


 , Maria Letizia Salvetat 3,†, Mutali Musa 4, Fabiana D'Esposito 5,6, Dario Rusciano 7,§, Antonino Maniaci 1, Francesco Pellegrini 3, Giuseppe Scibilia 8, Marco Zeppieri 9,*
, Maria Letizia Salvetat 3,†, Mutali Musa 4, Fabiana D'Esposito 5,6, Dario Rusciano 7,§, Antonino Maniaci 1, Francesco Pellegrini 3, Giuseppe Scibilia 8, Marco Zeppieri 9,*
1 Department of Medicine and Surgery, University of Enna “Kore”, 94100 Enna, Italy
2 Eye Clinic, Catania University San Marco Hospital, 95121 Catania, Italy
3 Department of Ophthalmology, Azienda Sanitaria Friuli Occidentale, 33170 Pordenone, Italy
4 Department of Optometry, University of Benin, 300238 Benin, Edo, Nigeria
5 Imperial College Ophthalmic Research Group (ICORG) Unit, Imperial College, NW15QH London, UK
6 Department of Neurosciences, Reproductive Sciences and Dentistry, University of Naples Federico II, 80131 Napoli, Italy
7 Fidia Ophthalmics, 95125 Catania, Italy
8 Department of Obstetrics and Gynecology, “Giovanni Paolo II” Hospital, 97100 Ragusa, Italy
9 Department of Ophthalmology, University Hospital of Udine, 33100 Udine, Italy
†These authors contributed equally.
§Retired.
Abstract
This review explores the connection between the ocular surface microbiome and glaucoma, highlighting its impact on disease progression. Beginning with an overview of global glaucoma significance, it emphasizes the importance of understanding the cellular characteristics and microbiology of the ocular microbiome. A search was conducted on the PubMed and Cochrane Library databases using the phrase “ocular microbiome glaucoma”. 0 records were returned from the Cochrane Library while 21 were returned from PubMed. A total of 21 results were retrieved from 2017 to 2024. This comprised one opinion paper, four original research articles, and 16 reviews. This review covered the anatomy of the ocular surface, advanced analysis methods, and the ocular microbiome. It also delved into dysbiosis in glaucoma, addressing altered microbial communities and their potential role in disease progression. The intricate interplay between the ocular microbiome and the host’s immune system is explored, emphasizing crosstalk and inflammatory responses. The review concludes by discussing therapeutic implications, including modulating ocular microbiota and potential future treatment strategies. Understanding the microbiome in healthy and glaucomatous eyes can help researchers and clinicians in innovative approaches to ocular health.
Keywords
- microbiome
- ocular surface
- glaucoma
- tear film
- cornea
- microbiome analysis
- treatment strategies
Glaucoma, a disease that leads to irreversible blindness, is a significant global health issue [1, 2]. It is estimated that by 2040, over 111.8 million individuals will be affected by this condition. The disease is characterized by the progressive and irreversible loss of retinal ganglion cells (RGCs), primarily due to increased intraocular pressure (IOP). Glaucoma encompasses open-angle glaucoma (OAG) and angle-closure glaucoma (ACG) and can be further categorized as primary or secondary [1, 2]. The total prevalence of primary open-angle glaucoma (POAG) was about 3.1%, while 0.5% of people had primary angle-closure glaucoma (PACG) [3, 4].
Based on these results and observations of normal tension glaucoma (NTG) and ocular hypertension (OHT), high IOP is not the sole risk factor or the mechanism of RGC death and glaucomatous optic neuropathy. It is neither necessary nor sufficient to cause glaucoma [5]. Indeed, RGC degeneration can be mediated by other non-IOP signaling cascades, including vascular, metabolic, oxidative, neurotrophic factor deprivation, and immune and inflammatory components. In addition, aging, ethnic background, sex, lifestyle, diet, body mass index (BMI), obesity, and depression also contribute to the development of glaucoma [5]. In this regard, glaucoma is a complex, multifactorial neurodegenerative disease involving many triggers, cell types, and signaling pathways [6]. This complexity underscores the need for comprehensive research and understanding.
It’s important to clarify the often-interchanged terms ‘microbiota’ and ‘microbiome’. The term ‘microbiota’ refers to the living microorganisms found within a specific environment, such as the human body or its organs and tissues [7]. On the other hand, ‘microbiome’ encompasses the genetic material of these microorganisms and their surrounding environmental conditions. In other words, the microbiome includes the microbiota and their genetic elements, metabolites, and structural components. Dysbiosis, a term used to describe an imbalance or disruption in the composition and function of the microbiota, can lead to adverse effects on health when harmful microorganisms proliferate or beneficial ones decrease in number [8].
Exploration into the diversity of the human microbiota traces back to Antonie van Leeuwenhoek, who, as early as the 1680s, observed and compared the microbial communities present in his oral and fecal samples [9, 10]. He noted significant disparities between these habitats and observed variations between individuals in states of health and illness in these regions. This highlights that investigations into microbial differences across body sites and health conditions are as ancient as the field of microbiology itself. What distinguishes modern research is not merely the ability to perceive these apparent distinctions but rather the utilization of advanced molecular techniques to delve into the reasons behind these variances and to comprehend how they can influence our health status and how we can influence transitions from one state to another.
Indeed, nowadays, we know that the human body is home to trillions of microorganisms, collectively known as microbiota, which play a crucial role in maintaining overall health and well-being [8]. Factors such as the external environment, dietary habits, and lifestyle choices significantly shape the composition and robustness of the microbiota. Recent research has highlighted the microbiota’s profound effects on health and disease. These microorganisms’ population, structure, variability, and viability are in constant flux, playing a pivotal role in the onset, progression, and management of numerous health situations. Notably, conditions ranging from cancer, metabolic disorders, and cardiovascular diseases to psychological disorders like schizophrenia can be influenced by the microbiota [10]. The microbiota can directly affect health by modulating physiological processes such as metabolism, immune function, and neurotransmitter production. Dysbiosis, or microbial imbalance, may lead to health problems like inflammation, metabolic disorders, and immune dysregulation. Microbiota can indirectly influence health by producing metabolites and signaling molecules interacting with host tissues and systems. These metabolites can affect cellular function, inflammation, and disease susceptibility. Additionally, dysbiosis-induced changes in the microbiota composition may alter susceptibility to infections, medication response, and risk of chronic diseases [10]. Understanding the significance and relevance of human microbiota is essential in comprehending their impact on human health and disease.
The gut is the best-known and the most populated site, harboring a diverse array of microorganisms, predominantly bacteria, that aid digestion, produce essential vitamins, and modulate the immune system. The gut microbiota, once overlooked, has now emerged as a central focus of medical and scientific research, revealing its crucial role beyond the gastrointestinal tract. Interdisciplinary studies have uncovered key pathways and molecular factors linking the gut microbiota to various bodily systems, including the central nervous system [11, 12, 13]. This complex dialogue profoundly influences host physiological functions, particularly central nervous system (CNS) development and health. In recent years, extensive research has unveiled a more comprehensive range of influences exerted by the gut microbiota beyond the digestive system, impacting various organ systems [14, 15, 16]. Imbalances in the gut microbiota have been associated with many brain disorders, encompassing neurodevelopmental, behavioral, and neurodegenerative diseases [17]. The skin is also inhabited by a complex ecosystem of bacteria, fungi, and viruses, contributing to skin health and immune responses. The skin microbiota is a barrier against pathogens, regulates inflammation, and influences skin conditions such as acne and eczema [18].
The oral cavity hosts a diverse microbial community pivotal in oral health and systemic well-being [19]. The oral microbiota is implicated in dental and periodontal diseases and is associated with systemic conditions such as cardiovascular diseases and diabetes [19]. The respiratory tract’s microbiota likely serves as a protective barrier, guarding against the colonization of harmful respiratory pathogens. Additionally, it may play a role in the development and upkeep of normal respiratory function and immune balance [20]. The nose also hosts a complex microbiota [21], which can affect neural signaling and brain function and behavior [22]. Most interestingly, a clinical study reported the efficacy and safety of a probiotic using Streptococcus salivarius 24SMBc and Streptococcus oralis 89a to prevent upper respiratory tract infections (URTIs) in children with recurrent respiratory infections (RRIs). The treatment (given as a nasal spray), administered over three months to 91 children, significantly reduced symptoms such as fever, cough, otalgia, and rhinorrhea compared to baseline. Younger children (1–3 years old) showed improvement after three months, while older children (3–6 and 6–12 years old) improved progressively from the first month. The treatment was generally well-tolerated, with only nine children experiencing temporary discomfort [23]. An association has also been described between the ear canal microbiome and generalized anxiety disorder (GAD), acting through the activation of inflammatory cytokine responses. The characterization of microbiota changes in the ear canal of GAD patients identified an increased microbial diversity and an altered abundance of specific bacteria in the ear canal, potentially involving upregulated inflammatory reactions. Additionally, the study explored the microbiota-ear-brain interaction and its potential role in the pathogenesis of GAD [24]. The human vaginal microbiota, closely associated with the vaginal epithelium, is a crucial determinant of vaginal health. The interaction between the human host and the vaginal microbiota is highly dynamic, shaped by transitional periods such as puberty, menopause, and pregnancy, as well as daily life activities and behaviors. Variability in microbial communities among women has been associated with an increased risk of adverse health outcomes, yet the mechanisms driving these associations remain fully elucidated [25, 26]. Finally, the eye, the subject of this review, also hosts its peculiar microbiota, and its physiological state is also influenced by both the local and the gut microbiota [27]. Understanding the literature on the ocular microbiota characteristics in glaucoma is vital to future interventions in early detection and prevention of glaucomatous damage.
The eyelids, cornea, sclera, and conjunctiva constitute the ocular surface. It is bathed by the tear film, which has an interdependent role and is an essential component for its integrity and function. The cornea is a transparent avascular tissue located in the anterior part of the eye. It represents a fundamental structure for focusing images, has specific characteristics of transparency and refractive index, and contributes to about 70% of total refraction [28, 29]. A diagram of the human eye is shown in Fig. 1.
 Fig. 1.
Fig. 1.
Diagram of the human eye.
Corneal transparency results from a specific organization of cellular (epithelial cells, keratocytes, and endothelial cells) and acellular (collagen glycosaminoglycans) components. It comprises five layers: epithelium, Bowman layer, stroma, Descemet’s membrane, and endothelium [29].
The conjunctiva plays a crucial role in supporting and protecting the cornea. It consists of a surface layer of nonkeratinized stratified squamous epithelium covering a vascular stroma of loose connective tissue [29]. The bulbar conjunctiva loosely connects with the underlying Tenon’s capsule, which covers the sclera [28]. One particular characteristic of the conjunctiva is the presence in its structure of the goblet cells, which are apocrine cells producing mucine, a fundamental component of the tear film. The conjunctiva has an essential role in the immune system of the ocular surface, with the presence of the so-defined eye-associated lymphoid tissue (EALT) [30].
The ocular surface is exposed to the external environment and in contact with many different microorganisms and antigens. Mechanical protection and immune regulation are critical to preserving its anatomic and functional characteristics [30]. The immune balance has to be efficient against potentially pathogenic microorganisms but also finely regulated to respect the delicate structures of the ocular surface.
The tear film has several crucial functions. It is the first thing seen on the ocular surface, nourishing the cornea. Therefore, its integrity and smooth constitution are essential for proper image focusing. It is also the first structure to come in contact with antigens and microorganisms, with a significant role as a buffer and containing the normal ocular microbiota’s antimicrobial and anti-inflammatory mediators [30].
The tear film traditionally consists of three layers: mucin, aqueous, and lipid [31]. It is now known that the first two have mixing levels, creating a gel structure overlayed by the lipid layer. The ocular anatomic surface and the overlying tear film are highly interdependent. Their proper functioning is vital not only for guaranteeing the integrity of the refractive pathway but also as they represent the environment for the residency of ocular microbiota, affecting those variations that can ultimately affect eye pathologic conditions such as glaucoma.
Previous studies have demonstrated the interactions between ocular surface microbiota (OSM) composition and tear film proteome in humans [32]. Since approximately 80% of the most abundant human tear proteins (namely lactoferrin, lipocalin-1, lysozyme, and IgA) have antimicrobial activity, it has been suggested that the antimicrobial components of the tear film may influence the OSM load and composition and that an altered tear film, such as in dry eye syndrome or in contact lens (CL) wearing, may lead to an OSM dysbiosis [32]. Authors have hypothesized that the OSM, especially the Corynebacterium genus, may be involved in the metabolism of amino acids on the ocular surface (OS), influencing the composition of the tear film with amino acids [33]. Previous authors found that some microbes found on the OS are involved in the metabolism of inorganic ions and lipids, suggesting that the OSM may influence tear film stability through these pathways [32]. The clarity in defining the human microbiome has been challenged by semantic confusion, particularly surrounding terms like “microbiota” (referring to microbial taxa associated with humans) and “microbiome” (representing the collective genome of these microbes immersed in their environment), often used interchangeably [34, 35, 36, 37, 38].
The authors searched the PubMed database (https://pubmed.ncbi.nlm.nih.gov/) and the Cochrane Library (https://www.cochranelibrary.com/search) using the phrase “ocular microbiome glaucoma”. 0 records were returned from the Cochrane Library. The following PubMed search string was hereby created viz; (“ocular”[All Fields] OR “oculars”[All Fields]) AND (“microbiome s”[All Fields] OR “microbiomic”[All Fields] OR “microbiomics”[All Fields] OR “microbiota”[MeSH Terms] OR “microbiota”[All Fields] OR “microbiome”[All Fields] OR “microbiomes”[All Fields]) AND (“glaucoma”[MeSH Terms] OR “glaucoma”[All Fields] OR “glaucomas”[All Fields] OR “glaucoma s”[All Fields]). A total of 21 results were retrieved spanning from 2017 to 2024. This comprised of 1 opinion paper, 4 original research articles and 16 reviews. The corresponding author checked each article for relevance to the topic and content. A PRISMA [39] diagram showing the search procedure is shown below as Fig. 2.
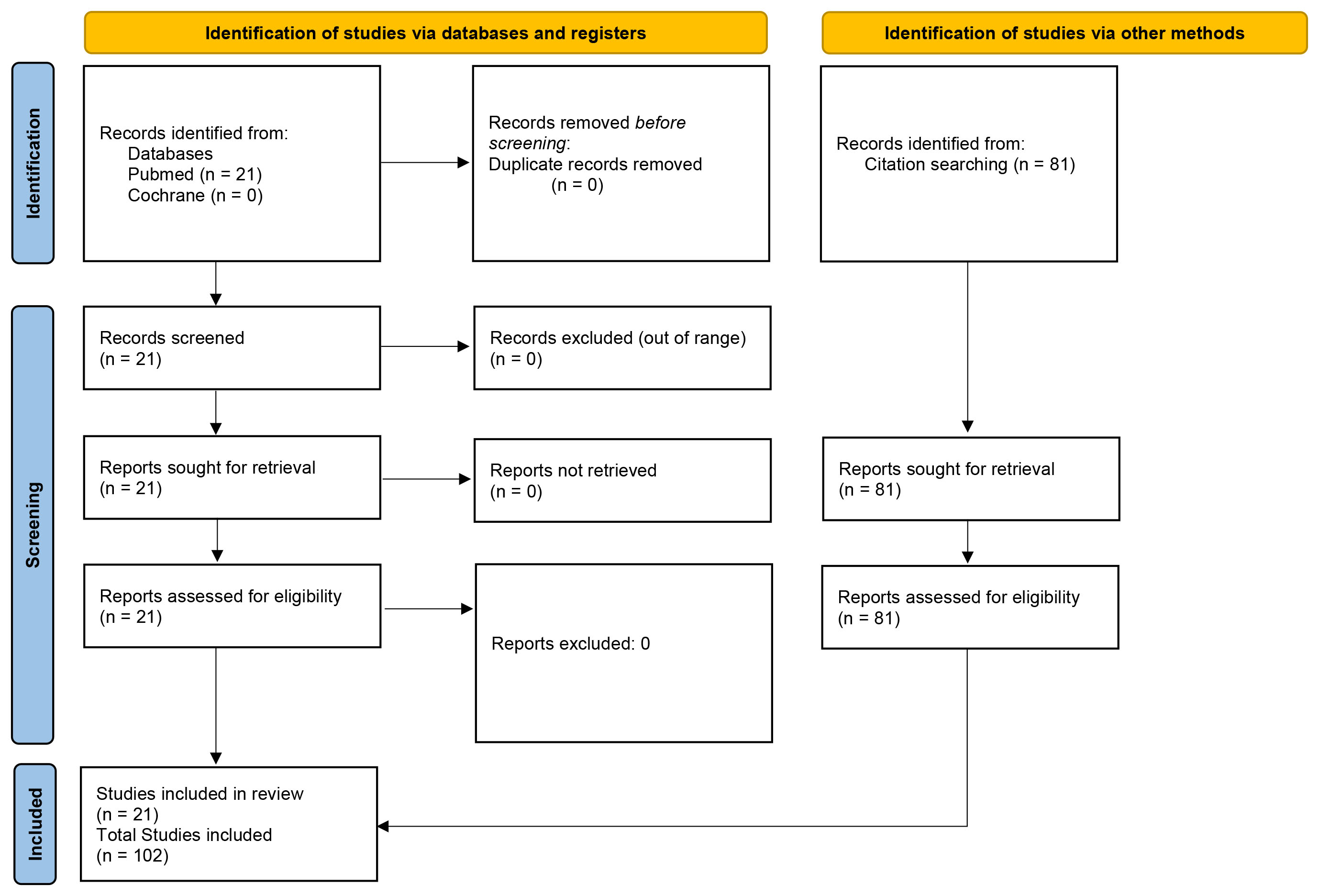 Fig. 2.
Fig. 2.
Preferred Reporting Items for Systematic reviews and Meta-Analyses (PRISMA) flow diagram of included studies.
Research has advanced significantly since the systemic microbiota were delineated in 2008, and the ocular surface microbiota was identified in 2011 [40, 41].
However, traditional microbiological techniques previously employed to characterize the ocular microbiota have limitations, with culture-based methods capturing only a fraction of its composition. Typically, less than 20% of bacteria from the environment, spanning various branches of the phylogenetic tree, can be successfully cultured in laboratory settings [42]. However, recent studies suggest that despite notable variations between samples, approximately 35 to 65% of molecularly detected species through sequencing have corresponding strains that can grow and propagate under laboratory conditions [43]. Classically, culture-based techniques for microbiome analysis involve growing microorganisms from a sample on various types of agar plates in the laboratory. These techniques have been widely used historically and are still employed in specific scenarios due to their ability to isolate and identify particular microorganisms. A schematic view of how the process typically works, starting from its application to the study of the ocular microbiota, is detailed below.
Sample Collection: In ocular microbiome studies, samples are collected from the ocular surface using swabs or other methods. These samples may include conjunctival swabs, tear fluid, or corneal scrapings.
Inoculation: The collected samples are streaked onto different agar plates, each designed to promote the growth of specific microorganisms. For example, blood agar plates can support aerobic and anaerobic bacteria growth, while selective media like MacConkey agar can selectively grow Gram-negative bacteria.
Incubation: The inoculated agar plates are then placed in an incubator set at the optimal temperature and conditions (aerobic or anaerobic) for bacterial growth. Incubation times can vary depending on the types of microorganisms being targeted.
Colonial Morphology: After incubation, the plates are examined for bacterial colonies. Each colony represents a population of bacterial cells derived from a single bacterial species.
Subculturing: If necessary, individual colonies can be subcultured onto fresh agar plates to obtain purer cultures for further analysis or identification.
Identification: Once the colonies have grown, various techniques can be employed to identify the bacterial species present. This can include visual inspection of colony morphology, biochemical tests, and, in some cases, molecular techniques such as polymerase chain reaction (PCR) or sequencing of specific genes.
Understanding the human microbiota, especially in the gut, mouth, and skin, has advanced significantly in 21st-century medicine. New techniques like polymerase chain reaction (PCR) and 16S ribosomal DNA sequencing have revealed a diverse community of microbes in various body sites, far more extensive than previously thought. Dysregulation of the gut microbiome has been linked to multiple pathologies like Clostridium difficile colitis, irritable bowel syndrome, and inflammatory bowel disease, suggesting therapeutic potential through fecal microbiota transplantation [44].
In Ophthalmology, there’s an ongoing debate about whether the ocular surface hosts a resident microbiota and its role in ocular physiology [45]. Studies using modern techniques have revealed a potential core ocular surface microbiome, but its nature and stability are still uncertain. Viruses and fungi on the ocular surface have also been observed, raising questions about their significance.
Understanding the ocular surface microbiome is crucial for addressing idiopathic ocular surface disorders with inflammatory components. A suggested framework for characterizing the ocular surface microbiome involves careful exclusion of contamination in molecular studies, distinguishing between viable and nonviable organisms, assessing stability over time, and generating testable hypotheses about microbial roles in health and disease. Applying high-throughput DNA sequencing techniques appropriately is essential for advancing ocular microbiology [44].
However, cultivating microbes in controlled conditions remains challenging due to various factors. Some low-abundant and slow-growing microorganisms struggle to thrive as more abundant and faster-growing species often outcompete them. Additionally, certain microbes fail to grow on standard culture media due to unfavorable conditions such as pH levels, redox state, temperature, or nutrient availability. Complex interactions, like interspecies electron transfer observed in symbiotic relationships facilitating organic matter decomposition, highlight the intricate metabolic connections in natural habitats [44]. Several strategies have been devised to culture previously unculturable microorganisms, mainly from environmental microbiology. These methods included mixed culture or cocultivation with helper strains to enhance growth [45, 46]. Moreover, the addition of signaling molecules like cAMP or homoserine lactones and cell-free supernatants have successfully promoted the development of previously unculturable microbes [47]. Another successful approach involves mimicking the native environment using diffusion chambers, enabling the growth of bacterial species that were previously difficult to culture [48].
In ocular microbiome analysis, culture-based techniques have been used to identify and characterize bacterial species inhabiting the ocular surface. Studies have cultured bacteria from conjunctival swabs or other ocular samples to investigate the diversity and composition of the ocular microbiota in healthy individuals and those with ocular diseases such as conjunctivitis or keratitis. While culture-based methods provide valuable information about cultivable microorganisms, they have limitations, such as the inability to culture all organisms in a sample and biases towards certain types of bacteria. Therefore, culture-independent methods, such as DNA-based techniques, have replaced traditional culture-dependent methods, allowing for a deeper understanding of microbial communities’ taxonomic composition and functional metagenomics. These methods, particularly those based on 16S ribosomal RNA gene sequencing, have revolutionized microbial ecology studies, providing insights into microbial diversity and relationships [49]. Large-scale research initiatives like the MetaHIT project [50] and the Human Microbiome Project [51] have further propelled microbiome research, shedding light on the microbial composition of healthy individuals and diseased states.
Despite the advantages of culture-independent methods based on 16S rDNA sequencing, these methods also have limitations, including the underestimation of low-abundant organisms and the inability to detect intraspecies variations [52]. Culture-based approaches are being used again to address these limitations, but in a different context. Hence, culturomics has been developed to cultivate previously unculturable microorganisms.
Culturomic Analysis
Culturomics, introduced in 2012 by the group of Didier Raoult and Jean-Christophe Lagier, represents a sophisticated strategy for microbial isolation and cultivation [53]. It has significantly enriched our understanding of the human microbiota, fueling databases like matrix assisted laser desorption ionization time-of-flight (MALDI-TOF) and microbial gene banks. Additionally, it has provided microbial entities for subsequent biochemical studies, enabling the cultivation of diverse microbes previously neglected due to low abundance or biased culture-independent analyses [54]. As of 2021, the number of identified bacterial species in humans has reached 3253, with an increase of 477 species compared to 2018. A substantial 63% of this increment is attributed to culturomics [55]. Similarly, 66.2% of new species added in 2018 relative to 2015 were obtained through this method [56].
Generally, the culturomics procedure encompasses several steps, starting with sample collection. For gut microbiome analysis, feces are the most common choice due to their convenience, cost-effectiveness, and reproducibility [57]. The sample is then processed by dilution and homogenization to facilitate microbial growth. Next, techniques like centrifugation and filtration are employed to separate microbes from other sample components. Isolated microbes are then cultured in specific media tailored to their growth requirements. Finally, bacterial identification is achieved through techniques like MALDI-TOF mass spectrometry or 16S rRNA sequencing. Identified microbial strains can be maintained in appropriate cultures for future studies.
Two primary approaches exist in culturomics. Non-targeted strategies aim to isolate and cultivate the broadest range of microorganisms possible. The processed sample is inoculated onto various media with different characteristics. After several days of cultivation, colonies of interest are selectively identified [53]. Targeted strategies leverage specific information about particular species to direct isolation and cultivation [58]. Information can be derived from scientific literature, metagenomic data, or other sources. Utilizing high-throughput cell sorting helps obtain the desired microorganisms with increased efficiency [59].
However, despite its contributions, culturomics also faces challenges. Non-targeted strategies can be labor-intensive and may not always capture desired taxa, leading to resource wastage [60]. Targeted strategies, while potentially more efficient, may face obstacles such as technical limitations and equipment setup complexity, particularly in anaerobic workstations, potentially hindering the isolation of certain microorganisms [61]. Additionally, the death of microorganisms before entering the culturomics procedure and unresolved symbiotic relationships pose further challenges [61, 62]. To address these constraints, culturomics procedures are continually optimized. The optimization of culturomics involves several key aspects, including sample collection and processing, microbial enrichment, anaerobic protection, sample preservation, microbial isolation, and cultivation techniques.
Concerning sample collection, while fecal sampling is convenient and widely used, it may not fully capture the microbial diversity, particularly from intestinal mucosa [63]. Endoscopic approaches offer a more direct sampling method but are costly and cumbersome [64]. Although technical and cost barriers remain, intelligent capsules have emerged as a promising alternative, enabling precise sampling from different intestine sites without invasion [65].
Sample processing is crucial for both culture-independent and culture-dependent studies of gut microbes. Microbial enrichment, achieved through selective conditions or media, enhances access to microbial resources, particularly for slow-growing species [66, 67]. Anaerobic protection is essential due to the gut’s anaerobic nature, and innovative methods like micro-particle systems have been developed to regulate dissolved oxygen concentration, facilitating microbial culture [68, 69]. Proper sample preservation is vital for maintaining sample integrity, with various methods available depending on storage duration and conditions [70].
Microbial isolation involves techniques like two-dimensional cell separation and droplet-based approaches, which have shown promise in increasing microbial diversity captured from single-growth mediums [62, 71]. Gene-targeted microfluidic isolation and automated microbiome imaging and isolation (CAMII) leverage advanced technologies like machine learning and microfluidics to improve efficiency and reduce labor [72, 73]. Live-FISH and metabolite labeling offer targeted sorting approaches [74], while Raman-activated microbial cell sorting (RACS) allows for sorting based on metabolic activity using stable isotope probes [75]. Antibody labeling and gene-targeted microfluidic isolation enable sorting based on specific genes or epitopes [76].
In cultivation optimization, selecting suitable growth media is critical, with extensive testing helping identify optimal conditions for microbial growth [77, 78]. Coculture systems and single-cell cultivation techniques, facilitated by microfluidic devices, enhance the cultivation of symbiotic or slow-growing microorganisms [79, 80, 81]. Overall, these advancements in culturomics offer promising avenues for expanding our understanding of gut microbiota and unlocking the potential of previously unculturable organisms [73, 75].
Culturomics has unlocked new frontiers in human gut microbiome research, offering a key to understanding its complexity and relevance to health and disease. Culturomics has successfully isolated novel bacterial species and elucidated physiological host-microbe relationships. As demonstrated by targeted cultivation methods, integrating molecular and culture-based approaches offers a promising new strategy for further understanding microbial communities and their roles in health and disease [53]. The development of innovative technologies and optimization of culture procedures will continue to fuel this scientific revolution [72], holding significant implications for personalized medicine and treating diverse pathologies [82].
Next-generation sequencing is a valuable tool in ophthalmology, offering more profound insights into the microbial landscape of the ocular surface [83]. It involves the analysis of thousands of genetic units, usually in a shorter period, and is similar to whole genome sequencing [84, 85]. This technology can refine our understanding of ocular infections and improve the diagnostic precision of infectious diseases by enabling the simultaneous analysis of genome information from a plethora of microorganisms in clinical specimens compared to 16s RNA, which deals with a single organism at a time [86, 87, 88]. This comprehensive approach, metagenomics, is particularly beneficial for examining microbial communities. NGS’s rapid, agnostic characterization capabilities have positioned it as an indispensable tool in various clinical settings, offering a swift and cost-effective means of microbial assessment [89, 90].
3.1.2.1 Conjunctival Sac Flora Analysis in Pre-Surgical Patients
The exploration of conjunctival sac flora using NGS in patients poised for cataract surgery, particularly those of advanced age who have not been subjected to any preoperative medication, presents intriguing findings [91, 92, 93]. Notably, the NGS methodology paints a different microbial landscape than traditional cultivation techniques [94]. This discrepancy is illuminated by the prevalence of Pseudomonas DNA in the NGS results, a genus not typically dominant in conventional reports, offering a more intricate view of the ocular microbiome than previously understood [95, 96]. Such insights call into question our reliance on classical cultivation methods and suggest a potential shift in our understanding of the microbial populations in the conjunctival sac, particularly in the context of pre-surgical assessments [97]. A deeper discussion into the realm of NGS studies on the conjunctival sac microbiome reveals a tapestry of microbial composition that varies from one study to another. Dong et al. [41], for instance, identified an unclassified bacterium as the most copious in their samples, with Pseudomonas spp also prominently featured, suggesting a diverse microbial environment [41, 95]. However, the presence of Pseudomonas aeruginosa on the ocular surface, as detected by NGS, is the subject of an ongoing debate. The significance of this debate is amplified by the fact that Pseudomonas aeruginosa is an opportunistic pathogen associated with severe ocular infections. Two primary perspectives exist: one suggesting that NGS accurately identifies Pseudomonas aeruginosa as a genuine component of the ocular microbiome, and the other positing that its detection may result from contamination during the PCR amplification step of the NGS process. The contention highlights the challenges of NGS data interpretation and the importance of ensuring that sequencing results accurately represent the actual microbial communities, especially when these findings have potential clinical implications. This is the challenge of identifying organisms via DNA in low-biomass environments. DNA contamination from other more populated organisms in low biomass environments has also been reported as a challenge by Pollock et al. [95]. Conversely, Zhou et al.’s findings [96] resonate more with traditional cultivation-based studies, where common culprits such as Corynebacterium, Streptococcus, Propionibacterium, and Staphylococcus dominated, comprising a significant portion of the bacterial DNA. In addition, Huang et al.’s study [98] further complicated the narrative by highlighting Corynebacterium as the most abundant, followed by Pseudomonas, which aligns with the previously mentioned NGS findings but diverges from conventional cultivation results. These comparative analyses underscored the variability and complexity of the conjunctival sac microbiome and emphasized the need for a more standardized approach to truly understand the microbial dynamics at play, as well as their implications for ocular health, particularly in the surgical context.
3.1.2.2 NGS Implications for Diagnosis and Treatment of Ocular Infections
In the realm of ocular health, NGS has become a pivotal tool, casting light on the intricate biodiversity of the ocular surface microbiome. The variability observed in the NGS results is not just a testament to the complexity of the microbiome but also a clarion call for more sophisticated analysis techniques [99, 100]. The necessity for refined methods is underscored by the potential for an expanded spectrum of bacterial species residing in the conjunctival sac. This spectrum might have been underappreciated or overlooked by traditional cultivation-dependent methodologies [101, 102]. The distinct findings gleaned from NGS analyses have profound implications for clinical diagnostics and treating ocular infections. By utilizing NGS, clinicians, and researchers can detect a diverse array of bacterial DNA, encompassing species known to be recalcitrant to standard laboratory cultivation [103, 104, 105]. This enhanced sensitivity and comprehensiveness provided by NGS could pave the way for improved diagnostic accuracy, thereby facilitating the development of targeted and more effective treatment regimens for ocular infections. Furthermore, NGS can potentially revolutionize our understanding of the core microbial inhabitants of the ocular surface. Such an understanding is imperative for distinguishing pathogenic bacteria from benign commensal flora during clinical evaluations [106, 107]. By establishing a clearer picture of the ocular microbiome’s baseline state, NGS aids in discerning deviations that may signal infection, thus enabling prompt and appropriate therapeutic interventions. The clinical ramifications of these advancements are significant. With the aid of NGS, clinicians could potentially tailor antibiotics to the specific pathogens present. This strategy might reduce the occurrence of antibiotic resistance by avoiding broad-spectrum antibiotic use [100, 108]. Moreover, the ability to detect bacterial DNA from non-cultivable organisms might also lead to the discovery of novel pathogens and reevaluate the pathogenesis of ocular infections [109].
Metagenomics is the study of genetic material obtained from microorganisms collected in a specific environment, in the natural state, not deriving from isolation or culture [110, 111]. Such an approach focuses on microbial diversity and its consequences on the interactions and relationships with the environment. It is a rapidly expanding field with many applications, from medicine to agriculture, pollution control, energy, farming, and wine-making [112, 113]. While 99% of the microorganisms in the environment cannot be cultured [114], the metagenomic approach isolates an impressive amount of microbial genetic material, mostly from bacteria but also from viruses and fungi [51].
The extraction of metagenomic DNA is crucial, as it involves collecting all the microorganisms in the sample and maintaining the integrity and purity of the fragments, ideally with minimal contamination from the host DNA, especially when collecting samples from human biological districts.
Different approaches can be used to proceed with metagenomic characterization, from shotgun sequencing to NGS. The advent of Next-Generation Sequencing (NGS) technology has offered the possibility of performing large-scale sequencing of microorganisms’ genetic material (metagenomic NGS, mNGS) [115]. The possibility of splicing a large amount of genetic material quickly and cost-effectively is particularly important, considering that while the human genome contains about 20,000 genes, the total microbiome is estimated to contain about 8 million genes [51].
Functional metagenomics can provide valuable information about bioactive substances and new microbial functional genes by characterizing specific enzymes or their functional variations under particular physicochemical conditions. There are some limitations to this kind of investigation, essentially represented by the fact that less abundant microorganisms may be challenging to extract and by a potential loss of DNA fragments in the gene cloning process. One further critical aspect of the metagenomic approach is that most studies have been performed on samples collected in developed countries, while the microbiota present in developing countries is underexploited [111].
In perspective, metagenomics, primarily if combined with metatranscriptomics, proteomics, and metabolomics, could provide valuable insights in the elucidation of missing links in the complex scenario of glaucoma pathogenesis.
The ocular surface (OS) complex system includes corneal epithelium, conjunctiva, tear film, eyelids, lacrimal gland, and nasolacrimal duct. The OSM is defined as the whole of the resident commensal non-pathogenic and pathogenic microbes that colonize cornea, conjunctiva, and eyelid margins, including low abundance taxons and proteins, which are species that make up less than 1% of the total microbiome population [116]. Bigger molecules and more abundant species usually mask low-abundance microbes and proteomic markers. However, specific testing techniques like 16s-rRNA sequencing and metagenome shotgun sequencing testing can identify them [100]. The existence of a resident commensal ocular surface microbiome (OSM) has been questioned for a long time by the ophthalmological community. The OS, being a physiological barrier against the external environment and therefore continuously exposed to a variety of microbes, allergens, and toxins, has indeed developed several mechanisms that limit infections and allergies, including:
(i) the mechanical effect of eyelids blinking and tears washing;
(ii) the anti-microbial substances present in the tear film, such as lysozyme, lactoferrin, immunoglobulin A, lipocalin, mucins, defensins, interleukins, and other peptides that have shown the ability to kill or inhibit microbial growth or modulate the immune response of the OS;
(iii) the presence of a conjunctiva-associated lymphoid tissue (CALT), composed by immune competent cells and lymphocytes, that can trigger an innate or adaptive immune response;
(iv) and the corneal epithelial cells and keratocytes can release interleukins and defensins to recruit neutrophils [100, 115, 116, 117].
Although the first description of the OSM dates back to the 1930s [118], it is more recently, after the publication of the results of the Human Microbiome Project [51], that ophthalmologists have become increasingly interested in identifying the quantity and composition of the OSM and its role in ocular pathophysiology [101].
The definition of the abundance and composition of the OSM in healthy subjects is highly variable in different studies and strictly dependent on the methods used, which include cultured-dependent and the more recent culture-independent methods. The OSM identified in healthy subjects by the cultured-dependent techniques appears almost entirely composed of bacteria, mainly including the genera coagulase-negative Staphylococci, Staphylococcus aureus, Propionibacterium, Corynebacterium and Streptococcus; other microbes isolated from the OS include Diphtheroid bacteria, Microcossus, Escherichia, Enterococcus, Lactobacillus, Bacillus, Haemophilus, Neisseria, Pseudomonas and fungi [118, 119, 120]. Low-abundance bacteria such as Chryseobacterium, Rothia, Massilia, Moraxella, Neisseria, Paracoccu, and Ralstonia exist among the OSM [100]. The most common OS bacteria are Gram-positive coagulase-negative Staphylococci, which are present in 20–80% of the conjunctival swabs and in 30–100% of the lid swabs [118, 119, 120]. No difference between fellow eyes was found in the OSM using the traditional culture-dependent techniques [118, 119, 120].
The overall positivity rate for fungi on the healthy OS based on the culture-dependent techniques ranges between 3 and 28% in the different studies [121]. Fungi identified on the OS with the culture-dependent methods include: Alternaria sp, Fusarium sp, Aspergillus niger, Aspergillus flavus, Curvularia sp., Penicillium sp., Helmintosporium sp., Candida albicans, Candida guilliermondii, Candida parapsilosis, Saccharomices cerevisiae, Hormodendrum sp., Rhodotorula rubra [121].
Although culture-based methods have the advantage of isolating living microbes only [122], they are time-consuming and unable to quantify and to precisely describe the composition of complex microbiomes [122]. Indeed, these traditional techniques only allow the identification of bacteria and fungi and their classification by the genus level according to their physiochemical characteristics. Moreover, they introduce a bias towards more easy cultivable species in standardized laboratory conditions. Still, cultivable species have been demonstrated to represent only a tiny proportion of the natural microbial populations present in the samples [122].
In 2011, Dong et al. [41] provided the first study about the OSM of healthy subjects using the amplicon sequencing method. Later, several other authors have studied the OSM using this technique [122, 123, 124, 125, 126].
The investigation of the healthy OSM with the 16S rRNA amplicon sequencing method has led to the identification of more than 600 different genera of bacteria belonging to 25 different phyla, with an average of more than 200 species of bacteria per subject [123, 124, 125, 126]. Anyway, it has been demonstrated that approximately 95% of the bacteria present on the OS can be classified into 4 phyla, i.e., Proteobacteria (65%), Firmicutes (4–15%), Actinobateria (15–20%) and Bacteroides (1–5%); and that 14 genera account for the 96% of the OSM of healthy subjects, mainly including Corynebacterium, Streptococcus, Proprionibacterium, Bacillus, Staphylococcus, Acinetobacter, Pseudomonas and Ralsontia, followed by the less common genera Streptophyta, Methylobacterium, Bradyrhizobium, Brevindimonas, Aquabecterium and Sphyngomonas. Pathogens, such as Pseudomonas aeruginosa or methicillin-resistant Staphylococci, can be also sporadically found on the OS of healthy individuals [123, 124, 125, 126, 127, 128, 129, 130].
Although some authors suggest that the core of the OSM in adult healthy subjects may be represented by the bacteria of the genera Corynebacterium, Acinetobacter, Staphylococcus, Pseudomonas, Proprionibacterium and Streptococcus [123, 125, 127, 129], the high inter-individual and inter-study variability found in the OSM composition when the bacteria classification is led to a genus level makes the existence of stable and unique OSM core still controversial [123, 127, 131].
The Corynebacterium has been found to be the most abundant germ present in the OSM of healthy subjects, systematically detected in all studies, even if is not detected in all subjects examined [123, 124, 125, 126, 127, 128, 129, 130].
Corynebacterium, Staphylococcus, and Proprionibacterium are also part of the normal periocular skin nasal cavity microbiome, which may suggest a possible transition, due to the anatomical contiguity, between normal skin and ocular surface microbiota [123, 124, 125, 126, 127, 128, 129, 130].
Moreover, sampling bias related to touching the periocular skin while sampling the conjunctiva cannot be excluded.
The fungi represent a minor proportion of the OSM of the healthy eye. Traditional culture-dependent techniques and genome sequencing methods have provided discordant results, considering that many fungi are not cultivable or not classifiable, so the culture-dependent methods showed a lower positivity rate (% of samples in which fungi are detected) and lower spectrum of fungi as compared to sequencing methods [121]. Comparing the results obtained with culture-dependent and sequencing methods in the same healthy subjects, previous authors have demonstrated that culture methods may detect at least four fungal genera with a positivity rate of 9–12%, whereas the amplicon sequencing method was able to detect between 65 and 94 genera, with a positivity rate of 40–74% [121].
Amplicon sequencing studies about the OS’s fungal microbiome identified fungi in 40–73% of the swabs [121, 132]. The most prevalent fungal phyla were Ascomycota, Basidiomycota, Zygomycota and Neocallimastigomycota, with Basidiomycota and Ascomycota resulting the most abundant species on healthy eyes [121, 132]. Although 94 fungal genera were isolated, 5 genera, including Aspergillus, identified as the most abundant genus, Malassezia, Rhodotorula, Davidiella, Alternaria, represented more than 80% of the fungal microbiome in more than 80% of the tested subjects and may be considered as the “core fungal taxa” on the normal OS [121, 132]. Anyway, the relative abundance and distribution of the fungal genera varied significantly amongst individuals and in different studies [121, 132].
The ability of the metagenomic sequencing technique to analyze both OSM and human host cells represents a challenge in OSM identification, considering that, in conjunctival swabs, the average percentage of non-human reads represents only 3.5% [123], so the relative abundance of human versus microbial mass makes difficult the proper assessment of the microbial component. The high sensitivity of this method has the limitation of being exposed to short random DNA contaminations, which may lead to artefactual identification of microbes; moreover, this method is costly, time-consuming, and requires complex data analysis [123]. For these reasons, most current published studies about the OSM are based on amplicon sequencing.
Studies using whole-metagenome shotgun sequencing showed that the OSM of a healthy eye is composed of the great majority of bacteria (median 93%, range of 78–98%), followed by viruses (median of 1%, range of 1–7%) and fungi or other eukaryotes (median of 4%, range of 0.02–20%) [123, 133, 134].
The bacterial DNA was detected in all subjects, whereas viruses and fungi were identified in 41% and 35% of subjects, respectively [135]. The dominant bacterial phyla were Actinobacteria, Firmicutes and Proteobacteria, whereas the most abundant genera were Propionibacterium, Agrobacterium and Corynebacterium [134].
Viruses are the second most common kingdom on the healthy OS, representing
approximately the 5% of the commensal microbes of the OSM and found in
approximately 40% of samples [123, 133, 134, 135, 136]. The presence of viruses on
the OS of healthy subjects has been studied using the metagenomic sequencing, the
Biome representational in silico karyotyping (BRiSK) and viral-detected PCR, that
can detect only DNA viruses, whereas the RNA viruses require the RNA sequencing
to be identified, which has not been addressed in studies to date, so that poor
information is currently available on the OS virome composition and function. The
viruses found on the healthy OS include phages, i.e., Bacteria-infecting viruses,
of the genera Siphoviridae, and the genera Anelloviridae,
Alphatorquevirus, Multiple Sclerosis-associated retrovirus,
Human endogenous retrovirus K, Human papilloma virus,
Abelson murine leukemia virus and Merkel cell polyomavirus. The
presence of phages on the OS, that are one of the main regulators of the
bacterial population density and distribution, may suggest the role of the
viruses in maintaining the OSM homeostasis. The meaning of the presence of
pathogenic and oncogenic viruses on the healthy OS is still unclear and requires
further explanations [135]. Table 1 (Ref. [121, 132, 134, 135, 137, 138]) summarizes
the composition of the OSM in healthy subjects. Microbes with a relative
frequency of
| Microbial type | Phyla and/or genera |
|---|---|
| Bacteria | Phyla: Proteobacteria, Firmicutes, Actinobateria, Bacteroides [134] |
| Genera: Corynebacterium, Streptococcus, Proprionibacterium, Bacillus, Staphylococcus, Acinetobacter, Pseudomonas, Ralsontia, Streptophyta, Anaerococcus, Neisseria, Veillonella, Prevotella, Actinomyces, Rothia, Lactobacillus, Finegoldia, Gemella, Methylobacterium, Bradyrhizobium, Brevindimonas, Aquabecterium, Sphyngomonas, Pseudomonas aeruginosa, methicillin-resistant Staphylococci [137, 138] | |
| Viruses | Genera: Siphoviridae, Anelloviridae, Alphatorquevirus, Multiple Sclerosis-associated retrovirus, Human endogenous retrovirus K, Human papilloma virus, Abelson murine leukemia virus, Merkel cell polyomavirus [134, 135] |
| Fungi | Phyla: Ascomycota, Basidiomycota, Zygomycota and Neocallimastigomycota [121, 132] |
| Genera: Aspergillus, Malassezia, Rhodotorula, Davidiella, Alternaria [121] |
The normal OSM represents a dynamic ecosystem, and its abundance and composition
have been shown to be influenced by several host, environmental, and iatrogenic
factors, which include. Previous authors have found that infants born by vaginal
delivery had an OSM similar to that found in the uterine cervix (i.e., rich of
Lactobacillus, Bifidobacterium, Escherichia coli,
Staphylococcus epidermidis, Bacteroides and
Diphtheroides), whereas infants born via cesarean section showed
conjunctival bacteria more similar to those of the normal facial skin (i.e.,
Propionibacterium, Corynebacterium, Diphtheroids,
Staphylococcus epidermidis) [137]. Richness refers to the abundance of
different species in the area of study. Data on the OSM richness and composition
in different age groups are contradictory, so it isn’t easy to draw conclusions.
Some authors found higher bacterial richness and diversity of the healthy OSM in
the pediatric population (i.e., in subjects younger than 18 years) as compared to
adults [138]. On the contrary, other authors found a larger bacterial amount in
subjects older than 60 years compared to those younger than 30 years [133, 136] or
no age-related differences [126]. Studies analyzing the OSM in healthy subjects
aged from 3 to 90 years have shown a prevalence of aerobic cocci and
Propionibacterium in the younger age groups and an increased proportion of
anaerobic cocci and Corynebacterium in older subjects [137]. Comparing
subjects younger and older than 18 years, other authors found that the phyla
Proteobacteria, Firmicutes, Bacteroides and
Fusobacteria and the genera Streptococcus,
Staphylococcus epidermidis and Brachybacterium were
prevalent in the pediatric population. In contrast, the phylum
Actinobacteria and the genera Corynebacterium,
Propionibacterium, Paracoccus, Streptococcus,
Micrococcus and Staphylococcus aureus were more abundant in
adults [136, 138]. Moreover, previous authors found that the microbiome of the
adult population was significantly richer in several antibiotic-resistance genes
than children [136]. On the other hand, the fungal OS microbiota appears to be
significantly richer in younger and older age groups as compared to ages between
30 and 50 years [132]. Although several studies did not find significant
gender-related differences in richness or diversity of the OSM in healthy
subjects [133, 138, 139], other authors noticed distinct OSM profiles based on sex,
especially at the genus level, with a relative decrease of
Propionibacterium acnes, Anaerococcus and
Staphylococcus epidermidis and higher abundance of
Acinetobacter, Enterobacteriaceae and Escherichia coli
in post-menopausal females as compared to males and younger females
[123, 126, 136]. The post-menopausal hormonal imbalance has been, therefore,
demonstrated to induce a profound modification of the OSM composition, which is
thought to incline old women to several inflammatory, autoimmune, and allergic
diseases of the ocular surface, including dry eye syndrome [34]. On the other
hand, the fungal microbiota showed no gender differences [132]. Differences in
the OSM composition have been identified amongst different ethnicities, with
higher similarities between Asian, Unite States and European populations [123],
whereas Australian subjects showed a lower abundance of Actinobacteria
and a higher presence of Firmicutes [126], and a Gambian cohort had a
higher prevalence of Actinobateria and Firmicutes [139]. The
differences amongst populations may suggest some dependence of the OSM on the
ethnic and environmental background of the host [123]. Differences in temperature
and humidity may alter the OSM composition. Previous authors found a
significantly higher abundance and diversity of the OSM in children
The ocular surface and its microbiome seem to form a physiological community, where the OSM is thought to play an important role in the maintenance of the OS homeostasis and the prevention of pathogens proliferation [17, 19, 111, 115, 116, 117, 127, 129, 130, 131, 141, 148, 149]. Although the interaction between OSM, ocular surface components and the immune system is complex and yet not wholly elucidated, some points need to be highlighted viz:
(a) The immune system tolerates the commensal OSM: it is well demonstrated that,
in the presence of potential pathogen antigens, the OS epithelium produces
pro-inflammatory cytokines, chemokines, growth factors, adhesion molecules, and
interferons that can activate the innate and adaptive immune system cells present
on the OS. These include macrophages, dendritic cells, neutrophils, and
lymphocytes, which can elicit an inflammatory and immune response against
pathogens [115, 116, 117]. The commensal microbes are a large source of intrinsic
antigens that continuously stimulate the immune system but usually do not elicit
inflammation or immune response. The immune system’s tolerance against the
commensal OSM is called “immune silence”. It may result from different
mechanisms, being still not completely elucidated, including the presence of a
physical barrier separating the OSM from the host epithelium, composed of tear
film, antimicrobial peptides, immunoglobulins, and immune cells, is thought to
prevent excessive innate or adaptive immune reactions of the host immune
competent tissue against the OSM [115, 116, 117, 149]. In vitro, studies
have demonstrated that when exposed to pathogens, such as Pseudomonas aeruginosa,
healthy cultured corneal and conjunctival epithelial cells can produce
pro-inflammatory cytokines (including interleukin (IL)1
(b) Anti-infective and immunomodulatory roles of the OSM on the OS: animal model
and clinical studies suggest the existence of a crosstalk between OSM and both
local and systemic innate and adaptive immune systems and a role of the OSM in
the defense against pathogens. The most important studies in this field include
germ-free mice predisposed to more severe Pseudomonas aeruginosa
keratitis, showing a significantly lower level of secretory IgA, complement
components, and of IL-1
(c) The intraocular microbiome in healthy eyes: although historically, the human intraocular environment of healthy subjects was always considered to be sterile [166, 167], recent studies have suggested the existence of an intraocular commensal microbiota mainly composed of rare and anaerobic microbes which are extremely difficult to culture [168, 169]. Deng et al. [169] have demonstrated that commensal microbes may be a part of the retinal ecosystem, influence the retinal microenvironment, and regulate the retinal immune response. The 16S rRNA amplicon sequencing method allowed the identification of some bacteria (Proprionibacterium acnei, Moraxella catarrhalis and Massilia timonae) in 3 samples of 69 donor eyes unsuitable for corneal transplantation [168]. More recently, the metagenomic sequencing analysis of the anterior aqueous humor specimens from 1000 cataract patients found 134 different bacterial species, including Enterococcus faecalis and Staphylococcus epidermidis, of which the Propionibacterium acnes was the most abundant [169]. The authors suggested that the intraocular microbes may originate from the gut microbiota passing through the mucosal barriers into the blood or lymphatic system in cases of increased intestinal and ocular-blood barrier permeability.
Recent scientific evidence has linked dysbiosis of the human microbiome with the onset and progression of glaucoma [109, 111, 165, 170, 171, 172, 173, 174, 175, 176].
The existence of ocular hypertensive and regular tension glaucoma patients has indeed underlined that the intraocular pressure (IOP) elevation, although considered the most crucial risk factor for the onset and progression of glaucomatous optic neuropathy (GON), is neither necessary nor sufficient to cause the retinal ganglion cells (RGCs) death in glaucoma. The hereditability of POAG, calculated by the concordant rate between identical twin pairs, is low (13%), suggesting that other non-genetic factors, such as age, ethnicity, sex, lifestyle, diet, smoking, obesity, depression, may be significant risk factors for the disease development [177]. Glaucoma should now be regarded as a complex multifactorial neurodegenerative disease involving IOP-dependent and independent pathways, including vascular, metabolic, oxidative, neurotrophic, immune, and inflammatory components, where the inflammation seems to play a crucial role in the pathophysiology of the RGCs glaucomatous damage [178].
The results of recent experimental and animal model studies and several clinical observations suggest that the dysbiosis of the commensal flora can trigger metabolic, immune, and inflammatory pathways recognized as possible pathogenetic mechanisms of glaucoma [110, 112, 114, 165, 174, 175]. The large majority of the available scientific data have associated the prevalence of primary open-angle glaucoma (POAG) and, less frequently, with that of normal tension glaucoma (NTG) and primary chronic angle-closure glaucoma (PACG), with dysbiosis of same extraocular sites, including:
(a) the oral microbiome dysbiosis: the oral microbiota, whose richness and diversity are second only to the gut microbiome, includes several microbes [101]. The most common bacterial species of the oral microbiota are Firmicutes, Bacteroides, Actinobacteria, Fusobacteria, Proteobacteria, and Spirochaetes [170]. A strong link between glaucoma, worse oral health, tooth loss and periodontitis has been found [170];
(b) the gastric microbiome dysbiosis: the gastric commensal bacterial community mainly comprises the phyla Proteobacteria, Firmicutes, Bacteroides, Actinobacteria and Fusobacteria [179]. Helicobacter pylori is a gram-negative bacterium and an obligatory colonized organism of the gastric mucosa. The H. pylori gastric infection affects over 50% of the world’s population [179]. Although the issue is still debated, several authors have reported a statistically significant association between POAG or NTG and chronic gastric colonization by Helicobacter pylori [179, 180], which has been confirmed by recent meta-analyses [171, 172].
(c) the gut microbiome (GM) dysbiosis: Studies have underscored the therapeutic potential of interventions to restore gut microbiota balance. Probiotics, prebiotics, dietary modifications, and fecal microbiota transplantation (FMT) have shown promise in addressing dysbiosis and its associated health consequences. Moreover, advancements in sequencing technologies have enabled a deeper understanding of the gut microbiome composition and its functional implications, paving the way for personalized approaches to dysbiosis management [181]. However, further research is needed to elucidate the intricate mechanisms underlying dysbiosis and to develop targeted interventions that can effectively restore microbial equilibrium and promote overall health and well-being. By addressing gut dysbiosis, healthcare practitioners may unlock new avenues for disease prevention and treatment, ultimately improving the quality of life for individuals affected by these conditions. The dysbiosis of the GM has been indeed associated with several metabolic, inflammatory, autoimmune, and degenerative diseases, including irritable bowel syndrome, inflammatory bowel diseases, cardiovascular diseases, obesity, type 1 diabetes, chronic liver diseases, multiple sclerosis, rheumatoid arthritis, depression, autism, Parkinson’s disease and other neurodegenerative and neuropsychiatric diseases [181].
Both animal models and clinical studies have indicated a possible causal association between GM dysbiosis and the development and progression of several ophthalmic pathologies, including blepharitis, Meibomian gland syndrome, Sjogren-associated dry eye, infectious keratitis, scleritis, episcleritis, uveitis, age-related macular degeneration, diabetic retinopathy and glaucoma [110, 112], suggesting the existence of a gut-eye axis [112, 113, 114], having a role in the glaucoma pathogenesis [173, 174, 175, 176, 182].
Glaucomatous eyes have a different ocular OSM composition, with the prevalence of Gram-negative microbes. Several studies support this finding using various methods. The results showed that the healthy eye OSM were characterized by a prevalence of aerobic Gram-positive bacteria, with Actinobacteria, Firmicutes, and Deinococcota being the most abundant phyla, and Corynebacterium, Cutibacterium, Blautia, and Gordonia the prevalent genera. On the other hand, both eyes of the glaucomatous patients, regardless of the use of anti-glaucoma eyedrops, showed a greater microbial diversity and a higher relative abundance of anaerobic Gram-negative microbes, with a prevalence of the phyla Firmicutes, Proteobacteria and Verrucomicrobiota, and the genera Akkermansia, Faecalisbacterium, Lachnospiraceae, Komagataeibacter and Clostridia. Only the phyla Actinobacteria were significantly lower in glaucomatous eyes treated with eyedrops than in healthy and untreated glaucomatous eyes [143]. The authors hypothesized that the OSM differences between glaucomatous and healthy eyes may be related to using hypotensive eyedrops rather than to the glaucoma diagnosis. They suggested that eyedrop components instilled in one eye or microbes may migrate to the fellow eye by eye-rubbing or via the lacrimal and nasal mucosa linking the fellow eye’s OSM. Furthermore, considering that different hypotensive active medications lead to similar OSM alteration, the dysbiosis should likely be related to the ingredient common to all eyedrops, i.e., the preservative benzalkonium chloride (BAK) [143].
The Gram-positive communities’ disruption and the Gram-negative microbes’ prevalence may favor the infection and inflammation of the OS. Animal model studies have demonstrated that some common Gram-positive commensals of the OS, such as the Corynebacterium mastitidis and Streptococcus, can stimulate the conjunctival T-cells to produce IL-17, leading to the recruitment of a greater number of neutrophils and a higher synthesis of antimicrobial substances into the tear film, enhancing the defense against pathogens [155]. Moreover, the prevalence of Gram-negative organisms on the OS has been associated with increased frequency of infective keratitis in CL-wearers [157], suggesting replacing Gram-positive with Gram-negative microbes may predispose patients to OS infections.
Glaucomatous eyes have a different OSM metabolite composition. Metagenomic sequencing studies have demonstrated that the OSM of glaucomatous patients, regardless of the use of hypotensive eyedrops, was associated with an increased synthesis of lipopolysaccharides (LPS) and anaerobic unsaturated and saturated fatty acids and with an enhanced metabolism of hydrogen sulfide and sulfate and that these metabolic pathways were related to the level of the genera Akkermansia and Lachnospiraceae; on the other hand, healthy control eyes showed a prevalence of carbohydrate synthesis, glycolysis and oxidative phosphorylation, and these metabolic pathways were linked with the abundance Corynebacterium and Cutibacterium [143, 183].
The LPS is a component of the microbial cytoplasmic membranes and it acts as an endotoxin because it is recognized by the Toll-like receptor of the human immune cells. On the OS, the LPS has been demonstrated to interact with the Toll-like receptor 4, stimulating the corneal and conjunctival epithelial cells to produce pro-inflammatory cytokines (IL-1beta, TNF-alfa) [184] and also activating the retinal complement components and microglia [173].
Glaucomatous eyes have signs of OS inflammation and an altered tear film composition, especially those treated with BAK-preserved eyedrops. Chang et al. [146] have demonstrated that eyes exposed to BAK-preserved eyedrops had a significantly lower tear meniscus height and tear break-up time compared with both healthy controls and untreated glaucomatous eyes [143], and that tear film alterations were directly related with the levels of some OSM Gram-negative microbes, including Akkermansia, Lachnospiraceae and Clostridia [143].
Although several studies have found an altered OS microbial flora in glaucomatous patients, it is still unclear whether the OSM dysbiosis is related to the glaucoma diagnosis or to the use of eyedrops, especially those preserved with BAK [143]. BAK is the most frequently used preservative in glaucoma eyedrops and artificial tears, aiming to prevent the growth of pathogens in medication bottles [185]. It acts as a detergent disrupting the bacterial cell walls with consequent release of the cytoplasmic content and, at low concentrations, it primarily inhibits Gram-positive organisms [185]. BAK has been associated with tear film alterations and ocular surface inflammation that can affect the long-term therapy compliance [185]. Considering that the aqueous layer of the tear film is fundamental for the transfer of oxygen and nutrients across the OS, the tear film disruption caused by BAK is thought to reduce the oxygenation of the OS selecting preferentially Gram-negative anaerobes. Anyway, a recent study has found that BAK had no effects on the OSM of glaucoma patients [186].
(d) Intraocular microbiome dysbiosis: Little is known about the existence of an intraocular microbiome and its involvement in the development of glaucoma. Some finding support this hypothesis, including:
- the histologic studies have found the presence of Helicobacter pylori coccoid in the trabecular meshwork and iris samples of POAG patients [187], suggesting that in the presence of a chronic inflammation, bacteria of the gastrointestinal tract may migrate and overpass the blood-ocular barriers;
- The metagenomic sequencing analysis of the anterior aqueous humor specimens from 1000 cataract patients found that cataract patients affected by glaucoma had a significantly higher level of Propionibacterium acnes and lower level of Staphylococcus warneri [169];
- The Posner-Schlossman Syndrome (PSS) is a unilateral eye disease characterized by acute anterior uveitis and ocular hypertension, classified as inflammatory glaucoma. Hypertensive anterior uveitis and Posner-Schlossman syndrome have been associated with the presence, in both aqueous humor and trabecular meshwork, of several viruses, including Cytomegaloviru (CMV) Herpes simplex virus, Varicalla-Zoster virus and Rubella virus, although CMV localization and uveitic cell infiltration seem to be the primary cause of the trabecular meshwork impairment in PSS [188].
This begs the question if there is a causal relationship between dysbiosis and glaucoma with respect to microbes, microbial metabolites or inflammatory and immune components.
Although the precise mechanisms linking dysbiosis and glaucoma is still unknown, experimental animal models and clinical studies have suggested that the prevalence of pathogens induced by the dysbiosis may cause a local subclinical chronic low-grade phlogosis, disruption of the epithelial barriers, increased vasopermeability, and final systemic endotoxemia, with migration of microbes, their metabolites, inflammatory mediators or aberrantly microbes-activated immune cells into the blood circulation. In the presence of a breakdown of the blood-retina barriers, all these components may enter the eye and reach the trabecular meshwork, the retina and the optic nerve [165, 174, 175].
(1) Direct dissemination of the microbes into the eye; Helicobacter pylori [87], and Propionibacterium acnes and Staphylococcus warneri [69] have been found in the intraocular tissues of patients with glaucoma; moreover, hypertensive anterior uveitis and Posner Schlossman syndrome have been associated with the presence, in both aqueous humor and trabecular meshwork, of several viruses, including Cytomegalovirus (CMV), Herpes simplex virus, Varicalla-Zoster virus and Rubella virus [88]. These microbes may damage trabecular meshwork, retinal ganglion cells (RGCs), and optic nerve (ON) axons directly or indirectly by the activation of inflammatory/immune cascades;
(2) Dissemination of microbiota components, microbial genome fragments, microbial metabolites, or aberrantly microbes-activated immune cells into the eye via blood circulation; the relationship between microbiome and host is mainly mediated by microbes-derived metabolites, the most important of which include short-chain fatty acids (SCFAs), bile acids, tryptamine and histamine, GABA, serotonin, and dopamine [101].
Several findings support this hypothesis:
(i) Activation of the lipopolysaccharide (LPS) and Toll-like receptors 4 (TLR4): glaucoma patients are characterized by increased lipopolysaccharide (LPS) synthesis of the microbiome of the OS [143] and of the gastrointestinal tract [174, 175]. The LPS is the major component of the outer membrane of the Gram-negative bacteria, and it acts as an endotoxin because it is recognized as a pathogen-associated molecular pattern (PAMP) by the pattern recognition receptors (PRRs), a surface protein of the antigen-presenting cells. The innate immune response of the eye, indeed, is mainly mediated by a subgroup of PRRs, the Toll-like receptors (TLRs), that are located on the OS, on corneal, conjunctival, and immune cells, and also in the retina on RPE cells, microglia, astrocytes, and Mueller cells. The TLRs recognize pathogen-associated molecular patterns (PAMPs), i.e., microbial structures/nucleic acid sequences, or damage-associated molecular patterns (DAMPs), that are usually molecules released from the host cells following a tissue damage. The LPS is known to bind specifically the Toll-like receptors 4 (TLRs4), that begins inflammatory and immune-response processes on the OS by up-regulating the secretion of pro-inflammatory cytokines (IL-1beta, TNF-alpha) by the corneal and conjunctival epithelial cells [184]. Moreover, animal models of glaucoma have demonstrated that the administration of a low-dose of subcutaneous LPS induced an up-regulation of the complement and the activation of the retinal and ONH microglia in the retina and optic nerve [173], that are thought to have a role in the RGCs and ON axons neurodegeneration [178]. Furthermore, TLR4 of retinal microglia and astrocytes, that respond to several endotoxins, including the bacterial LPS, are up-regulated in both glaucoma animal models and glaucomatous donor eyes [189]. Finally, several TRL4 polymorphisms have been associated with an increased risk of POAG and NTG [190];
(ii) The Heat shock proteins (HSPs) and auto-reactive CD4+ T cells activation: HSPs are highly conserved from bacteria to humans and seem to be involved in the autoimmune pathogenesis of glaucoma [191]. Mice models of hyperbaric glaucoma have demonstrated that increased IOP and exposure to the commensal microbes are both necessary to induce the up-regulation of CD4+ T cells that specifically target commensal bacterial HSPs (HSP27), cross-react with the same HSPs present on the host RGCs and ON axons, infiltrate the retina and ONH and induce RGCs death and ON damage. Chen et al. [191] suggested a two-step model of glaucoma pathogenesis: the auto-reactive CD4+ T cells are trained or pre-sensitized by the commensal resident micro flora to recognize bacterial antigens (HSPs) that can cross-react with RGCs antigens; secondarily, an elevated IOP (or another insult) may favor the CD4+ T cell entrance and infiltration into the retina and the optic nerve by damaging the blood-retinal barriers and stimulate the retinal cells to express stress factors that become the target for an immune response that lead the RGCs degeneration. This cascade of events can persist despite IOP normalization and is blocked in germ-free animals [191]. Moreover, another study has demonstrated that both glaucomatous patients and glaucomatous animal models show higher blood levels of auto-antibodies against specific heat shock proteins (HSP), such as anti-HSP-27, as compared to controls [192]; similarly, Chen et al. [191] found a significant increase of HSP-27 and HSP-60 responsive T cells in primary open-angle glaucoma (POAG) and normal tension glaucoma (NTG) patients when compared to controls;
(iii) The short-chain fatty acids (SCFAs): SCFAs are commonly considered as beneficial microbial metabolites for human health [101]. Recent animal model studies have demonstrated that SCFAs may induce the activation of the retinal and cerebral microglia, with consequent neuroinflammation and RGCs and neuron damage [173]. POAG patients have shown higher levels of SCFAs, especially propionate, isovalerate, butyrate, and caproate, in serum and in fecal specimens [173]. Butyrate has been implicated in lowering IOP in normotensive murine models [193].
(iv) The involvement of other microbiome metabolites: The chronic gastric infection of Helicobacter pylori has been strongly associated with glaucoma [171, 172]. Helicobacter pylori stimulates the gastric secretion of proinflammatory factors, including TNF-alpha, iNOS, endothelin-1, reactive oxygen species (ROS) and other pro-inflammatory cytokines; furthermore, it can cause a chronic atrophic gastritis that induces a reduced absorption of vitamin B12 and folate, with consequent hyperhomocysteinemia [194]. The migration of proinflammatory cytokines and homocysteine from the altered gastric mucosa and their accumulation into the blood circulation may induce a vascular endothelial dysfunction and enhance the blood-retinal barrier permeability. Several proinflammatory molecules may thus reach the retina, may activate the retinal microglia and its differentiation into phagocytic macrophages or may directly induce the apoptosis of the RGCs or cause vasoconstriction of the ONH vessels; on the other hand, the vascular endothelial dysfunction itself has been linked with the development of POAG and NTG [180, 194]. The GM metabolite trimethylamine N-oxide can induce the synthesis of a mutant myocilin protein, which is associated with the development of a juvenile-onset POAG [195].
(v) The presence auto-anti-bodies: previous studies have shown that glaucomatous patients had significantly higher of IgG anti-Helicobacter pylori in blood and aqueous as compared to control subjects, which appeared to correlate with OD damage [196]. Moreover, several authors have found an association between glaucoma diagnosis and the presence of blood and aqueous auto-antibodies auto-reactive T cells anti retinal and optic nerve antigens (glycosaminoglycans, myelin essential protein, vimentin, glial fibrillary acidic protein, retinal S-antigen retinaldehyde-binding protein) [197].
Considering the relevance of the microbiota (ocular surface or intestinal) for eye health, alternative approaches such as probiotics, their metabolites, and bacteriophage therapy have emerged as potential therapeutic methods for preventing and treating ocular diseases [198, 199, 200]. Probiotics, live microorganisms that confer health benefits when consumed in adequate amounts, and prebiotics, non-digestible fermentable oligosaccharides that promote beneficial bacteria growth, are being investigated for their potential in disease management [201]. Moreover, the intricate relationship between the gut microbiota and the immune system has garnered significant research interest in recent years. The human gut microbiota plays a crucial role in maintaining overall health and well-being, with a growing body of evidence highlighting its bidirectional relationship with the immune system. The gut microbiota can influence immune function through various mechanisms, including the production of metabolites, modulation of the intestinal barrier, and shaping of the immune response [202]. Conversely, the immune system can regulate the composition of the gut microbiota.
Along this line, probiotics have emerged as a promising avenue for influencing the gut microbiota. Evidence suggests that probiotics can modulate the composition of gut bacteria, potentially promoting the growth of beneficial bacterial strains while suppressing the growth of harmful ones [203]. This shift in the gut microbial landscape is hypothesized to contribute to enhanced immune function through several mechanisms. Firstly, probiotics may directly stimulate the production and activation of immune cells in the gut, thereby bolstering the first line of defense against invading pathogens and promoting tolerance towards harmless antigens while enhancing the immune response against genuine pathogens [204]. Secondly, they may enhance the gut barrier function, which is critical in preventing the entry of harmful substances and pathogens from the gut into the bloodstream. Probiotics achieve this by promoting mucus production and strengthening the tight junctions between intestinal epithelial cells [205]. Therefore, probiotics can modulate the gut microbiota and influence immunity, a valuable tool for promoting immune function and overall health.
Probiotic treatments have gained popularity as a dietary approach due to their temporary and non-invasive nature, which aids in sustaining a healthy immune system by enhancing gut health. Besides regulating immune responses, probiotics are believed to shield against physiological stress, inhibit pathogen invasion, adjust microbiota, and enhance the gut epithelium’s barrier function [206]. For instance, Lactobacilli have been shown to reduce neutrophil extracellular traps, while Bacillus fragilis offers protective effects against autoimmune diseases through its polysaccharide capsule [207, 208]. Ongoing research explores the efficacy of an eye drop postbiotic formulation containing Lactobacillus sakei in patients with dry eye disease. Patients were given either L. sakei ophthalmic bacterial lysate drops, oral probiotic capsules, and corresponding placebos. Results showed that active postbiotic drops significantly decreased dry eye disease (DED) signs and symptoms. Both active products were well tolerated. However, the oral intake of L. sakei as a probiotic capsule showed no significant effect on DED symptoms [209]. However, the specific metabolization of prebiotics in the gut by host microorganisms is believed to determine the growth and function of local bacteria, influencing human health [210]. Positive results have also been reported with their use in inflammatory pathologies [211]. Experimental studies suggest that probiotics and prebiotics could restore microbiota balance and modulate immune responses, potentially alleviating autoimmune symptoms without the side effects of immunosuppressive drugs [212]. Probiotics are believed to regulate the immune system, mitigate physiological stress, inhibit pathogen invasion, balance the microbiota, and enhance the integrity of the gut epithelium [202]. For instance, Lactobacilli have been observed to reduce the formation of neutrophil extracellular traps (NETs). NETs are crucial for fighting microbial invaders, but their excessive release can harm surrounding tissues, contributing to various diseases. Intestinal disorders, including chronic inflammatory bowel disease, are linked to gut microbiota changes and inflammation. Probiotics are known to benefit gut health, and this effect might occur because nonpathogenic, enteropathogenic, and probiotic bacteria affect NET formation. The probiotic Lactobacillus rhamnosus GG has been shown to inhibit NET formation induced by phorbol 12-myristate 13-acetate (PMA) and Staphylococcus aureus. Additionally, it reduced neutrophil oxidative stress and phagocytic activity while protecting against cell damage, suggesting a novel mechanism for probiotics to modulate local immune responses in the gut [213]. In another study, a probiotic mixture containing Streptococcus thermophilus ST10 (DSM 25246), Lactococcus lactis LLC02 (DSM 29536), and Lactobacillus delbrueckii (DSM 16606) significantly shortened the time for small chalazia to resolve compared to standard treatment alone [214]. Similarly, the well-known human symbiotic bacterium Bacteroides fragilis shields animals from intestinal ailments like ulcerative colitis. Its capsular polysaccharide TP2 is pivotal in decreasing inflammation, thereby contributing significantly to its protective effects [215]. Probiotics, known for their efficacy in alleviating allergic inflammation, were investigated for their potential in managing vernal keratoconjunctivitis (VKC). In a study involving seven patients with mild to moderate VKC, Lactobacillus Acidophilus eye drops were administered four times daily for four weeks. Significant improvements in both clinical signs and symptoms were observed after two weeks and further enhanced by the end of the treatment period. Photophobia was notably reduced at two weeks, while itching, tearing, conjunctival hyperemia, and chemosis showed significant improvement at four weeks. Additionally, down-regulation of ICAM-1 and TLR-4, indicative of reduced inflammation, was observed in patients with clinical improvement. This open pilot study suggests that probiotic eye drops could effectively improve signs and symptoms in VKC patients, warranting further double-blind controlled trials with larger sample sizes to confirm these effects [216]. Separately, probiotic therapy emerges as a promising alternative for preventing and treating corneal infections. An in vitro study aimed to evaluate the preventive effects of Lactobacillus reuteri and Bifidobacterium longum subsp. infantis against Pseudomonas aeruginosa infection in cultivated human corneal epithelial (HCE) cells. Pretreatment with L. reuteri or B. longum significantly protected HCE cells from infection, enhancing cell viability. Moreover, both probiotics exhibited anti-inflammatory activities, reducing inflammatory cytokine levels and restoring cellular homeostasis. This research underscored the protective effects of L. reuteri and B. longum in combating P. aeruginosa-induced corneal infections by preserving cell viability and modulating inflammatory responses [217]. Additionally, Bifidobacterium has been found to facilitate the production and utilization of short-chain fatty acids (SCFAs), which directly influence gut mucosal immunity [218] and alleviate the severity of experimental autoimmune uveitis (EAU) [219, 220]. Combining probiotics like Lactobacillus gasseri KS-13, Bifidobacterium bifidum G9-1, and B. longum MM-2 has shown promise in improving rhino-conjunctivitis during allergy season by increasing serum total IgE levels and the percentage of regulatory T cells (Tregs) [221]. Chisari and colleagues [222] studied patients with DED treated with Bifidobacteria and Lactobacillus and concluded that this therapy can improve DED. In another study, DED treatment with probiotics that contained Saccharomyces boulardii MUCL 53837 and Enterococcus faecium LMG S-28935, administered orally or topically, mixed with artificial tears produced a noticeable improvement in patients without significant complications [223, 224]. Lactobacillus has also been studied as a probiotic. In patients with type 2 diabetes, a significant decrease in low-density lipoprotein and cholesterol and better disease control were demonstrated [225, 226]. Studies on IRT5 treatment, a blend of five probiotic strains, indicate its potential to alleviate symptoms in autoimmune dry eye models by modulating immune processes and reducing pathogenic CD8+ T cells in drainage lymph nodes [227, 228]. Notably, Lactobacillus paracasei KW3110 has been found to enhance the function of retinal pigment epithelial (RPE) cells under chronic inflammation and safely alleviate eye fatigue in humans [229].
Phage therapy, characterized by high specificity and safety in humans, offers the potential to modulate microbiota-associated diseases [230]. Phages have shown promise in treating bacterial infections, including those causing keratitis and endophthalmitis [231, 232]. They also demonstrate holistic effects on the microbiome, influencing host immune responses and disease progression [233, 234].
Advancements in intraocular drug delivery, such as utilizing stable RNA nanoparticles derived from bacteriophages, may offer innovative approaches for ocular therapies [235]. However, despite the known involvement of the host microbiota in disease pathogenesis, mechanisms of action remain elusive, warranting further investigation into the intricate relationships between phages, microbiota, and hosts [230].
Accordingly, diet is known to play a crucial role in shaping the composition of the human gut microbiota within the gastrointestinal tract (GIT), leading to the production of secondary metabolites such as short-chain fatty acids, neurotransmitters, and antimicrobial peptides [236]. Prebiotics, such as galacto-oligosaccharides, fructans, inulin, and others, serve as nutrients for the gut microbiota, promoting the growth of beneficial bacteria and potentially aiding in the prevention or treatment of various conditions, including Crohn’s disease, skin allergies, cardiovascular diseases, and neurological disorders [237]. While research on prebiotics’ role in eye disorders is limited, innovative contact lens formulations releasing the prebiotic resveratrol have shown promise in reducing harmful ocular bacteria growth and local inflammation [238]. Hyaluronic acid may also act as a prebiotic, enhancing antioxidant capacity and modifying gut bacteria composition [239]. Clinical trials combining probiotics and prebiotics (symbiotics) have effectively reduced inflammatory markers and improved symptoms in uveitis and dry eye patients. These symbiotic formulations, including species like Lactobacillus, Bifidobacterium, and Streptococcus, have shown positive outcomes in increasing tear secretion and alleviating dry eye symptoms [224, 240, 241]. Moreover, fermented products containing prebiotic strains and dietary fibers show promise in alleviating immune system disorders and metabolic diseases, including cardiovascular disease [242]. B-group vitamins and vitamin E are identified as potential prebiotic candidates, suggesting their beneficial effects on gut microbiome composition and overall gut health [243, 244]. Furthermore, investigations into dietary supplements like Durvillaea antarctica highlight their potential in modulating gut microbiota composition and improving immune and cardiometabolic health [245]. Additionally, the role of vitamin D and its receptor in shaping the gut microbiota composition suggests a potential interplay between vitamin D, gut microbiota, and human health [246]. Lastly, studies exploring the effects of specific food supplements, such as a hop extract standardized in 8-Prenylnaringenin, on bone health and gut microbiome in postmenopausal women underscore the potential impact of food supplements on gut microbiota composition and its implications for various aspects of human health, including cardiovascular health, immune function, and aging-related conditions [247]. These findings collectively underscore the potential of food supplements as prebiotics in modulating the human gut microbiota and influencing various aspects of human health [248], potentially including also eye diseases such as POAG [249, 250, 251, 252] and NTG [253]. Moreover, patients must undergo pertinent testing, examinations, and management in the differential diagnosis of glaucoma and other ocular diseases to ensure the best treatment options and clinical outcomes [254, 255, 256].
In conclusion, our knowledge about human-associated microbial communities is still limited, but they have increased our understanding of human health and pathophysiology. The ocular microbiome research in healthy subjects and in different systemic and ocular pathologies is an emerging and interesting field. There is evidence that the OSM is involved in various host processes and that dysbiosis of the OSM is associated with the development and progression of several eye diseases. Further investigations with large sample sizes under standardized conditions are required to define a core microbiota, to understand the relationship between OSM and host OS, to find out the host, environmental, and iatrogenic factors that can influence the OSM load and composition, to define how the OSM may have a role in the OS health and disease. Finally, little is known about the possible presence of an intra-ocular microbiome. Recent studies have suggested the existence of an intraocular commensal microbiota, which is likely composed of rare and anaerobic microbes that are extremely difficult to culture. A better understanding of the role of the ocular microbiome in the host pathophysiology may lead to new therapeutic strategies targeting the human microbiome to prevent or treat ocular diseases.
The gut microbiota plays a crucial role in mammalian development and homeostasis, potentially influencing ocular health. Therefore, considering probiotics in clinical practice may offer a promising avenue for intervention. Probiotics could modulate dysbiosis and mitigate inflammatory responses, offering therapeutic benefits in managing ocular conditions. However, despite mounting evidence indicating a tight correlation between the microbiota and primary open-angle glaucoma (POAG), no preclinical or clinical data have yet demonstrated that interventions targeting the gut or ocular microbiota can influence the occurrence or progression of POAG. Both preclinical and clinical data have been generated regarding the role of food supplements in mitigating the progression of POAG and protecting retinal ganglion cells (RGC) from apoptotic death caused by glaucoma-related factors. While a direct effect of the components of these food supplements on glaucoma neuroprotection is described, it is also possible that these supplements may function as prebiotics, altering the microbial balance within the gut microbiota and promoting a healthier combination of gut microbes. For instance, a study compared the gut microbiota composition and metabolite profiles between glaucomatous rats and controls. Glaucomatous rats exhibited reduced microbial diversity, increased Firmicutes/Bacteroidetes ratio, Verrucomicrobia phylum, and certain bacterial genera like Romboutsia, Akkermansia, and Bacteroides, which correlated negatively with RGC survival. Metabolomic analysis identified 284 differentially expressed metabolites, with enrichment mainly in bile secretion pathways. Glutathione (GSH), an antioxidant metabolite, was negatively correlated with certain gut bacteria and positively correlated with RGC survival. Reduced GSH levels in glaucomatous rats’ blood were associated with specific bacterial genera and the Firmicutes/Bacteroidetes ratio. These findings suggest a potential link between the gut microbiota, metabolites, and glaucoma, highlighting the possibility of gut microbiota-targeted interventions for the disease. Therefore, food components influencing the presence of glutathione in the organism might also affect gut microbiota composition.
CG and MZ designed the research study. CG, MS, MM, FP, AM, GS and MZ performed the research. CG, MS, FD, MM, FP, AM, DR and MZ analyzed the data. CG, MS, MM, DR, FP, and MZ wrote the manuscript. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This research received no external funding.
The authors declare no conflict of interest. Dario Rusciano worked for Fidia Ophthalmics before retirement and declares no conflict of interest. Given his role as Guest Editor and Editorial Board member, Dario Rusciano had no involvement in the peer-review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process for this article was delegated to Adrian Gericke.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.