


 , Yongchun Cui 1,*
, Yongchun Cui 1,*1 Beijing Key Laboratory of Preclinical Research and Evaluation for Cardiovascular Implant Materials, Animal Experimental Center of Fuwai Hospital, State Key Laboratory of Cardiovascular Disease, and National Center for Cardiovascular Diseases, Chinese Academy of Medical Sciences & Peking Union Medical College, 102300 Beijing, China
2 Department of General Surgery & Pharmacy, The Affiliated Suqian First People's Hospital of Nanjing Medical University, 223800 Suqian, Jiangsu, China
3 Department of Colorectal Surgery, The First Affiliated Hospital of Nanjing Medical University, 210003 Nanjing, Jiangsu, China
†These authors contributed equally.
Abstract
Background: Vascular smooth muscle cell (VSMC) intimal migration,
proliferation, and phenotypic transformation from a contractile to a synthetic
state are hallmarks of the progression of atherosclerotic plaques. This study
aims to explore the effects of exosomes derived from M2 macrophages (ExoM2)
on the pathological changes of VSMCs in atherosclerosis (AS). Methods:
Cell Counting Kit-8 (CCK8) and wound healing assays were used to examine the impact of ExoM2 on
platelet-derived growth factor-BB (PDGF-BB)-induced VSMC
proliferation and migration, respectively. Western blotting was employed to
analyze changes in the expression levels of contractile markers (e.g.,
alpha-smooth muscle actin [
Keywords
- M2 macrophages
- exosomes
- atherosclerosis
- smooth muscle cells
Cardiovascular disease (CVD) is the leading cause of death worldwide, with
atherosclerosis (AS) recognized as the common pathological basis for fatal CVD
[1]. The formation of atherosclerotic plaques involves the activation of various
cells in the arterial intima, including endothelial cells, smooth muscle cells,
lymphocytes, and macrophages, which collectively contribute to a localized
inflammatory response [2]. Vascular smooth muscle cells (VSMCs) are key
components of atherosclerotic plaques and play a crucial role in AS progression
[3]. The proliferation and migration of VSMCs are critical events in AS
progression [4]. Additionally, VSMCs may undergo a phenotypic transformation from
a contractile state, characterized by markers such as alpha-smooth muscle actin
(
Macrophages in AS are heterogeneous and can be primarily polarized into M1 and M2 types [6]. M1 macrophages are pro-inflammatory and exacerbate AS by secreting inflammatory cytokines [7], while M2 macrophages are anti-inflammatory and maintain normal metabolic homeostasis [8]. The growth rate and stability of atherosclerotic plaques are positively correlated with pro-inflammatory M1 macrophages and negatively correlated with anti-inflammatory M2 macrophages. However, the effects and underlying mechanisms of M2 macrophages on the fate regulation of VSMCs during AS remain elusive.
Exosomes, ranging from 40 to 160 nm in diameter, are tiny membrane-enclosed vesicles produced by virtually all prokaryotic and eukaryotic cells and can be released into extracellular spaces through exocytosis [9]. Recently, exosomes have emerged as significant mediators of intercellular communication, transporting enclosed cargo like bioactive lipids, as well as noncoding RNAs, miRNAs, and proteins from donor to recipient cells [7, 8, 9, 10]. Significant research has investigated the role of exosomes derived from M2 macrophages (ExoM2) in diabetes [11], cancer [12], spinal cord injury [13], autoimmune diseases [14], and myocardial injury reperfusion [15].
However, the role of exosomes in the communication between M2 macrophages and VSMCs in AS remains unresolved. This study aims to explore the effects of ExoM2 on VSMC fate using an in vitro atherosclerotic VSMC model induced by platelet-derived growth factor-BB (PDGF-BB), and on the progression and stability of atherosclerotic plaques in high-fat diet ApoE-/- mice. Our findings will comprehensively elucidate the mechanisms through which M2 macrophages modulate atherosclerosis.
Twelve six-week-old male ApoE-/- mice (C57BL/6), weighing 20 to 22 g, were purchased from Huafukang Bioscience (Beijing, China) and randomly assigned into two groups: the High-fat diet (HFD) group (n = 6) and the HFD+M2 macrophage-derived exosomes (HFD+ExoM2) group (n = 6). After 6 weeks on a high-fat diet (H10540, Beijing HFK Bioscience Co. Beijing, China , Protein:Carbohydrate:Fat = 20:40:40 [kcal%]), mice in the HFD+ExoM2 group received tail vein injections of ExoM2, 200 µg per mouse, twice a week for 4 weeks. Mice in the HFD+PBS group were administrated an equivalent of phosphate-buffered saline (PBS) via the tail vein. The dosing regimen was based on previous studies [16, 17]. Mice were killed by cervical dislocation under isoflurane anesthesia, followed by exsanguination and organ isolation.
All animal experiments were conducted in accordance with the Guide for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee (IACUC) of Fuwai Hospital, Chinese Academy of Medical Sciences.
The mouse macrophage cell line RAW264.7 was purchased from the Cell Resource Center of Peking Union Medical College (PCRC). When cell confluency reached 50%–60%, the cell culture medium was replaced with exosome-free Dulbecco’s Modified Eagle Medium (Gibco BRL, Carlsbad, CA, USA), supplemented with 20 ng/mL lnterleukin 4 (IL-4) (Peprotech, Rocky Hill, NJ, USA). After 24 hours, the polarization of macrophages towards the M2 phenotype was assessed using immunofluorescence staining for CD206, an M2 macrophage-specific protein marker.
Mouse aortic smooth muscle cells (MOVAs) were purchased from
Fenghui Biotechnology (Beijing, China). When MOVAs reached 60%
confluency, 30 ng/mL platelet-derived growth factor-BB (PDGF-BB) was added to
induce atherosclerotic pathological changes [18]. After 24 hours, cells were
lysed to collect proteins, and the expression levels of contractile phenotype
markers of
All cell lines were validated by STR profiling and tested negative for mycoplasma. Cells were all cultured in a humidified incubator at 37 °C and 5% CO2.
Serum-free M2 macrophage cell culture supernatant was collected and centrifuged
at 4 °C, 2000
Exosome ultrastructure and size distribution were assessed using transmission electron microscopy (Hitachi Model H-7650 TEM, Tokyo, Japan) and Nanoparticle Tracking Analysis (Malvern, UK). Drawing from previous studies [19], the MicroBCA Protein Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA) was used to determine the protein concentration of the isolated exosomes. Immunoblotting was performed to detect specific exosome markers, including tumor susceptibility gene 10 (TSG101), ALG-2 interacting protein X (ALIX), heat shock protein 70 (HSP70), and Calnexin.
For the in vitro exosome uptake assay, purified ExoM2 were labeled
with the membrane-labeling dye PKH67 (Sigma-Aldrich, Eschenstr, Taufkirchen,
Germany) according to the manufacturer’s instructions. Next, VSMCs were incubated
with 100 µg/mL PKH67-labeled exosomes for 0, 1, 2, 3, and 4 hours, and
stained with 4
Purified ExoM2 were labeled with PKH67 (Sigma-Aldrich, Eschenstr, Taufkirchen, Germany) following the manufacturer’s instructions. ApoE-/- mice in the PKH67-labeled ExoM2 group were injected intravenously with 200 µg of exosomes every other day for 2 weeks [16]. Animals in the blank control group received an equal volume of PBS. At the endpoint, major organs (the aorta, lung, spleen, kidney, liver, and heart) were harvested to assess the in vivo distribution of PKH67-labeled ExoM2. The fluorescence intensity was quantified using the IVIS® Spectrum system and Living Image® Software (Version 4.4, PerkinElmer, Shanghai, China). Additionally, fluorescence quantification of tissue sections was performed using fluorescence microscopy. Cell nuclei were stained with DAPI (Sigma-Aldrich, Eschenstr, Taufkirchen, Germany).
Purified exosome pellets or cultured MOVAs were harvested and lysed in RIPA
(Beyotime Biotechnology, Shanghai, China) buffer supplemented with complete
protease inhibitor cocktail tablets (Roche, Basel, Swiss). The lysates were
separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gels, transferred to Polyvinylidene Fluoride (PVDF) membranes (Bio-Rad, Hercules, CA,
USA), and respectively incubated with antibodies against TSG101 (ab125011), Alix
(ab275377), HSP70 (ab2787), and Calnexin (ab22595) (for exosome, Abcam, London,
UK), or antibodies against ɑ-SMA (55135-1-AP), SM22ɑ (10493-1-AP), OPN
(22952-1-AP) and ɑ-Tubulin (11224-1-AP) (for MOVAs, Proteintech, Wuhan, Hunan,
China). The Bicinchoninic Acid Protein Assay Kit (Thermo Fisher Scientific,
Waltham, MA, USA) was used to calculate the protein concentration. The signal
intensity of primary antibody binding was quantitatively analyzed using ImageJ
software (Version: 2.3.0/1.53q, LOCI, University of Wisconsin, Madison, WI, USA)
and normalized to the signal intensity of GAPDH or
The wound healing assay is a well-established method for measuring cell
migration [20]. To evaluate the effects of ExoM2 on the PDGF-BB-induced
migration of VSMCs, MOVAs (2
VSMCs were seeded into 96-well plates (Beyotime Biotechnology, Shanghai, China)
at a density of 2
At the endpoint of the study, the animals were euthanized by CO2
suffocation. Arterial tissues containing plaques were collected, and a
comprehensive analysis of the effects of ExoM2 on the progression and
stability of atherosclerotic plaques was performed using Oil Red O (ORO)
staining, hematoxylin-eosin (H&E) staining, and Masson’s trichrome staining. To
explore the cellular mechanisms underlying the atheroprotective effects of
ExoM2, immunohistochemistry analysis was used to detect the expression
levels and spatial distribution of OPN, CD68, MMP-9, and
Quantitative analyses were conducted with at least three independent
experiments. Results are presented as the mean
The ExoM2 used in this study was derived from IL-4 (20 ng/mL)-induced RAW264.7 cells. As shown in Fig. 1A, CD206 expression in RAW264.7 cells reached 100%, indicating successful polarization into M2 macrophages. To ensure the quality and purity of the exosomes, we used transmission electron microscopy, NanoSight, and Western blotting to identify their morphological structures and protein markers, respectively. The results showed that ExoM2 had a diameter ranging from 30 nm to 150 nm and a ‘round-shaped’ morphology (Fig. 1B,C). Western blotting revealed that the ExoM2 expressed TSG101, Alix, and HSP70, but lacked calnexin, an endoplasmic reticulum-specific marker (Fig. 1D), consistent with typical exosome characteristics.
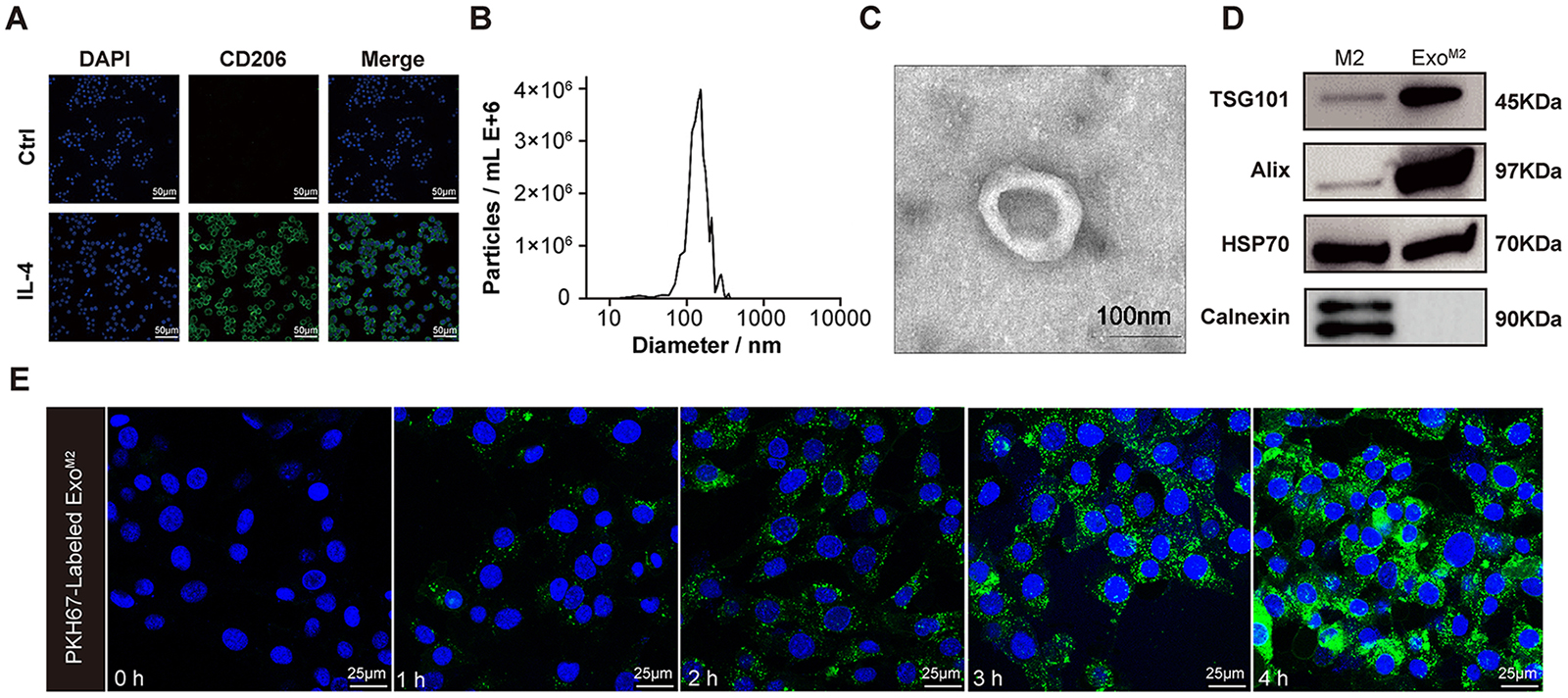 Fig. 1.
Fig. 1.
Isolation and identification of Exo𝐌𝟐 in vitro.
(A) Identification of M2 RAW264.7 macrophages induced by IL-4. Fluorescence
staining showed that CD206 (green) was expressed in all cells. Scale bar: 50
µm. (B) Size distribution profile of ExoM2 as determined by Nanosight.
(C) Ultrastructure of ExoM2 observed by transmission electron microscopy.
Scale bar: 100 nm. (D) Western blot was used to detect the levels of exosome
markers TSG101, Alix, HSP70, and Calnexin. (E) Representative fluorescence images
of VSMCs loaded with PKH67-labeled ExoM2 (green) (100 µg/mL) after 0,
1, 2, 3, and 4 h of co-incubation. Nuclei were stained with DAPI (blue). Scale
bar: 25 µm. ExoM2, exosomes derived from M2 macrophages;
IL-4, lnterleukin 4; DAPI, 4
To detect the manner of interactions between ExoM2 and recipient VSMCs, ExoM2 were labeled with PKH67 and co-cultured with VSMCs, and then visualized with confocal microscopy. Results revealed an increase in the number of internalized vesicles increased over time (Fig. 1E). ExoM2 were observed around the nuclei or lining the inner surface of the cell membrane after internalization into VSMCs, suggesting that ExoM2 can be effectively taken up by VSMCs, and membrane fusion may be the primary mechanism through which VSMCs internalize exosomes.
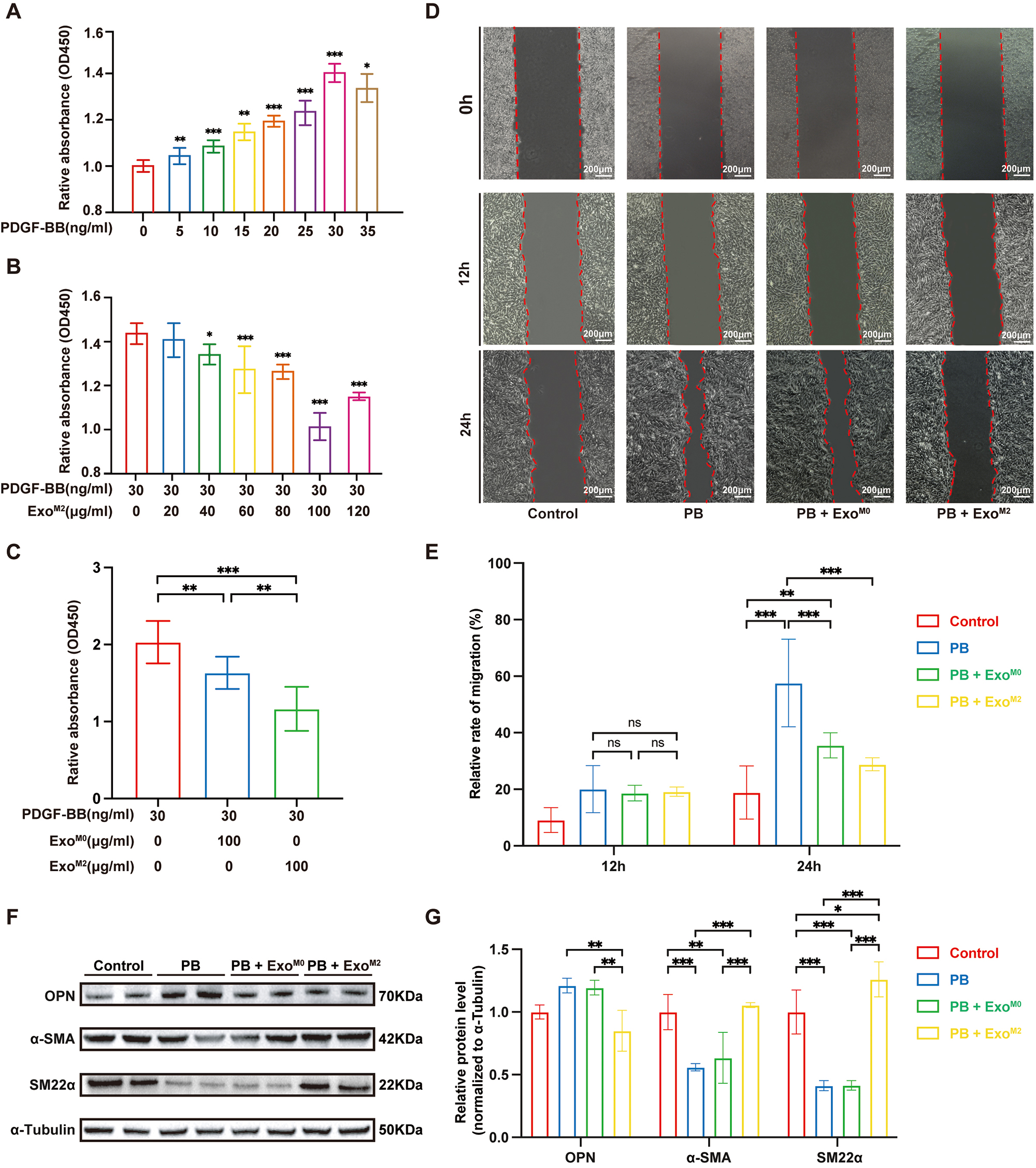
PDGF-BB is a commonly used inducer to replicate in vitro the
pathological changes of VSMCs associated with atherosclerotic cardiovascular
disease, such as proliferation, migration, and transformation from a contractile
to a synthetic phenotype [21]. To investigate whether ExoM2 regulate these
atherosclerotic pathological changes in VSMCs CCK8 and wound-healing assays were
conducted using mouse aortic vascular smooth muscle cells (MOVAs). CCK8 assay
results showed that ExoM2 (100 µg/mL) attenuated PDGF-BB-induced
proliferation by approximately 30% (Fig. 2A,B). Compared to ExoM0,
ExoM2 more significantly reversed the proliferation of MOVAs induced by
PDGF-BB (Fig. 2C). In the wound-healing assays, neither ExoM0 nor ExoM2
significantly changed cell mobility at 12 h compared to the control (PDGF-BB)
group. However, at 24 h of co-incubation, ExoM2 treatment significantly
inhibited PDGF-BB-induced migration of MOVAs (p
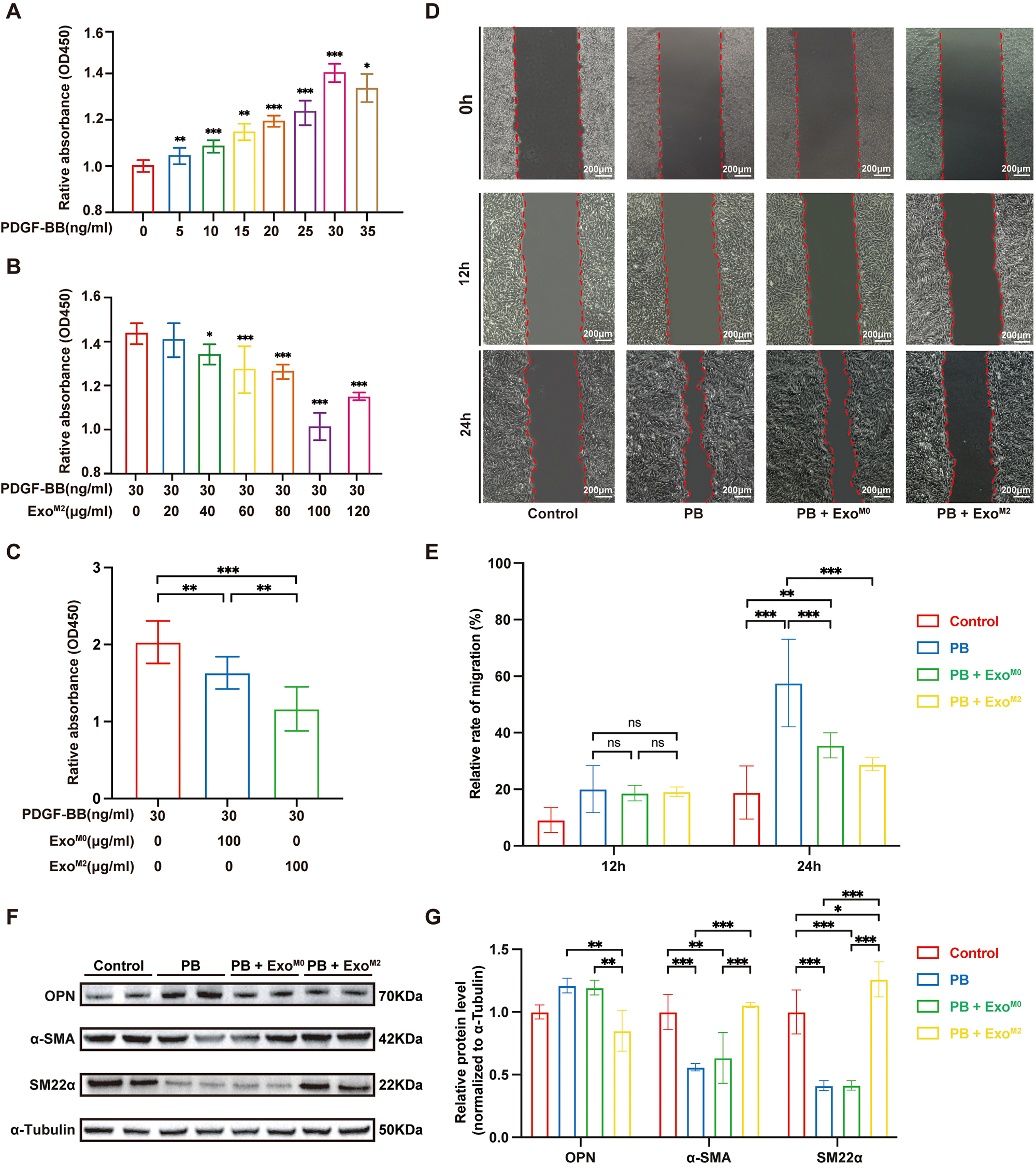 Fig. 2.
Fig. 2.
The effects of Exo𝐌𝟐 on PDGF-BB-induced proliferation,
migration, and phenotypic transformation of MOVAs. CCK8 assays were used to
assess cell proliferation. (A) PDGF-BB-induced MOVA proliferation in a
dose-dependent manner (n = 6 per group). (B) ExoM2 (40–100 µg/mL)
effectively inhibited PDGF-BB (30 ng/mL)-induced MOVA
proliferation (n = 6, per group). (C) ExoM2 (100
µg/mL) had a more significant inhibitory effect on PDGF-BB-induced MOVA
proliferation compared to the same dose of ExoM0 (n = 6 per group). (D)
MOVAs were incubated with ExoM0 or ExoM2 for 4 h and then were
stimulated with or without PDGF-BB (30 ng/mL) for 12 or 24 h. Differences in the
relative migration rate of MOVAs were detected by wound-healing assays (n = 6 per
group; Scale bar: 200 µm). (E) Quantification of the VSMC migration area
using ImageJ software. (F) Western blotting bonds represent the regulatory
effects of ExoM0/ExoM2 on the expression of
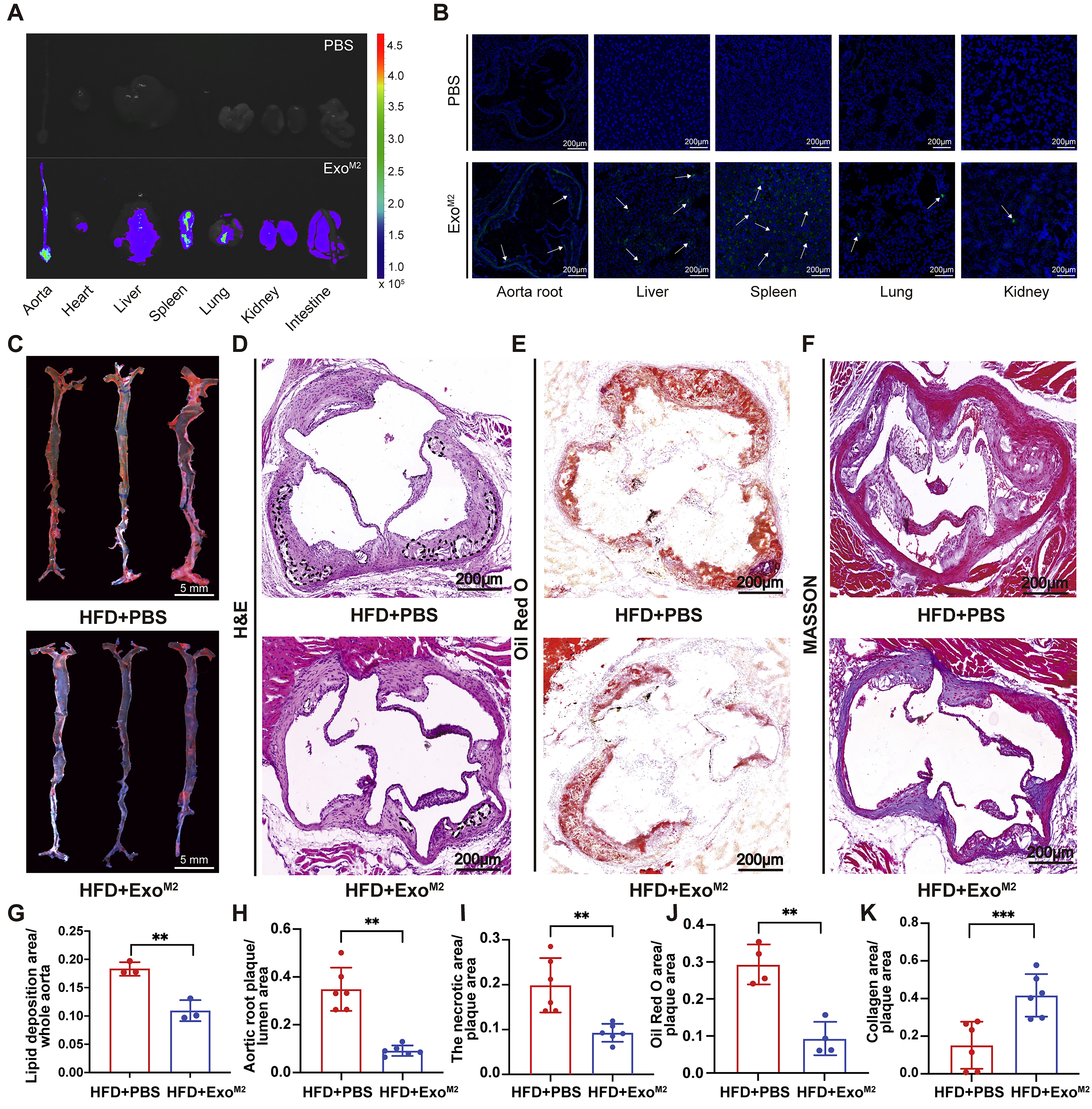
To explore the lesion-targeting ability of ExoM2, we labeled purified ExoM2 with PKH67 and administered it into ApoE-/- mice via tail vein injections (200 µg of exosomes per mouse) every other day for 2 weeks [16]. As shown in Fig. 3A, fluorescence-labeled ExoM2 mainly accumulated in the liver, spleen, and aorta, consistent with the results of fluorescence imaging of tissue sections (Fig. 3B), indicating that ExoM2 could be successfully taken up by histiocytes, including those within plaque tissue at the aortic root.
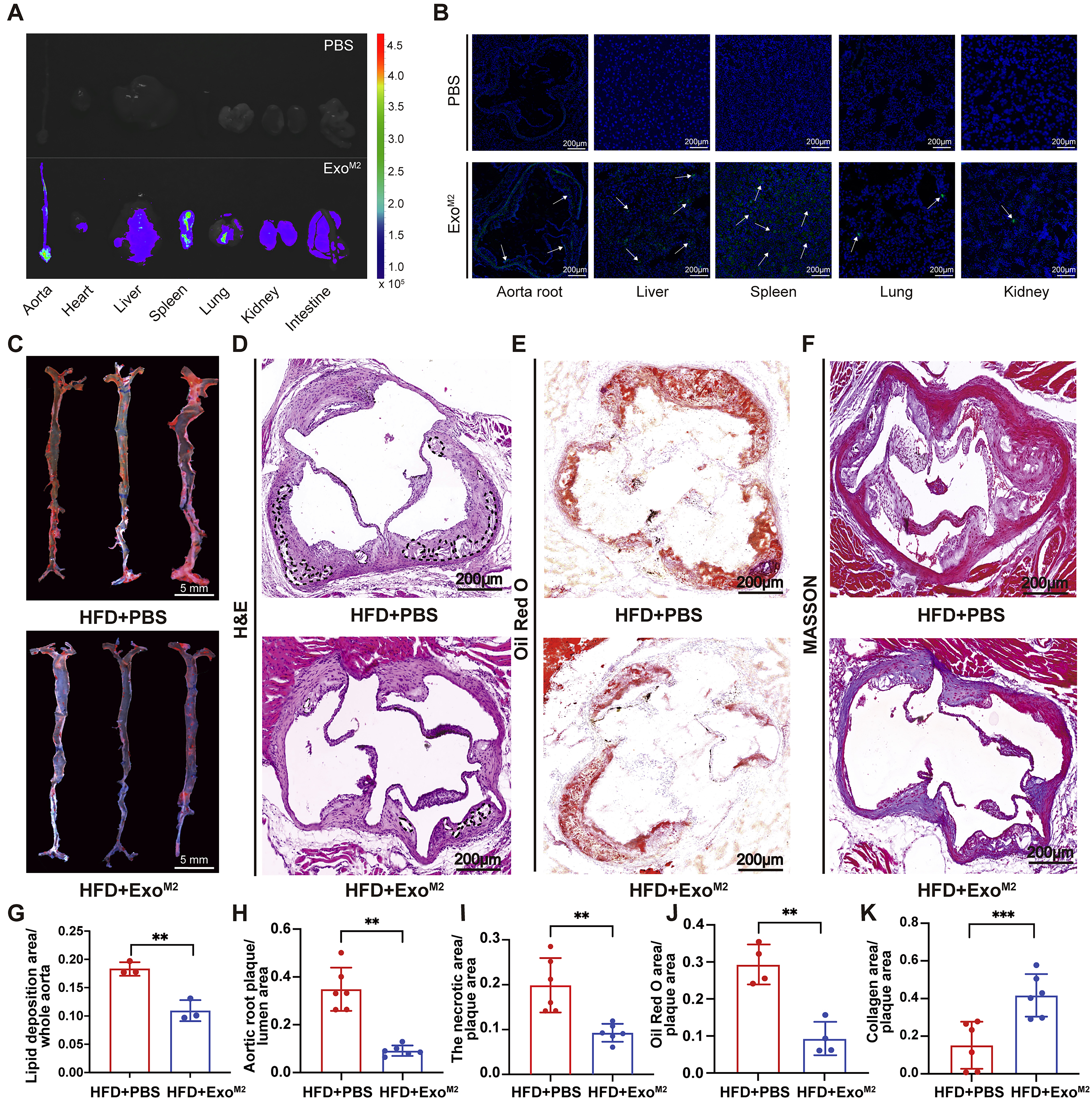 Fig. 3.
Fig. 3.
The effect of Exo𝐌𝟐 on plaque progression in ApoE-/-
mice. (A) Ex vivo fluorescence bio-imaging of major organs after
ApoE-/- mice were administrated PKH67-labeled ExoM2 (200 µg per
mouse, once every other day for 2 weeks). (B) Representative images of
PKH-67-labeled ExoM2 in different tissue sections. Nuclei were stained with
DAPI (blue). The arrows indicate PKH-67-labeled M2 macrophage derived exosomes. Scale Bar: 200 µm. (C) Oil Red O staining aortic tissues from atherosclerotic mice with
or without ExoM2 treatment (n = 3 per group). Scale bar: 5 mm. (D–F)
Representative images of H&E staining, Oil Red O staining, and Masson staining
in cross-sections of the proximal aorta. Scale Bar: 200 µm. (G)
Quantitative analysis of atherosclerotic lesion size in the whole aorta (n = 3).
(H) Quantification of plaque area in the aortic root in H&E-stained proximal
aorta (n = 6). (I) Quantification of necrotic area in H&E-stained proximal aorta
(n = 6). (J) Quantification of positive area in Oil Red O-stained proximal aorta
(n = 4). (K) Quantification of collagen content in aorta root sections stained
with Masson’s trichrome (n = 6). Statistical analysis was conducted using
Student’s t-test. ** p
ExoM2 inhibited plaque progression in ApoE-/- mice fed a high-fat
diet (HFD). Oil red O staining of longitudinal aortic sections revealed that
compared to the HFD+PBS group, ExoM2 treatment significantly reduced the
percentage of the plaque area relative to the total aortic area (10.94%
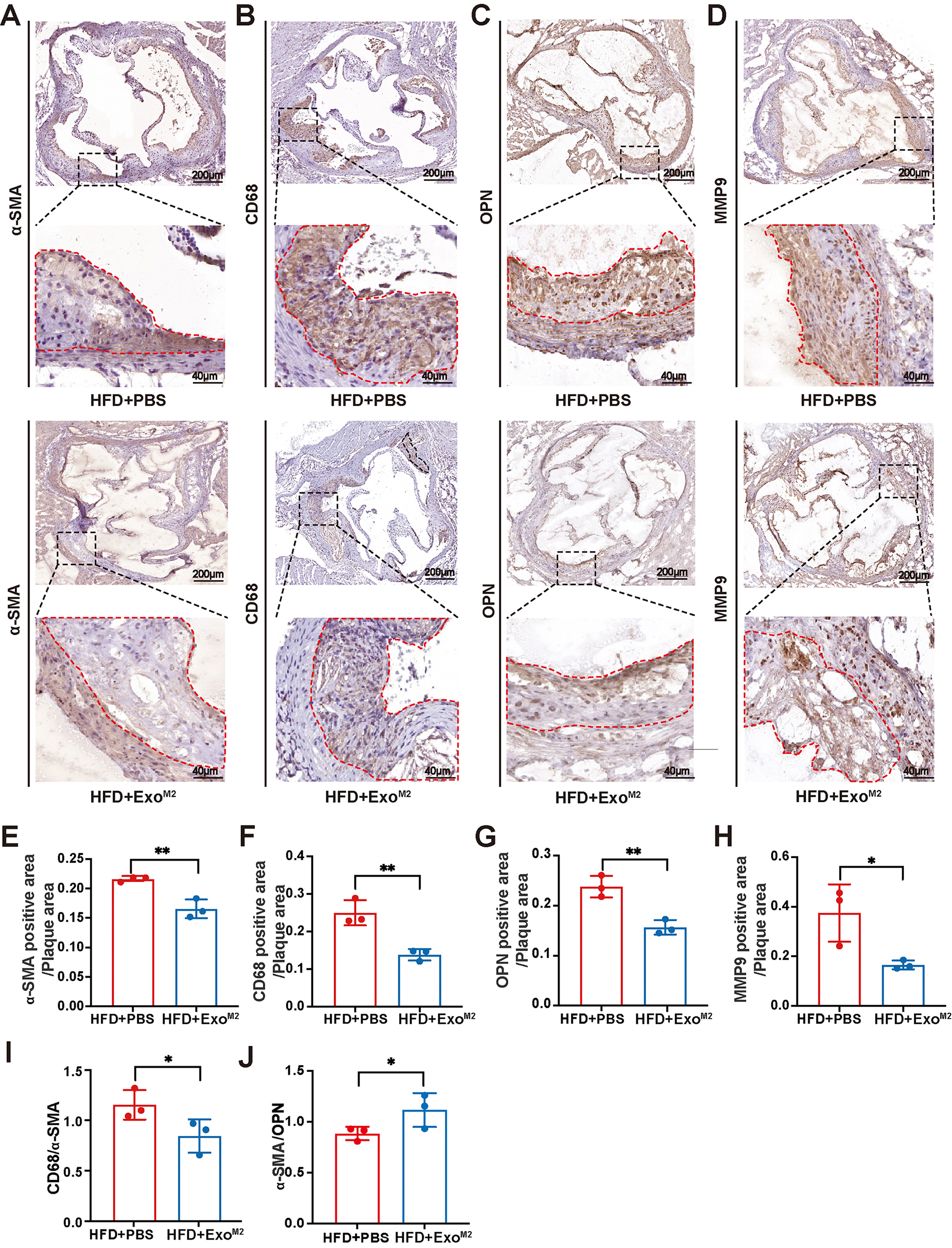
To elucidate whether the atheroprotective role of ExoM2 was associated with
smooth muscle cells, further histopathological analysis of the plaques was
performed. Immunohistochemical staining results showed that compared to the
HFD+PBS group, the areas of
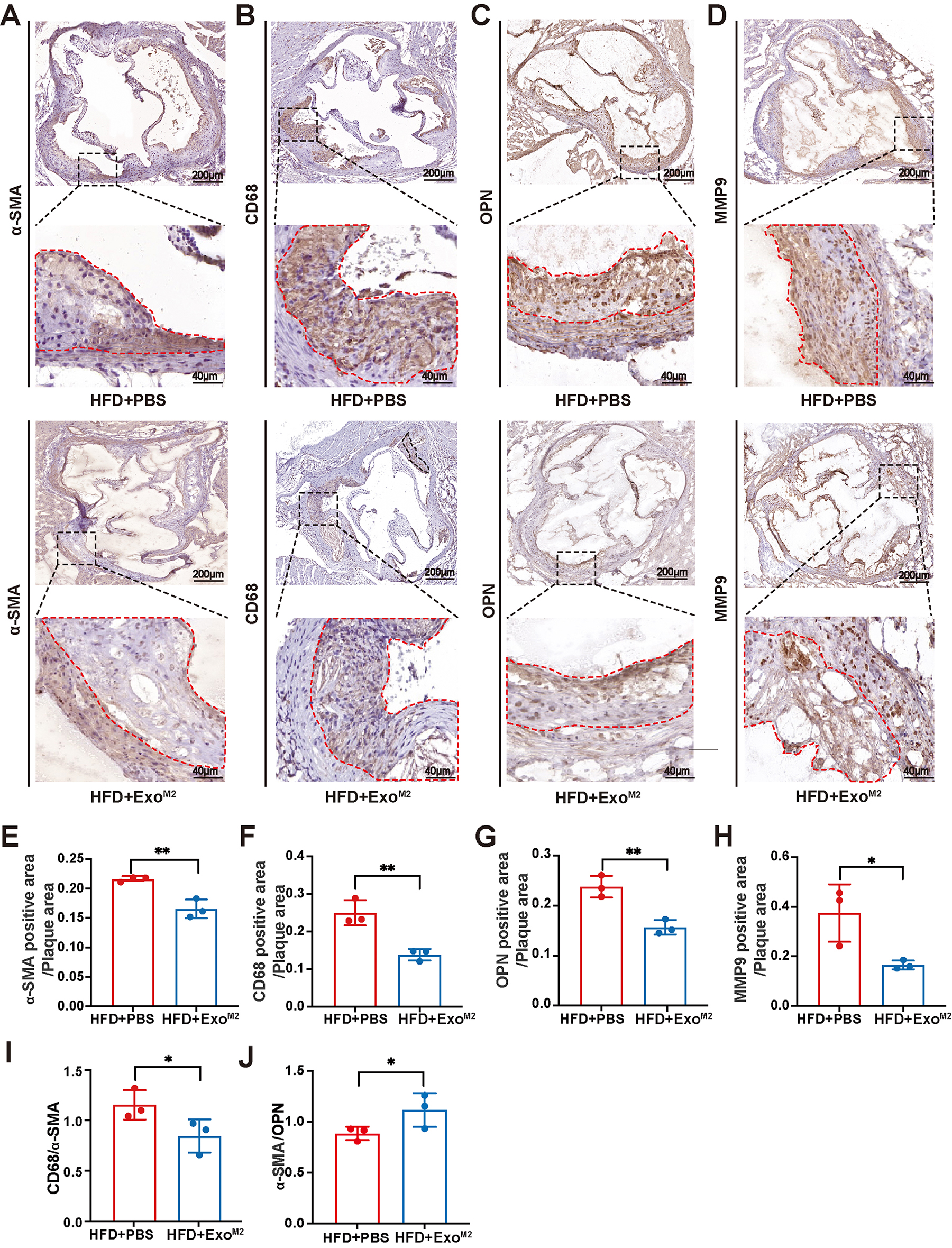 Fig. 4.
Fig. 4.
The effect of Exo𝐌𝟐 on atherosclerotic plaque stability.
(A–H) Representative images and quantitative analysis of aorta root sections
stained with antibodies against
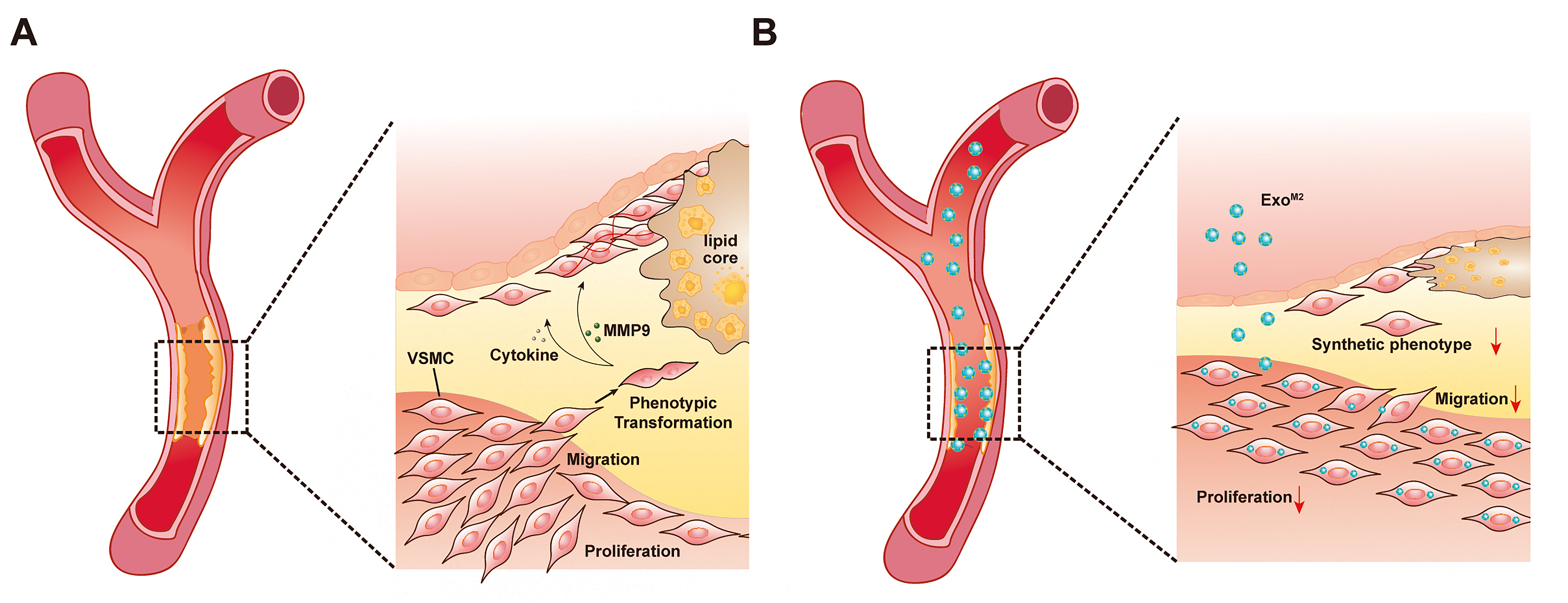 Fig. 5.
Fig. 5.
Schematic diagram of the cellular mechanism of Exo𝐌𝟐 regulating atherosclerosis. (A) In atherosclerotic plaque progression, smooth muscle cells proliferate and migrate from the media to the intima, where they transform into a synthetic phenotype, secreting high levels of MMP9 and cytokines, which promotes plaque progression and reduces plaque stability. (B) The above process was considerably suppressed by ExoM2. MMP9, matrix metalloproteinase 9; VSMC, Vascular smooth muscle cell.
Macrophages play a crucial role in AS progression by secreting various substances that affect neighboring cells, such as VSMCs, throughout all stages of AS [22]. They can be classified into pro-inflammatory M1 and anti-inflammatory M2 types [23]. While numerous previous studies have reported the role of macrophages and VSMCs in the development of atherosclerotic diseases [24, 25], research on the effects of M2 macrophage-derived exosomes on the atherosclerotic pathological changes in smooth muscle cells remains underexplored. In this study, we found that ExoM2 inhibit AS progression by regulating the proliferation, migration, and phenotypic transformation of VSMCs.
Aortic medial smooth muscle cells constitute the main part of atherosclerotic plaques. A large number of studies have demonstrated the correlations of AS development with abnormal proliferation, migration, and phenotypic transformation of smooth muscle cells [26, 27]. It is widely accepted that the phenotypic transformation of VSMCs from a contractile to a synthetic phenotype indicates atherosclerotic disease progression.
Previous studies have demonstrated that this phenotypic transformation triggers the release of various pro-inflammatory factors and destabilizes atherosclerotic plaques [28, 29, 30]. Therefore, inhibiting VSMC proliferation, migration, and phenotypic transformation has become a key strategy for AS management. However, an ideal solution for addressing this pathological change has yet to be identified.
Recent reports have indicated that smooth muscle cell proliferation can be inhibited when co-cultured with macrophages [22]. However, the underlying mechanisms remain unknown. Exosomes, carrying a variety of bioactive substances, are critical mediators for cell-to-cell communication [31]. The question arises whether M2 macrophages regulate their biological function of smooth muscle cells through exosome secretion. To investigate this, we examined the effects of ExoM2 on the proliferation, migration, and phenotypic transformation of VSMCs. We firstly utilized a recognized PDGF-BB-induced phenotypic transformation model of VSMCs to simulate their pathological changes in atherosclerotic conditions. Then we applied the well-identified ExoM2 to regulate the above pathological processes. Compared to ExoM0, treatment with 100 µg/mL ExoM2 can significantly inhibit VSMC proliferation and migration, and reverse the phenotypic transformation from a contractile to a synthetic phenotype.
Plaque stability is generally assessed by indicators such as plaque volume,
structure, and composition. Plaque stability is inversely correlated with plaque
size [32], the ratio of lipid core to fibrous caps [33], the ratio of macrophages
to contractile phenotype smooth muscle cells (CD68/
The present study has some limitations. While we have uncovered a novel mechanism by which ExoM2 exerts its anti-atherosclerotic effects, this study was limited to the cellular level and did not extend to the molecular level. Subsequent studies will focus on what cargos in ExoM2 mediate the effects of ExoM2 on VSMC proliferation, migration, and phenotypic transformation. Additionally, the atheroprotective effects of ExoM2 require validation through clinical evidence.
M2 macrophage-derived exosomes can significantly inhibit the progression and improve the stability of atherosclerotic plaques in ApoE-/- mice. The atheroprotective effects of ExoM2 are associated with the inhibition of VSMC proliferation, migration, and phenotypic transformation. Despite challenges such as large-scale production, exosomes remain a promising new avenue for the preventive therapy of AS.
VSMCs, vascular smooth muscle cells; MOVAs, mouse aortic smooth muscle cells;
HFD, high fat diet; PBS, phosphate buffer saline; ExoM2, exosomes derived
from M2 macrophage/M2 macrophage-derived exosomes; H&E, hematoxylin and eosin;
DMEM, Dulbecco’s Modified Eagle’s Medium; FBS, fetal bovine serum; TEM,
transmission electron microscopy; DIO, 3,3
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
YC and TC designed the research study, reviewed and revised the manuscript. ShuoW, XW, and YL performed the research, analyzed the data, and wrote the manuscript. ZZ and ZY provided help on the preparation of figures. ShuangW, CW, KZ, TH, XH and JM provided help and advice on the animal and cell experiments. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All animal experiments were conducted in accordance with the Guide for the Use and Care of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee (IACUC) of Fuwai Hospital, Chinese Academy of Medical Sciences (IACUC reference number: FW-2020-0018).
Here we acknowledge all members of the Animal Experimental Center of Fuwai Hospital, Chinese Academy of Medical Sciences for their support with animal experiment facilities, equipment, and technologies.
This research was funded by the National Natural Science Foundation of China (Grant number: 81970387, 81200213), the Medical Scientific Research Project of Jiangsu Provincial Health Commission (Z2021069).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.