

1 Department of Biology and Biotechnology, University of Pavia, 27100 Pavia, Italy
2 Department of Pharmaceutical Sciences, University of Perugia, 06126 Perugia, Italy
Abstract
Bulimia nervosa (BN) is a condition marked by a typical cyclical behavioural activity, characterized by restrictions, binges and vomiting, as well as a disturbance of the emotional value of food. Food stimuli acquire excessive relevance, giving rise to a succession of states of excitement and anxiety. The depressive condition accompanies very often BN. Most people with BN also experience one or more anxiety disorders. The aim of the review is to identify a link at a central and peripheral level that connects an eating disorder with a mood state. Altered nervous mechanisms are involved in BN. Among the cerebral areas, the insula is functionally compromised in BN. The insula is also implicated in depressive states. The insula is the primary gustatory cortex, where gustatory sensory information such as taste discrimination and higher cognitive functions such as food anticipation and reward are processed. The insula is anatomically connected to a wide range of cortical, limbic and paralimbic structures, and functionally implicated in high-order cognition, emotional responses, and empathic processes. The insula plays a crucial role in empathy, or in the ability to share the emotional states of others, and in particular negative emotions. In fact, the insular cortex is also activated in conditions of anxiety and depression. One of the pathophysiological factors that influences bulimia and depression is the composition of gut microbiota, as there is a strong association between the microbial signature and the brain function. Gut dysbiosis condition may contribute to the development of eating disorders, including BN. Dysbiosis may promote intestinal inflammation, alter gut permeability, and trigger immune reactions in the hunger/satiety regulation center contributing to the pathophysiological development of eating disorders. From this emerges the importance of adequate probiotic integration as a preventive and/or therapeutic tool in these pathologies.
Keywords
- bulimia nervosa
- depression
- insula
- opioid
- microbiota
- microbiota-gut-brain axis
- psychobiotics
It is common opinion that when trying to explain why some people eat too much food, phrases like “I eat because I feel depressed” emerge.
It is said that those who have problems with food control have “nervous hunger”, indicating some form of mental malaise, anxiety or depression, but the qualification “nervous” calls to mind the concept of frenzy, eagerness a ravenous hunger given that there is a voracity disproportionate to the actual need for refreshment. Being nervous does not come from hunger, but rather from appetite, and the lack is not of energy and nutrients, but of balance in the gratification-seeking system. Food is seen as a tool to fill an internal void or as a means of comfort. So, is it depression or rather a form of agitation that drives you towards food when you are not mentally at ease [1, 2]?
Bulimia nervosa (BN) is considered an eating disorder (ED) in accordance with the Diagnostic and Statistical Manual of Mental Disorders (DSM-5).
It is a serious psychological disorder characterized by frequent binge eating (consuming excessive quantities of foods in brief periods of time) accompanied by compensatory purging behaviour as, i.e., abuse of laxatives and diuretics, self-induced vomiting, fasting and very intense exercise, occurring at least once a week over the course of three months [3]. The frequency of purging behaviour determines the severity of BN. According to the World Health Organization, the prevalence of BN in adults is 4% to 6.7% considering the DSM-5 criteria [4]. BN develops mainly in adolescence or young adulthood and affects both sexes, with a higher prevalence in girls and young women [5]. More than 70% of people with BN report concomitant psychiatric comorbidity comorbidities—i.e., affective disorders, anxiety and drug use [6]; precocious psychiatric illness is considered a predisposing factor for BN symptoms [7].
The aetiology of BN is multifactorial, and is determined by a complex combination of environmental and genetic factors, including psychosocial influences and neurobiological factors [8]. Environmental factors contributing to aetiology, gravity and treatment comprise socio-cultural factors such as the ‘Western’ ideal of thinness and attractiveness [9, 10, 11]. Population-based studies of BN highlight that the prevalence of BN is elevated in their relatives of BN compared with the relatives of controls [12], with heritability estimates ranging from 54 to 83% [13, 14].
The mortality rates in BN subjects are significantly elevated from 1.5% to 2.5% in great part due to the medical implications, among which ischaemic heart disease [11]. In addition, suicide was considered the most important cause of death in individuals with BN; furthermore, a very low body mass index (BMI) was, also, found to be an important death predictor and this could be the result of a previous anorexia nervosa (AN) history. In fact, evidence shows that 9/10 of the deceased patients were known to have been AN before being BN [15].
According to reports, clinical identification of EDs, such as BN, in male patients presents less often than in female patients [16, 17]. This difference is likely a result of previous diagnostic criteria, which included amenorrhea as one of the diagnostic criteria in the Diagnostic and Statistical Manual of Mental Disorders Fourth Edition (DSM IV) [16]. In addition, male individuals with eating disorders may exhibit increased muscularity instead of a low BMI and a focus on exercise [16, 17]. Furthermore, the presence of an obesity history in male patients [18] makes it difficult to detect EDs. The link between EDs and depression is multifaceted and characterized by interconnected physiological and behavioural factors that influence aetiology, symptoms, and response to treatment. The rate of depression disorders (DD) is higher than 40% among EDs, EDs which is the second most common psychiatric comorbidity after anxiety disorders [19, 20].
EDs, such as BN, were found to have a mutual influence with depression, according to longitudinal studies. Indeed, in a large sample of ED patients, one-third of patients with a comorbid diagnosis of major depressive disorder (MDD) evolved MDD before the start of their ED, and two-thirds of the patients in the same year or after the ED onset [21].
Recognized studies have shown that DDs comorbid with EDs are linked to lower rates of remission after treatment [22] and persist even after an ED has remitted [23]. Other studies have shown that treatment of BN leads to a decrease in depression [24, 25].
In EDs body dissatisfaction and self-esteem are the main causes of depressive symptoms [26, 27]. The association between EDs and depression among females has been extensively documented by research from both clinical and epidemiological studies [28, 29].
Depression in comorbidity with ED can enhance the gravity and chronicity of both conditions [30, 31]. Indeed, 85.6% of female adolescents who were hospitalized for ED treatments had a history of depressive episodes throughout their lives, which were linked to a greater degree of disorder severity and a poorer prognosis [32]. A high percentage of male adolescents with EDs have comorbid depression and their rate of having a history of suicidal ideations or attempts is twice as high as that of female adolescents [33].
BN is a condition characterized by a typical cyclical behavioural activity of food consumption, as well as a disturbance of the emotional value of food. It appears that in bulimia there may be a lack of interoceptive sensations. BN frequently coexists with states of anxiety and depression. Mood dysregulation and EDs appear to be potential moderators of anxiety and depression conditions [34].
However, scientific research in this field is still relatively limited and little is known about which BN symptoms contribute to the psychopathology of BN and how they are associated with symptoms of depression and anxiety. Certainly, reduced self-esteem and high impulsivity are central factors in the psychopathology of BN [35].
Various studies have reported evidence that most subjects suffering from BN also have one or more anxiety disorders. Obsessive-compulsive disorder, social phobia, and specific phobia are among the most common anxiety disorders in bulimics [36].
Avoidance of social eating is a very common symptom of EDs in general and BN in particular, and it may partly explain the overlap between EDs and anxiety-provoking states. This is explained by the fact that anxiety disorders can involve the fear of eating in a social context. Low self-esteem is an aspect present in both anxiety and bulimic subjects as they use physical appearance as a tool to evaluate and regulate their self-esteem [37]. One strategy currently being conducted to study this type of association uses network analysis. This approach allows you to analyse within a mathematical model a large number of symptoms, such as body dissatisfaction, perfectionism, social insecurity, which interact simultaneously. Studies have shown that the central factors of BN are the fear of gaining weight and the strong desire to lose weight. From this model it emerges that the bulimic subject maintains a state of activation [38, 39, 40].
A brain region involved in eating behaviour and EDs is the insula. The insula is a portion of the cerebral cortex that receives multisensory information and is involved in a variety of functions including sensory processing. It is the primary gustatory cortex, where gustatory sensory information such as taste discrimination and higher cognitive functions such as food anticipation and reward are processed. The insula region represents a crucial centre of integration between the external environment and internal processing [41]. Food assumes excessive importance, giving rise to a succession of states of excitement and anxiety, and this is probably due to the fear, real or perceived, of being or becoming fat [42, 43, 44].
The insular cortex is implicated not only in other addictive, reward-driven behaviours, including drug abuse and gambling, but it has also been the most consistently activated area in studies of neuroimaging on anxiety [45, 46, 47].
Altered signaling at the insula level could translate into an alteration of the hedonic process. In bulimic subjects, an increase in the activation of the insula could lead to an exaggerated interoceptive perception of hunger. This would lead to amplifying hedonic regulation systems, leading to greater food intake.
Although various brain regions including the amygdala and prefrontal cortex are involved in emotional states, the anterior insular cortex appears to be a rather critical area both for depressive condition and for BN [48, 49]. In particular, the insula is a brain area implicated in depressive states. Studies have reported that the depressive condition is accompanied by morphological alterations at the insula level. Furthermore, it was reported that insula metabolism was altered in depressed patients. In depressed subjects, a reduction in the connections between the insula and the amygdala has also been observed [50].
Various studies have highlighted greater atypical activity in depressed subjects. The insula is anatomically connected to a wide range of cortical, limbic and paralimbic structures, and functionally implicated in high-order cognition, emotional responses, and empathic processes [51, 52, 53]. The insula projects to the ventral striatum, which is crucial region for dopamine-mediated reward learning [54]. In addition, the insula projects to the orbitofrontal cortex (OFC), which plays an important role in the control of the striatal activation and motivation to approach food [55].
Finally, it has been posited that the network involving the insula may mediate the potential association between early stress and future depression [56].
The close anatomical proximity of the insula, the OFC and the ventral striatum can also explain the close functional interconnections that are established between these areas. An alteration in the organization of white matter fibers could change the neural connectivity between these regions and alter the taste and processing of food rewards in subjects with EDs. In fact, it is believed that the cortical insula is an area responsible for conscious perception and discrimination between various flavours. In particular, the insula, ventral striatum, and OFC have been repeatedly implicated in functional taste reward and structural brain imaging studies in AN [57, 58, 59]. In this context, it emerges that the insula is a multifunctional, visceral-sensory-motor region. Recent studies have demonstrated its activation even following unfavorable events. The insula also plays a crucial role in empathy, or in the ability to share the emotional states of others, and in particular negative emotions. The insula is also activated by pleasant stimuli, thus becoming the centre that mediates craving, or the irrepressible desire of physical dependence to obtain a substance of abuse, such as smoking, drugs or food [60, 61, 62].
Furthermore, the lateral asymmetry noted in the insular region was consistent with findings from previous cerebral blood flow and metabolism studies on subjects with EDs [63, 64].
Studies reported that chocolate lovers showed insular activation only as long as they described the experience as rewarding, but no longer beyond the point of satiety. Therefore, modulation in cortical chemosensory areas, including the insula and the caudomedial and caudolateral OFC, confirming here to be the site of reward [65]. In addition, an increase in cerebral blood flow at the level of the insula was observed in starved controls, that were fed a liquid meal [66]. Insular activity is therefore confirmed to be linked to food reward.
It is well known that eating behaviour is under the regulation of a complex nervous and humoral system. The fulcrum of this control is carried out by the cerebral cortex as well as by the reward areas of the limbic system, which detect stimuli coming from the environment; they include both food, such as look, taste and smell of food, and non-food, such as emotional distress and stress stimuli. Stimulation of the ventromedial nuclei of the hypothalamus is accompanied by a reduction in appetite, while stimulation of the lateral nuclei is accompanied by its increase.
It has been suggested that in bulimic subjects there may be a state-related
down-regulation of opioid receptors as result of a chronic increase in endogenous
opioid release. Opioids are endogenous mediators released by neurons of the
arcuate nucleus on the ventral tegmental area and nucleus accumbens. It is known
that endogenous opioid peptides include endorphins, enkephalins, dynorphins, and
endomorphins and act through three types of receptors: µ,
Otherwise, decreased MOR binding may be a trait-related marker that increases the reward value of dieting and bulimic behaviours in susceptible individuals [46].
Finally, it has been reported that MOR agonists can increase the intake of palatable food and increase the consumption of more preferred food [68]. Molecules have been identified in milk and wheat proteins which, due to their structure, resemble endogenous opioids. Studies on rats have confirmed that following the ingestion of these foods there is an analgesic effect similar to that of morphine.
Interestingly, a majority of subjects suffering from BN show antibodies against melanocortin peptide alpha- Melanocyte Stimulating Hormone (MSH), which is responsible for reducing food intake, and is affected by pre- and post-synaptic action of endogenous opioids [69]. As mentioned below, BN is characterized by a driven behavioral cycle of restricting, bingeing, and vomiting and by a disturbance in the affective value of food whereby food stimuli acquire excessive salience and induce subjective experiences of increased arousal or anxiety. This emotional response presumably arises from the psychopathologic fear of being or becoming fat that is a hallmark of the disorder.
Some authors have found that bulimic individuals have reduced binding to the opioid receptor MOR in the left insular cortex. This decrease is proportional to fasting behaviour. Specifically, cyclical food deprivation followed by food access alters MOR binding in limbic regions, suggesting that repeated activation of the opioid system through restriction and binge eating may help support bulimic behaviour [70]. A neurotransmitter that is involved in appetite control, ED control and depression is serotonin (5, hydroxitryptamine, 5-HT), whose activity in the brain contributes to a sense of satiety. In theory, bulimic behaviour is consistent with reduced 5-HT activity, whereas anorexigenic behaviour is consistent with increased 5-HT activity. In animal models, studies have revealed inhibitory effects of 5-HT on eating behaviour, suggesting that 5-HT acts as an anorexigenic mediator [71].
Further evidence supports the hypothesis that the reduced post-ingestion satiety in the bulimic subject is linked to a reduced serotonergic reactivity in the hypothalamus [72, 73]. The decrease in 5-HT signaling contributes to the bulimic subject’s binges [74]. On the other hand, dysregulation in the serotonergic system implicated in MDD would explain the increased prevalence of depressive episodes in patients with EDs. Clinical and preclinical studies support the evidence that low 5-HT levels are associated with a depressive condition [75]. The best evidence for the role played by 5-HT in the pathophysiology of depression is confirmed by the depletion of tryptophan, the precursor of 5-HT, a crucial player in depression [76, 77].
There are various pathways by which tryptophan can be metabolised, thus influencing its availability to cross the blood brain barrier. Among these pathways is the kynurenine pathway which is formed through the enzyme indoleamine-2,3-dioxygenase (IDO) [78]. The induction of IDO determines a shift in tryptophan metabolism towards the production of kynurenine and its metabolites; activated IDO accelerates the degradation of 5-HT which produces by-products of reactive oxygen species and subsequently inflammation. This further intensifies the 5-HT deficiency, leading to disruption of neurotransmission and depression [79].
Some neurotrophic factors are highly connected to depression, notably brain-derived neurotrophic factor (BDNF). There is evidence that BDNF acts in accordance with 5-HT in synaptic plasticity phenomena [80, 81].
BDNF, expressed ubiquitously in the brain, controls several behaviours associated with the regulation of food intake by acting in both appetite and satiety brain centers through a complex interactive network [82]. Evidence revealed that serum levels of BDNF in ED patients were found to be significantly lower compared to healthy controls [83]. BDNF displays a close relationship with the depressive condition. In animal model of stress, BDNF levels are reduced in cortex and in hippocampus [84]. In addition, the expression of BDNF and its receptor TrkB was reduced in post-mortem brain tissues from depressed subjects [85], and the deletion of BDNF increases depression-like behaviour [86].
Finally, dopamine (DA) is a neurotransmitter that contributes significantly to EDs. DA is synthesized and released in the midbrain area, including the ventral tegmental area, whose neurons send projections to the ventral striatum which constitutes the cerebral region crucial for food craving and the mesolimbic circuits which are associated with motivation. This system, when hyperactive, leads to an increase in the desire for food-related rewards, thus contributing to the initiation of food consumption [87]. DA is also released in the substantia nigra, whose neurons project to dorsal striatum, ventromedial striatum (including the nucleus accumbens), and the prefrontal cortex [88, 89]. Animal models of depression showed altered mesolimbic DA system function. In addition, depression is associated with a reduced striatal response to reward. Lesions of dopaminergic neurons in the substantia nigra and in the ventral tegmental area enhance depressive-like behaviour in rats [90]. The downregulation of the dopaminergic system is in accord with existing evidence of impaired dopamine release in the nucleus accumbens in animals exposed to chronic stress [91].
Increasing numbers of studies report that one of the pathophysiological factors that influences both bulimia and depressive state, both appetite and mood, is the composition of gut microbiota, as there is a strong association between the microbial signature and the brain function. The gastrointestinal tract hosts the microbiota which is a complex microbial ecosystem consisting of bacteria, but also of viruses, fungi and other microorganisms. The human gut microbiota performs important biological functions that are responsible for the maintenance of the organism homeostasis [92].
A characteristic of BN is uncontrolled eating, which involves intermittent
excessive consumption of palatable foods high in saturated fats and refined
sugars [93]. On the other hand, chronic consumption of a high-fat diet is
associated with peripheral and central inflammation. Proinflammatory cytokines,
including interleukin-(IL)-1
It is known that the intestinal microbiota, the microbial community that populates the gastrointestinal canal and establishes a condition of fundamental mutualistic symbiosis with the host, produces a plethora of neuroactive compounds, toxins and metabolites that are released and act on a broad spectrum of various organs and structures, from heart to liver, from pancreas to brain. These molecules act through the enteric nervous system, circulatory and immune systems to modulate brain signaling pathways that can regulate mood, cognition, social behaviour and memory [97, 98].
Over the last two decades, research has also highlighted that the gut microbiota is an important factor capable of modulating bidirectional communication between the gut and the brain, by contributing to the direct neurobiochemical signaling between the gastrointestinal tract and the central nervous system, called the “gut-brain axis”. This intricate network of communication occurs through the autonomic nervous system, enteric nervous system, neuroendocrine system, and immune system [99].
Ample scientific evidence enables us to link various psychiatric conditions, such as anxiety and depression, to significant alterations in the biodiversity of the intestinal microbiota that could compromise the microbiota-gut-brain axis. This allows the identification of new potential targets for the therapeutic treatment of these pathologies [100].
Probiotics are defined as “live microorganisms which when administered in adequate amounts confer a health benefit on the host” [101, 102]. Specific bacterial strains may themselves improve certain psychiatric states. The term “psychobiotic” was therefore coined [98]. In particular, these probiotic strains improving cognitive functions, reducing stress levels, anxiety and improving mood.
Several studies show that probiotics provide benefits to human health and play a crucial role in the prevention of various diseases [103]. They seem to have an important role in preventing and improving intestinal diseases such as irritable bowel syndrome, metabolic diseases such as diabetes, obesity and cardiovascular diseases [104, 105].
Recently, the identification of a microbiota-gut-brain axis has triggered
numerous studies with the aim of verifying whether probiotics are able to
influence the mental health of the host. What emerges from preclinical and
clinical studies is that the therapeutic effects are involved in reducing
symptoms, for example, in patients with diagnosed anxiety and major depression
[106]. Psychobiotics help produce specific neurotransmitters such as
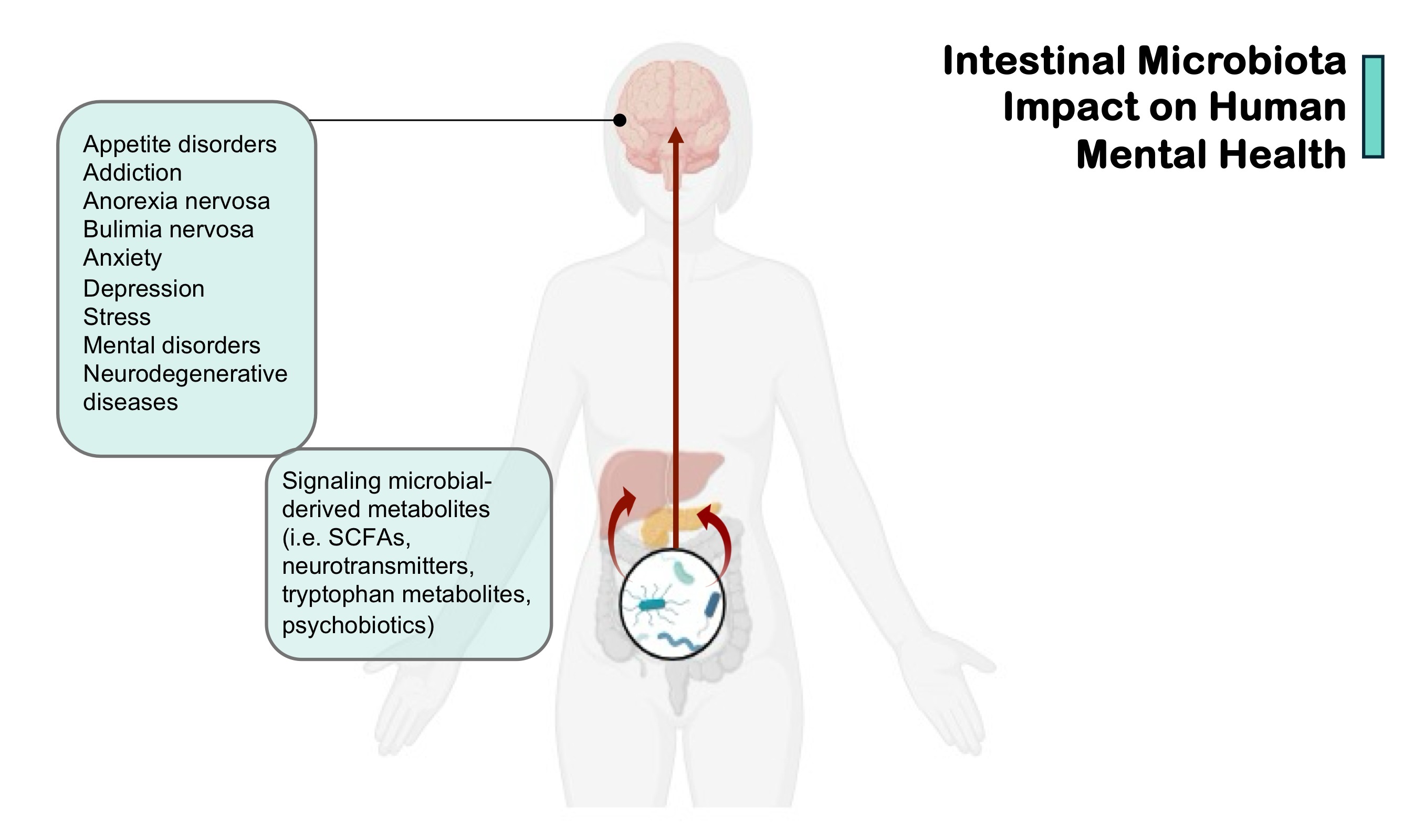
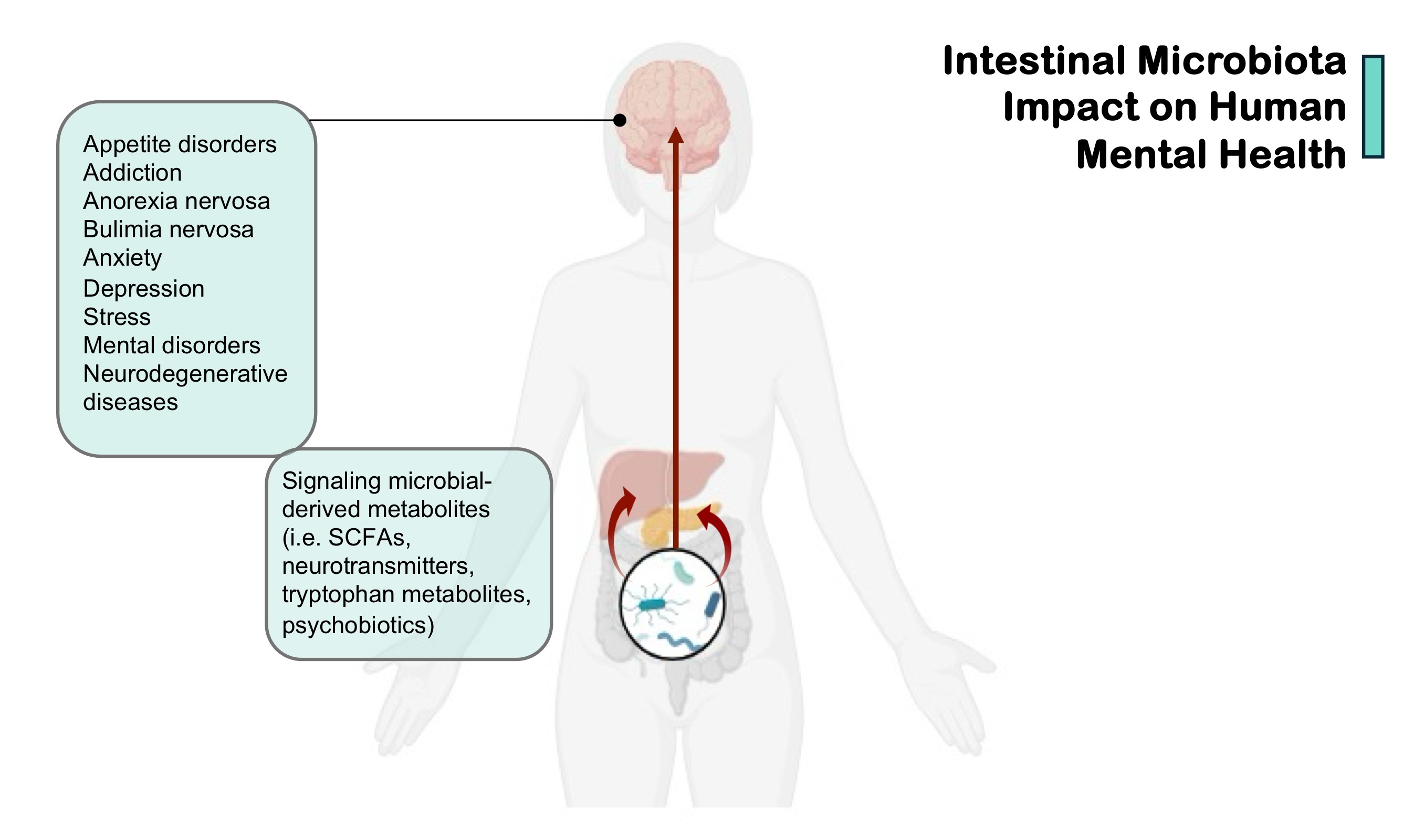 Fig. 1.
Fig. 1.
Intestinal microbiota impact on human mental health. SCFAs, short-chain fatty acids. (Created in BioRender.com.)
An important contribution of gut microbiota dysbiosis to the genesis of EDs has been observed. Dysbiosis, associated with a specific genetic susceptibility, may contribute to the development of EDs. In contrast to AN and binge eating disorder, to date only one study focused on microbiota changes in BN. A relevant study has shown that caseinolytic protease B (ClpB), a protein produced by common gut bacteria is present in human plasma and its concentrations can be elevated in ED subjects, comprise BN cases, and associated with ED-related psychopathological traits. This supports the existence of a link between bacterial ClpB and the ED pathophysiology [115].
Although EDs, anorexia and BN have an etiopathogenesis that is still not entirely clear, recent studies show the involvement of the gut microbiota and the gut-brain axis [116]. Gut dysbiosis could contribute to the onset of inflammation, alteration of the gut permeability and immune hyperactivation with an involvement, at the level of the central nervous system, of the centres for regulating of hunger and satiety, by favouring the onset of EDs [114].
Interestingly, much evidence reports that both the gut microbiota and the immune system are involved in psychiatric disorders, including mood disorders and EDs. Therefore, alterations in the composition of the intestinal microbiota and immune system dysfunctions can maintain and exacerbate dysregulated eating behaviours, characterized by binge eating as in BN, as well as mood and behaviour [110, 117, 118, 119].
The modulation of the microbiota-gut-brain axis through specific biomodulators with prebiotic and probiotic actions could therefore represent a possible therapeutic target in the clinical management of patients with EDs [119].
Knowledge of the biological mechanisms underlying BN and the associated depressive condition, as well as the state of dysbiosis of the intestinal microbiota, the state of peripheral and central inflammation could together serve to identify biomarkers of increased risk of developing a bulimia status.
From this perspective, reducing the depressive condition by modulating the intestinal microbiota could be a strategy to control EDs.
GT, conceptualization; GT, writing-original draft preparation; MR, GC, GT, writing-revision and editing; MR, GC, GT, literature collection; GT, visualization; GT, supervision. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript.
Not applicable.
The authors thank the reviewers for their opinions and suggestions.
This research received no external funding.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.