts to culture media can affect yeast cells in many ways, leading to the changes in yield discussed above. Potential causes of these effects include selenium toxicity, alterations in yeast metabolism or cell structures and interactions between selenium and other media components, leading to complex effects on eukaryotic cells [29]. Excessive selenium can cause metabolic and structural disturbances in cells, negatively impacting their ability to grow and divide. Selenium can influence changes in yeast cell metabolism by disrupting the redox balance and affecting the expression of genes related to metabolism and growth, which in turn can lead to differences in biomass production and cell shape [30, 31]. The biomass of yeast cells cultivated in selenium-enriched media showed significant differences compared to the biomass from the control media. This phenomenon is likely due to toxic metabolites (hydrogen selenide H2Se; selenodiol H2SeO2) produced during selenium transformation in yeast cells, including potentially toxic elemental red selenium (Se0). The generation of such substances can significantly disturb redox balance and lead to the oxidation of essential cellular components. Cultivating microorganisms in stressful environmental conditions can enhance the expression of genes responsible for carotenoid biosynthesis, as observed in the red yeast R. glutinis CCY 20-2-26 studied [32]. Selenium addition can affect cell structure by modifying the chemical composition of the cell wall or cellular organelles. This, in turn, can affect cell shape and their ability to maintain structural integrity. It is important to conduct thorough research to identify or attempt to identify the specific mechanisms and pathways by which selenium affects different types of yeast cells.
The presence of various elements in the culture medium can lead to changes in microorganism morphology. According to the literature [12, 33], the presence of selenites (IV) and selenates (VI) in the culture environment can cause changes in yeast cell sizes owing to adaptive processes to unfavourable environmental conditions. The results of cell length and width measurements are presented in Tables 1,2. Yeast cells of S. cerevisiae ATCC 7090 cultured in the control medium after 24 h had an average surface area of 30.66 µm2, which slightly decreased to 26.36 and 25.42 µm2 after two additional days of cultivation. Selenium doses from 0.5 to 10 mg/L did not significantly affect the dimensions of S. cerevisiae ATCC 7090 cells. However, adding 20 mg Se4+/L to the medium and cultivating for 72 h resulted in significant changes in the morphology of S. cerevisiae ATCC 7090 cells, increasing the surface area to 41.42 µm2. Similar results were obtained for selenium doses of 30 and 40 mg Se4+/L after 72 h of cultivation.
| Selenium content in the medium (mg/L) | Culturing time (h) | Length (µm2) | Width (µm2) | Cell surface area (µm2) | Cell perimeter (µm2) |
| 0 | 24 | 6.92 bcdef | 5.75 defg | 30.66 defg | 20.04 bcdefg |
| 48 | 6.26 defg | 5.46 defg | 26.36 efg | 18.38 defg | |
| 72 | 5.79 g | 5.60 cdef | 25.42 efg | 18.36 efg | |
| 0.5 | 24 | 6.67 bcdefg | 5.22 efg | 29.18 defg | 19.78 cdefg |
| 48 | 5.89 efg | 4.66 g | 24.92 efg | 18.42 efg | |
| 72 | 6.87 bcdefg | 5.23 fg | 31.80 cdefg | 20.69 bcdef | |
| 1.0 | 24 | 6.37 defg | 5.21 efg | 27.28 efg | 19.09 defg |
| 48 | 6.70 bcdefg | 5.10 fg | 27.38 efg | 19.27 defg | |
| 72 | 6.75 bcdefg | 5.40 defg | 29.20 defg | 19.80 bcdefg | |
| 2.0 | 24 | 6.73 bcdefg | 5.06 fg | 28.82 defg | 19.74 cdefg |
| 48 | 6.62 cdefg | 5.47 defg | 29.18 defg | 19.80 cdefg | |
| 72 | 6.30 defg | 5.03 fg | 27.04 efg | 19.03 defg | |
| 5.0 | 24 | 6.59 cdefg | 5.43 defg | 27.49 efg | 19.29 defg |
| 48 | 6.86 bcdefg | 6.02 cdef | 32.04 cdefg | 20.41 bcdef | |
| 72 | 6.18 defg | 5.62 defg | 26.83 efg | 18.73 defg | |
| 10 | 24 | 5.81 efg | 4.93 fg | 23.34 fg | 16.98 fg |
| 48 | 7.14 abcd | 6.18 abcd | 33.20 bcde | 20.93 bcde | |
| 72 | 6.34 cdefg | 5.75 bcdef | 27.89 efg | 19.27 defg | |
| 20 | 24 | 5.90 fg | 4.97 g | 22.69 g | 17.16 g |
| 48 | 6.51 cdefg | 5.58 defg | 28.52 efg | 19.36 defg | |
| 72 | 7.88 ab | 6.96 abc | 41.42 ab | 22.98 ab | |
| 30 | 24 | 6.63 bcdefg | 5.74 cdef | 29.53 defg | 19.59 bcdefg |
| 48 | 6.49 defg | 5.58 defg | 26.97 fg | 18.76 fg | |
| 72 | 8.52 a | 7.22 a | 47.52 a | 24.88 a | |
| 40 | 24 | 6.87 bcdefg | 6.28 bcdef | 33.31 cdef | 20.72 bcdef |
| 48 | 6.90 bcde | 6.10 abcde | 35.15 bcd | 21.01 defg | |
| 72 | 7.68 abc | 7.10 ab | 43.23 abc | 23.37 bcd |
a-gMeans with the same letter did not differ significantly.
| Selenium content in the medium (mg/L) | Culturing time (h) | Length (µm2) | Width (µm2) | Cell surface area (µm2) | Cell perimeter (µm2) |
| 0 | 24 | 6.90 ab | 4.63 a | 26.23 abc | 19.33 abcde |
| 48 | 6.54 ab | 4.69 a | 27.14 ab | 19.32 ab | |
| 72 | 6.59 abcd | 4.79 a | 27.22 ab | 19.33 abcd | |
| 0.5 | 24 | 6.91 ab | 4.49 ab | 24.96 abc | 19.00 abcde |
| 48 | 6.17 abcd | 4.44 a | 23.68 abc | 18.06 abcde | |
| 72 | 8.65 ab | 4.54 a | 24.61 abc | 18.35 abcde | |
| 1.0 | 24 | 6.46 abcd | 4.40 ab | 24.12 bc | 18.39 abcde |
| 48 | 6.27 abcd | 4.29 ab | 22.94 bc | 17.78 cde | |
| 72 | 5.97 cd | 4.36 ab | 23.06 bcd | 17.76 cde | |
| 2.0 | 24 | 6.37 abcd | 4.35 ab | 23.70 bc | 18.17 abcde |
| 48 | 6.77 a | 4.54 a | 26.67 ab | 19.15 abc | |
| 72 | 6.75 abcd | 4.71 a | 27.05 ab | 19.28 abcde | |
| 5.0 | 24 | 6.49 abcd | 4.60 a | 25.67 abc | 19.02 abcde |
| 48 | 6.09 bcd | 4.25 ab | 22.97 cd | 17.83 def | |
| 72 | 6.47 abcd | 4.54 ab | 25.23 abc | 18.66 abcde | |
| 10 | 24 | 6.92 ab | 4.73 a | 30.08 a | 20.36 a |
| 48 | 6.76 abc | 4.52 ab | 24.38 abc | 18.45 abcde | |
| 72 | 6.38 abcd | 4.25 ab | 23.24 bc | 18.16 bcde | |
| 20 | 24 | 6.28 abcd | 3.94 bc | 21.17 abc | 17.46 cdef |
| 48 | 5.88 abcd | 4.36 ab | 21.25 cd | 17.13 def | |
| 72 | 6.52 abcd | 4.52 a | 24.67 abc | 18.53 abcde | |
| 30 | 24 | 6.59 abcd | 4.34 ab | 24.28 abc | 18.52 abcde |
| 48 | 5.85 abcd | 4.29 ab | 20.31 cd | 16.83 ef | |
| 72 | 6.45 abcd | 4.50 ab | 25.35 abc | 18.71 abcde | |
| 40 | 24 | 6.72 abcd | 4.49 ab | 25.30 abc | 18.92 abcde |
| 48 | 5.65 d | 3.38 c | 16.53 d | 15.40 f | |
| 72 | 6.61 abcd | 4.47 ab | 23.88 abc | 18.17 abcde |
a-fMeans with the same letter did not differ significantly.
Yeast cells of R. glutinis CCY 20-2-26 were smaller than those of
S. cerevisiae ATCC 7090. In the control sample, the surface area of
cells ranged from 26.23 to 27.22 µm2 with no significant
differences observed in each day of cultivation. Like S. cerevisiae ATCC
7090, selenium concentrations in the medium
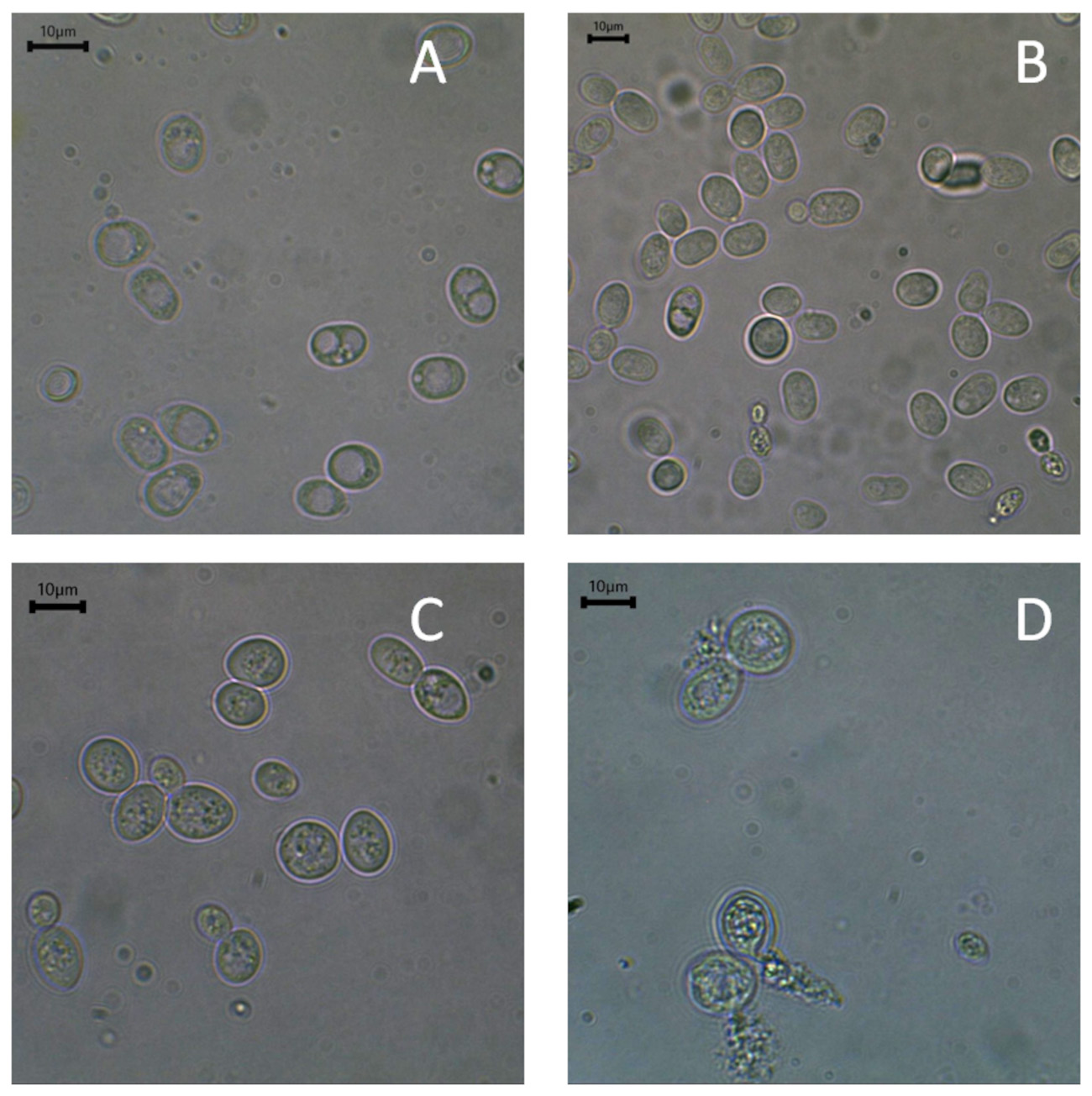 Fig. 4.
Fig. 4.
Microscopic photographs of R. glutinis. (A) Yeast peptone dextrose (YPD) control, and (B) experimental medium enriched with 40 mg Se4+/L and S. cerevisiae (C) YPD control, and (D) experimental medium enriched with 40 mg Se4+/L. Scale bar: 10 µm.
It is worth noting that the tolerance level of cells to selenium ions may vary depending on the type or species of yeast, related to the potential induction of oxidative stress. When yeasts exhibit low tolerance to selenium in the environment, it can lead to changes in the cell wall structure, affecting the overall cell architecture. Research by Kieliszek et al. [12] showed that in the cultivation of C. utilis ATCC 9950 in media containing selenium ions, intracellular structures degraded. This resulted in loss of membrane integrity and cytoplasmic leakage, especially at a selenium concentration of 40 mg Se4+/L after 72 h of cultivation. A significant reduction in yeast cell size due to selenium presence at doses of 30 and 40 mg/L was observed in R. glutinis CCY 20-2-26 yeasts from the first day of cultivation and on each subsequent day. Based on a study by Kieliszek et al. [12], it can be assumed that yeast cell size increase results from selenium accumulation in cellular structures such as vacuoles. The gradual reduction in yeast cell size in the stationary phase can be explained by the progressive detoxification process of selenium in cellular organelles. This mechanism involves the gradual reduction of Se4+ to volatile hydrogen selenide (H2Se) and selenium compounds bound to glutathione. Another possible detoxification method that may lead to yeast cell size reduction is the transport of selenium compounds first to the cytoplasm and then outside the cell. This process may involve transport proteins and Golgi apparatus vesicles [34]. A study by Rajashree and Muthukumar [35] demonstrated that S. cerevisiae NCYC 1026 yeast cells cultivated in selenium-enriched medium showed a size reduction of 0.88 µm compared with control cells not exposed to this element.
Based on the conducted studies, it can be assumed that S. cerevisiae ATCC 7090 and R. glutinis CCY 20-2-26 yeast strains respond differently to the presence of selenium in the environment. S. cerevisiae cells increased their surface area, probably owing to increased selenium compound accumulation in their intracellular structures. An opposite trend was observed for R. glutinis cells, probably owing to an enhanced detoxification system. Interestingly, literature results provide conflicting information about yeast cell morphology changes due to selenium ions in the culture medium, making this topic highly relevant [21, 35]. More research results on various yeast strains are needed.
Selenium accumulation in yeast cells depended on the selenium concentration in
the culture medium. The selenium content in S. cerevisiae ATCC 7090
yeast biomass after the first day of cultivation was lowest in the medium with
the minimum selenium dose (0.5 mg Se4+/L) compared with the other selenium
doses, amounting to 0.021 mg Se4+/g (Fig. 5). Adding selenium to the medium
at a dose of 10 mg Se4+/L resulted in yeast accumulating 1.88 mg
Se4+/g. A significant increase in selenium content in S. cerevisiae
ATCC 7090 cell biomass after the first 24 h of experimental cultivation was
obtained in the medium with an initial selenium concentration of 20 mg/L,
reaching 4.27 mg/g compared with media with selenium content
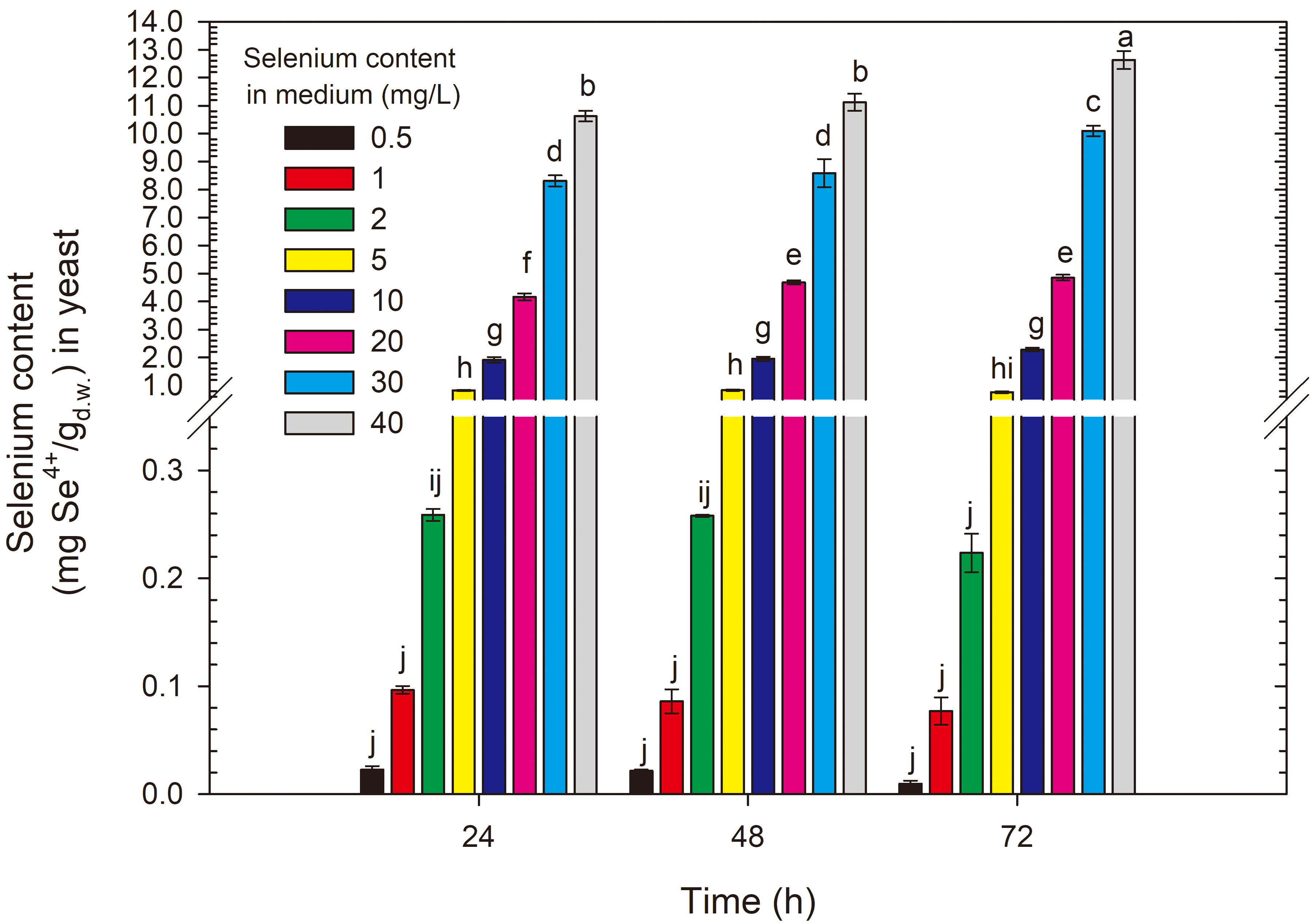 Fig. 5.
Fig. 5.
Selenium content in S. cerevisiae yeast cells grown in medium with different selenium additions in three time units. a-jMeans with the same letter did not differ significantly.
For the R. glutinis CCY 20-2-26 yeast strain, after the first 24 h of
cultivation, selenium concentrations of 0.5–5 mg/L resulted in selenium
accumulation in the biomass of the red yeast at a level of 0.01–0.29 mg
Se4+/g (Fig. 6). An initial selenium concentration of 10 mg/L led to the
binding of this element by the yeast cells in the amount of 0.64 mg/g, and
doubling the selenium addition to the experimental medium significantly increased
selenium accumulation (compared with the 10 mg/L dose) to 1.4 mg/g (for 20 mg/L).
Further increases in selenium concentration in the medium resulted in a
proportional and significant increase of this element in the yeast cell biomass
compared with the results for media with selenium content
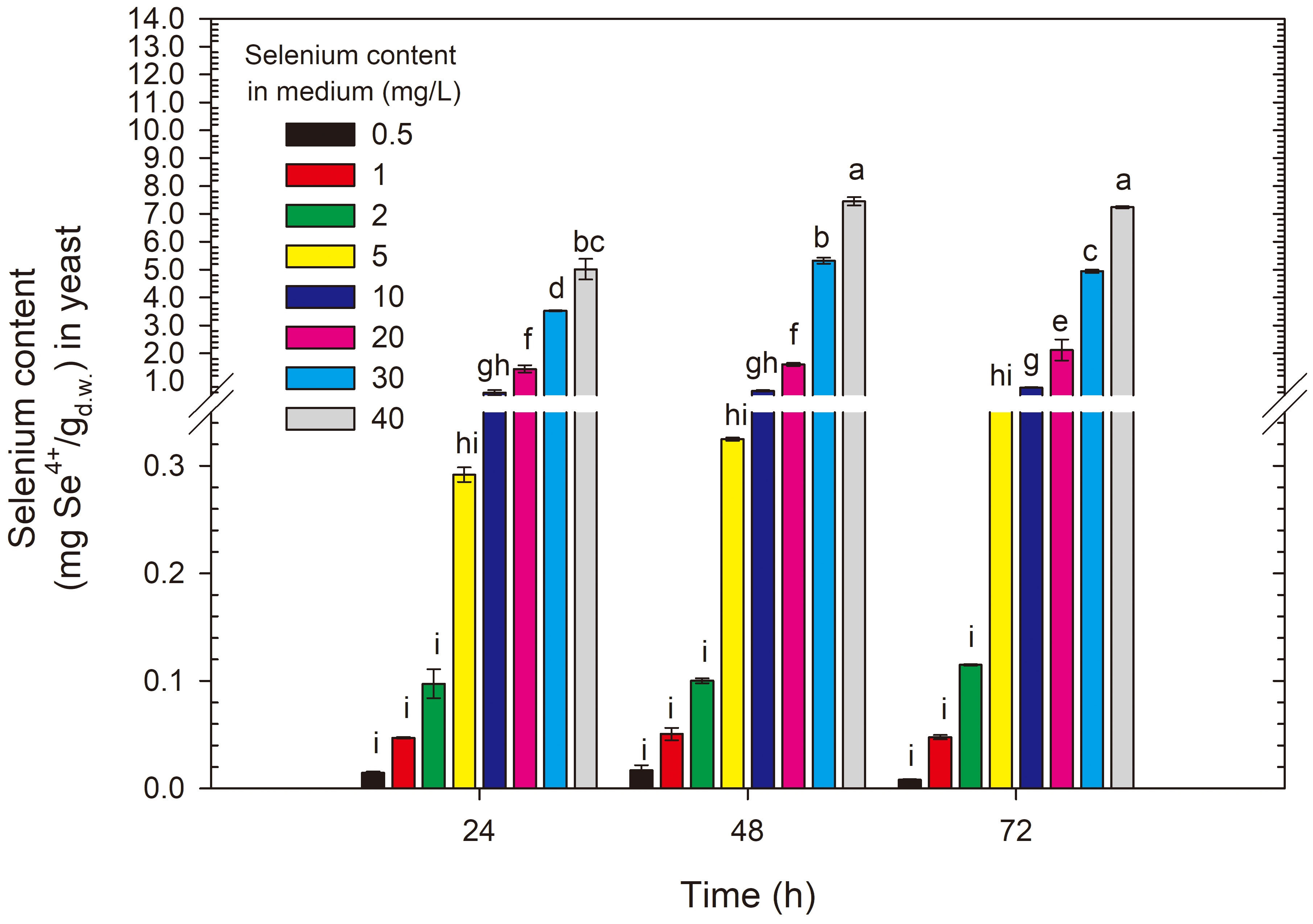 Fig. 6.
Fig. 6.
Selenium content in R. glutinis yeast cells grown in medium with different selenium additions in three time units. a-iMeans with the same letter did not differ significantly.
Comparing the results obtained for the two tested strains, it can be observed that the Rhodotorula yeast showed just over half selenium accumulation but exhibited better tolerance to higher selenium doses owing to twice the biomass yield compared with the results for S. cerevisiae ATCC 7090.
Several factors influence the ability of yeast cells to accumulate selenium – cultivation conditions, initial selenium concentration in the experimental medium and the type of microorganism studied [10]. In the study by Martiniano et al. [36], analyses was conducted using two different strains of S. cerevisiae (Lalvin ICV D47, 174, 193 and 405). A selenium dose of 15 mg Se4+/L resulted in the accumulation of this element at levels of 0.66 and 1.1 mg/g, respectively. Other two strains of S. cerevisiae (EC1118 and AW106) in the study by Yoshinaga et al. [37] showed binding of 3.1 and 3.5 mg Se/g, respectively, in a medium with an initial selenium dose of 34.58 mg/L. The study conducted by González-Salitre et al. [10] on the S. boulardii strain demonstrated selenium accumulation of 0.5, 0.79 and 3.4 mg Se/g after 3, 6 and 9 h of cultivation, respectively, with an initial selenium concentration in the medium of 74 mg/L. In the study conducted by Kieliszek et al. [23], the best selenium accumulation results were achieved in the cultivation of the S. cerevisiae ATCC MYA-2200 strain, with 1.61 mg Se4+/g obtained after adding 10 mg Se4+/L over a 72-h period. In the case of the C. utilis ATCC 9950 strain, 1.84 mg Se4+/g was obtained after adding 20 mg Se4+/L over 72 h. Analysing the efficiency of selenium utilization, yield and biomass content, the optimal selenium amounts were evaluated at approximately 15 mg Se4+/L for S. cerevisiae and 25 mg Se4+/L for C. utilis. The optimal cultivation time in both cases was 48 h. The study by Nam et al. [38] showed that the addition of selenium to the YM culture medium at concentrations ranging from 30 to 125 mg Se4+/L resulted in an increase in selenium content in the cell biomass of S. cerevisiae 6M from 1.04 to 5 mg/g. The study on R. glutinis yeast provided results confirming selenium accumulation at a level of 5.3 mg/g [13]. The study by Kieliszek et al. [21] reported new information on selenium accumulation by red yeast. The highest efficiency in selenium absorption from the environment was observed on the fourth day of cultivation, where statistically significant differences between doses influenced the amount of this element in the biomass of the tested yeast strain, especially in the case of the maximum selenium dose (20 mg Se4+/L). As a result, after four days of cultivation, the yeast showed gradual selenium utilization efficiency from the experimental environment in the following proportions: 78.04%, 53.48% and 57.50% at selenium concentrations of 10, 15 and 20 mg Se4+/L in the medium, respectively [21].
Krausova et al. [39] investigated the selenium bioaccumulation capacity of commercial lactic acid bacteria strains Streptococcus thermophilus CCDM 144 and Enterococcus faecium CCDM 922A in the presence of various sodium selenite concentrations (0–50 mg/L). The researchers noted that the ability of lactic acid bacteria to accumulate selenium increased with increasing selenium concentration in the culture medium. Both strains showed the highest selenium accumulation using an initial dose of this element at 50 mg/L–7.3 and 6.5 mg/g for CCDM 144 and CCDM 922A strains, respectively. Pieniz et al. [40] also reported high selenium accumulation by the Enterococcus durans LAB18s strain. Enterococcus durans was cultivated in BHI medium (35 °C, 24 h, anaerobic conditions) containing selenium in the form of sodium selenite at concentrations of 15, 30, 60 and 120 mg/L. The studies showed a significant correlation between selenium bioaccumulation in the biomass and the concentration of this element in the culture medium. Selenium concentration in bacterial biomass increased from 8.4 to 61.5 mg/g with increasing sodium selenite concentration added to the medium, ranging from 15 to 120 mg/L. The rate of selenium bioaccumulation reached 5.68% at a sodium selenite concentration of 60 mg/L. The analysis of the percentage of selenium binding suggests that the degree of bioaccumulation of this element increases until the sodium selenite concentration reaches 30 mg/L.
Hyrslova et al. [27] evaluated the ability of various bacterial strains from the genera Streptococcus, Lactococcus and Enterococcus to accumulate selenium in a medium containing 10 mg/L and 50 mg/L Na2SeO3. At a concentration of 10 mg Se4+/L in the culture medium, selenium content ranging from 2.5 to 7.4 mg/g was observed in bacterial cells. Only the Streptococcus thermophilus CCDM 561 strain had significantly higher selenium content, reaching approximately 45 mg/g. As in the above example, higher selenium concentrations (50 mg/L) in the culture medium resulted in a 4.5 to 7.8-fold increase in total selenium content in bacterial cells (17–201 mg Se4+/g).
Ponce et al. [41] conducted the study on various methods of selenium enrichment in S. cerevisiae yeast. They found that the best approach is to use lower concentrations of Na2SeO3, ranging from 10 to 50 mg/L, in the early logarithmic growth phase of the yeast. Additionally, temperature and pH are important factors influencing the selenium enrichment process. Yin et al. [42] demonstrated that the optimal conditions for selenium enrichment of S. cerevisiae are a temperature of 27 °C and a pH environment of 5.8. The culture was conducted in PDB (Potato, dextrose, broth) medium at 160 rpm. The study by Khakpour et al. [43] found that for the biomass of baker’s yeast S. cerevisiae PTCC 5010, the highest selenium binding level was 12.5 mg Se4+/g at a pH of 5.0. Kieliszek et al. [33] reported that pH in the range of 4.0–6.0 does not significantly affect selenium binding by yeast cells, hence the diversity of starting pH in different researchers’ studies, and in this study, the pH of the experimental medium was set at 4.8, which falls within the above range.
Differences in selenium accumulation by the same species of yeast may be due to metabolic variability [10]. Phosphate transporters and the regulation of their activity through phosphate concentrations are another factor influencing the interaction between selenium ion resistance and its accumulation. It has been shown that conditions characterized by low phosphate levels in the culture medium increase the activity of low-affinity phosphate transporters, leading to increased selenium uptake by yeast cells [14, 37].
One of the yet uncharacterized compounds is elemental selenium (Se0). There are reports in the available literature about the synthesis of selenium nanoparticles (SeNP) by microorganisms through enzymatic reduction of Se2- to Se0 [2, 10]. Yeasts absorb selenium from the medium and transport it through the membrane, where it reacts with glutathione (GSH) to form selenodiglutathione (GSSeSG). GSH continues the reduction process, converting it into selenenyl sulphide glutathione (GSSeH). In the next step, this selenenyl sulphide glutathione (GSSeH) undergoes spontaneous dismutation, leading to the formation of elemental selenium (Se0) and the regeneration of GSH [44].
Elemental selenium production likely occurs when yeasts are unable to biotransform selenium into organic compounds (selenoamino acids) owing to excessively high concentrations of this element in the culture medium. It has been observed that selenium concentration in the medium is directly proportional to the aggregation of elemental selenium, although this mechanism has not been thoroughly studied and remains partially unclear, associated with the Ostwald mechanism where smaller particles coalesce into larger aggregates without control over their size due to significant surface energy [45]. In the study by González-Salitre et al. [10] on S. boulardii, an initial selenium concentration of 74 mg/L resulted in the production of elemental selenium. For the S. cerevisiae ATCC 7090 yeast examined in this study, the threshold dose causing Se0 production is 30 mg/L. According to the study by Kieliszek et al. [23], using selenium concentrations exceeding 40 mg Se4+/L in experiments is uneconomical because a significant amount of selenium remains unused in the culture fluid. Under such conditions, a red precipitate can be observed, suggesting the presence of large amounts of reduced selenium. Biomass obtained under such conditions contains increased amounts of elemental selenium, which is characterized by low bioavailability [44].
This study concludes that the optimal conditions for producing selenium-enriched yeast from the S. cerevisiae ATCC 7090 strain involve culturing for 24 h in a medium with an initial selenium dose of 20 mg/L. Under such conditions, the yeast can bind 4.27 mg/g, and this amount of accumulated element, considering the literature, falls within the upper range of results. The upper intake limits for selenium in humans are not directly regulated at the European level. Relevant organizations, such as the WHO (World Health Organization) and EFSA (European Food Safety Authority), recommend a daily selenium intake for adults of 30–40 µg. The Food and Nutrition Board of the National Academy of Sciences notes that daily selenium requirements vary by age, being 40–70 µg for men and 45–55 µg for women [8, 46]. Proper preparation of selenium-enriched yeast biomass could easily help mitigate selenium deficiencies worldwide. Culturing for 24 h allows for additional economic benefits compared with cultures conducted for 48 h or more. Higher selenium doses lead to the precipitation of elemental selenium, and a longer culturing time does not significantly impact the selenium accumulation results. According to Kieliszek et al. [23], using selenium at concentrations above 40 mg Se4+/L in experimental conditions is uneconomical because a high level of selenium remains unused in the culture medium. Under these conditions, a red precipitate can be observed, suggesting the presence of large amounts of reduced selenium. Selenium in this form is biologically inactive due to its low bioavailability.
For the red yeast R. glutinis CCY 20-2-26, optimal selenium binding conditions were obtained by culturing for 48 h in a medium with an initial selenium ion content of 40 mg/L. This yeast strain proved to be more resistant to high selenium doses, and at the highest studied selenium concentration, it accumulated 7.53 mg Se4+/g. Considering the literature used to write the discussion of this part of the study, this result is record-breaking and is 42% higher than the result (5.3 mg/g) obtained by Wang et al. [13]. Such a high selenium binding result by the biomass and greater tolerance to high selenium doses compared with the S. cerevisiae strain may be related to the natural ability to produce carotenoid pigments.
Owing to the documented health benefits of carotenoids (combating neurological
diseases, peptic ulcers, cancer), scientists are continually seeking new ways to
obtain them [47]. Many carotenoids are currently produced through chemical
synthesis, but as demand for natural colourants grows, production via
microorganisms is gaining significance. Among microorganisms capable of producing
carotenoids, yeasts of the genus Rhodotorula are particularly
interesting. These organisms primarily synthesize
The carotenoid content in R. glutinis yeast biomass during cultivation in a control medium without selenium addition was the highest (Fig. 7). After 24 h of cultivation, the carotenoid content in the control sample was 163.92 µg/g, and after 48 and 72 h, it significantly increased to 205.07 and 200.14 µg/g, respectively. The addition of the lowest selenium dose (0.5 mg Se4+/L) after 24 h of cultivation caused a significant decrease in carotenoid content in the biomass (130.54 µg/g), and subsequent days of cultivation did not significantly affect their content in the biomass. The addition of selenium to the medium in doses of 1 and 2 mg Se4+/L significantly reduced carotenoid content in the biomass to 75.32–85.24 µg/g compared with the control sample. Selenium in amounts above 5 mg Se4+/L also negatively impacted the biosynthesis of carotenoids, limiting their amount in the biomass to only 16.14–40.05 µg/g.
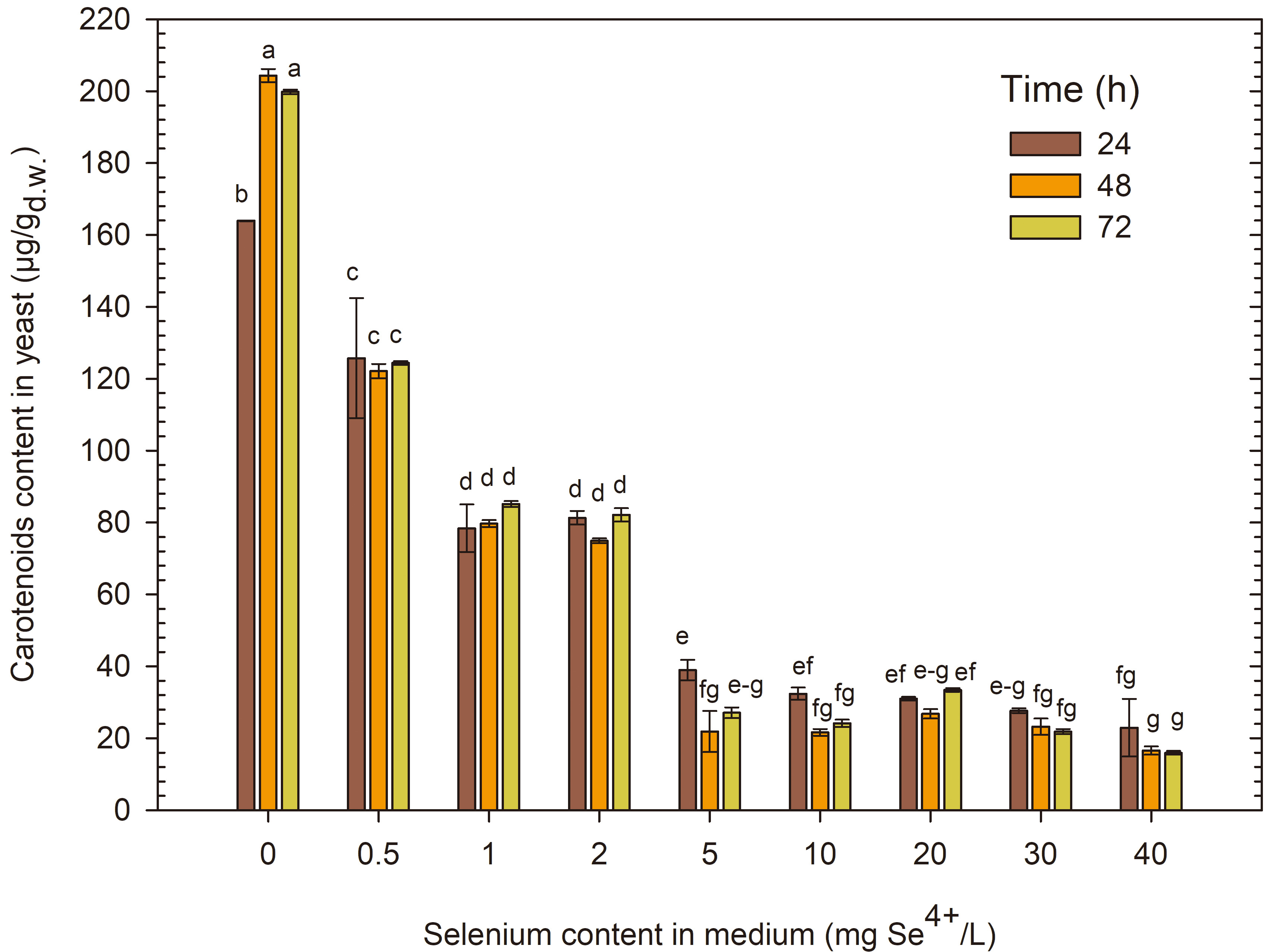 Fig. 7.
Fig. 7.
Content of carotenoids extracted from R. glutinis yeast cells grown in medium with various amounts of selenium in three time units. a-gMeans with the same letter did not differ significantly.
Carotenoids are a key group of low-molecular-weight antioxidants in the non-enzymatic process. Many microorganisms, such as bacteria, molds and yeasts, can produce various carotenoids, including gamma and beta-carotene, lycopene, torulene, torularhodin and astaxanthin [47]. It is particularly noteworthy that yeasts belonging to the genera Rhodotorula, Rhodosporidium, Sporobolomyces and Phaffia demonstrate an exceptional ability to produce carotenoids even under harsh conditions such as UV radiation, high temperature, or the presence of solvents or heavy metals. The study by Bogacz-Radomska and Harasym [48] showed that the presence of trace elements in the medium has a varied impact on the profile and the quantity of carotenoids synthesized by Rhodotorula graminis DBVPG 7021. For example, the presence of aluminium (III) and zinc (II) stimulated the synthesis of beta-carotene and gamma-carotene, while zinc (II) and manganese (II) inhibited the production of torulene and torularhodin. Another study by Bhosale and Gadre [49] demonstrated that divalent cations such as zinc (II), cobalt (II), iron (II) and copper (II) had a stimulating effect on the production and accumulation of carotenoids in R. glutinis CCY 20-2-26 yeast cells.
Research conducted by Nechay et al. [50] on Phaffia rhodozyma
yeast showed that as selenium concentration in the medium increased, the quantity
of carotenoids in the yeast biomass proportionally decreased. During these
studies, various doses of selenium in the form of sodium selenite were used,
including
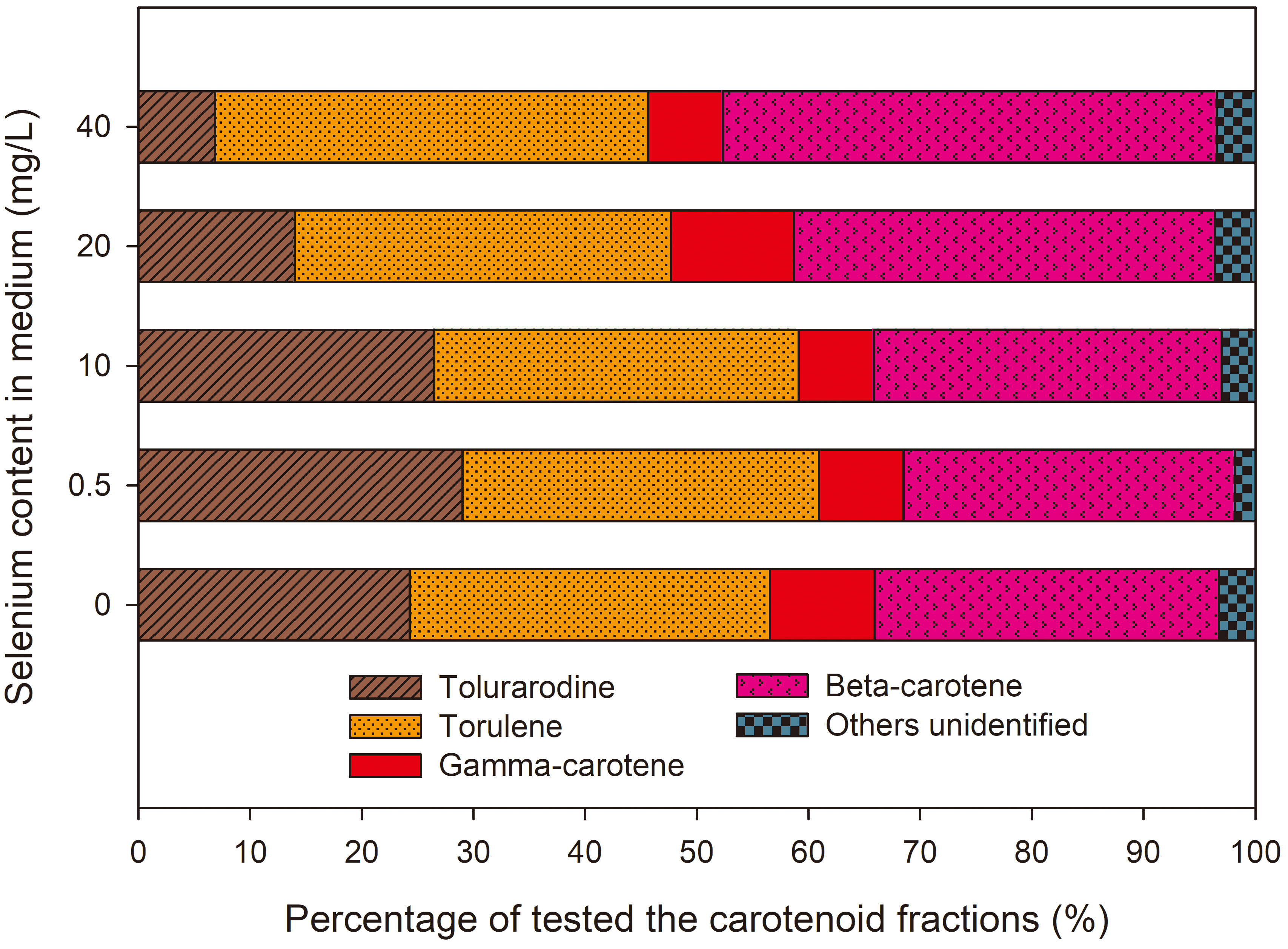 Fig. 8.
Fig. 8.
Percentage of carotenide fractions from R. glutinis yeast cells grown in medium with various amounts of selenium in three time units.
It was found that the presence of selenium in the medium in amounts
The study by Díaz-Navarrete et al. [51] have shown that R. mucilaginosa 6S responds to the presence of selenium with a varied production of biomass, carotenoids, and other biochemical components. Selenium supplementation in the form of sodium selenite (Na₂SeO₃) showed that concentrations above 10 mg/L significantly limited biomass growth, which may be the result of selenium-induced oxidative stress. An increase in selenium concentration disrupted the oxidation-reduction balance of cells and led to lower carotenoid production. In the control sample, where yeast was cultivated without selenium supplementation, the carotenoid content was 221.3 µg/g of biomass. After introducing selenium at a dose of 30 mg/L at the beginning of the culture, the carotenoid content decreased to 63.3 µg/g of biomass, a decrease of about 71% compared to the control sample. When selenium was added after 48 hours, when the cells were already in the logarithmic phase, the carotenoid content was 163.3 µg/g biomass, which means a decrease of about 26% compared to the control. The obtained results indicate that adding selenium at the late stage of the logarithmic growth phase can promote the bioaccumulation of selenium while minimizing its negative impact on carotenoid biosynthesis.
Moreover, studies on the effect of selenium on carotenoid biosynthesis are scarce, so to complement the discussion, we cite the results of studies on other metal salts that may have a similar effect on producing these compounds in yeast. The addition of zinc ions to the medium significantly affects the biosynthesis of carotenoids by the yeast R. toruloides MH023518, especially at appropriate concentrations [52]. The study showed that using zinc oxide nanoparticles at a dose of 50 mg/L increased the total carotenoid production to 264 mg/L, while the control without the addition of ZnO reached only 124 mg/L. However, with a further increase in the ZnO concentration to 200 mg/L, the carotenoid content decreased to 152 mg/L. These results suggest that zinc supports carotenoid production, but only up to a certain level, above which the production of pigments is inhibited. Another study by Elfeky et al. [53] presents the results on the effect of aluminum ions on carotenoid production in R. glutinis (AS 2.703). The culture medium was supplemented with aluminum sulfate Al₂(SO₄)₃ at a concentration of 0.7 mM, which resulted in an increased total carotenoid content of 2.21 mg/L. Increasing the dose of aluminum sulfate resulted in a decrease in carotenoid production. The study’s results indicate that selenium may be an essential factor modulating carotenoid production, but its presence at too high concentrations inhibits this production. This finding is crucial for further studies on optimizing the cultivation conditions of yeasts of the genus Rhodotorula to obtain biomass rich in carotenoids. The results suggest that to increase the efficiency of carotenoid biosynthesis by yeast, it is essential to precisely adjust the selenium concentration in the cultivation environment, which may be the basis for new bioproduction strategies [51].
The study provided evidence that selenium negatively affects the concentration of carotenoid pigments in the analysed Rhodotorula yeast. The type, oxidation state and amount of this element present in the culture medium can influence the regulation of carotenoid enzyme activities, especially specific desaturases responsible for carotenoid biosynthesis. This enzymatic regulation occurs in red yeast cells, according to the works of Kot et al. [26].
Combining studies on selenium accumulation assessment and changes in carotenoid content in yeast cell biomass is a significant gap in the available literature. The selection of appropriate cultivation parameters and selenium concentrations in the starting medium can contribute to optimal selenium binding in yeast biomass while increasing desired carotenoid fractions (e.g., astaxanthin). Such advancement would undoubtedly represent a significant step in the discussed research field.
Microorganisms can synthesize various valuable compounds, such as lipids (especially single-cell oil, SCO). Oleaginous microorganisms are characterized by their ability to produce and accumulate over 20% lipids in dry cell mass. Oleaginous yeasts are rare among all yeast species, constituting only about 5% of the total population. The genera of oleaginous yeasts that have attracted the most attention from researchers are Trichosporon, Cryptococcus, Rhodotorula, Rhodosporidium, Candida, Yarrowia and Lipomyces [54]. The fat content in their biomass can reach up to 60% (Yarrowia lipolytica) [55]. Yeasts mainly produce palmitic acid (16:0), oleic acid (18:1), linoleic acid (18:2) and linolenic acid (C18:3) [26].
After 24 h of cultivation, it was observed that the studied yeasts, S. cerevisiae and R. glutinis, showed no significant signs of fat production. The results of the quantitative analysis of extracted lipids (2.5–4.9% for S. cerevisiae (Fig. 9) and 2–3% for R. glutinis (Fig. 10)) suggest that these amounts are so small that they are not likely to be accumulated as storage substances. Rather, these lipids are present in cell structures, such as the cell wall-membrane complex. To better understand this issue, it is worth mentioning that cellular fats can be stored in various organelles within the cell. The main locations where intracellular lipids can be present are primarily cell membranes, where phospholipids form an essential part of the structure. The fats in yeast cells can also include lipids of the internal membranes of organelles, such as mitochondria, where they play important metabolic functions (beta-oxidation, lipid synthesis, apoptosis regulation) [56].
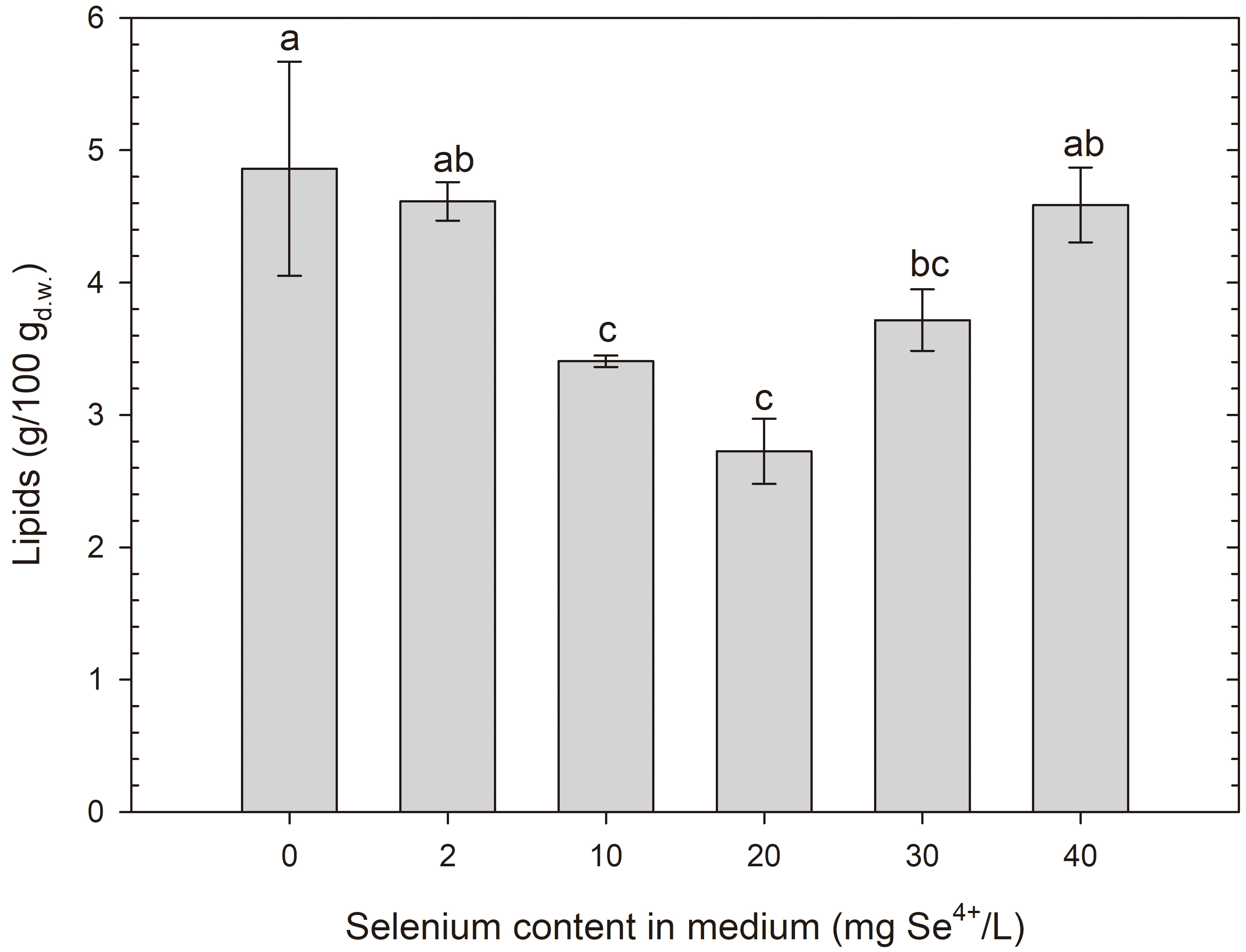 Fig. 9.
Fig. 9.
Lipid content in S. cerevisiae yeast cells grown in medium with different selenium additions in three time units. a-cMeans with the same letter did not differ significantly.
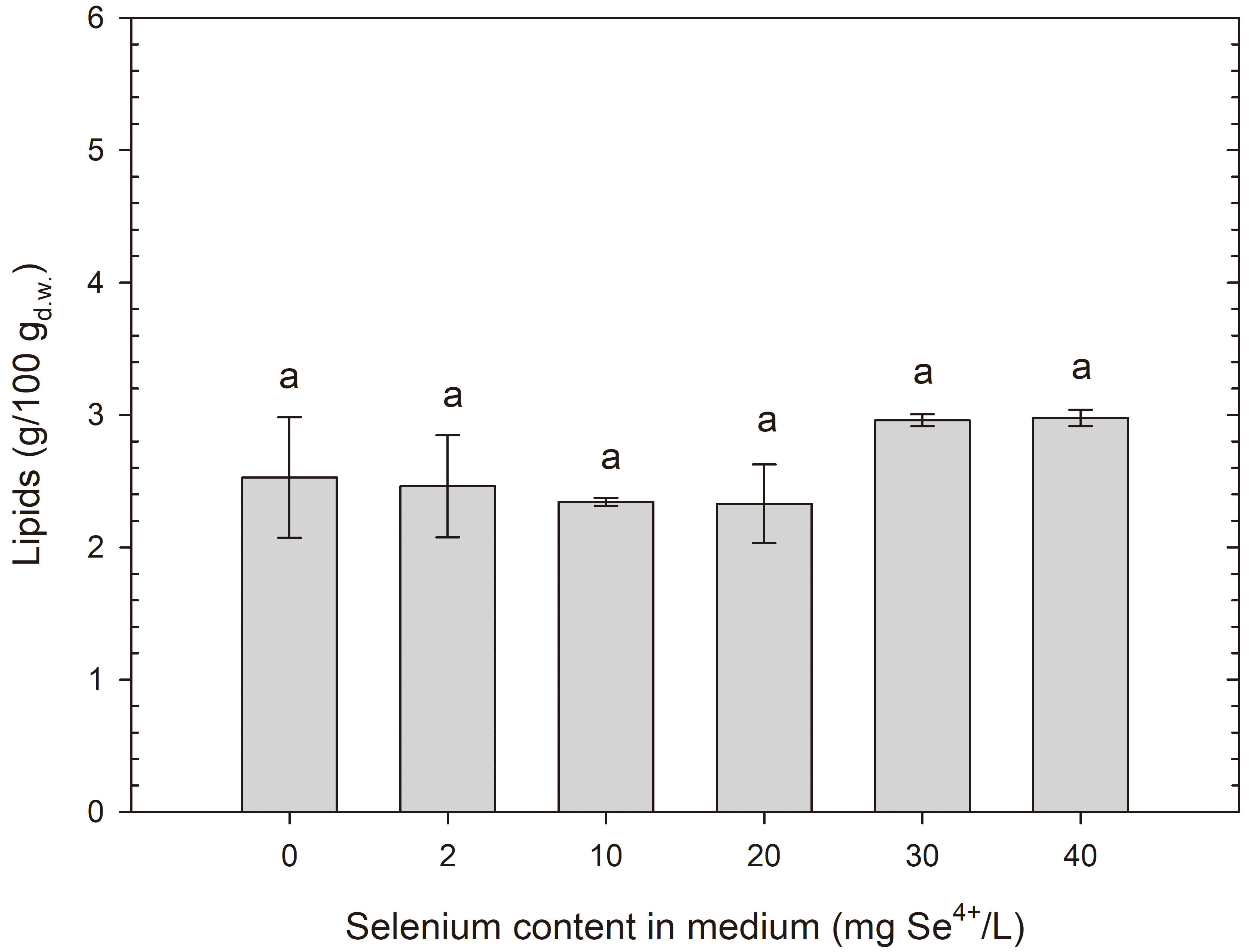 Fig. 10.
Fig. 10.
Lipid content in R. glutinis yeast cells grown in medium with different selenium additions in three time units. aMeans with the same letter did not differ significantly.
The study by Tahmasebi et al. [54] on the simultaneous production of carotenoids and lipids by Rhodosporidium babjevae yeasts showed that with a glucose content of 60 g/L in the medium, 10.5% lipids were detected, while the highest carotenoid concentration (312 µg/g) was observed at a glucose content of 10 g/L. A summary of lipid production by other yeast strains in the available literature is as follows: Rhodotorula mucilaginosa CCT 7688 (no lipid production capability) [57], Yarrowia lipolytica Po1g (up to 58.5%) [58], Rhodosporidium toruloides 5149 (up to 30.3%) [59], Yarrowia lipolytica 5054 (up to 29.3%) [59], Rhodosporidium toruloides RP (up to 59.8%) [60].
In conclusion, selenium does not affect the fat accumulation process in the microorganisms, indicating that this element has no significant impact on lipid biosynthesis in yeast cells.
Environmental condition changes during cultivation can affect gene expression related to the production of new proteins in yeast cells. An example of such changes is the addition of selenium to the cultivation medium. The study indicates [44] that the expression of selenoproteins in organisms reaches its maximum at moderate selenium concentrations. However, higher doses of selenium can lead to lipid peroxidation in cell membranes. This underscores the importance of precise control of microbial cultivation conditions to optimize the production of desired proteins and minimize potential negative effects of this biotechnological process.
In this study, the impact of increasing selenium doses on changes in protein content in the cellular biomass of tested yeast strains after 24 and 48 h of cultivation was examined (Figs. 11,12). The biomass of S. cerevisiae ATCC 7090 in the control sample contained 59.65% total proteins after 24 h of cultivation, and 52.23% after 48 h. A selenium dose of 10 mg/L caused a significant increase in proteins after 24 h compared with the control sample (69%); however, after another day of cultivation, this value returned to the level observed in the control sample after 24 h of cultivation. Selenium doses of 20, 30 and 40 mg/L did not cause significant changes in total protein content compared with the medium with an initial selenium concentration of 10 mg/L.
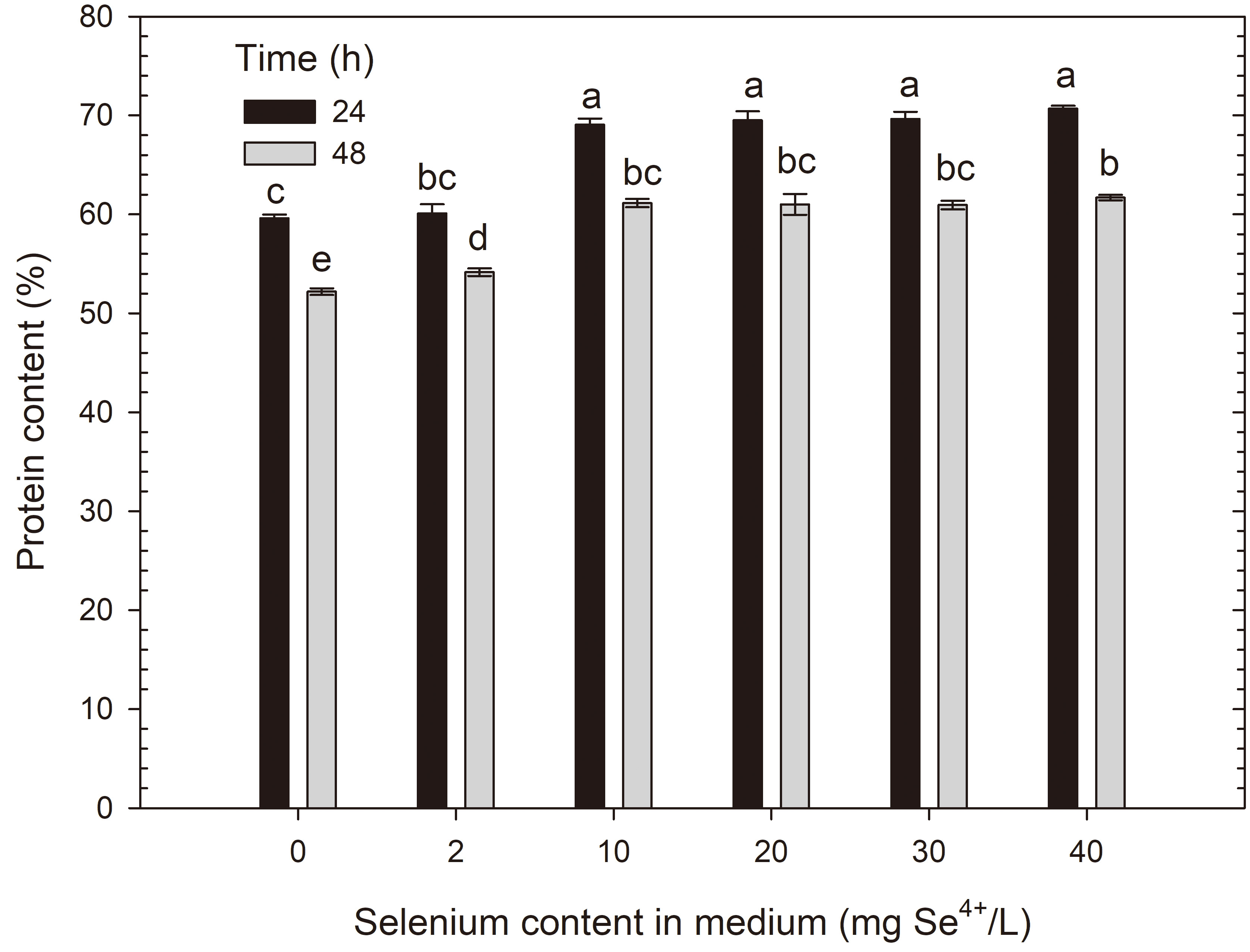 Fig. 11.
Fig. 11.
Protein content in S. cerevisiae yeast cells grown in medium with different selenium additions in three time units. a-eMeans with the same letter did not differ significantly.
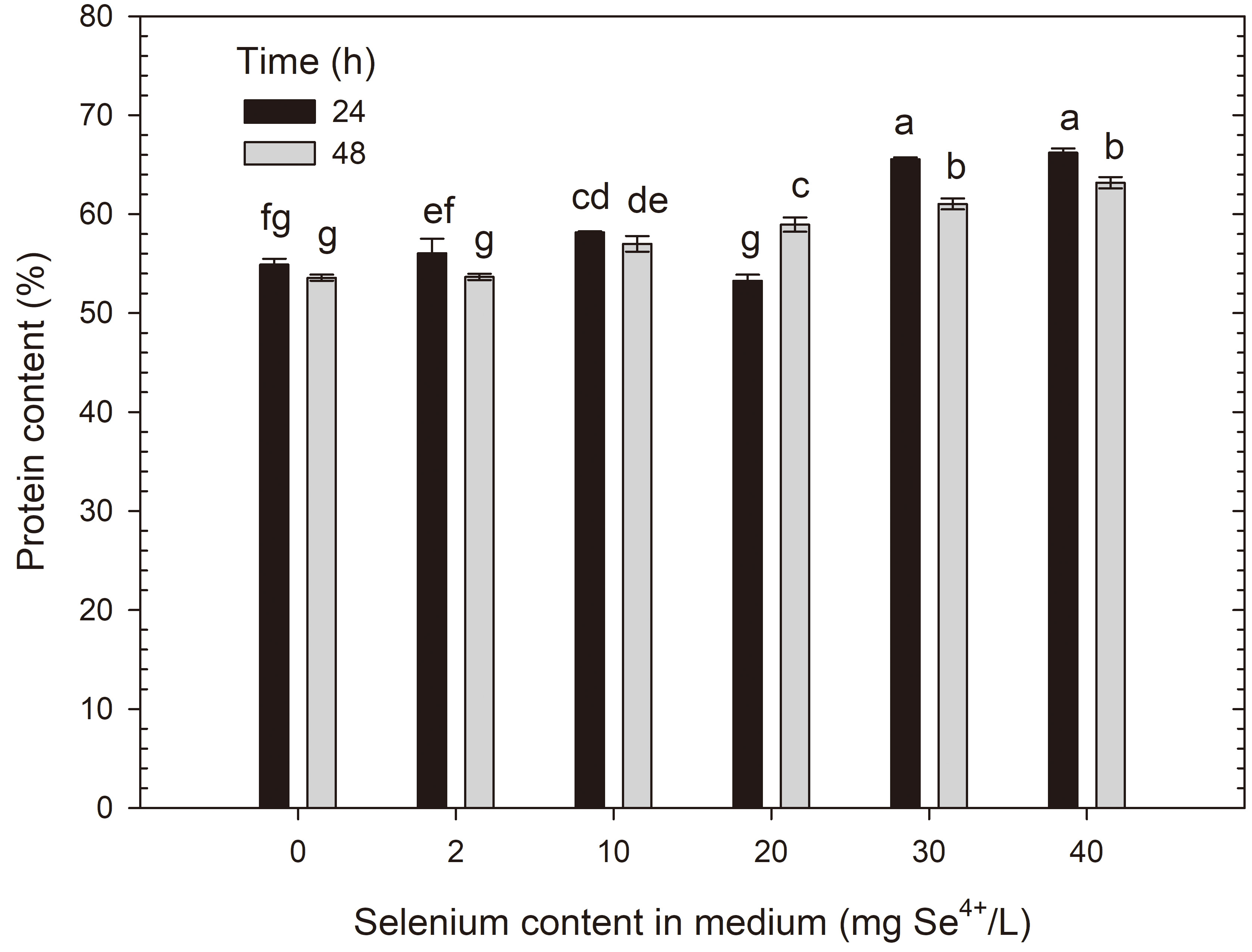 Fig. 12.
Fig. 12.
Protein content in R. glutinis yeast cells grown in medium with different selenium additions in three time units. a-gMeans with the same letter did not differ significantly.
The biomass of R. glutinis CCY 20-2-26 yeast cells in the control
medium had a total protein content of 55.02% after 24 h of cultivation, and
53.58% after 48 h. Selenium doses of
The results showed an increase in protein content during the cultivation of Saccharomyces and Rhodotorula yeasts in selenium-containing media. The presence of selenium in the medium seems to stimulate the biotransformation of this element into its organic forms, such as selenomethionine, selenocysteine or selenomethylselenocysteine. This, in turn, can result in an increase in the number of proteins involved in these processes, including homocysteine synthase, methionine synthase and selenomethyltransferase [44].
In another study conducted by Kieliszek et al. [33], selenium supplementation of C. utilis ATCC 9500 and S. cerevisiae MYA-2200 cultures at a concentration of 20 mg Se4+/L contributed to increased levels of essential amino acids, including lysine, leucine and valine, compared with the those in the control culture. Parrondo et al. [61] in their studies showed that longer cultivation times of Kluyveromyces marxianus CECT 1123 yeast are associated with autolysis in the stationary growth phase, leading to a significant decrease in soluble protein content in cellular structures. For K. marxianus yeast, Aggelopoulos et al. [62] and Yadav et al. [63] report protein content ranges from 43 to 59%. In contrast, Agboola et al. [64] report that the protein content in Saccharomyces cerevisiae and Cyberlindnera jadinii yeast is 50.1% and 46.3%, respectively.
Guaragnella and Bettiga [65] described the effect of acetic acid as a stress factor suggesting an increase in the expression of genes involved in transcription and protein synthesis in the initial growth phase (first 24 h) and stimulation of protein metabolism in the later growth phase (48 h) because of S. cerevisiae yeast cells responding to stress caused by the addition of acetic acid (1.2–12 g/L). According to Yuan et al. [29], in response to various stress conditions (thermal, osmotic, metals, acids, salts, H2O2), S. cerevisiae yeast can adapt to unfavourable conditions by remodelling the cell wall and membrane, reprogramming gene expression and making metabolic adjustments (e.g., increasing antioxidant production, changing lipid profiles, producing protective substances). It is worth noting that according to the literature, the high protein content in dried yeast biomass typically ranges from 35% to 53% [33]. However, the studied yeast strains showed even higher protein content, up to 70% (S. cerevisiae strain after 24 h in a medium with an initial selenium content of 40 mg/L) after selenium supplementation. These results suggest that selenium may affect yeast metabolic processes and can serve as a starting point for further research on the biochemical mechanisms of selenium’s impact on these organisms. Further research will allow for a better understanding of new metabolic pathways involving selenium or those it may intensify.
This study provides new insights into selenium accumulation by model yeast S. cerevisiae and red yeast R. glutinis, as well as the impact of this element on the synthesis of carotenoid pigments and proteins. The use of a natural method to enrich yeast biomass with selenium leverages the biological potential of microorganisms.
Optimal conditions for obtaining selenium-enriched yeast from the tested S. cerevisiae strain involve cultivation for 24 h in a medium with an initial selenium dose of 20 mg Se4+/L. For red yeast R. glutinis, optimal selenium binding conditions were achieved by cultivating for 48 h in a medium with an initial selenium ion content of 40 mg Se4+/L. This increased resistance to the toxic effects of selenium in red yeast may be due to their natural ability to produce carotenoids. Carotenoids have the ability to neutralize free radicals in organisms, which is a key defensive mechanism against oxidative damage.
Additionally, the study results suggest that selenium increases the protein content in cellular biomass without affecting the production of intracellular fat. The increased protein content during the cultivation of S. cerevisiae and R. glutinis in media supplemented with selenium may be due to the enhanced activity of enzymes involved in intracellular transport and the conversion of selenium into various chemical compounds with antioxidant properties. The presence of selenium in the medium may stimulate the growth of protein structures involved in the biotransformation of this element into organic forms.
The detoxification mechanism in response to increased selenium content appears to differ between the two yeast species. In S. cerevisiae, this leads to the reduction of selenium to its elemental form and an increase in cell surface area. Conversely, R. glutinis copes better with oxidative stress caused by the presence of selenium, likely due to more efficient detoxification systems and a decrease in cell surface area with increasing selenium concentration.
Nevertheless, to fully understand the selenium metabolism process in S. cerevisiae ATCC 7090 and R. glutinis CCY 20-2-26, further research is necessary. Despite this, the results open new perspectives in the fields of biotechnology and food technology (technological processes related to the production of dietary supplements, food enrichment with nutrients, or technologies for preparing special food products for individuals with specific nutritional needs) and indicate further potential challenges to be addressed.
The datasets used and analyzed during the current study are available from the first author upon reasonable request.
WS: resources, investigation, formal analysis, writing — original draft; AMK: investigation; MK: conceptualization, formal analysis, methodology, investigation, resources, writing-review and editing, supervision. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Our research did not include any human subjects or animal experiments. Neither ethics approval nor consent is needed. According to local rules, it was not necessary to obtain ethical approval.
The research was carried out using the equipment purchased as part of the “Food and Nutrition Centre—modernization of the WULS campus to create a Food and Nutrition Research and Development Centre (CŻiŻ)” co-financed by the European Union from the European Regional Development Fund under the Regional Operational Programme of the Mazowieckie Voivodeship for 2014–2020 (Project No. RPMA.01.01.00-14-8276/17).
This work was funding by the Preludium Bis 2 (2020/39/O/NZ9/00639). Marek Kieliszek declared that the funds were supported by the National Science Centre (NCN), Poland.
Given his role as the Guest Editor and Editorial Board member, Marek Kieliszek had no involvement in the peer-review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process for this article was delegated to Rosa Alduina. The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.